
Growth and physiological acclimation to shade in young plants of Adesmia bijuga Phil., a critically endangered species in central Chile
iForest - Biogeosciences and Forestry, Volume 14, Issue 4, Pages 307-312 (2021)
doi: https://doi.org/10.3832/ifor3640-014
Published: Jul 01, 2021 - Copyright © 2021 SISEF
Research Articles
Abstract
Adesmia bijuga Phil. is an endemic and endangered shrub species of central Chile. Its potential shade intolerance is one of the leading hypotheses for its vigor loss when the species grows beneath closed canopies. The objective of this study was to assess the growth and physiological acclimation to the shade of young plants of A. bijuga. A nursery experiment was established with three light levels based on the interception of photosynthetic active radiation (PAR) (TRT0: control at full sun; TRT60: 60% shaded; TRT90: 90% shaded), and maintained for 71 days during the summer season. Growth and leaf morpho-physiological responses were evaluated at the beginning, at the middle, and at the end of the experiment. The shading treatment increased plant height (H), live crown percentage (Lcrown), and specific leaf area (SLA) compared to the control treatment at full sun. However, light-saturated photosynthetic rate (Amax), dark respiration rate (Rd), and light compensation point (Γi) were higher in TRT60 than in the other treatments, while no differences were found among treatments for the apparent quantum yield (α). At this stage of plant development, our results suggest high acclimation plasticity of A. bijuga to light levels; however, a semi-shade environment (i.e., TRT60) favored a better performance of the species.
Keywords
Adesmia bijuga, Shade Tolerance, Photosynthesis, Light Acclimation, Forest Restoration
Introduction
Central Chile (30°-38° S) contains more than 60% of the country’s total flora and endemic species, and is listed among the 25 global biodiversity hot spots due to its high endemism and loss of habitat ([20], [8]). Currently, while this area has a highly fragmented landscape, due to the extent of Pinus radiata D. Don plantations, it still contains some important natural biocoenoses such as the Maulino forest, the sclerophyllous forest, and species of the ecologically important South American genus Adesmia, whose populations are characterized by small size and low genetic diversity ([8], [10]).
The genus Adesmia (Fabaceae) comprises 230 species distributed in South America ([2]). In Chile, there are 130 species, of which 85 are endemic ([19]) with social and agricultural importance ([6]). Adesmia bijuga Phil. is one of the most endangered species of the genus, as it is spatially restricted over an area of 4 ha in the Maule Region of Central Chile ([9]). Despite the species has been listed as “Critically Endangered” by the International Union for Conservation of Nature (IUCN - [12], [8]), it is not currently protected by the Chilean Government. The species has small compound leaves with leaflets of 28-mm length and 7-mm wide. It has a shrubby habit and can reach up to 2 m height with long and straight branches ([12]).
Currently, the habitat of A. bijuga is very fragmented and largely immersed in P. radiata plantations. Because of its floristic and biogeographical importance, as well as its conservation status, there is an interest in the ecological restoration of the species, but there is still little knowledge about to what extent P. radiata plantations might modify the environmental and ecological conditions of A. bijuga habitat. It is known that some native species might positively benefit from growing beneath P. radiata canopies ([27]), especially shade-tolerant species ([11], [9]). Brockerhoff et al. ([1]) reported a decrease in light-demanding pioneer species and an increase in shade-tolerant species with canopy closure and stands age. In the case of A. bijuga, the species seems to have a better recruitment performance in open sites, but little is known about the specie’s responses to shade when it grows beneath plantation canopies.
Light is the driving force for photosynthesis, and plants acclimate their leaf physiology and morphology to optimize carbon assimilation. Acclimation to irradiance is typically addressed by studying photosynthetic light response curves. Leaf acclimation from high to low irradiance involves a reduction in the light-saturated photosynthetic rate, respiration rate, light compensation point, photoprotective compounds, palisade tissue, mesophyll cells, stomata per unit of leaf area, and chlorophyll a/b ratio, but also implies an increase in the apparent quantum yield, and in the nitrogen allocated to light-harvesting organs ([7], [15], [23], [21]). To our knowledge, there are no studies on A. bijuga assessing the acclimation of the species to different light levels, but it has been observed that light is an important limiting factor for the development of understory vegetation in other conifer plantations ([14]). In this study, a nursery experiment was established with young plants of A. bijuga grown under three different light levels and maintained under the warm conditions of the summer months in Central Chile. The objective of the study was to assess the growth and physiological acclimation of A. bijuga to shade at the early growing stage. The results of this study might provide information for the conservation and ecological restoration of the species.
