
Tropical seedling performance under drought: a functional trait approach for species selection in restoration
iForest - Biogeosciences and Forestry, Volume 19, Issue 1, Pages 9-17 (2026)
doi: https://doi.org/10.3832/ifor4713-018
Published: Jan 10, 2026 - Copyright © 2026 SISEF
Research Articles
Abstract
Tropical rainforests experience seasonal variations in water availability, with dry periods resulting in severe soil water scarcity that critically affects seedling performance. A functional trait-based approach, focusing on morphological characteristics, provides a practical means of identifying drought-tolerant tropical woody species for restoration initiatives. This study aimed to evaluate the effects of drought stress on the early establishment of seven tropical woody seedling species and to identify the traits that contribute to seedling performance under these conditions. A greenhouse experiment was conducted to assess seedling responses to varying drought levels, measuring seedling performance (survival rate, relative growth rate, and wilting state) along with traits related to resource acquisition and allocation. The 12-week experiment revealed that drought significantly reduced soil moisture and seedling performance, with species-specific responses. Principal component analysis reveals a shift in trait importance under drought, suggesting altered resource allocation strategies. The root-to-shoot ratio and leaf mass per area emerged as critical indicators of seedling survival during drought, with larger seed and higher leaf dry matter content initially promoting growth but diminishing in effectiveness under severe drought conditions. Larger seeds exhibited greater drought tolerance, as evidenced by lower wilting rates. Although a functional trait-based approach offers valuable information, our results indicate that it may not be fully predictive of seedling performance under all drought scenarios. These findings underscore the importance of selecting species based on their sensitivity to drought stress for specific restoration strategies.
Keywords
Direct Seeding, Drought Stress, Forest Restoration, Functional Traits, Seedling Survival, Tropical Forest
Introduction
Tropical rainforests are characterized by distinct rainy and dry seasons, resulting in considerable fluctuations in water availability ([45]). During dry periods, soil water potential can drop to -2.3 MPa, resulting in severe water scarcity, particularly in upper soil horizons ([5]). Due to their underdeveloped root systems, seedlings are especially susceptible to drought during this phase ([29]). The marked decline in soil moisture intensifies desiccation and other related stresses, hindering seedling survival and growth ([13], [18]). Given the critical role of seedlings in forest regeneration, understanding drought tolerance is essential for successful and resilient forest restoration efforts ([24]).
The high biodiversity of tropical rainforests makes traditional, species-specific drought-tolerance assessments difficult, as it requires considering a wide variety of species with potentially diverse responses to drought. A functional trait-based approach offers a more efficient alternative by focusing on measurable traits that can be used across species to assess drought tolerance ([16]). These traits can rapidly predict a species’ drought tolerance without extensive testing ([16]). Although several studies have explored the effects of drought on tropical species ([13], [36], [14], [1], [7]), there is a notable lack of studies linking functional traits to seedling performance under drought conditions. This knowledge is crucial for developing effective species-selection criteria for targeted restoration and enrichment-planting initiatives. Despite its advantages, the trait-based framework has limitations, including difficulties in linking morphological traits to specific functions, challenges in quantifying physiological traits, and strong interdependencies among traits ([16]). Furthermore, inconsistencies between trait-based and species-based models add complexity to predictive analyses ([28]).
Plants have evolved various adaptations to cope with drought, from structural modifications to physiological processes ([12], [29]). Seedling establishment under drought depends on germination, a water-sensitive stage influenced by evolutionary history, trait variation, and plastic responses to environmental conditions ([25]). Seedling performance under drought is shaped by resource acquisition traits (e.g., specific root length, leaf area ratio, root length), which enhance water uptake, and resource allocation traits (e.g., root-to-shoot ratio, leaf mass per area, seed mass, and dry matter content), which regulate biomass investment and structural resilience ([36], [29]). Understanding these trait interactions is essential for predicting seedling success and species’ adaptive strategies under water-limited conditions.
Trait trade-offs under drought stress are crucial in shaping plant survival and growth strategies ([51]). Drought often requires balancing resource allocation between growth and survival, leading to trade-offs in physiological and morphological traits ([33]). For example, plants may prioritize root development over aboveground biomass to enhance water uptake ([37]), or invest in roots at the expense of leaf area, reducing photosynthetic capacity but improving water acquisition ([29]). Additionally, optimizing water-use efficiency often results in slower growth rates ([33]). These trade-offs, such as prioritizing root growth for water uptake or investing in water-use efficiency at the expense of slower growth, underscore plants’ complex adaptive strategies for surviving water-limited conditions.
Submontane forests (1000-1500 m a.s.l.) serve as transitional ecosystems between lowland rainforests and montane forests. In Indonesia, these forests are characterized by a declining dipterocarp abundance and an increasing dominance of Fagaceae, Lauraceae, and Myrtaceae, driven by climatic and latitudinal variation ([43]). Unlike seasonal montane forests of Africa and South America, Indonesian submontane forests have a year-round wet climate that supports tropical taxa at higher elevations ([2], [9], [8]). However, these ecosystems face increasing threats, including habitat destruction ([26]) and pronounced fluctuations in water availability ([7]), making them highly susceptible to drought ([48]), particularly for seedlings with underdeveloped root systems.
To address these challenges, we conducted a controlled drought stress experiment on tropical woody seedling species to evaluate functional traits associated with resource acquisition and allocation. By simulating the first dry season - a critical survival bottleneck for seedlings ([49]) - this study examines seedling performance, specifically survival rate, relative growth rate, and wilting state, under varying drought conditions.
The aims of this study are to (i) assess the seedling performance (i.e., survival rate, relative growth rate, and wilting state) of seven tropical woody species under varying levels of drought stress; (ii) explore the effects of drought stress on functional traits and to assess how these traits differ among species in their adaptive responses to water limitation; and (iii) identify critical functional traits that contribute to drought tolerance in seedlings. We examined the following hypotheses: (1) seedlings of seven tropical woody species exhibit different responses to drought stress; (2) under drought conditions, resource allocation traits show adaptive patterns, with increased biomass investment in roots to enhance water uptake; and (3) resource allocation traits can predict seedling performance under drought stress, as they determine the efficiency of resource acquisition and stress tolerance. This research addresses the critical challenge of selecting drought-tolerant species for targeted restoration strategies and provides recommendations for species selection based on drought resilience.
