Needle traits of understory Silver fir, Norway spruce, and Scots pine in response to increased canopy openness in a birch-dominated stand
iForest - Biogeosciences and Forestry, Volume 18, Issue 5, Pages 273-282 (2025)
doi: https://doi.org/10.3832/ifor4909-018
Published: Oct 14, 2025 - Copyright © 2025 SISEF
Research Articles
Abstract
The study investigates the impact of varying environmental treatments on the needle morphology, chemistry, and photosynthetic performance of Silver fir (Abies alba), Norway spruce (Picea abies), and Scots pine (Pinus sylvestris) undergrowth in a birch (Betula pendula)-dominated young forest. The experiment analyzed the effects of different levels of birch canopy reduction. The treatments included a birch preparatory stand managed through positive tending of promising birch trees (PS), a restored small clearing (RC), and a former clearing overgrown by birch sprouts (OC), while the control treatment was left unmanaged. The aim was to evaluate the rate of adaptation to irradiation changes among these species. Fir and pine exhibited significant needle morphological changes within the first year after canopy opening, while spruce needles remained relatively stable. In the first year after release, pine and fir new needles in the RC treatment had reduced K (and Mg in fir), while spruce needles increased N and Ca concentrations. Chlorophyll a fluorescence measurements indicated that photosynthetic efficiency tended to slightly decrease across all species when exposed to higher irradiance. The results show high plasticity of the species in response to the changes in light conditions of the nurse stand. The means of the adaptation were specific for each species, but the new conditions did not limit the adaptation potential for any of them.
Keywords
Acclimation, Needles, Morphology, Chemistry, Chlorophyll Fluorescence, Light Environment, Two-phase Forest Regeneration
Introduction
Climate change poses new challenges for forest management in many regions with deteriorating forests ([18]). To improve the stability of future forests, more diversified stands should be established, particularly by incorporating a wider variety of site-specific species ([25]), and a greater diversity of forest structures ([4]).
Each tree species has a specific range of environmental conditions that are suitable for its growth. One of the key environmental factors is light, which is also closely related to both surface and air temperature ([34]). Tree species with pioneer traits tend to establish in sunny habitats ([55], [39]). On the other hand, natural regeneration of shade-demanding species often avoids intensively irradiated locations and can survive with reduced light availability for a certain period of time ([26]). The impact of radiation intensity on vegetation influenced by both the quantity and quality of the light, as well as the ratio of direct to diffuse radiation. Under forest cover, diffuse radiation prevails. Moreover, tree crowns affect the overall spectral image of radiation ([29], [38]), which also influences plant photosynthetic processes ([10]).
The properties and photosynthetic activity of plant tissues tend to adapt to the light conditions of their growth environment ([11]). To introduce shade-demanding species, such as Silver fir, in large clear-cut areas, a two-phase forest regeneration approach is recommended ([9], [30]). A nurse stand formed by pioneer tree species improves conditions for shade-loving or shade-tolerant species, allowing for their more successful natural or artificial regeneration beneath. Pioneer tree species also modify soil nutrient availability ([31]). Such preparatory stands need special management to minimize competition and enhance the growth of the target underplanted trees ([16], [22]). This management must cope with sprouting (rapid forming of stump or root sprouts) of the nurse tree species ([17]).
Releasing and tending interventions in a preparatory forest stand, even when intended to support target trees, increases light penetration beneath the canopy. ([15], [35]). Underplanted trees must respond to these changes. In the case of intensive canopy changes, this response can also be associated with tree stress ([1], [2]). One of the common reactions to stress by irradiance is photoinhibition, where increased light exposure intensifies processes that reduce the efficiency of solar energy conversion into electron transport ([49]). Furthermore, the morphology of new leaves can be affected, as plants often increase leaf mass per area (LMA) in sun-exposed leaves ([6]). Light plays a crucial role in determining the metabolic activities of plants, influencing nutrient uptake, assimilation, and distribution within the leaves, which can result in varying nutrient concentrations in plant tissues ([42], [56]).
It is expected that individual underplanted tree species respond specifically to changes in the light environment. The survival and growth of individual underplanted tree species beneath forest canopies have been frequently studied ([20], [8], [46]). However, the response of the photosynthetic apparatus of different tree species grown under the same conditions of tended nurse-crops has not yet been compared.
The most common pioneer species of the nurse-crops in Europe is birch. The main reasons are the frequent presence of individual birch seed trees within forest stands or on forest edges, the extended range of their seed dispersal, and their ability to establish a seed bank that germinates in 1-5 years ([50]). The viability of naturally dispersed seeds of spruces (Picea) and many pines (Pinus) typically extends into the next growing season and rarely into the second growing season ([44]). The growth environment of a birch stand, characterized by reduced forest weed competition and changing topsoil conditions, can enhance the germination and survival of young seedlings of many tree species, including birch itself.
This study aims to evaluate the rate and speed of adaptation of three target conifers to changes in irradiation while growing in different treatments of a birch preparatory stand. Scots pine (Pinus sylvestris), European silver fir (Abies alba), and Norway spruce (Picea abies) were investigated, and their needles were analyzed in terms of morphology, chemistry, and chlorophyll a fluorescence parameters. The working hypotheses were that (i) new needle properties would tend to adapt directly to the current light conditions, even when growing from the buds formed in a previous environment, and (ii) each species would adapt to every tested treatment specifically but adequately.
Material and methods
Study site and experimental design
The experiment was conducted in NE Bohemia (Czech Republic) on the Nemojov research plot (50.48° N, 15.77° E). The plot covers more than 6 ha of a formerly clear-felled area, which originated with hurricane Kyrill in 2007 (Fig. 1, Fig. 2). The wood biomass was removed from the clearing, and individual windthrown stumps were straightened. Permian sandstone forms the geological basis of the site, on which luvic cambisol has formed. The type of natural vegetation community is predominantly Fagetum illimerosum-acidophilum (4I), according to the Czech System of Forest Sites ([54]). The original mature stand before Kyrill was dominated by spruce, with individual mixtures of pine, oak, birch, and aspen ([43]).
Fig. 1 - The location and layout of the Nemojov experimental plot overlaid on an aerial photograph from 2021. Map of Europe taken from ⇒ https://alabamamaps.ua.edu/contemporarymaps/world/europe/index.html.
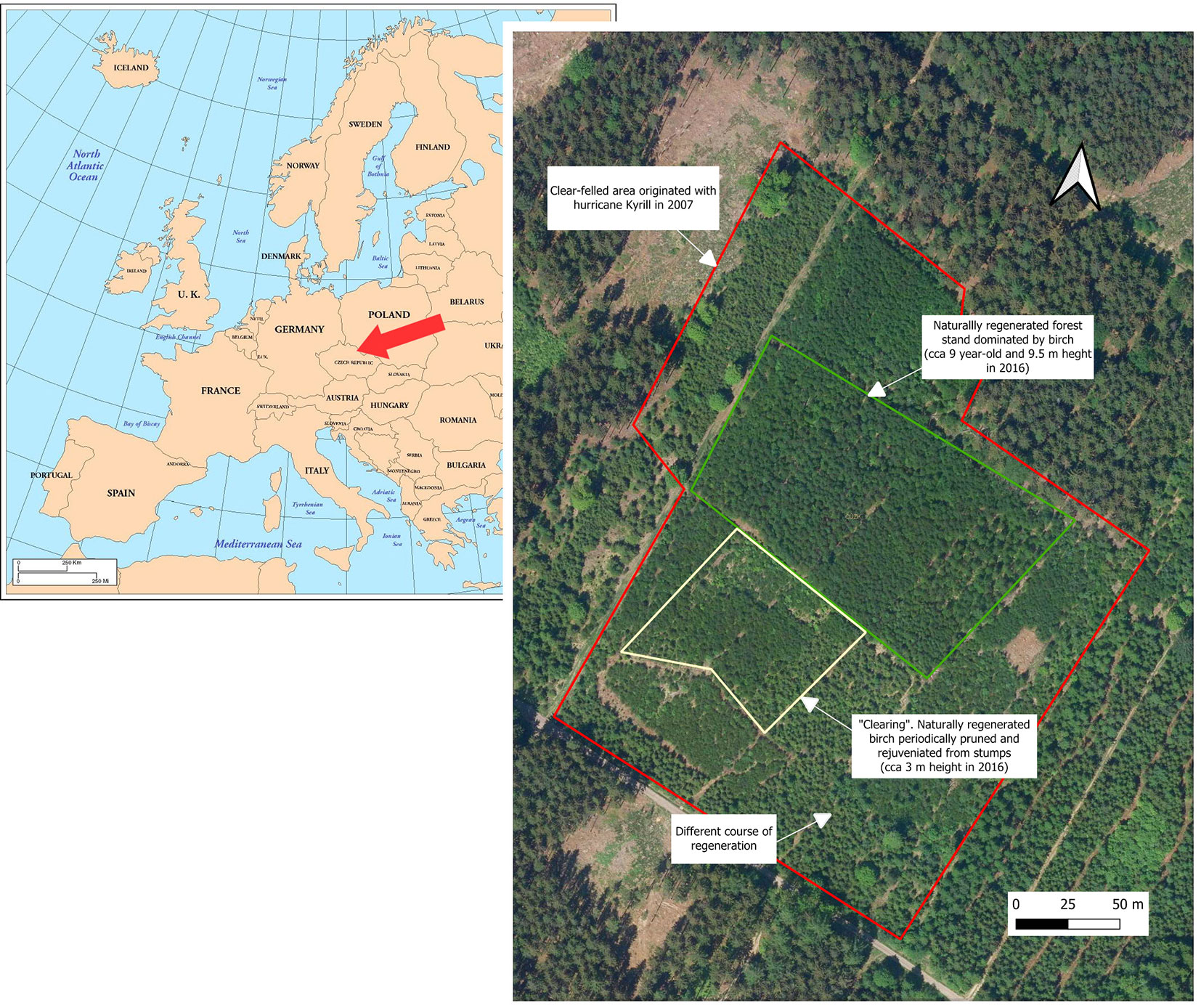
Fig. 2 - Experimental plot genesis schedule (above) and establishment of the treatments and investigations schedule (below).