Materials and methods
Plant material and study design
In winter 2016, A. bijuga cuttings were collected in the field from its natural population in the Coastal Range of the Maule Region in Central Chile (35° 27′ S, 72° 27′ W; elevation 150 m a.s.l.). Cuttings were obtained from healthy plants with no herbivore damage and were 33 cm long, with the presence of ax”illary buds. After collection, cuttings were planted in 140 mL pots containing composted bark of P. radiata and grown for two growing seasons in facilities of the Company “Forestal Mininco” S.A. Then, the plants were transferred to the nursery facilities at the Universidad de Talca, city of Talca, Chile (35° 24′ S, 71° 38′ W; elevation 112 m a.s.l.). Plants were transplanted to 1 L plastic bags filled with local topsoil and grown for six months in a nursery and covered with an 80% black polyethylene mesh (Raschel®, Santiago, Chile). The soil water content was maintained between 26% to 45% and monitored using a ThetaProve® soil moisture sensor (Delta-T Ltd, Cambridge, UK). No fertilizers were added to the growing substrate.
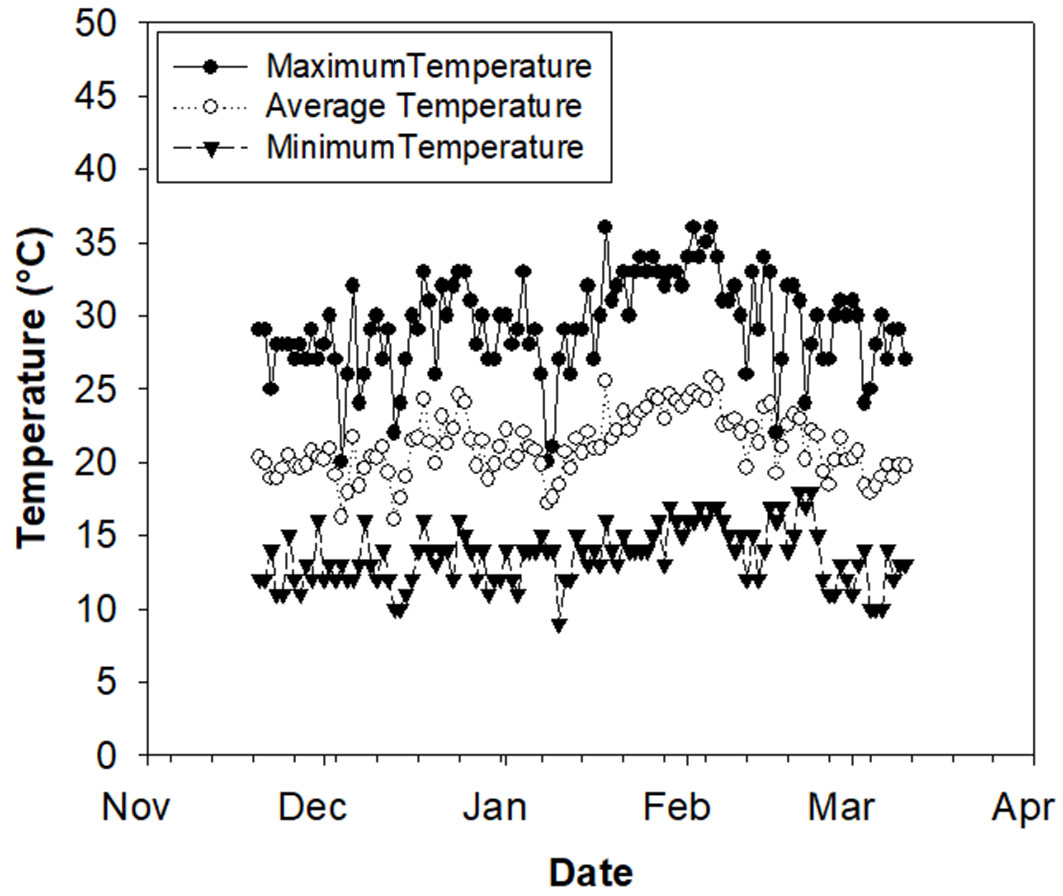
The study was a completely random design with two shading treatments and a control treatment at full sun. Two 1 m3 domes differing in light interception were built for each shading treatment using black polyethylene mesh (Raschel®, Santiago, Chile) and secured with 1.3 cm diameter polyvinyl chloride (PVC). Each treatment had two replicates that were randomly assigned within each dome and six plants were randomly assigned to each replication. Shading treatments were defined as the percentage of photosynthetic active radiation (PAR) intercepted by the domes relative to the maximum PAR recorded at midday during the summer months (about 2100 µmol m-2 s-1), which was determined by using a quantum sensor LI-190® (LICOR Inc., Lincoln, NE, USA). Thus, the sun-exposed treatment was named TRT0, while the shading treatments were named TRT60 and TRT90 (60% and 90% of PAR interception, respectively). Light level at TRT90 represents closed-canopy on mature P. radiata stands. Light treatments were imposed on November 27, 2018, and maintained until March 7, 2019. Fig. 1shows the minimum, average, and maximum temperatures recorded during the study period. Because the material available comprised a pool of genotypes, and there was a high variation in plant height, plants for the experiment were selected based on their growth uniformity and visual healthy condition.
Fig. 1 - Daily maximum, minimum, and mean air temperature for the duration of the experiment.
Growth and physiological measurements