Materials and methods
Study system
We conducted the experiment from April to December 2023 in a controlled greenhouse at the Cibodas Botanical Garden, Indonesia (06° 44′ 00.4″ S, 107° 00′ 18.9″ E; 1350 meters a.s.l.), which is included in the Cibodas Biosphere Reserve. The average greenhouse conditions were a temperature of 18.0 ± 1.6 °C, relative humidity of 99.1 ± 0.6%, and light intensity of 1.194 ± 2552 lux.
Seven submontane tropical woody species native to West Java, Indonesia, were chosen for this study: Alangium rotundifolium (Hassk.) Bloemb., Engelhardia serrata Blume, Ficus ribes Reinw. ex Blume, Heptapleurum lucescens (Blume) Lowry & G.M.Plunkett, Leea indica (Burm.f.) Merr., Neolitsea cassiifolia (Blume) Merr., and Pyrenaria serrata Blume (Tab. 1). These species represent diverse families and growth forms and encompass a range of plant functional types. Their nomenclature was verified using the International Plant Names Index (IPNI - ⇒ https://www.ipni.org/) and Plants of the World Online (POWO - ⇒ https://powo.science.kew.org/).
Tab. 1 - List of woody species included in the study.
| Species | Family | Growth form |
|---|---|---|
| Alangium rotundifolium (Hassk.) Bloemb. | Cornaceae | Tree |
| Engelhardia serrata Blume | Juglandaceae | Tree |
| Ficus ribes Reinw. ex Blume | Moraceae | Tree |
| Heptapleurum lucescens (Blume) Lowry & G.M.Plunkett | Araliaceae | Scrambling tree |
| Leea indica (Burm.f.) Merr. | Vitaceae | Shrub or tree |
| Neolitsea cassiifolia (Blume) Merr. | Lauraceae | Tree |
| Pyrenaria serrata Blume | Theaceae | Tree |
Seed collection, germination, and seedling growth
Seeds were collected from natural forests within the Cibodas Natural Reserve, West Java, following the methodology described by Schmidt ([40]). The collection was conducted from the tree canopies one month before the experiment. Prior to germination, seeds were extracted from fleshy fruits, washed, air-dried, and stored at room temperature.
Germination occurred in containers filled with organic soil from April to May 2023. The time required for seedling emergence and the appearance of the first leaf were tracked for each species. Following successful germination, seedlings entered the growing phase (June-August 2023). Each seedling was transplanted into pots containing a 1:1:1 mixture of organic soil, cocopeat, and rice husk ash. Seedlings were regularly watered to keep the soil moisture at field capacity. Soil nutrients were maintained to prevent deficiencies by applying 1 g of slow-release fertilizer per pot (NPK 17:11:10, Osmocote®, Geldermalsen, NL).
Determination and maintenance of soil field capacity
Soil field capacity (FC) was determined using the gravimetric method ([20]). Soil samples were oven-dried at 105 °C for 24 hours to obtain their dry weight (Wdry). The dried soil was then placed in pots, and water was gradually added until saturation was reached. Excess water was allowed to drain freely for 48 hours to achieve equilibrium moisture content. The final soil weight after drainage was recorded as WFC, and field capacity was calculated using the following formula (eqn. 1):
Based on preliminary measurements, 100% FC corresponded to 351 mL of water per pot. To simulate drought stress, soil moisture was maintained at four FC levels: 100% FC (351 mL), 80% FC (281 mL), 60% FC (211 mL), and 30% FC (105 mL). Daily pot weight measurements were conducted to monitor water loss due to evaporation and plant uptake, and water was replenished accordingly to maintain target FC levels. For example, if a pot at 100% FC exhibited a 50 g (mL) weight reduction, an equivalent volume of water was added to restore the initial FC.
Experimental design and trait measurements
The experiment, conducted from September to November 2023, evaluated drought stress tolerance in woody tropical species. Soil moisture was maintained at four FC levels: well-watered (100% FC), mild drought (80% FC), moderate drought (60% FC), and severe drought (30% FC), representing a gradient of drought stress relevant to plant physiology and adaptation ([47]). Each species was subjected to all four watering treatments, with sixteen replicates per treatment, and plants were randomly arranged across three greenhouse benches. Soil moisture was monitored daily using pot weight measurements, with water added as needed to maintain the assigned FC levels.
Nine key functional traits related to resource acquisition and allocation strategies were measured throughout the experiment (Tab. 2). The selected traits, i.e., specific root length (SRL), root length, root-to-shoot ratio (RSR), leaf area ratio (LAR), leaf mass per area (LMA), seed mass, leaf dry matter content (LDMC), stem dry matter content (SDMC), and root dry matter content (RDMC), were assessed following Pérez-Harguindeguy et al. ([32]).
Tab. 2 - Plant traits measured in this study.
| Traits | Variables | Abbr. | Units | Definition |
|---|---|---|---|---|
| Resource acquisition traits |
Specific root length | SRL | cm g-1 | Root length per root dry mass |
| Leaf area ratio | LAR | cm g-1 | Total leaf area relative to plant dry mass | |
| Root length | - | cm | Final root length | |
| Resource allocation traits |
Leaf mass per area | LMA | g m-2 | Ratio of leaf dry mass to fresh leaf area |
| Root-to-shoot ratio | RSR | - | Ratio of root mass (g) to shoot mass (g) | |
| Leaf dry matter content | LDMC | g g-1 | Leaf dry mass per fresh mass | |
| Stem dry matter content | SDMC | g g-1 | Stem dry mass per fresh mass | |
| Root dry matter content | RDMC | g g-1 | Root dry mass per fresh mass | |
| Seed mass | - | mg | Dry mass per seed | |
| Seedling performances |
Seedling survival | - | % | (N2/N1) × 100 |
| Relative growth rate | RGR | mg g-1 day-1 | ln(X2 - X1)/(t2 - t1) | |
| Wilting states | - | - | Categorized into six stages ranging from normal to dead (Tab. S1) |
These functional traits are essential for understanding plant growth strategies, resource allocation, and ecological adaptations, particularly under drought conditions. The selection of root-shoot allocation traits aligns with functional equilibrium theory, which explains how plants allocate biomass between roots and shoots in response to resource availability ([1]). SRL reflects plant efficiency in soil exploration and resource uptake, whereas root length determines soil foraging capacity ([23]). LAR and LMA provide insights into photosynthetic capacity and leaf economic strategies, with LMA often linked to leaf toughness and stress tolerance ([50], [35]). The RSR represents biomass partitioning between above- and below-ground structures and influences drought resilience ([23]). Seed mass affects seedling survival and establishment success ([30]), while LDMC, SDMC, and RDMC are indicators of tissue robustness, resource conservation, and drought resistance ([29]).