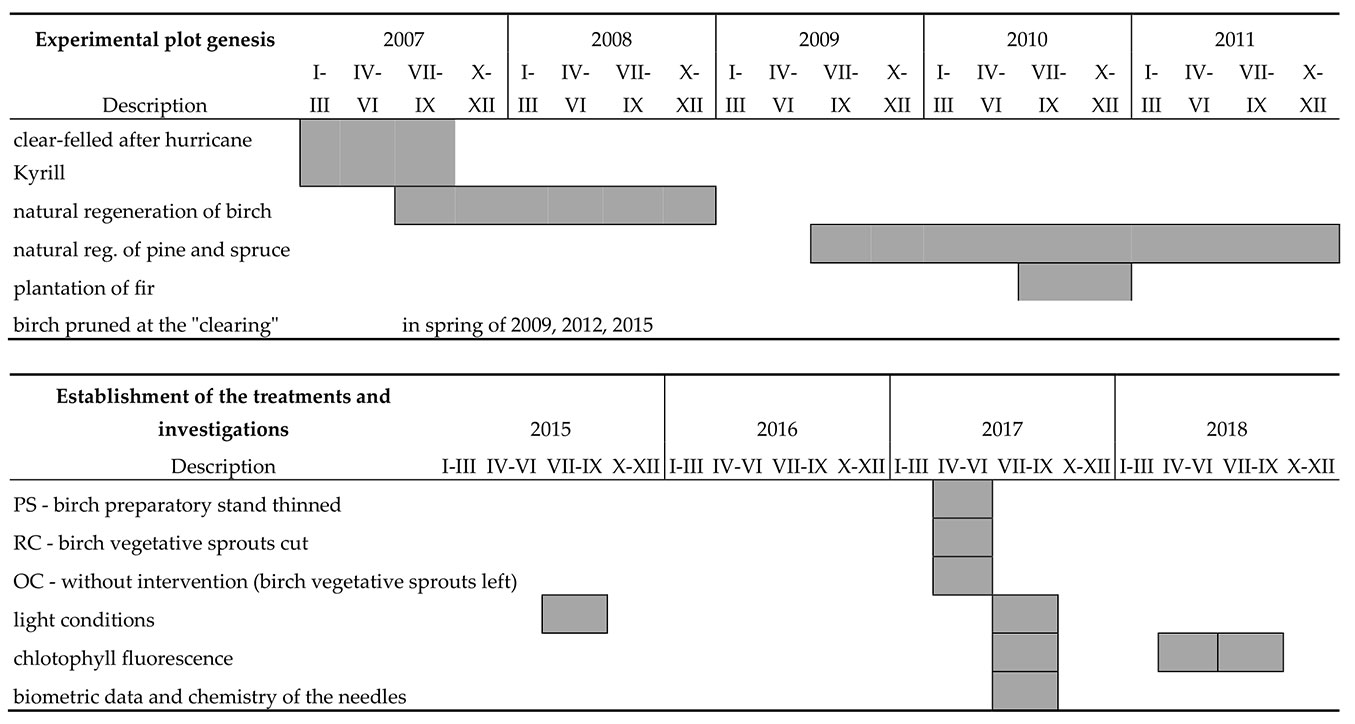
The experimental plantings were carried out under an approximately 9-year-old birch stand created by natural regeneration, which had a height of around 10 m in 2017, and in an adjacent “clearing”. In the clearing, the naturally regenerated birch trees were periodically pruned, but repeatedly rejuvenated from stumps (Fig. 1, Fig. 2). At the time the experiment was set up (2017), the birch there formed a stand of around 3 m in height with a relatively dense canopy.
Firs were planted both under the birch stand and in a periodically restored clearing 3 years after the natural regeneration of the birch (in 2010). The spruce and pine trees originated from natural regeneration from a similar period (Fig. 2). At the time of the experiment (2017), the pines were from 1.8 to 2.8 m tall, the firs between 0.3 and 0.9 m, and the spruces between 0.5 and 1.4 m.
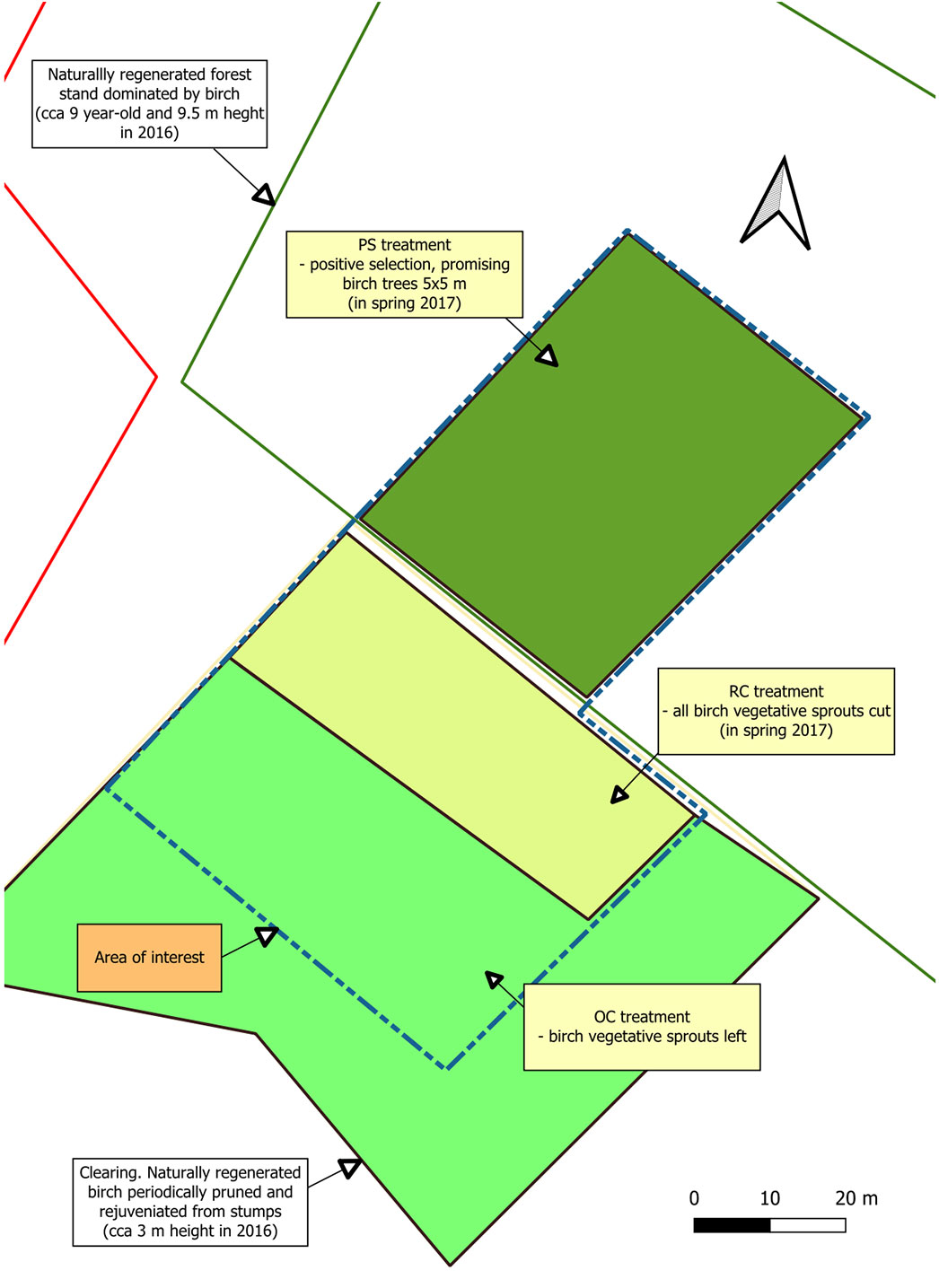
In spring 2017, before the start of the experiment, the birch preparatory stand was thinned over an area of 40 × 50 m (0.2 ha). A positive selection of promising birch trees in a patch of approximately 5 × 5 m was carried out. The birches were released by 2 to 5 competing trees, and the understory was left in place. The treatment was designated the birch preparatory stand (hereafter PS treatment). During the same period, all birch vegetative sprouts were cut in an area of 20 × 60 m of the overgrown clearing (NW-SE orientation of the cut), thus restoring the character of the clearing (restored clearing: RC). In the neighboring area, the birch sprouts were left (areas cleared of overgrown birch sprouts: OC - Fig. 2, Fig. 3).