Growth and photosynthetic-light response curves were measured on three dates: Date 1, a baseline measurement a day before assigning the plants to the treatments (November 27, 2018); Date 2, mid-summer (February 1, 2019); and Date 3, at the end of summer (March 7, 2019). Plant height (H) of the dominant shoot was measured using a metric tape, and increments of this variable were determined among measurement dates. Light-response curves were made using a portable gas exchange system LI 6800® (LICOR Inc., Lincoln, NE, USA). Initial chamber conditions were set up at ambient conditions, with a temperature of 20 °C, CO2 concentration of 400 ppm, a relative humidity of 50%, and PAR of 1800 µmol m-2 s-1. These parameters were kept steady during the measurements and controlled by the LI 6800. The light-response curve was obtained in a ten-step reduction process (1800, 900, 600, 300, 200, 150, 100, 50, 10, 0 μmol m-2 s-1). Five plants per treatment were randomly selected at each date. At each plant, two leaflets of a compound leaf from the upper third were carefully placed in the chamber to avoid self-shading and acclimated to the initial conditions for 2 minutes. Measurements were performed between 09:00 and 13:00 hours local time. Because of the small leaf size, additional measurements of leaf size (i.e., lamina’s length and wide) were used to calculate leaf area in the chamber (L) as (eqn. 1):
where l is the lamina length, w is the lamina width and n the number of leaves within the chamber. Similarly, the mean area of the lamina was calculated as LAlamina = L/n. The maximum photosynthetic rate (Amax), dark respiration rate (Rd), light compensation point (Γi), and apparent quantum yield (α) were estimated fitting the non-rectangular hyperbola ([13]).
In the third measurement date, the foliage was collected from plants on each treatment and scanned for leaf area determination using a bed scanner and the software Image J (developed by the National Institute of Health). Then, the foliage was dried at 65 °C to a constant weight to determine the specific leaf area (SLA, leaf area to dry weight ratio). Similarly, on the second and third measurement dates, the live crown percentage was calculated as the portion of the shoot with foliage divided by the total plant height.
Statistical analysis
All the analyses were made in the software SAS® version 9.2 (SAS Institute Inc., Cary, NC). Light-response curves parameters were obtained using non-linear least squares in the PROC NLIN procedure. Traits that were measured only at the end of the study were subjected to a one-way analysis of variance, with the light treatment as the main factor, whereas in the other traits, the model was extended to a repeated measures analysis. We used the unstructured (UN) function to model the variance-covariance matrix of the error terms. All the variables were analyzed using the PROC MIXED procedure. Mean comparisons were made using the Tukey’s adjustment test. Treatment effects were considered significant at an alpha level of 0.05.