In September 2023, before the drought experiment began, sixteen additional seedlings per species were randomly selected for initial measurements (outside the drought treatment groups). Plants were removed from pots, roots rinsed to remove soil, and seedlings divided into roots, stems, and leaves for fresh-weight measurement on an analytical balance. Digital images were captured for analysis using ImageJ software ([41]). The samples were oven-dried at 80 °C for 48 hours to obtain their dry mass, facilitating initial calculations of relative growth rate and root length.
Each species comprised 64 seedlings subjected to four drought-stress treatments for 12 weeks. Seedling survival was monitored daily, and wilting stages were assessed weekly (Tab. S1 in Supplementary material) according to Engelbrecht & Kursar ([12]). At the end of the experiment, all remaining seedlings were collected and photographed, and measurements of root, stem, and leaf characteristics were taken to quantify functional traits and assess seedling performance.
Data analyses
Data were analyzed using R ver. 4.4.0 ([38]). We compiled a matrix of seedling performance (i.e., survival rate, relative growth rate, and wilting state) and functional traits (i.e., SRL, LAR, root length, RSR, LMA, seed mass, LDMC, SDMC, and RDMC) for seven tropical woody species. Because the data on soil volumetric water content and seedling performance did not meet the assumption of normality, nonparametric tests were employed. To assess overall differences in soil volumetric water content among treatment groups, we performed a Kruskal-Wallis test (α = 0.05). Significant differences among specific treatments were identified using Wilcoxon rank-sum tests with a Bonferroni correction. Similarly, Kruskal-Wallis tests were utilized to analyze seedling survival, growth rate, and wilting point under drought conditions, with treatment groups serving as the explanatory variable.
We used principal component analysis (PCA) to assess relationships among traits across seven species (Tab. S2), employing the “FactoMineR” package ([19]). Spearman correlations were computed to aid the interpretation of trait associations. Given the strong influence of water availability on plant traits, we further examined its role in shaping trait trade-offs. PCA with Varimax rotation was applied to identify key axes summarizing trait variation and relationships. To analyze trait responses to drought, we fitted linear mixed-effects models using the “lme” function in the “lme4” package ([4]), with treatment, species, and their interaction as fixed effects, and greenhouse bench as a random factor.
Seedling responses to drought were analyzed using generalized linear mixed models (GLMMs). Model optimization was performed using the “optimx” package ([31]), and models were subsequently fitted using the “glmer” function in the “lme4” package ([4]). The normality of response variables and predictor variables was assessed using the Shapiro-Wilk test, while Levene’s test was used to evaluate homoscedasticity, both implemented via the “rstatix” package ([22]).
The models were used to examine the relationship between response variables, seedling survival, RGR, and wilting states, and predictor variables, which included resource acquisition and allocation traits, while incorporating a random factor by species. Multicollinearity among predictor variables was assessed using Spearman’s correlation coefficient, with a threshold of ρ > 0.7 indicating high collinearity ([11]). As LMA and SDMC were highly correlated across all drought treatments, SDMC was excluded from the final model to avoid redundancy.
GLMMs with binomial (link = logit) distributions were applied to analyze trait-survival associations, while Gaussian (link = identity) and Poisson (link = log) distributions were used for RGR and wilting states, respectively. Key functional-trait predictors of seedling performance were identified using backward multiple regression ([52]).
Results
Drought treatment effects on seedling performance
During the 12-week drought experiment, soil moisture significantly decreased from 17.5 ± 1.2% volumetric water content (VWC) in the control to 7.7 ± 1.3% VWC in the severe drought treatment. The Kruskal-Wallis test (df = 3) showed a highly significant effect of drought stress on soil water content (p < 2.2e-16 - Fig. S1 in Supplementary material), indicating a gradient of decreasing moisture with increasing drought stress. Soil water content across the four treatments was consistently maintained throughout the experiment (Fig. S2).
Seedling survival declined with increasing drought stress (control 95%, mild 93%, moderate 88%, and severe 82% - Fig. 1). Severe drought significantly reduced survival (df = 3, p < 0.05). No seedling mortality occurred in H. lucescens, L. indica, A. rotundifolium, and P. serrata under drought stress (Fig. S3 in Supplementary material). In contrast, E. serrata, F. ribes, and N. cassiifolia exhibited low survival rates (<80%) under drought conditions. RGR also declined with increasing drought stress. Significantly lower values were shown by moderate and severe drought conditions compared to the control (p < 0.05 - Fig. 1). RGR under drought varied between species, ranging from 26.33 ± 13.25 to 63.77 ± 13.34 mg g-1 day-1 for E. serrata and A. rotundifolium, respectively.
Fig. 1 - Drought treatment effects on seedling performance (A) seedling survival, (B) relative growth rate, and (C) wilting states. P-value is based on comparison with control using the Kruskal-Wallis test followed by the Wilcoxon post-hoc rank sum test. (*): p < 0.05; (***): p < 0.001; (****): p < 0.0001.