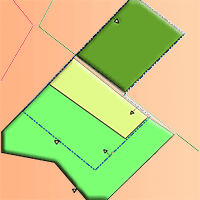
Fig. 3 - Detailed area of interest (compare with Fig. 1) and the three experimental treatments following establishment in 2017. Treatments: PS = birch preparatory stand; RC = restored clearing; OC = clearing overgrown by birch sprouts.
Soil of the OC and PS treatment was sampled in 2022, i.e., 5 years after the release of PS. The L+F soil horizon was analyzed for total nitrogen (Kjeldahl method) and other macronutrient content, according to Zbíral ([57]). The F+H and A horizons were analyzed for pH, nitrogen and macronutrients (Mehlich 3 method - [24]). Three soil pits were randomly selected within each treatment. Data were analyzed in R v. 4.4.2 ([33]) using a t-test. Significant differences between the treatments were found only in the L+F horizon, where PS had higher concentrations of P (0.02) and K (0.005 - Tab. 1).
Tab. 1 - Soil acidity and nutrient content of PS and OC treatments in 2022. (PS): preparatory stand; (OC): overgrown clearing.
| Horizon | Treatment | pH (H2O) |
N | P | K | Ca | Mg |
|---|---|---|---|---|---|---|---|
| mg kg-1 | mg kg-1 | mg kg-1 | mg kg-1 | mg kg-1 | |||
| L+F | PS | - | 17.763.2 | 1200.0 | 1566.7 | 6153.3 | 1230.0 |
| OC | - | 18.407.2 | 1036.7 | 1033.3 | 6750.0 | 1213.3 | |
| F+H | PS | 4.76 | 1.4 | 27.3 | 498.0 | 1806.7 | 328.7 |
| OC | 4.89 | 1.5 | 24.0 | 494.0 | 1792.7 | 426.0 | |
| A | PS | 4.63 | 0.7 | 6.3 | 239.3 | 792.7 | 147.7 |
| OC | 4.80 | 0.9 | 5.7 | 286.7 | 802.0 | 222.0 |
Parameters analysed
Light conditions
In late summer 2015 (i.e., before the canopy release in PS and the establishment of RC) and in late summer 2017, the light conditions were analyzed using the hemispherical photography method (Fig. 2). Photographs were taken in each environment (treatment) at a height of 130 cm above the ground at a regular distance of 3 × 3 m. The equipment used included: Olympus SP-570UZ® camera, DÖRR Digital Professional 0.19× DHG® Fisheye lens, and Olympus C5060 48.5-52® adapter and self-leveling tripod ensuring the vertical position of the lens. Between 11 and 35 pictures were taken for each treatment on both dates, considering the variability of the light regime.
The photos were retouched and thresholded through the software ImageJ v. 1.52d using the Minimum method ([32], [3]), which provided the most accurate results for the given lighting conditions. The images were then analyzed using the Gap Light Analyzer v. 2.0, and the process was automated using the AutoHotKey application (AutoHotkey Foundation LLC, Indiana, USA). The vegetation period was entered as the period from 15/4 to 15/10; the values for Cloudiness Index, Spectral Fraction, and Beam Fraction were left at the default value (0.5).
The evaluated light environment parameters were: Canopy openness (c.o.), LAI 4Ring (effective leaf area index integrated over zenith angles 0 to 60° - [45]), and the proportion of direct (transmitted direct, tdt), diffuse (transmitted diffuse, tde), and total penetrating radiation (transmitted total, tt).
Chlorophyll fluorescence
The chlorophyll a fluorescence of the needles was measured using the Plant Efficiency Analyzer® (PEA, Hansatech Instruments, UK) in May 2017 (i.e., before the interventions - treatment establishment). The measurement was repeated when the light conditions changed: in September 2017 on the needles of the new shoots, in June 2018 on the one-year-old needles, and in August 2018 on the new needles (Fig. 2). At least 12 trees from each tree species and treatment were measured in each measurement period. Needles were taken from three different lateral shoots from the upper part of the crown of each tree plant (i.e., three replicates per plant) and individually placed face up in shielding clips. The chlorophyll fluorescence parameters of the dark-adapted needles were measured; samples were shielded for at least 30 minutes before the measurement. The radiation intensity was set to 50% of the maximum, i.e., to approximately 1500 μmol m-2 s-1. The level was chosen to conserve the battery. A test with increasing light intensity confirmed that the value was sufficient for spruce and fir. In contrast, sun-adapted pine needles could reach the light saturation point at higher radiation levels.
The following parameters were analyzed: minimum fluorescence (F0), maximum fluorescence (Fm), and maximum quantum yield of photosystem II in the dark-adapted tissues (Fv/Fm - [23]). The vitality of the plants was characterized through the parameter performance index, which was calculated using the equations and the performance index on absorption basis (PIabs) previously reported by Strasser et al. ([47]).
Biometric data and the chemistry of the needles
In September 2017, new shoots were collected from the upper part of the crown of 10 seedlings of each species in each treatment to make a biometric and chemical analysis of the needles in the first year after the interventions, i.e., after changes in light conditions (Fig. 2). The needles were separated (20 per tree) and their biometric parameters (length, maximum width, area, and perimeter) were determined using digital image analysis methods in ImageJ software v. 1.52d with the ObjectJ extension. In addition, the basic nutrients (N, P, K, Ca, Mg; %) of three mixed needle samples per species (each sample from needles of three to four samplings) were analyzed for each treatment, according to the methodology proposed by Zbíral ([57]).
Statistical analysis
The hemispheric photography data were tested using the Kruskal-Wallis non-parametric test. The biometric data of the needles from individual tree species were analyzed using the Nested ANOVA method, with stand treatment as a fixed effect and individual trees as a random effect. The representation of nutrients in the needles and the chlorophyll a fluorescence parameters of each species in the stand treatments were compared using one-sided ANOVA followed by a post-hoc Tukey test. To ensure normality and stabilize the variances, the Fv/Fm data were transformed using the Box-Cox transformation. The tests were performed in the statistical environment R v. 4.4.2 ([33]). Differences were considered significant when p ≤ 0.05.