Results
Growth performance and leaf traits
Before imposing the light treatments, average plant diameter and height for the dominant branch were 3.7 mm and 45.4 cm, with coefficients of variation (CV) of 16% and 18%, respectively (Fig. 2). Plant survival within the domes varied from 41% to 75%. The results of the analysis of variance showed a significant interaction between the light treatment and measurement date for H (p=0.0037) and live crown percentage (p=0.0016 - Tab. 1). Differences among treatments for both traits were found since Date 2 (Fig. 3A). At the end of the experiment (i.e., Date 3), plant H was 41% higher in TRT90 than TRT0, whereas plants at TRT60 had an intermediate value (Fig. 3A). At Date 3, plants at TRT90 had 97% higher Lcrown than plants in the other treatments (Fig. 3B).
Fig. 2 - View of the A. bijuga plants under the different shading treatments (TRT0: full sun; TRT60: 60% shade; TRT90: 90% shade) during the second measurement.

Tab. 1 - P-values based on the analysis of variance for differences among treatments (TRT) and dates (DATE) in plant height, live crown percentage (Lcrown), specific leaf area (SLA), leaf area per lamina (LAlamina), maximum photosynthetic rate (Amax), dark respiration (Rd), light compensation point (Γi), and apparent quantum yield (α).
| Source of variation | Growth and leaf morphology | Photosynthetic capacity | ||||||
|---|---|---|---|---|---|---|---|---|
| Height | Lcrown | SLA | LAlamina | Amax | Rd | Γi | α | |
| TRT | 0.0210 | 0.0073 | 0.0011 | 0.7069 | 0.0414 | 0.1845 | 0.0879 | 0.3821 |
| DATE | <0.0001 | <0.0001 | - | - | 0.0272 | 0.0105 | 0.0124 | 0.3609 |