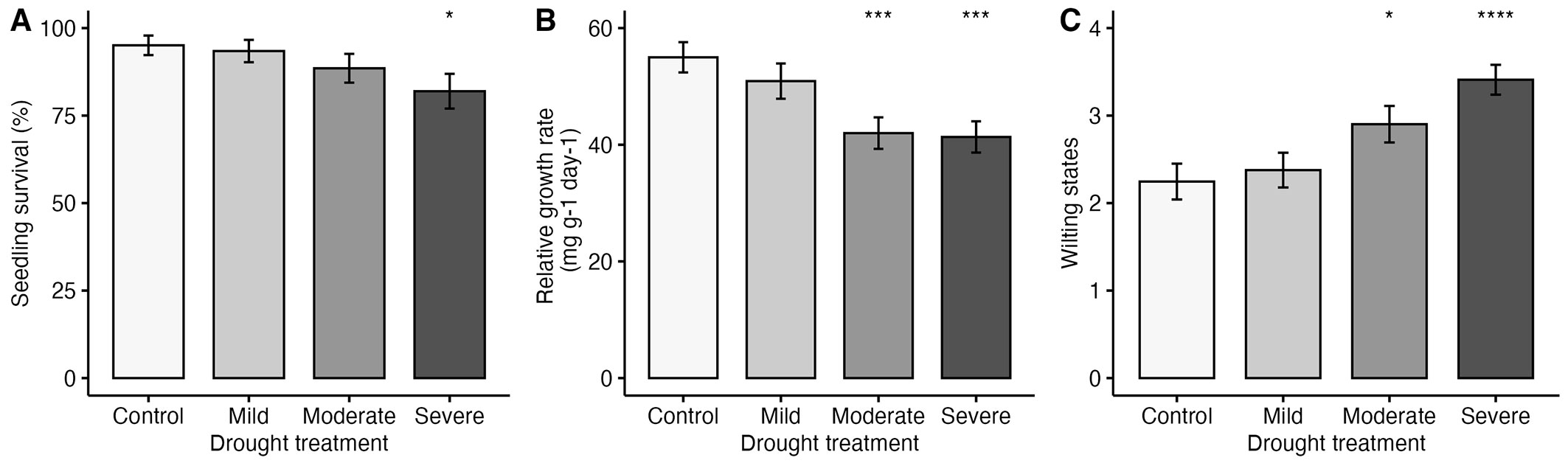
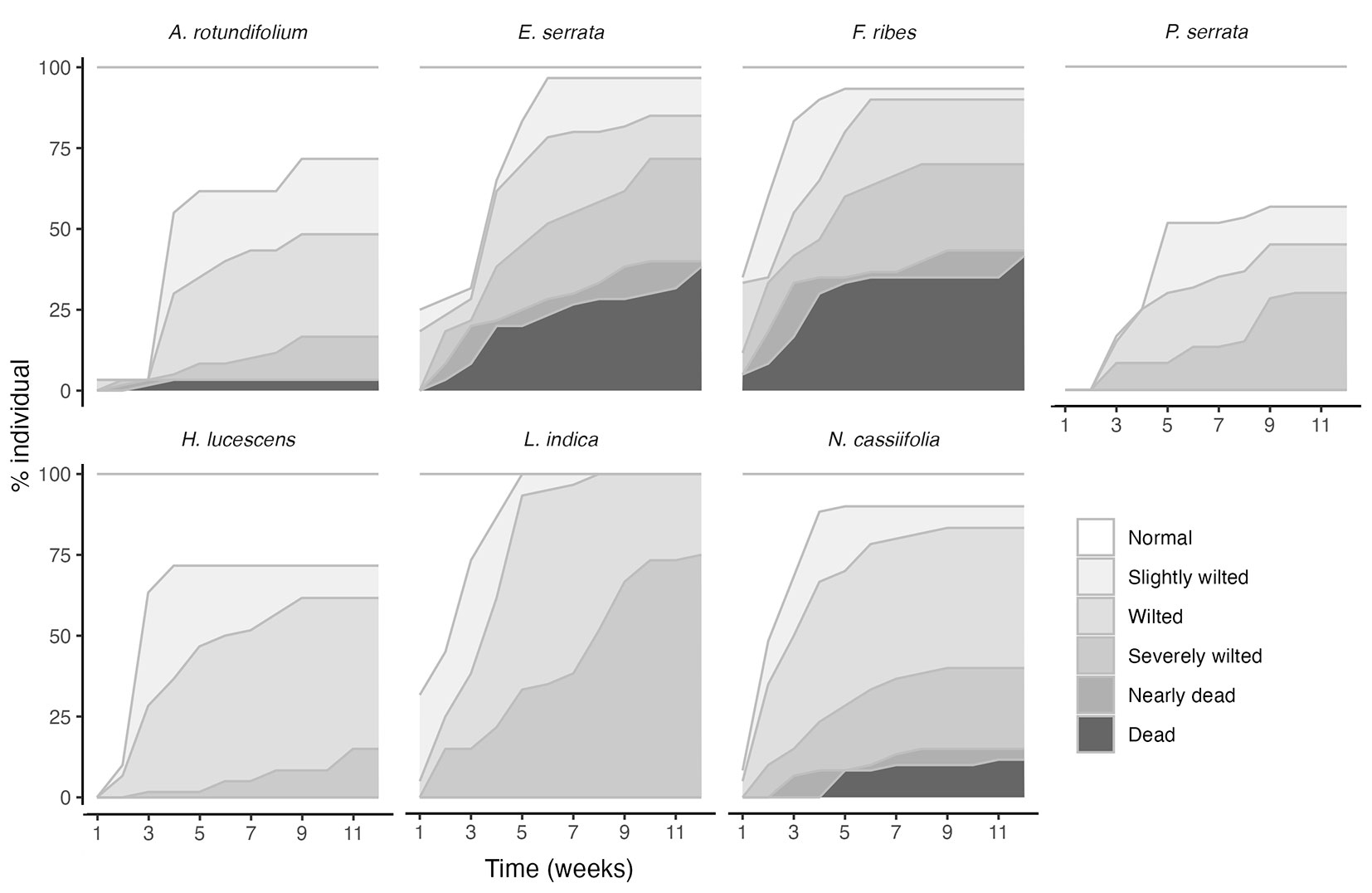
Wilting was significantly more pronounced under moderate and severe drought than under control conditions (Fig. 1, Fig. S4). In most species, drought treatments caused wilting, with varying effects among species (Fig. 2). E. serrata, F. ribes, and L. indica wilted early, with some individuals dying early except for L. indica. Conversely, A. rotundifolium, H. lucescens, and P. serrata showed minimal visible drought stress and low mortality. There was a continuum of responses, with species such as E. serrata, F. ribes, and N. cassiifolia exhibiting varying degrees of wilting and mortality.
Fig. 2 - Wilting states and mortality among species under drought stress conditions during 12 weeks of study (for the description of the wilting states, see Tab. S1 in Supplementary material).