Results
Light conditions/environment
Before the canopy opening (2015), the average c.o. of the PS and OC treatments was 15.7% and 19.2%, respectively. After the release (2017), the c.o. of the RC treatment was clearly the highest, as expected, reaching an average of almost 50%. The proportions of tdt and tde radiation in RC also reached about twice the values of the other treatments (Tab. 2). Despite the realized release in PS, the differences between the analyzed light parameters of the other two treatments (i.e., PS and OC) were not significant: the average total c.o. was less than 20%. The minimal increase in c.o. in the PS treatment compared to 2015 was likely due to the nature of the release, which maintained the birch understory.
Tab. 2 - Light characteristics of the canopy of the analyzed stand treatments after release interventions in 2017. (PS): preparatory stand; (RC): restored clearing; (OC): overgrown clearing. Different letters indicate statistically heterogeneous treatment groups within the given parameter (ANOVA, Tukey test). LAI 4Ring: effective leaf area index integrated over the zenith angles between 0° and 60°.
| Treatment | Canopy Open. (c.o.) |
LAI 4Ring (LAI) |
Transmit. direct (tdt) | Transmit. diffuse (tde) | Transmit. total (tt) |
|||||
|---|---|---|---|---|---|---|---|---|---|---|
| % | SD% | Mean | SD | % | SD% | % | SD% | % | SD% | |
| PS | 19.9 b | 4.8 | 1.78 b | 0.44 | 25.5 b | 11.5 | 29.0 b | 6.2 | 27.2 b | 8.2 |
| RC | 49.7 a | 5.5 | 0.61 a | 0.18 | 47.7 a | 14.0 | 63.0 a | 8.1 | 55.4 a | 10.6 |
| OC | 19.4 b | 6.6 | 2.19 c | 0.67 | 24.4 b | 7.0 | 28.6 b | 8.1 | 26.5 b | 7.0 |
A significant difference between PS and OC was only observed for the determined LAI (LAI 4Ring parameter), which was highest in the OC treatment. The medians of c.o., penetrating tde, and tt radiation were about 4% lower in the OC treatment compared to PS, although not significantly different (Tab. 2).
Needle morphology
In the first growing season, which began with the release intervention, the pines reacted to the increase in solar radiation by altering the shape of their new needles. The new needles of the pines from the restored clearing (RC treatment) had a significantly higher maximum width and area than the needles of the pines growing under the preserved canopy of the OC treatment (Fig. 4).
Fig. 4 - Biometric parameters of new pine, fir, and spruce needles according to stand treatments in the first year after the release interventions in 2017. (PS): preparatory stand; (RC): restored clearing; (OC): overgrown clearing. Different letters indicate statistically heterogeneous groups of the parameter for the respective tree species between the treatments (Nested ANOVA).
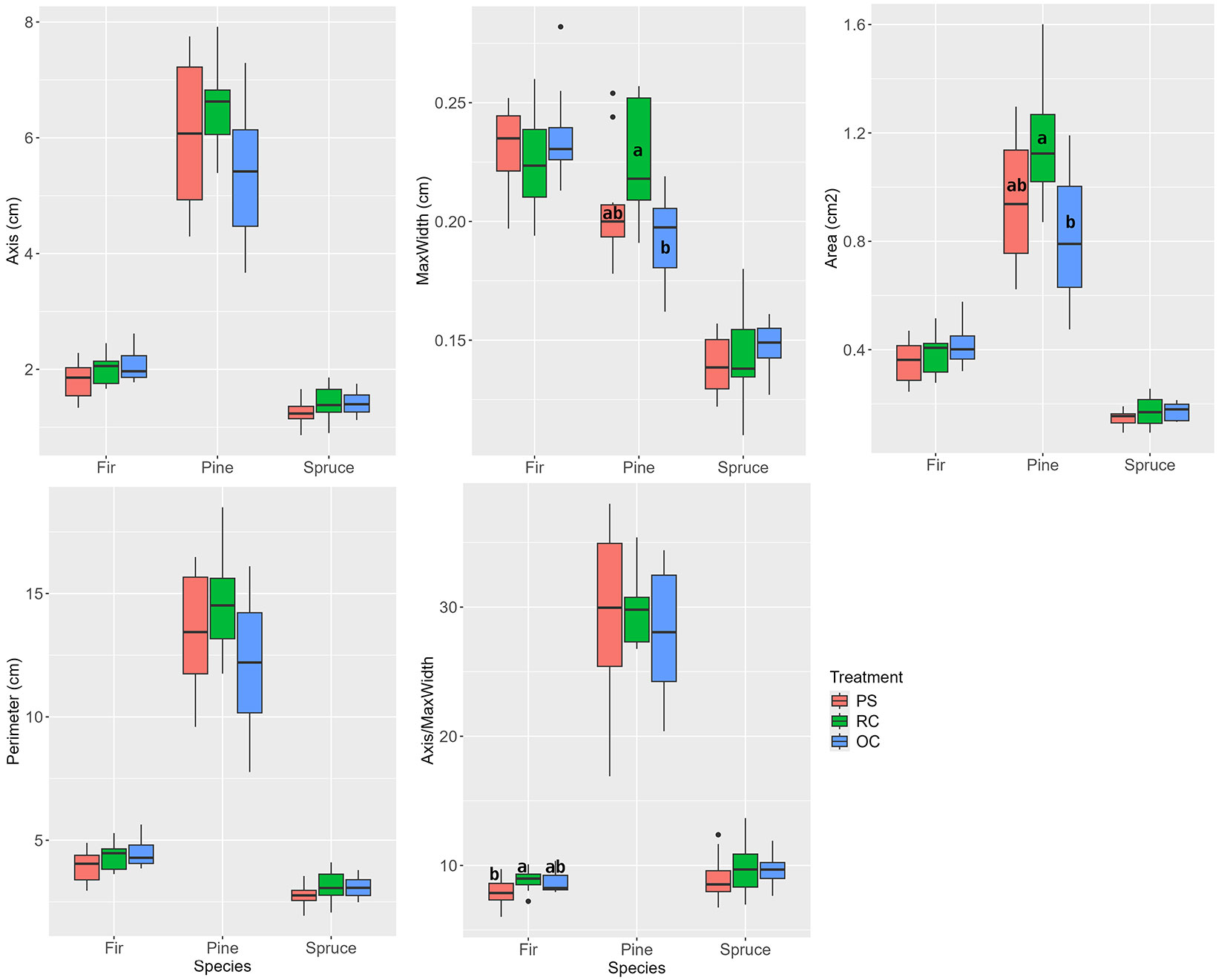
In the firs exposed to the increased radiation of the clearing (RC), the change in conditions led to a significant increase in the ratio of needle length to maximum width as compared to the PS. The dimensions of the needles of the new spruce shoots were not significantly affected by the increase in light conditions (Fig. 4).