| TRT × DATE | 0.0037 | 0.0016 | - | - | 0.0493 | 0.0056 | 0.0451 | 0.3490 |
Fig. 3 - Mean (± standard error) values per treatment (TRT0, TRT60, TRT90) and date for (A) height and (B) live crown percentage in young A. bijuga plants. (*): denotes significant differences among treatments. Different lowercase letters denote significant differences among treatments, according to the Tukey’s adjustment test. The inset graph shows the means for specific leaf area (SLA) per treatment at the end of the study period.
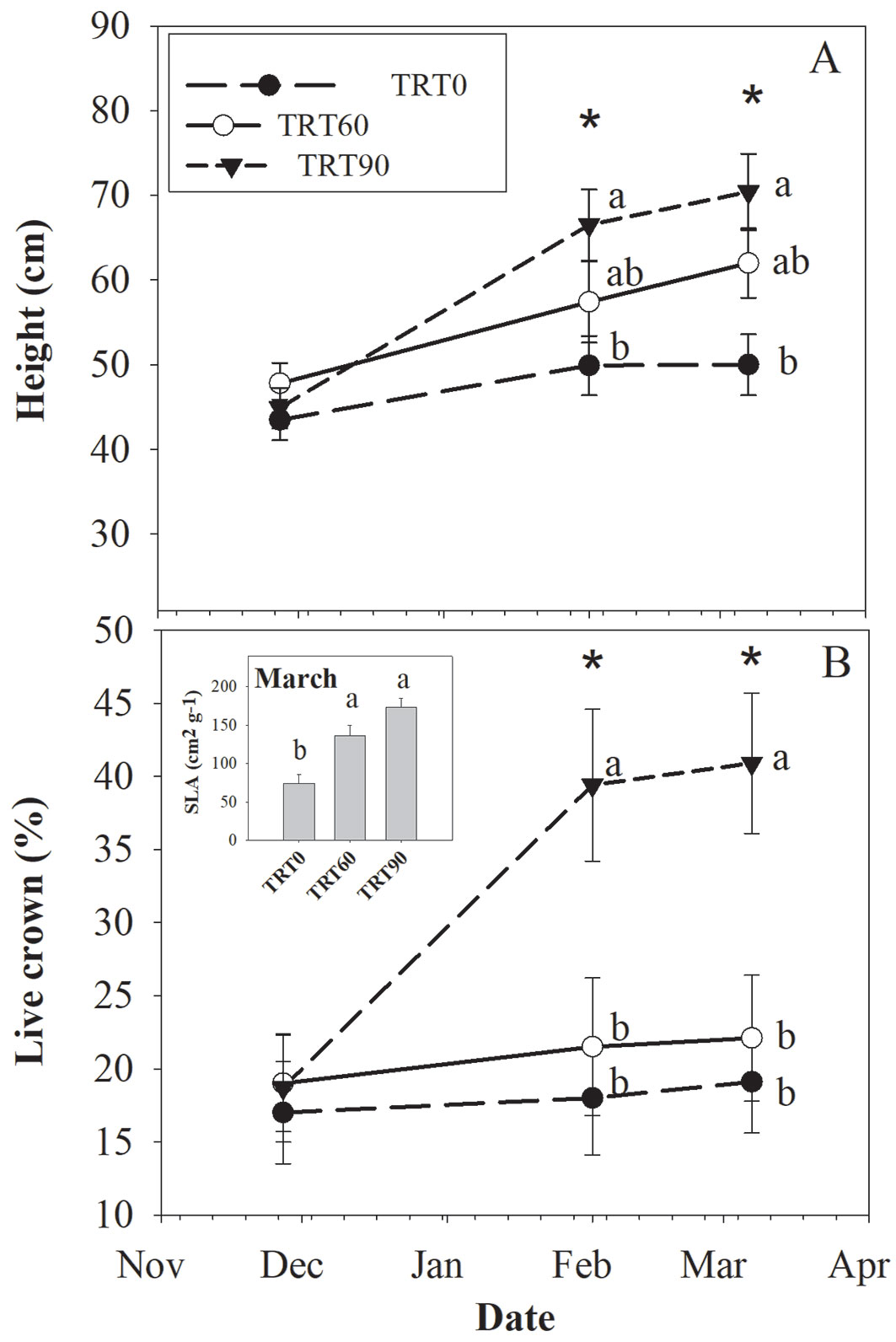
There was no effect of the treatments on LAlamina (p=0.7069), but increasing the shading level increased SLA (p=0.0011 - Tab. 1). SLA did not differ between TRT60 and TRT90, and on average, plants at these treatments had a SLA 108% higher than plants at TRT0 (Fig. 3B).
Photosynthetic light response curves parameters
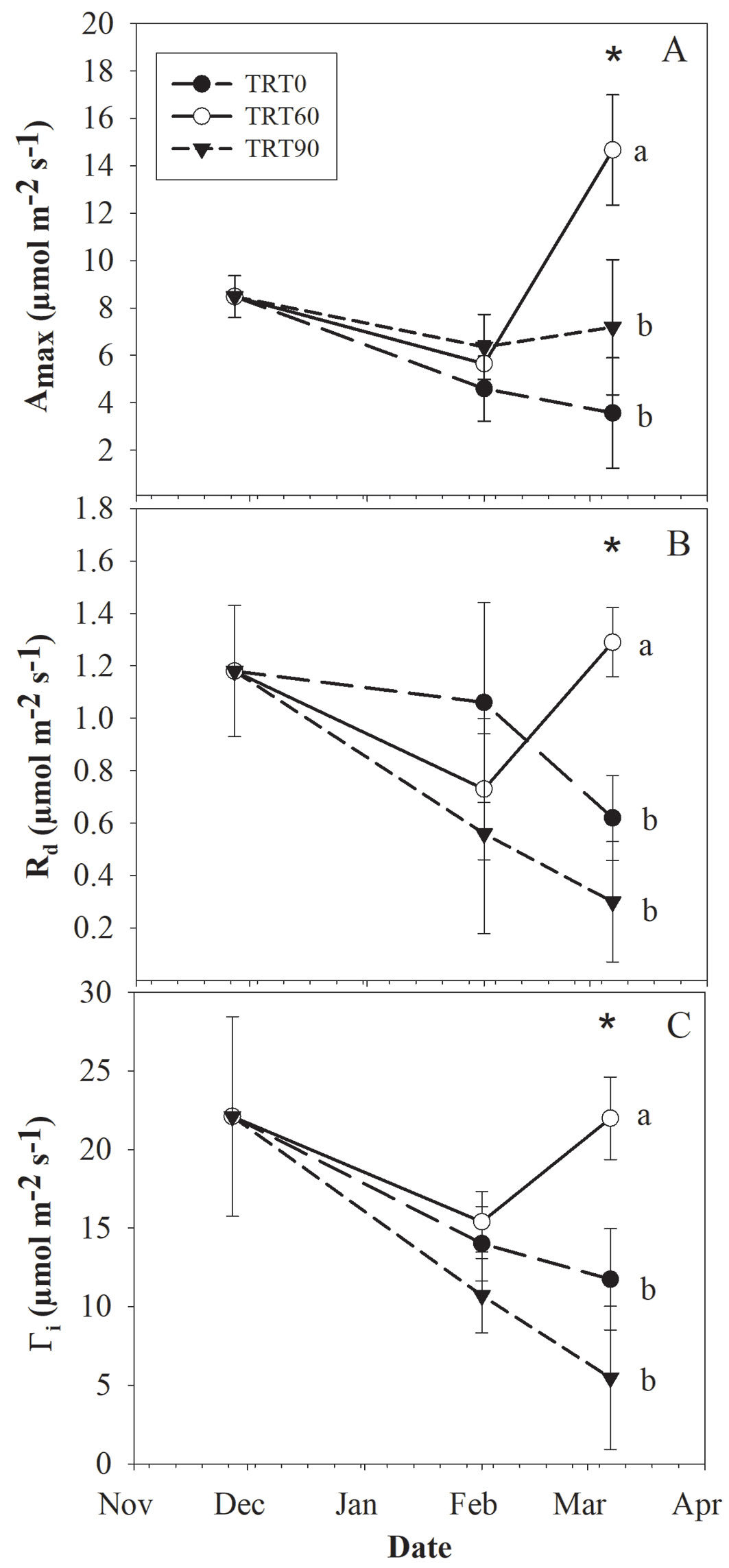
With the exception of α, there was a significant effect of the interaction between the light treatment and measurement date for all the variables derived from the photosynthetic light response curves (Tab. 1). The differences among treatments were found only at the end of the experiment, i.e., Date 3 (p<0.05 - Fig. 4). Amax, Rd, and Γi did not differ between TRT0 and TRT90, but these values were significantly lower than those on TRT60. Overall, these photosynthetic parameters tended to decrease with time for TRT0 and TRT90 (Fig. 4) and tended to decline in TRT60 toward Date 2 and then increased toward Date 3. Mean α for the first, second, and third measurement dates were 0.051, 0.070, and 0.058 µmol CO2 µmol photon-1, respectively. Overall, stomatal conductance (gs) ranged from 0.007 to 0.28 mmol m-2 s-1, whereas vapor pressure deficit (VPD) ranged from 1.29 to 1.79 MPa.
Fig. 4 - Mean (± standard error) values per treatment (TRT0, TRT60, TRT90) and (A) date for maximum photosynthetic rate (Amax), (B) dark respiration (Rd), and (C) light compensation point (Γi) in young A. bijuga plants. (*): denotes significant differences among treatments. Different lowercase letters denote significant differences among treatments according to the Tukey’s adjustment test.
Discussion
Several factors might be involved in the population decline of A. bijuga. There is thus a need to understand the ecological behavior of the species in order to develop strategies for its conservation. An important factor driving the population recruitment of the species is the ecological pressure that pine plantations might exert over the highly fragmented habitats where this species develops. Some studies have shown the negative effects of pine invasion on the richness of other native species ([4]), which is related to the competition for light and soil resources. We started under the assumption that, in its natural growing environment, the observed loss of vigor and growth of A. bijuga in the understory might be related to its shade-intolerant behavior, a fact that is not well understood in this species. In this study, we examined the growth and leaf-level physiological acclimation of young A. bijuga plants to different light treatments. Overall, our results indicate that the species had high phenotypic plasticity to light intensity. However, its higher physiological performance was in the partial shade treatment (TRT60) and not in the full-sun exposed treatment (TRT0).