Variation in traits
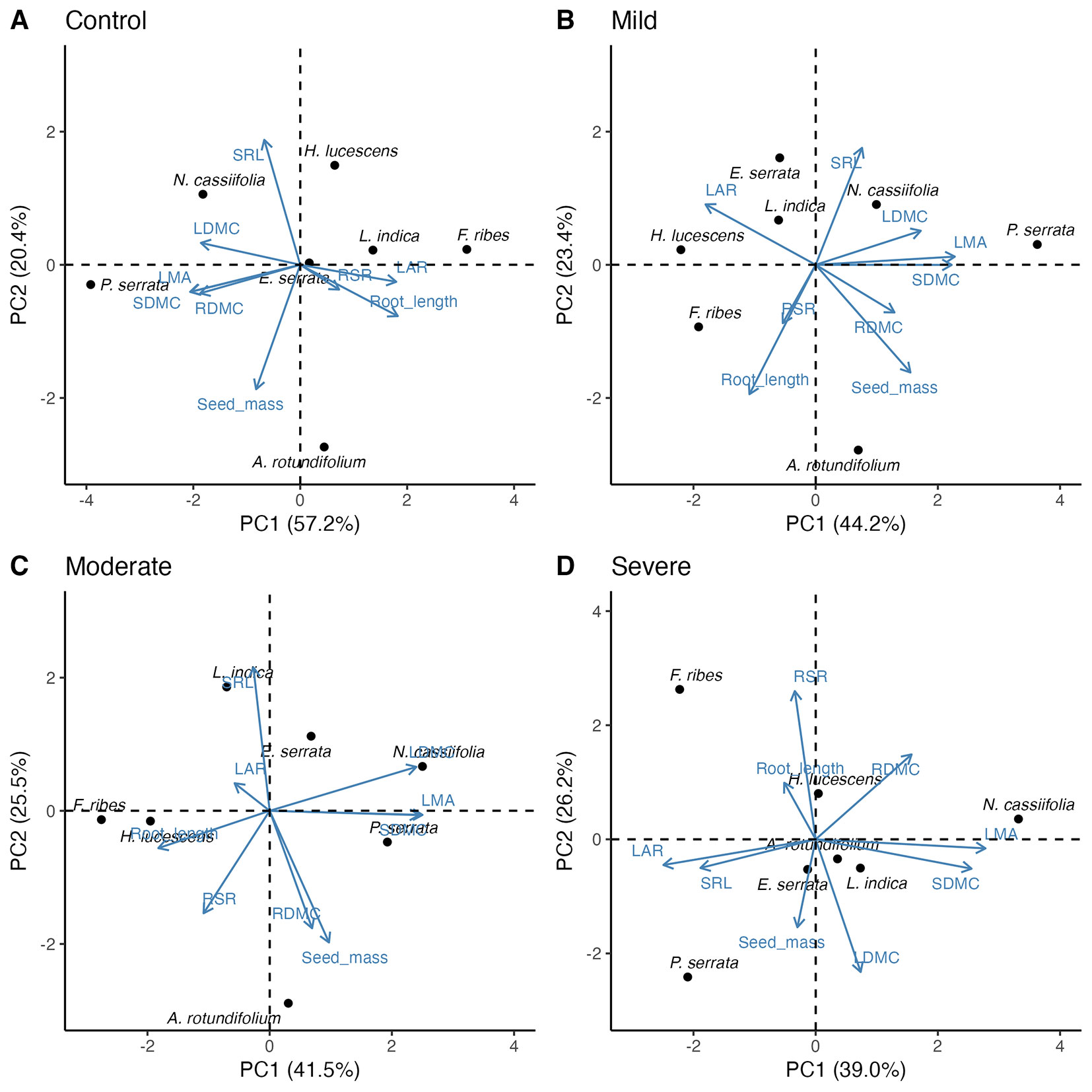
The Spearman correlation analysis revealed distinct relationships among traits across drought-stress levels (Tab. S3 in Supplementary material). Notably, LMA and SDMC exhibited robust positive correlations across all conditions. The first two principal components explained a substantial portion of the variation (65.16%-77.63% - Fig. 3, Tab. S4). PCA revealed that control plants prioritized light capture, exhibiting low LMA, whereas stressed plants adopted a water-use-efficiency strategy, with higher LMA. Root morphology shifted from longer, less dense roots in control plants to shorter, denser roots in stressed plants. LAR declined, and LDMC increased under stress, reflecting resource reallocation. Inter-specific trait variation was evident, with some species displaying consistent trait responses (e.g., A. rotundifoilum, H. lucescens) and others showing more complex patterns (e.g., E. serrata, N. cassiifolia). Notably, P. serrata exhibited extreme trait plasticity across drought gradients.
Fig. 3 - Biplot of principal component analysis for the first and second axes of 9 traits for control (A), mild (B), moderate (C), and severe drought treatments (D). Points are species in trait space. See Tab. 2 for trait abbreviations.
Trait relationships to seedling performance
The analysis identified RSR and LMA as negatively correlated with seedling survival (p < 0.05 - Tab. S5 in Supplementary material), with substantial interspecific variability observed, particularly under severe drought stress, where survival declined more with increasing RSR and LMA (Fig. 4). This high value of R2 conditional (R2 = 0.71) suggests that the random effects (i.e., species) contribute significantly to explaining the variance in seedling survival. However, further analysis revealed no significant interaction between species and treatment (p > 0.05).
Fig. 4 - The relationships between the probability of seedling survival (A-B), relative growth rate (C-E), wilting states (F), and the significant functional traits (root-to-shoot ratio, leaf mass per area, seed mass, root dry matter content, leaf dry matter content) across four treatment conditions (control, mild, moderate, severe). The colors represent drought treatments, the lines depict model predictions, and the scatter points represent individual data points for each treatment. All models were corrected for heteroscedasticity.