Needle chemistry
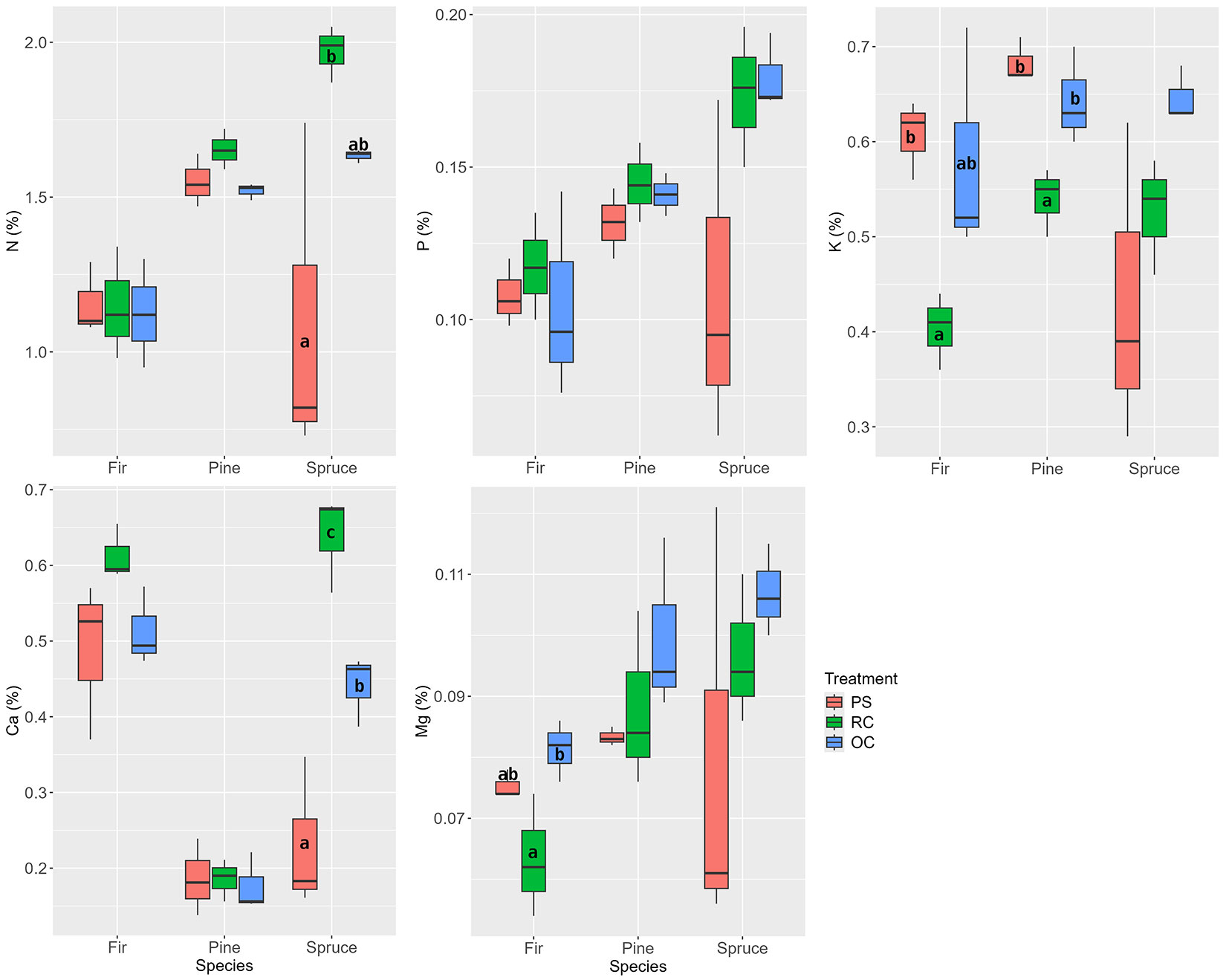
The release was reflected in the content of nutrients in the one-year-old needles. Compared to the other treatments, the pine needles in the RC treatment had a significantly lower K content. In fir, lower K and Mg contents were found in the RC treatment, but the differences were only significant in one of the other stand treatments. The spruce in the RC treatment, compared to the OC treatment, responded to the higher irradiance with a significant increase in N and Ca contents (Fig. 5).
Fig. 5 - Nutrient contents in one-year-old needles of pine, fir, and spruce according to stand treatments in the first year after the release interventions in 2017. (PS): preparatory stand; (RC): restored clearing; (OC): overgrown clearing. Different letters indicate statistically heterogeneous groups of the parameter for the respective tree species between the stand treatments (ANOVA, Tukey test).
Chlorophyll a fluorescence
The specific environmental conditions of the individual treatments before the release interventions (2015) showed treatment-specific differences in the values of Fo and Fm for pine and fir (Fig. 6). However, the maximum quantum yield Fv/Fm of all three tree species showed no significant differences between the stand treatments. On the other hand, PIabs values showed significantly higher photosynthetic efficiency in pines growing in the shadier PS treatment, whereas spruces had higher efficiency in the OC treatment.
Fig. 6 - Chlorophyll fluorescence parameters before the release (May 2017). (PS): preparatory stand; (RC): restored clearing; (OC): overgrown clearing. Different letters indicate statistically heterogeneous groups of the parameter for the respective tree species between the stand treatments (t-test).
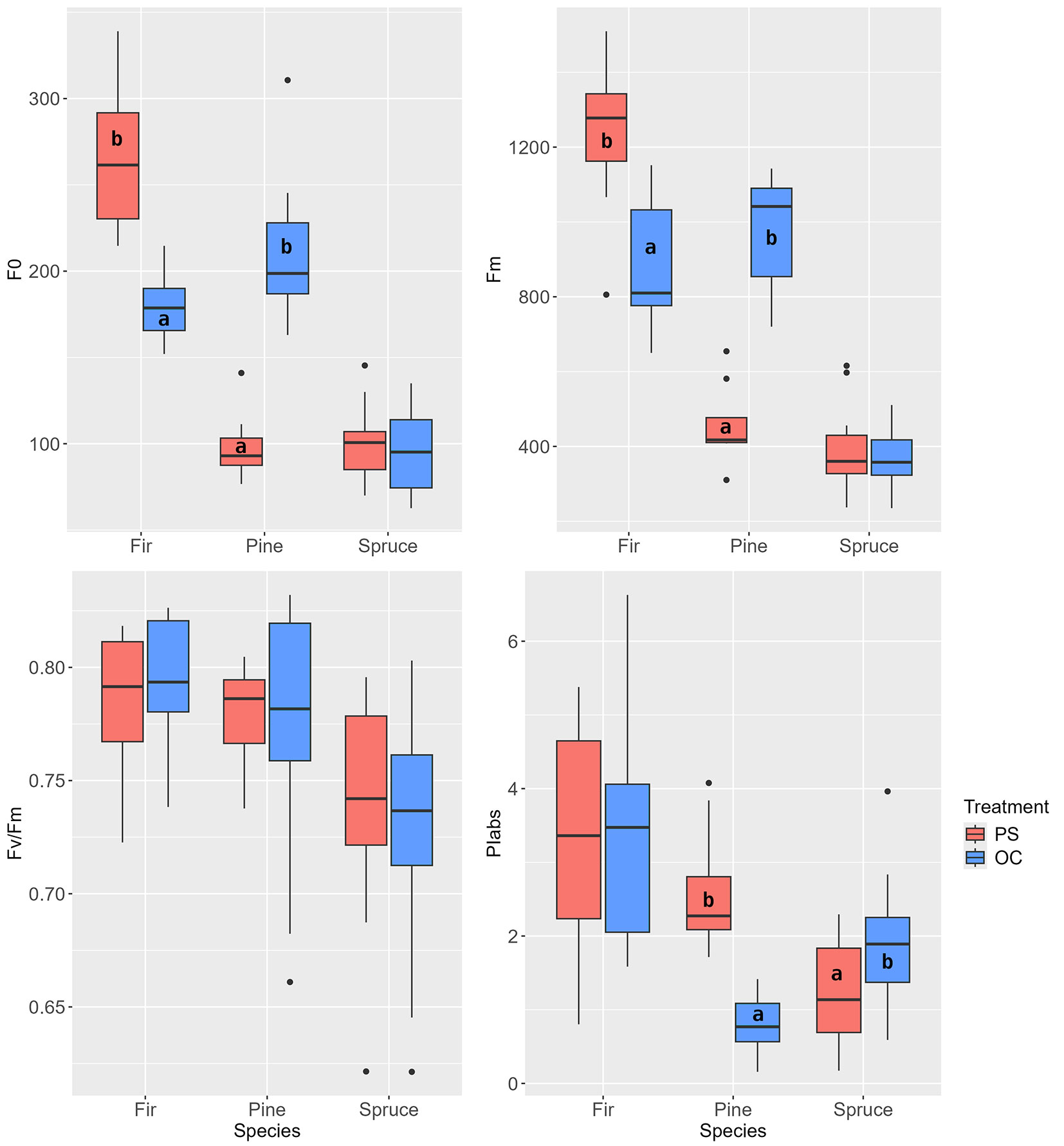
At the end of the first growing season after the release interventions (2017), significant differences between the individual stand treatments were only observed in the new needles of the pines and spruces (Fig. 7). In pine, the Fv/Fm parameter (also Fm) was highest in the PS treatment. According to the PIabs parameter, the trees from the PS and RC treatments grew better than those from the OC treatment. In spruce, the differences were only evident in Fv/Fm, with values for the PS and OC treatments being higher than in RC. For all tree species, the average PIabs value was highest in the RC treatment, but the differences were only significant for pine (Fig. 7).
Fig. 7 - Chlorophyll fluorescence parameters at the end of the first growing season after the release (September 2017). PS - preparatory stand, RC - restored clearing, OC - overgrown clearing. Different letters indicate statistically heterogeneous groups of the parameter for the respective tree species between the stand treatments (ANOVA, Tukey test).