Small compound leaves, like those of A. bijuga, might be considered as an adaptive characteristic of species growing under high light or dry environments ([29]). Small leaves are typically found in shade-intolerant species, while large leaves are found in shade-tolerant species ([26], [30]). Under high light, small leaves may improve the coupling with the atmosphere, reducing the boundary-layer resistance and consequently leaf temperature and water loss ([22], [26]). However, beyond their classification as shade-tolerant and intolerant species, each species may exhibit a different degree of acclimation to light ([16]). Some studies show that shade-intolerant species may have higher acclimation plasticity to different light levels than shade-intolerant species ([24]), which may be the case of A. bijuga. The latter author also mentioned that the ability for acclimation of SLA and leaf mass per area (LMA, the inverse of SLA) to shade might be a good predictor of the relative growth and survival rate. This agreed with our results because SLA was highly plastic and exhibited an increase over 2-fold in the shaded treatments relative to the control full sun-exposed. A high SLA also implies a higher number of chloroplasts and enzymes on leaves and an improved photosynthesis capacity ([3]). The fact that treatments did not differ in LAlamina suggests that differences in SLA (p=0.7069) are mainly due to the differences in dry mass, which implies a decreased leaf thickness, with lower investment in leaf construction and protective tissues ([24]).
We are aware of the low replication of the study, which likely influenced the lack of significance of some traits such as LAlamina (Tab. 1). Nevertheless, trends were clear for all the traits, and values for leaf morphological traits and height were lower at TRT0, which is consistent with the results found in other studies ([16]). However, Liu & Su ([16]) mentioned that a lower investment in shoot development might favor leaf production and its competitiveness capability, which disagree with our study. At the full sun, A. bijuga reduced growth and live crown percentage (i.e., leaf area). The lower Lcrown at TRT0 might be associated to faster senescence due to transient plant water stress ([18]). Because plants in the nursery were acclimated to shade before treatment imposition, the lower Lcrown at TRT0 was probably induced by the inability of older leaves to counteract photoinhibition ([18]). Otherwise, the new living foliage was developed and acclimated to the high light condition from the beginning. We argue that the high temperature and light conditions during the study period might have affected A. bijuga growth at TRT0. During the experimental period, maximum temperatures were over 30 °C most of the days (Fig. 1). Thus, although the water was kept over 26%, there likely was a thermal stress on those plants. The high thermal conductivity and the low volume of the plastic bags, likely affected the substrate heat dissipation and, consequently, the root functions and shoot growth.
The study of light response curves is used to assess the shade tolerance of the species ([5]) since shade tolerance is associated with several functional traits and is also influenced by other abiotic factors ([30]). In our study, A. bijuga plants exhibited high photosynthetic rates up to 15 µmol m-2 s-1 at the intermediate light level (i.e., TRT60 - Fig. 3). To our knowledge, little is known about the ecophysiology of Adesmia species. However, the photosynthetic rates found in this study were two-fold higher to the ones reported for other Adesmia species growing at high altitudes in the north-central Chilean Andes ([25]). Overall, sun acclimated leaves have high assimilation rates (Amax), dark respiration (Rd), and light compensation point (Γi), but lower quantum efficiency relative to shade adapted leaves ([23], [30], [17]). These findings partially agreed with our results as a greater physiological performance was obtained at TRT60, in which Amax, Rd and Γi were higher than in the other treatments, while there were no differences between the extreme light treatments TRT0 and TRT90. We argued that regarding the harsh environmental and experimental conditions (i.e., light, temperature, rooting volume) described in this study, the low values of the photosynthetic parameters obtained in TRT0 might be explained by damages to the plants photosynthetic machinery. Although it was not measured, we observed a higher yellowing in plants exposed to TRT0 than the other treatments, suggesting a denaturation of leaf pigments. However, this photochemical damage was not expressed on differences among treatments in α, a parameter that is altered when the excess of light leads to photoinhibition. On the other hand, the low values of the physiological parameters in TRT90 compared with TRT60 suggests that the photosynthetic capacity of A. bijuga is considerably compromised at the low light level. Therefore, more research is needed in this respect, especially in field trials. We speculate that the species has high plasticity to light levels, but this might be a specific feature of young plants. Torres et al. ([28]) mentioned that the acclimation capacity of a species to different light levels at early ontogenic stages is associated with its ability to prosper in the forest understory. Otherwise, as plants develop in the understory, they might also be limited for other resources than light, such as water acquisition and temperature. Pine species are good competitors for resources, and they decrease the composition and abundance of understory species as crown develops ([4]). Thus, the low vigor of A. bijuga growing under pine canopies might be associated with complex interactions among light, temperature, and water availability ([28]), which needs further investigations. Overall, species of the genus Adesmia have great potential for forage and for medicinal purposes ([6]), and are well adapted to the semi-arid condition in Mediterranean climates due to its high stomatal control ([25]). However, species such A. bijuga are under high anthropogenic pressure, thus basic information like the one provided by this study is needed for their restoration.