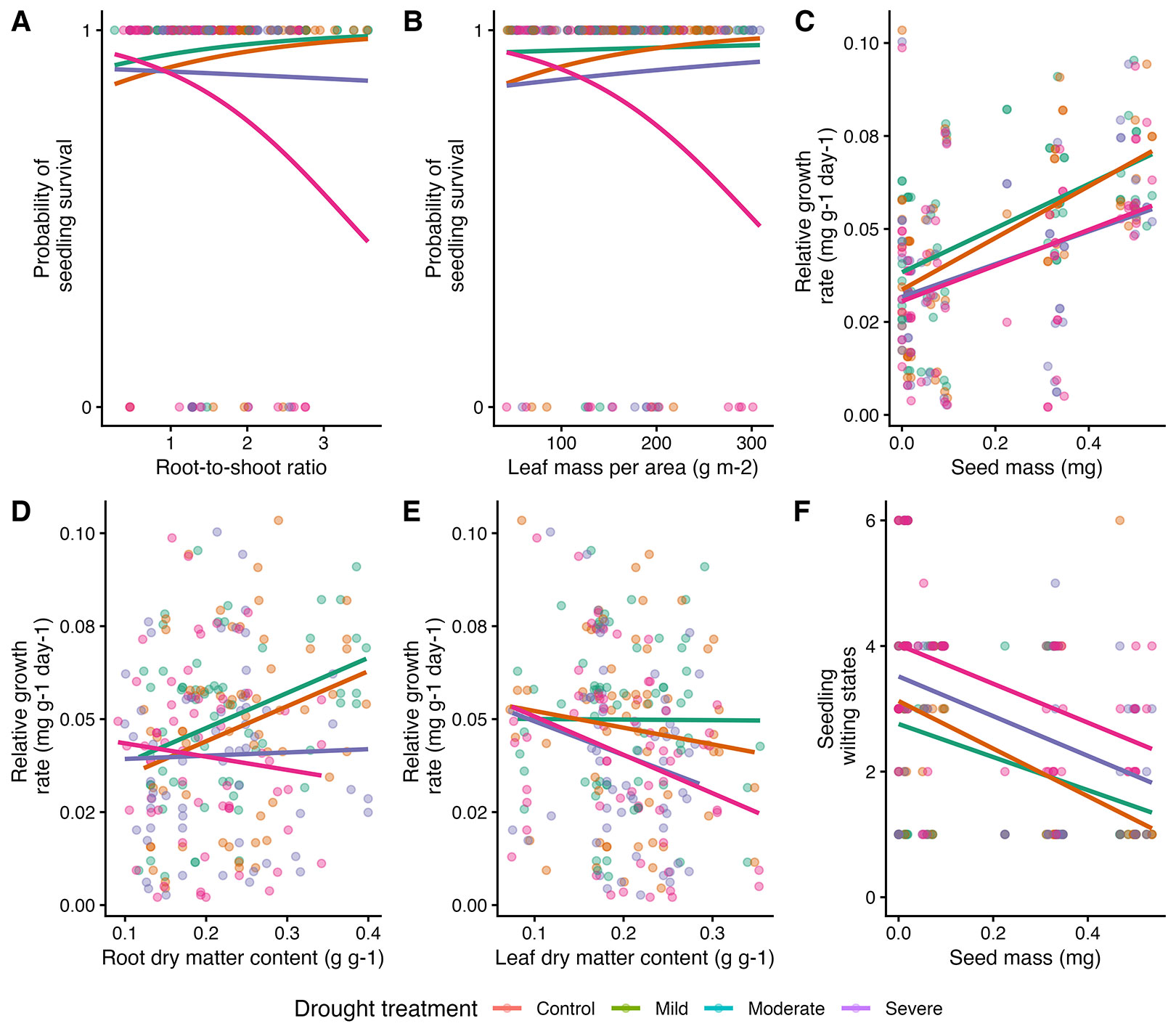
Seed mass and RDMC positively influenced relative growth rate (p < 0.01 and p < 0.05, respectively - Fig. 4, Tab. S5), indicating that larger seeds and higher RDMC contribute to faster growth. However, the beneficial impact of RDMC on growth was reduced under moderate and severe drought stress. Conversely, LDMC negatively affected growth rates (p < 0.05). Additionally, larger seeds exhibited lower wilting rates (p < 0.05 - Fig. 4, Tab. S5), underscoring their role in drought tolerance.
Treatment and species effects on traits
Drought stress significantly affected several seedling traits (Tab. S6 in Supplementary material). The most consistent responses to drought stress involved decreased LMA, indicating less investment in leaf structure, and reduced SDMC and RDMC, suggesting lower allocation to storage reserves. Species significantly influenced all traits (p < 0.01), including resource acquisition and allocation traits. Significant interaction terms across most traits (excluding root length, SRL, and seed mass) indicate species-specific drought-stress responses.
Discussion
Drought stress significantly reduced soil moisture and seedling survival, with severe drought causing the highest mortality. It also led to lower relative growth rates and increased wilting. Functional traits shifted under drought, with higher LMA and SDMC enhancing water conservation. RSR and LMA were negatively correlated with survival under severe drought, whereas seed mass and RDMC were positively correlated with growth. Species identity played a key role in drought responses, with species-specific adaptations in survival-related traits under water stress.
Seedlings response to drought stress
The results confirm the hypothesis that seedlings of the seven tropical woody species exhibit distinct responses to drought stress, as reflected in their survival rates, growth performance, and wilting. Overall, survival declined with increasing drought severity, with severe drought significantly reducing survival. However, H. lucescens, L. indica, A. rotundifolium, and P. serrata showed high drought tolerance with no mortality, while E. serrata, F. ribes, and N. cassiifolia had lower survival, indicating greater susceptibility. These findings align with previous studies on species-specific drought resistance of tropical species ([13]).
Drought reduced relative growth rates across species, with A. rotundifolium maintaining the highest and E. serrata the lowest. This pattern aligns with studies linking drought tolerance to water-use efficiency and resource allocation strategies ([33]). Under drought stress, stomatal closure limits CO2 uptake, reducing carbon assimilation and increasing the risk of photoinhibition, which can ultimately impair plant growth and survival ([10]). Additionally, prolonged water stress leads to the accumulation of reactive oxygen species (ROS), causing oxidative damage to cellular structures, including the photosynthetic apparatus, further constraining plant performance under drought conditions ([10]).
Wilting responses varied, with E. serrata, F. ribes, and L. indica wilting early, while A. rotundifolium, H. lucescens, and P. serrata exhibited minimal drought stress. Species with higher drought tolerance maintained leaf water status longer, delaying wilting and reducing mortality, whereas drought-sensitive species wilted early due to limited water retention capacity ([13]). Tolerance of low leaf water potential is crucial for drought survival, enabling desiccation-tolerant species to persist in drier environments while restricting drought-sensitive species to wetter habitats. Delayed wilting is often associated with physiological traits such as lower stem hydraulic conductance and greater resistance to xylem embolism, which enhance water transport efficiency under drought stress ([24]).
Functional trait variabilities and trade-offs under drought treatments
Our results support the hypothesis that resource allocation traits exhibit adaptive patterns under drought stress, particularly in traits related to water-use efficiency. While LMA and SDMC increased under drought conditions, indicating a shift toward greater resource conservation, root traits exhibited more complex responses. Instead of a uniform increase in root biomass, drought-stressed plants developed shorter, denser roots than the longer, less dense ones observed in control conditions.