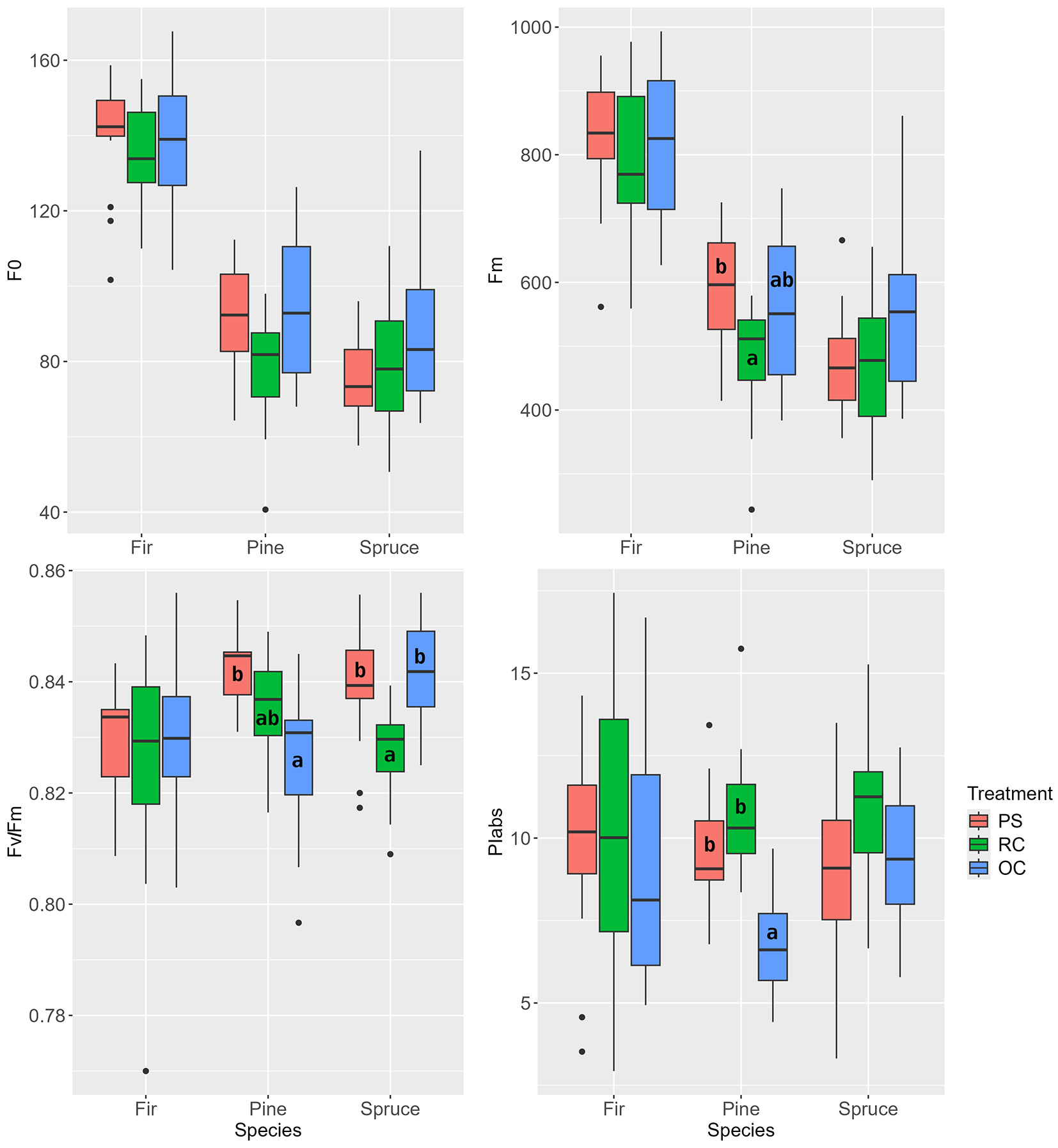
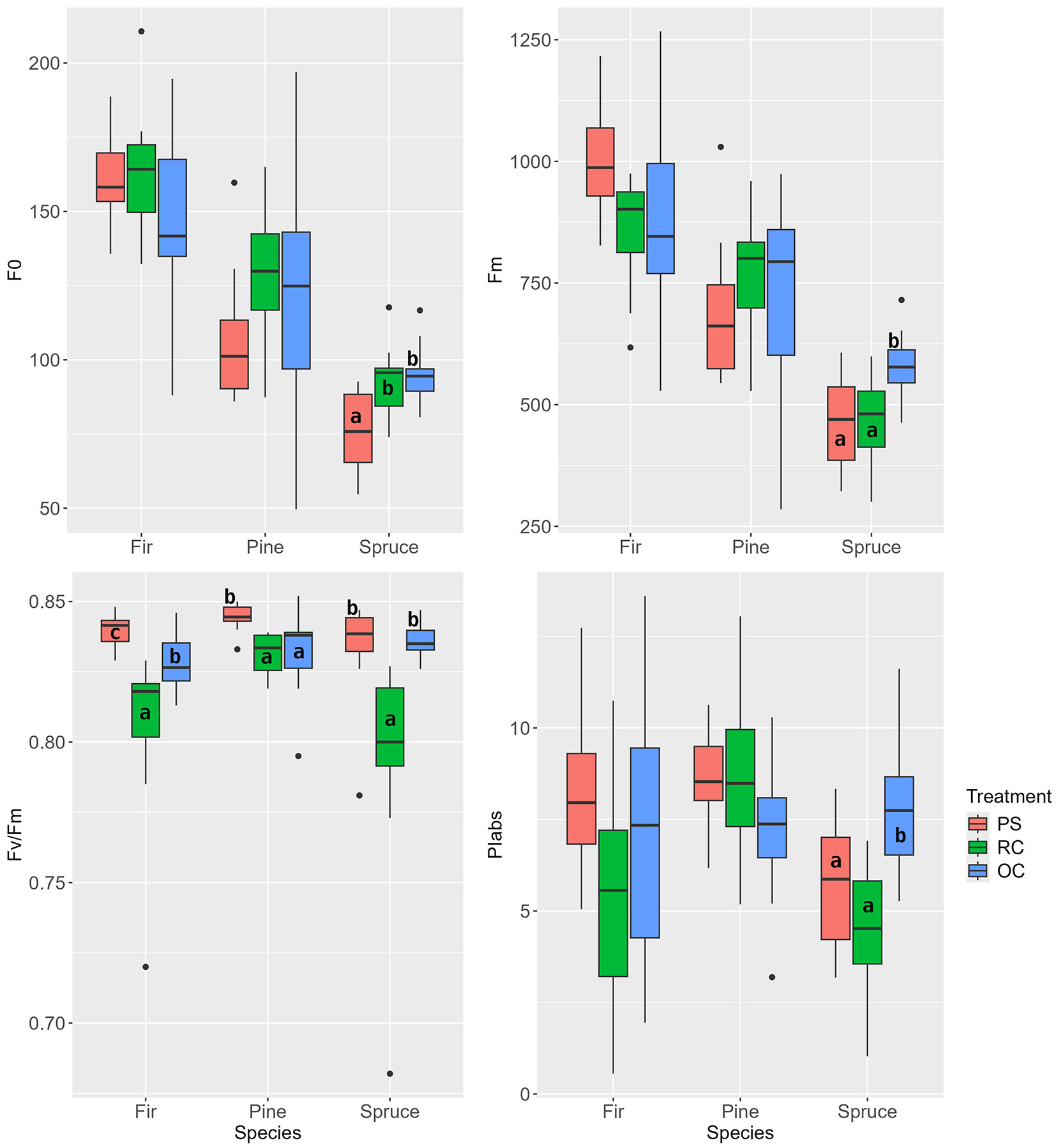
At the time of measurement at the beginning of the second vegetation period after the interventions, significantly lower Fv/Fm and PIabs values were observed in the RC treatment for one-year-old needles of all tree species (also Fm values for fir and spruce - Fig. 8). For pine needles, a decrease in the Fv/Fm ratio was observed with increasing irradiance in the following order: OC>PS>RC. In contrast, the Fo values showed the opposite trend. In pine, the Fv/Fm parameter indicated more intensive photosynthesis in the OC treatment. The PIabs index showed a similar trend in spruce, while the difference between PS and OC treatments could not be confirmed in fir.