Conclusions
Our study showed that A. bijuga exhibited acclimation plasticity to different light levels. At the early stages of development, a semi-shade environment favored a better physiological performance of the species, but growth and leaf morphological traits were benefited in an environment with higher shading. Thus, the results of this study contradict our assumption that light availability is the main factor influencing the vigor loss of the species when this develops beneath P. radiata plantations. However, this contention needs further research to explore the potential interaction between light and other abiotic factors affecting the species development.
Conflict of interest
The authors declare that there is no conflict of interest.
Acknowledgments
We thank the Poplar Technology Center (PTC) of the University of Talca (Chile) for providing the LI-6800.
References
Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Núcleo Científico Multidisciplinario. Universidad de Talca. 2 Norte 685, Talca. P.O. Box 747 (Chile)
Instituto de Bosques y Sociedad, Facultad de Ciencias Forestales y Recursos Naturales, Universidad Austral de Chile, Campus Isla Teja, Valdivia (Chile)
Facultad de Ciencias Agrarias y Forestales, Universidad Católica del Maule, Avenida San Miguel, 3605 Talca (Chile)
Corresponding author
Paper Info
Citation
Yáñez MA, Gómez P, Gajardo J, Espinoza S (2021). Growth and physiological acclimation to shade in young plants of Adesmia bijuga Phil., a critically endangered species in central Chile. iForest 14: 307-312. - doi: 10.3832/ifor3640-014
Academic Editor
Gianfranco Minotta
Paper history
Received: Sep 01, 2020
Accepted: Apr 19, 2021
First online: Jul 01, 2021
Publication Date: Aug 31, 2021
Publication Time: 2.43 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2021
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 34897
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 29186
Abstract Page Views: 2827
PDF Downloads: 2289
Citation/Reference Downloads: 1
XML Downloads: 594
Web Metrics
Days since publication: 1822
Overall contacts: 34897
Avg. contacts per week: 134.07
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2021): 3
Average cites per year: 0.60
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Conservation of Betula oycoviensis, an endangered rare taxon, using vegetative propagation methods
vol. 13, pp. 107-113 (online: 23 March 2020)
Research Articles
Growth, morphology, and biomass allocation of recently planted seedlings of seven European tree species along a light gradient
vol. 13, pp. 261-269 (online: 03 July 2020)
Short Communications
Effect of four levels of shade on survival, morphology and chlorophyll fluorescence of Nothofagus alessandrii container-grown seedlings
vol. 8, pp. 638-641 (online: 08 January 2015)
Research Articles
Light acclimation of leaf gas exchange in two Tunisian cork oak populations from contrasting environmental conditions
vol. 8, pp. 700-706 (online: 08 January 2015)
Research Articles
The effect of calcium on the growth of native species in a tropical forest hotspot
vol. 11, pp. 221-226 (online: 01 March 2018)
Research Articles
Using field and nursery treatments to establish Quercus suber seedlings in Mediterranean degraded shrubland
vol. 13, pp. 114-123 (online: 26 March 2020)
Research Articles
Linking nursery nutritional status and water availability post-planting under intense summer drought: the case of a South American Mediterranean tree species
vol. 9, pp. 758-765 (online: 03 June 2016)
Research Articles
Use of alternative containers for promoting deep rooting of native forest species used for dryland restoration: the case of Acacia caven
vol. 10, pp. 776-782 (online: 02 September 2017)
Technical Reports
Effects of different mechanical treatments on Quercus variabilis, Q. wutaishanica and Q. robur acorn germination
vol. 8, pp. 728-734 (online: 05 May 2015)
Technical Reports
De novo adventitious root formations in mini-cuttings of Azadirachta indica in response to different rooting media and auxin treatments
vol. 8, pp. 558-564 (online: 09 December 2014)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