This study revealed robust correlations between LMA and SDMC across all drought treatments, suggesting a coordinated plant response to water deficit. Linear mixed effects modeling confirmed the significant influence of drought on both traits. These findings align with previous research demonstrating the plasticity of shoot and leaf traits in response to water limitation ([15]). Consistent with prior research, plants exhibited elevated LMA and SDMC under drought conditions ([35], [1]). High LMA is often associated with resource conservation and photosynthetic efficiency ([35]), whereas increased SDMC enhances stem hydraulic function and mechanical stability ([1]).
The PCA results revealed a significant shift in plant traits from a growth-oriented strategy under control conditions to a water-conservation strategy under severe drought stress. PC1 captures this transition, with positive loadings on LMA (thicker leaves), SDMC (denser stems), and RDMC (root density), all of which are strongly associated with drought tolerance. High LMA is particularly notable, as leaves with high LMA typically feature thicker cell walls that help minimize water loss and maintain leaf function during dry conditions ([35]). Similarly, high SDMC, which is positively correlated with drought indices ([36]), reflects adaptations such as narrower vessels and thicker cell walls, reducing vulnerability to xylem cavitation - a primary factor contributing to tree death in drought-prone areas ([36]). High RDMC in plants, driven by increased nonstructural carbohydrates and inorganic nitrogen in roots under drought, enhances water uptake and reduces losses, thereby aiding adaptation to water scarcity ([39]).
Despite exhibiting high drought tolerance, as indicated by PCA results, species such as N. cassiifolia do not necessarily translate this tolerance into high seedling performance. In contrast, A. rotundifolium, which shows a more moderate drought response, consistently maintains high seedling performance across all drought conditions. This pattern, supported by linear mixed-effects models, suggests that most trait-performance relationships are independent of drought treatments. Such findings are consistent with previous studies indicating limited intraspecific variation in traits and local adaptation in seedling performance ([25]).
Key predictors of seedling performance under drought stress
The hypothesis that resource allocation traits predict seedling performance under drought stress is partially supported. While RSR and LMA were expected to enhance drought tolerance, they were negatively correlated with seedling survival under severe drought stress. This suggests that excessive allocation to roots and structural investment in leaves may reduce survival in extreme conditions. In contrast, RDMC positively influenced growth, whereas seed mass was associated with wilting, supporting the hypothesis that resource allocation traits affect plant performance.
The best predictors of drought-induced seedling survival are the RSR and LMA, both of which negatively affect survival under severe drought but contribute to tolerance under milder conditions. The decline in RSR under severe drought may be linked to reductions in root length and diameter, as observed in a meta-analysis of drought experiments ([44]). These findings suggest that extreme water stress limits root expansion and resource acquisition, particularly in trees, by increasing root tissue density while reducing overall root growth ([44]). As drought intensifies, balanced resource allocation becomes critical, as excessive root investment can enhance water absorption ([29]) but may reduce photosynthetic capacity and ultimately lower survival.
Similarly, higher LMA improves drought tolerance by reducing water loss through thicker leaves ([28]) but can limit light capture and photosynthesis efficiency ([35]). These findings align with studies in tropical dry forests, in which seedlings allocate more resources to structural adaptations that enhance survival at the expense of rapid growth ([18]). Balancing LMA for water conservation and resource acquisition is therefore crucial for seedling persistence under drought ([35]).
Species significantly contribute to the variance in seedling survival due to their diverse responses to environmental factors like drought stress and soil moisture. These differences are crucial for understanding species distribution and how species-specific responses to moisture availability shape plant communities at local and regional scales ([13]). Additionally, species exhibit different reactions to key abiotic and biotic factors, indicating niche differences that enable coexistence ([21]). This finding parallels studies in tropical dry forests, where species’ inherent hydraulic traits and carbon-investment strategies are stronger predictors of survival than short-term acclimation ([17]).
The positive association between relative growth rate and both seed mass and RDMC observed under optimal conditions was attenuated under both moderate and severe drought stress. These findings challenge the established notion that larger seeds are associated with slower growth rates ([46]). Given the absence of large seeds (>5 g), in this study, which are known to provide substantial reserves for initial growth and stress resilience ([30]), the smaller seed size of the studied species may account for the observed pattern.
Plant growth tends to increase with higher RDMC. However, this trend may shift under resource-limited conditions ([44]). Seedlings under drought stress exhibit increased RDMC as a strategy to enhance water acquisition under drought conditions. This adaptive response, characterized by greater biomass allocation to belowground structures and the development of deeper root systems ([29]), is consistent with previous findings indicating that RDMC is positively associated with drought resistance in tropical tree species ([36]). Although factors such as high turgor loss and nitrogen limitation may contribute to elevated RDMC ([28]), this metric remains a reliable and cost-effective proxy for fine root tissue density ([6]), providing valuable insights into plant drought responses.
Our study found that larger seed size reduces wilting frequency in seedlings, confirming previous research indicating that insufficient water induces wilting ([13]). Leaf water potential at the turgor loss point, a metric for plant drought response, indicates when stomata close to minimize water loss, leading to wilting and changes in leaf water content, potential, and hydraulic conductance ([3]). Wilting typically occurs under severe drought, with wilting points determining drought tolerance ([28]). Tropical trees exhibit varying drought tolerance, with wilting points ranging from -1.4 to -3.2 MPa, reflecting adaptations to dry seasons ([28]).
Our findings suggest that a functional-trait-based approach is limited in its ability to predict seedling performance under drought. While functional traits provide valuable insights into plant characteristics and stress adaptation, species-specific responses and environmental variability may also influence seedling performance. Functional traits are measurable characteristics reflecting plant-environment interactions and are commonly used to predict plant responses to stressors like drought ([17]). However, our study suggests that functional traits may not accurately predict seedling survival under drought conditions. Different species may exhibit unique drought tolerance strategies that are not fully captured by functional traits. For instance, some species may possess internal mechanisms for water conservation or stress resistance that are not reflected in their external traits and differ at the species level ([42]). Additionally, the severity and duration of drought events can impact seedling performance, and functional traits may not account for specific drought conditions ([44]). Furthermore, the complex interactions among traits are challenging to capture with a single trait-based approach ([34]).