Fig. 8 - Chlorophyll a fluorescence parameters of one-year-old needles at the beginning of the second growing season after the release (June 2018). (PS): preparatory stand; (RC): restored clearing; (OC): overgrown clearing. Different letters indicate statistically heterogeneous groups of the parameter for the respective tree species between the stand treatments (ANOVA, Tukey test).
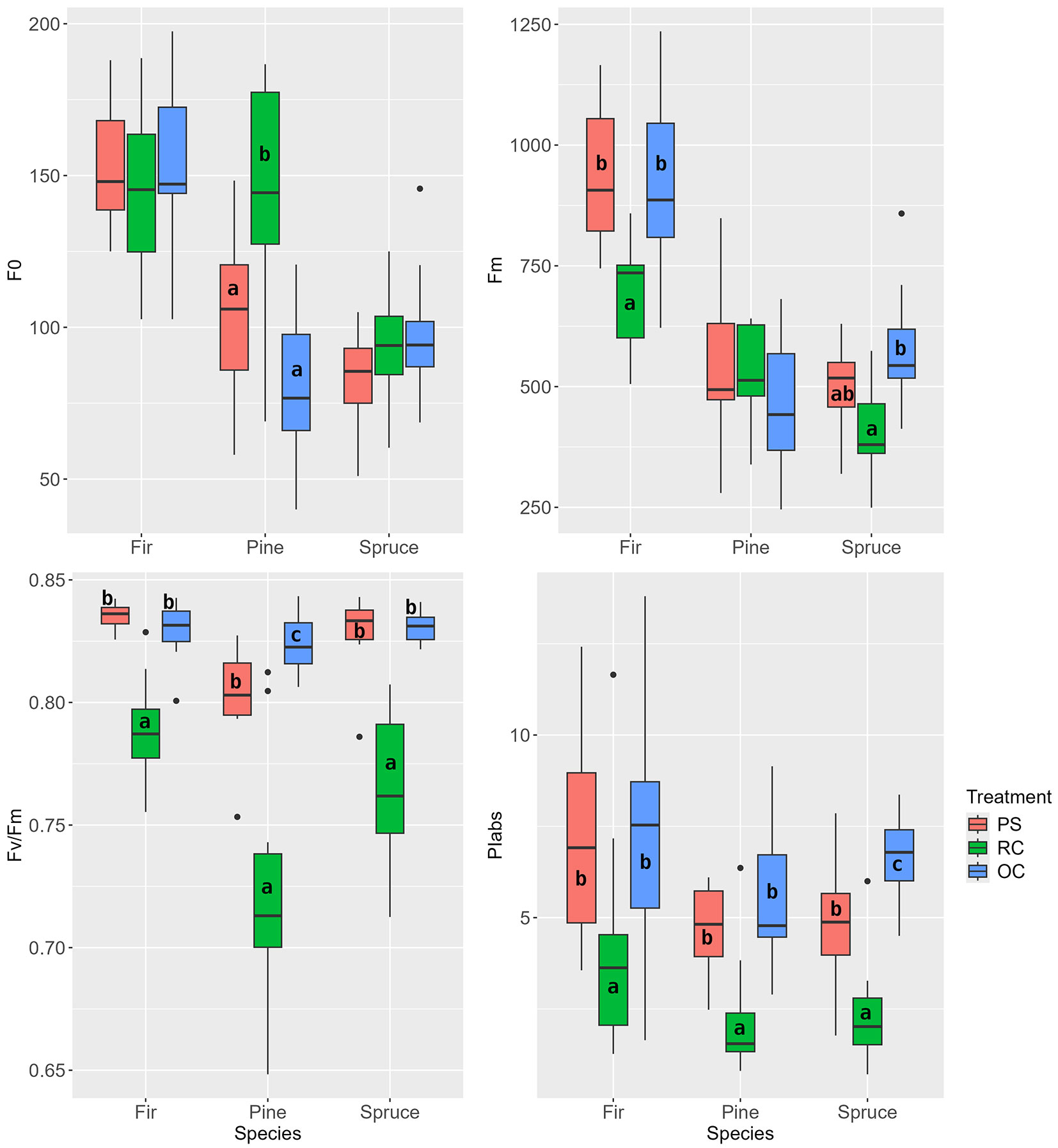
At the end of the second vegetation period after the interventions, the new needles of the pine in the PS treatment showed significantly higher Fv/Fm values. In contrast, the value decreased in fir in the order PS>OC>RC (Fig. 9). Spruce showed differences in all evaluated parameters: the Fv/Fm and PIabs values indicate a significantly lower performance/efficiency of chlorophyll fluorescence in spruce under the RC treatment at the given time.
Fig. 9 - Chlorophyll fluorescence parameters of new needles at the end of the second growing season after the release (August 2018). (PS): preparatory stand; (RC): restored clearing; (OC): overgrown clearing. Different letters indicate statistically heterogeneous groups of the parameter for the respective tree species between the stand treatments (ANOVA, Tukey test).
Discussion
Improving the environmental conditions in favor of the target species is the main objective of silvicultural measures in preparatory forest stands ([16], [40]). In addition, depending on the chosen methodology, a positive effect on promising trees in the upper layer can also be expected. In our experiment, this was the aim of the PS treatment. Increasing irradiation below the canopy also changes the temperature and the humidity regime inside the stand ([12]).
The interventions increased c.o. by 30% (RC) and 4% (PS), while OC remained unchanged as its canopy developed naturally. The restored clearing (RC) showed the highest c.o. and a significant increase in both transmitted direct (tdt) and total radiation (tt), indicating a dramatic change in light conditions. The hemispherical photography-derived parameters of the PS and OC treatments were found to be comparable, except for the LAI 4Ring parameter, which was highest for OC. The estimated effective LAI is smaller than the actual leaf area index due to leaf clumping ([45]), yet it shows the mutual relations. The LAI may be responsible for the spectral limitation of light under the tree canopy, as the leaves absorb strongly in the red and blue regions of the visible spectrum due to chlorophyll ([14]), while they reflect in the near infrared region ([53]). This could also affect specific reactions of the target species.
The morphological adaptation of the needles in the first year after the release of the canopy was most evident in Scots pine. This species is the most light-demanding of those studied, and therefore, the highest gain could be expected from the release. Increasing the total area of needles may be one of the adaptations to benefit from higher irradiance. Niinemets et al. ([27]) also observed that the width and length of pine needles were positively correlated with the integrated quantum flux density. Sun leaves are usually smaller than shade leaves ([21]); however, this was not confirmed in our study. The question arises as to the reason for the highest variability in the Axis/MaxWidth ratio in firs with an unchanging environment (OC). The OC conditions were relatively unique: the standard deviation of the c.o. was only insignificantly higher than the others.
With pine, it is assumed that the general anatomy of the new shoots fixed in buds formed in the shady conditions of the previous year limits the number of new needles, and pine aims to increase the total leaf area by needle elongation. With evergreen trees, the morphology at the leaf- and shoot-levels reflects the irradiance level prevailing at the time of shoot elongation ([41]). In our experiment, the highest morphological plasticity to the light environment (or stability) was observed in spruce, whose new needle shapes did not respond significantly.
Similar trends in species responses to needle morphology were also expected in the nutrient concentrations; however, the nutrient concentrations showed completely different patterns. Pine needles reacted to the intense release at RC only with a reduced K concentration, and firs lowered both K and Mg concentrations, but the values remained in the optimal range ([51]). This is the opposite result to that of Knapp et al. ([19]), who found that foliar N, P, and K concentrations of longleaf pine seedlings in mature stands of Pinus taeda generally increased with the intensity of the harvest treatment.
Furthermore, there was no difference in foliar nutrients between the maintained canopy (OC) and the light release (PS) in the pines and firs in our study. This may be due to the similar resulting light conditions prevailing in these two treatments (Tab. 2). Soil nutrients of these two treatments differed in L+F horizon only, the higher P and K concentrations of PS (Tab. 1) did not reflect in significant differences of nutrients in needles of any species (Fig. 5). In contrast, the needles of the released spruces showed increased concentrations of N (in agreement with [19]) and Ca. The N concentration of the PS treatment was below the limit of deficiency, while the average value in RC exceeded the limit of surplus ([51]). However, the Ca content in spruce needles increased in the order PS<OC<RC, i.e., the lowest Ca values were in the lightly released PS treatment. This probably indicates other factors besides the current year’s irradiation affecting Ca nutrition. The value in RC was also in the surplus range. Preparatory birch stand positively affects nutrient availability in the soil, particularly in the forest floor ([52], [13]). It can be expected that this effect gradually fades in the years following intensive canopy opening. Therefore, Ca availability in soil during the first year after the release may vary across different parts of the preparatory stand more than in the year of soil sampling, which could be reflected in Ca accumulation in spruce needles.