Our findings parallel the shift toward drought-tolerant species in Ghana’s forests ([14]), suggesting consistent adaptive mechanisms across developmental stages. In Ghana, prolonged drought favored deciduous, drought-resilient canopy species over evergreen sub-canopy species. Similarly, seedlings with drought-associated traits in our study showed greater persistence under water stress, emphasizing the role of functional traits in shaping forest composition. A comprehensive understanding of drought tolerance requires integrating trait-based approaches with species-specific physiology, environmental conditions, and genetic variation.
Recommendations for species selection in restoration projects
Active restoration, such as enrichment planting, is critical for recovering secondary and degraded tropical forests subjected to recurrent disturbances ([27]). Standard restoration methods, including planting nursery-raised seedlings and direct seeding, often encounter challenges, particularly during the establishment phase under dry conditions. Species selection is paramount for successful restoration in these degraded ecosystems. For direct seeding, prioritizing drought-resilient species is essential, including A. rotundifolium and P. serrata. Species with higher seed mass, favorable RSR, and elevated LMA typically exhibit superior performance across varying drought conditions, including severe drought. These traits promote robust seedling establishment and growth under water-limited environments. Direct seeding is particularly advantageous for these high-performing species because it requires less pre-sowing care and minimal intervention. This method is cost-effective for restoring isolated, degraded areas but requires careful consideration of potential challenges, such as reduced survival rates under drought stress.
Conversely, species with lower drought tolerance or greater variability should be propagated in nurseries, e.g., E. serrata, F. ribes, and N. cassiifolia. These species benefit from controlled conditions that enhance their growth and drought resilience before field planting. Focus on species with lower seed mass, reduced RSR, and less effective water-conserving traits. Nursery cultivation allows for better management and acclimatization, thereby improving the species’ survival and establishment in degraded habitats. This approach ensures that even species with less inherent drought resilience can contribute effectively to restoration efforts. While drought-tolerant species are crucial, incorporating a diverse array of species with varying drought-tolerance strategies can enhance long-term ecosystem resilience. Further research into the seed enhancement approach is needed to improve survival and growth outcomes.
Conclusion
Drought stress reduced soil moisture and seedling survival, with severe drought causing the highest mortality. Functional traits exhibited adaptive shifts, with increased LMA and SDMC enhancing water conservation, while root traits showed complex responses. Species-specific differences confirmed that seedlings respond variably to drought, supporting the first hypothesis. The second hypothesis was partially supported, as increased RSR was not consistently observed, and RSR and LMA were negatively correlated with survival under severe drought. The third hypothesis was supported, RDMC positively influenced growth, whereas seed mass was associated with wilting. Effective restoration efforts require selecting species based on their drought sensitivity and matching them with suitable restoration techniques to maximize success.
Acknowledgements
MZ, NNS, SPD, and ES contributed equally to this work. MZ, NNS, SPD, and ES designed the project. MZ conducted the experiment, performed the analysis, and wrote the first draft of the manuscript. MZ, NNS, SPD, and ES revised the manuscript and supervised. ES received the funding.
This study was funded by the Institut Teknologi Bandung, Indonesia, through Ganesha Talent Assistantship-Research Group (498/IT1.A/SK-KM/2021) and PPMI-SITH (32/IT1.C11/SK-TA/2021). We sincerely thank the Research Center for Applied Botany, National Research and Innovation Agency, Indonesia, for providing seed materials and laboratory access, as well as the volunteers who assisted with seed collection. We also appreciate the anonymous reviewers for their insightful comments, which greatly improved the manuscript.
References
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Authors’ Info
Authors’ Affiliation
Susana Paulina Dewi 0000-0002-5802-5660
Endah Sulistyawati 0000-0003-1843-0570
School of Life Sciences and Technology, Institut Teknologi Bandung, Jl. Ganesha 10, Bandung, West Java 40132 (Indonesia)
Research Center for Applied Botany, National Research and Innovation Agency, Jl. Raya Jakarta - Bogor Km. 46, Cibinong, Bogor, West Java 16911 (Indonesia)
MKi, Hungarian Research Institute of Organic Agriculture, Ráby Mátyás u. 26, 1038 Budapest (Hungary)
Corresponding author
Paper Info
Citation
Zuhri M, Setiawan NN, Dewi SP, Sulistyawati E (2026). Tropical seedling performance under drought: a functional trait approach for species selection in restoration. iForest 19: 9-17. - doi: 10.3832/ifor4713-018
Academic Editor
Tamir Klein
Paper history
Received: Aug 26, 2024
Accepted: Jun 10, 2025
First online: Jan 10, 2026
Publication Date: Feb 28, 2026
Publication Time: 7.13 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2026
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 4326
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 1863
Abstract Page Views: 1293
PDF Downloads: 1044
Citation/Reference Downloads: 1
XML Downloads: 125
Web Metrics
Days since publication: 183
Overall contacts: 4326
Avg. contacts per week: 165.48
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
(No citations were found up to date. Please come back later)
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Addressing post-transplant summer water stress in Pinus pinea and Quercus ilex seedlings
vol. 8, pp. 348-358 (online: 16 September 2014)
Short Communications
Differential adaptations in nursery seedlings from diverse Chilean provenances of Peumus boldus Mol.
vol. 9, pp. 409-413 (online: 29 January 2016)
Research Articles
Can the dynamics of forest restoration reduce landscape fragmentation in the Atlantic forest?
vol. 18, pp. 61-68 (online: 04 April 2025)
Review Papers
Problems and solutions to cork oak (Quercus suber L.) regeneration: a review
vol. 16, pp. 10-22 (online: 09 January 2023)
Research Articles
Effect of drought stress on some growth, morphological, physiological, and biochemical parameters of two different populations of Quercus brantii
vol. 11, pp. 212-220 (online: 01 March 2018)
Research Articles
Outplanting performance of three provenances of Quillaja saponaria Mol. established in a Mediterranean drought-prone site and grown in different container size
vol. 13, pp. 33-40 (online: 21 January 2020)
Research Articles
Oak sprouts grow better than seedlings under drought stress
vol. 9, pp. 529-535 (online: 17 March 2016)
Research Articles
Towards a functional phytosociology: the functional ecology of woody diagnostic species and their vegetation classes in Northern Italy
vol. 14, pp. 522-530 (online: 22 November 2021)
Research Articles
Matching seedling size to planting conditions: interactive response with soil moisture
vol. 12, pp. 220-225 (online: 25 April 2019)
Research Articles
Advantages of the point-intercept method for assessing functional diversity in semi-arid areas
vol. 8, pp. 471-479 (online: 31 October 2014)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