In the chlorophyll a fluorescence parameters, the slightly different initial light conditions (by 4% of c.o.) reflected in the treatment differences in the minimum and maximum fluorescence levels of fir and pine. Whereas Silver fir had higher values in the PS, Scots pine had higher values in the OC treatment (Fig. 6). Fluorescence parameters of the new needles after the release measure were species-specific. This finding can also express the diverse preference for irradiation during the youth stage of these two species, similarly to the new needle anatomy. PIabs, the summarizing parameter, indicated that pines performed better in more open environments, which aligns with Brichta et al. ([5]), who found that pine natural regeneration in a shelterwood system reacts positively to canopy release. Paradoxically, spruce had higher PIabs in the denser OC treatment, indicating better photosynthetic efficiency at the time of measurement there. For both summarizing parameters (Fv/Fm, PIabs) of Silver fir, the differences in irradiance did not manifest in the first year, but only in the second year after intervention. The state of the second year is in line with a previous study comparing shaded and sun-exposed firs; it confirmed higher Fv/Fm in shaded trees ([43]).
In the second vegetation period after the interventions, all three species indicated a mild stress reaction in the most open treatment (RC). During the night before the measurement (from 6 to 7 June 2018), the minimum ground air temperature in the open dropped to 5.4 °C, while the minimum was 9.1 °C under the dense birch stand (control plot). The difference at the time of the measurement was also significant: the temperature was around 27 °C under the birch, while it had already risen to 34 °C at the RC treatment. Temperature, especially temperature extremes, impacts chlorophyll fluorescence ([48]). Interestingly, Scots pine was the most irradiance-dependent when Fv/Fm was considered, with a negative correlation of the value to the respective treatments - the more light, the lower the value (Fig. 8). However, there is a risk of limiting the chosen saturation pulse intensity, which could be insufficient to reach the maximum fluorescence level of sun-adapted pine needles of RC treatment.
New needles of the trees growing in the highest irradiance (RC) also showed the lowest Fv/Fm values at the end of the second vegetation period after interventions. Still, these values were not within the range responding to stress. The question is whether the higher value in OC compared to the PS treatment for Silver fir is related to the different light parameters of these treatments. Similarly, there were statistically identical values in RC and OC for Scots pine (Tab. 2, Fig. 9). Both could respond to the physiological acclimatization of the new needles to the light conditions of the respective treatment. Woody plants require sufficient light for photosynthesis, but they also protect themselves from excessive solar energy ([28]). For Scots pine, higher irradiance is beneficial, whereas spruce might thrive better under slightly denser canopy conditions ([36]). Shade-demanding species like Silver fir may require a more cautious approach to canopy opening, ensuring gradual acclimatization to avoid stress responses ([37]). A gradual canopy release may help trees better acclimatize to increased light, reducing temperature stress and potential damage from sudden exposure, which benefits fir in particular ([7]).
Under the conditions of the tested treatments in our experiment, all species successfully acclimated, which is a promising sign for their prosperity. It is evident that in two-phase forest regeneration, pioneer light-loving tree species can be successfully complemented by target tree species with higher shading requirements. These results corroborate that environmental changes made by birch management were realized to a suitable extent to benefit the target species’ undergrowth. Future canopy release and tending interventions should also be planned carefully to avoid excessive stress due to sudden increases in light availability, especially for Silver fir. This approach can enhance the stability and resilience of future forests, particularly within the context of changing climatic conditions.
Conclusions
This study investigated the effects of different environmental treatments in a young birch forest on the needle morphology and photosynthetic performance of the undergrowth of Silver fir, Norway spruce, and Scots pine. The reactions of the species to different levels of canopy opening showed: (i) each species exhibited some degree of direct morphological adaptation of its new needles to changes in light conditions already during the first year after the release of the nurse stand canopy. Silver fir and Scots pine showed more significant changes, while Norway spruce needles remained relatively stable. (ii) All species showed changes in nutrient concentrations in needles in response to increased irradiance, though with different patterns. Silver fir and Scots pine showed a decrease in certain nutrients (K, Mg for fir; K for pine), while Norway spruce showed an increase in N and Ca in the most open treatment. (iii) According to the chlorophyll a fluorescence parameters of the needles, all species tended to be less efficient in photosynthesis when exposed to increased irradiance both during the first and second years after release. In the first summer after release, new needles of Scots pine benefited the most from the increased irradiation. Shade-loving Silver fir showed similar patterns of physiological acclimatization to spruce and pine.
The results indicate a high plasticity of the studied species to changes in light conditions of the nurse stand, which leads to adaptations in morphology, nutrient status, and physiology of new needles as early as the first year after changes in the nurse canopy. Although the means of adaptation were specific for each species, new conditions did not limit their adaptation potential. Increasing canopy openness of the birch stand from 16% to 20% and from 19% to 50% was positively accepted by two environmentally different species, Scots pine and Silver fir.
Acknowledgements
The study was supported by the Ministry of Agriculture of the Czech Republic, the institutional support no. MZE-RO0123. The author thanks librarian Mgr. Jitka Součková for her assistance with the citation format in the reference list. The author also thanks theBestTranslation agency for language proofreading.
References
Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Online | Gscholar
Gscholar
Online | Gscholar
Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Forestry and Game Management Research Institute, Research Station at Opočno, Na Olive 550, 517 73 Opočno (Czech Republic)
Corresponding author
Paper Info
Citation
Špulák O (2025). Needle traits of understory Silver fir, Norway spruce, and Scots pine in response to increased canopy openness in a birch-dominated stand. iForest 18: 273-282. - doi: 10.3832/ifor4909-018
Academic Editor
Claudia Cocozza
Paper history
Received: May 27, 2025
Accepted: Sep 16, 2025
First online: Oct 14, 2025
Publication Date: Oct 31, 2025
Publication Time: 0.93 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2025
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 4528
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 2454
Abstract Page Views: 1028
PDF Downloads: 888
Citation/Reference Downloads: 0
XML Downloads: 158
Web Metrics
Days since publication: 284
Overall contacts: 4528
Avg. contacts per week: 111.61
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
(No citations were found up to date. Please come back later)
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Light acclimation of leaf gas exchange in two Tunisian cork oak populations from contrasting environmental conditions
vol. 8, pp. 700-706 (online: 08 January 2015)
Short Communications
Effect of four levels of shade on survival, morphology and chlorophyll fluorescence of Nothofagus alessandrii container-grown seedlings
vol. 8, pp. 638-641 (online: 08 January 2015)
Research Articles
Wintertime photosynthesis and spring recovery of Ilex aquifolium L.
vol. 12, pp. 389-396 (online: 31 July 2019)
Research Articles
Growth and physiological acclimation to shade in young plants of Adesmia bijuga Phil., a critically endangered species in central Chile
vol. 14, pp. 307-312 (online: 01 July 2021)
Research Articles
Magnolia grandiflora L. shows better responses to drought than Magnolia × soulangeana in urban environment
vol. 13, pp. 575-583 (online: 07 December 2020)
Research Articles
Effects of substrate and ectomycorrhizal inoculation on the development of two-years-old container-grown Norway spruce (Picea abies Karst.) seedlings
vol. 8, pp. 487-496 (online: 10 November 2014)
Research Articles
Effects of mild drought on the morphology of sun and shade needles in 20-year-old Norway spruce trees
vol. 12, pp. 27-34 (online: 10 January 2019)
Research Articles
Relationships between leaf physiognomy and sensitivity of photosynthetic processes to freezing for subtropical evergreen woody plants
vol. 12, pp. 551-557 (online: 17 December 2019)
Research Articles
Growth, morphology, and biomass allocation of recently planted seedlings of seven European tree species along a light gradient
vol. 13, pp. 261-269 (online: 03 July 2020)
Research Articles
Calibration of a multi-species model for chlorophyll estimation in seedlings of Neotropical tree species using hand-held leaf absorbance meters and spectral reflectance
vol. 9, pp. 829-834 (online: 17 May 2016)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







