
Outplanting performance of three provenances of Quillaja saponaria Mol. established in a Mediterranean drought-prone site and grown in different container size
iForest - Biogeosciences and Forestry, Volume 13, Issue 1, Pages 33-40 (2020)
doi: https://doi.org/10.3832/ifor3142-012
Published: Jan 21, 2020 - Copyright © 2020 SISEF
Research Articles
Abstract
In degraded environments with restricted seed availability, like those burned by wildfires in central Chile during 2017, the use of plant material from outside its area of origin for restoration purposes needs to be accurately investigated. We assessed the early development of three Chilean provenances of Quillaja saponaria grown in different container sizes (140 vs. 280 mL) in the nursery and then outplanted in a common field trial at a site severely affected by fire under Mediterranean-type climate. We analyzed growth, biomass, and leaf-level physiological traits. In the nursery, there was a significant provenance by container type interaction for the biomass traits (P < 0.05). Seedlings from the Maule provenance cultivated in larger containers had the highest biomass, while the lowest biomass was observed for the Metropolitan provenance cultivated in small containers. Two years after outplanting, the provenance by container size interaction was significant for stomatal conductance and chlorophyll density. Seedlings from the Metropolitan provenance cultivated in larger containers exhibited a higher stomatal conductance, while those from the Maule provenance cultivated in small containers exhibited the highest chlorophyll density. Seedling height showed significant variation for provenance and container size. The tallest seedlings were those grown in larger containers from the Maule provenance; however, no differences in survival and height increment were found. Gas exchange parameters differed among provenances, the Metropolitan provenance had a low performance and the opposite was found for the Biobío and Maule provenances. This study demonstrated that different provenances of Q. saponaria have stable performances in a Mediterranean site, which support their use for restoration purposes outside their home area with no detrimental effects on outplanting performance.
Keywords
Restoration, Quillay, Dry Site, Water Stress, Pot Size, Seed Origin
Introduction
Quillaja saponaria Mol. is a sclerophyllous evergreen tree species widely distributed in Chile, extending longitudinally over 650 km from the xeric shrubland in the north to the Mediterranean and temperate forests of the south (latitudinal range from 30° to 38° S - [11]). The forest types where the species can be found have been degraded in the past by land conversion for agricultural uses, and more recently by the devastating fires that occurred in central Chile during 2017 ([13]), which burned almost half a million hectares of land, of which 17.4% corresponds to native forests ([10]). This has raised a significant concern about the factors involved in the successful broad-scale seed-based restoration of Q. saponaria and other sclerophyllus species.
Banister et al. ([3]) pointed out that the poor quality of nursery stock for native species is one of the main bottlenecks that constrain forest landscape restoration in Chile. Some studies have highlighted the importance of managing the stock quality on the successful early growth in restoration projects ([22], [23]), while other studies have focused on proper selection of the seed provenance ([4]). However, no previous investigations have examined the potential combined effects of these factors on Q. saponaria, which may provide guidance for restoration of the species.
It is well known that autochthonous seed sources are better adapted to local conditions, exhibiting higher relative fitness in their original habitat ([6], [7]). Performance increases as local seeds are planted near to their home site ([25]). This is likely related to specific genetic adaptations of a provenance to local conditions that confers the highest fitness in their home environment when compared to non-local populations. However, the use of strict local plant material may limit the scale of restoration projects in areas severely altered ([18]), due to restrictions in seed availability ([8]). In the case of Q. saponaria, Ovalle et al. ([31], [32]) found high outplanting survival rates when using local provenances (i.e., seeds collected in a range of 40 km from the field test site). In a nursery experiment, Magni et al. ([28]) found that different provenances of Q. saponaria exhibited a stable performance under controlled growth conditions. However, the extent to which the provenance effect contributes to the successful restoration of the species is unknown. In other sclerophyllus evergreen tree species, Gratani et al. ([20]) found superior morphological and physiological responses for Quercus ilex L. provenances established close to their seed collection site.
Nursery cultural practices determine the quality of the planting stock and their functional characteristics ([35]). A positive relationship between seedling quality and successful restoration has been reported for numerous tree species ([46]). Seedlings with a well-developed root system rapidly grow roots after planting ([12]) and tend to have higher survival than seedlings with poor developed root systems ([22]). This can be manipulated by the container size ([15]). As the root system size is proportional to the container dimensions ([2]), seedlings cultivated in deep containers result in higher outplanting survival than seedlings cultivated in shallow containers in Mediterranean sites, the formers developing a large root system that quickly reaches moisture in deep soil horizons ([17], [9]). In a nursery experiment applying water restriction, Espinoza et al. ([16]) found that survival, growth, and biomass of water-stressed Q. saponaria were higher in seedlings cultivated in large containers than in small ones (280 mL of volume). Ovalle et al. ([31], [32]) found a high outplanting survival (80 to 100%) for Q. saponaria seedlings that were cultivated in 400 mL containers.
In this study we examined the performance of three provenances of Q. saponaria from different sites characterized by semiarid, humid, and very humid climate. Seedlings were cultured in the nursery in two container sizes and then outplanted in a site with Mediterranean-type climate. We aimed to assess the effect of stock quality and seed provenance on survival, growth, biomass, and physiological traits. The results of this study may guide the restoration decisions of Q. saponaria under Mediterranean-type climatic conditions.
Material and methods
Plant material
Three provenances of Q. saponaria located over 200 km from each other, and showing latitudinal-related climate differences, were studied (Tab. 1). In April 2016, dominant and co-dominant trees with well-developed crown, health condition, and abundant seed production were randomly selected from each provenance. Ripen fruits were collected from 10 to 20 mother trees per provenance. Seed quality was checked after collection according to ISTA standards ([24]).
Tab. 1 - Location and climatic parameters for the nursery and field experiments, and the seed collection sites of the three provenances of Q. saponaria under study. (MAP): mean annual precipitation (mm); (MAT): mean annual temperature (°C). The De Martonne aridity index was estimated as MAP/(MAT + 10).
| Experiment | Provenance | Location | Latitude S |
Longitude W |
MAP | MAT | De Martonne aridity index |
Climate type |
|---|---|---|---|---|---|---|---|---|
| Seed collection | Metropolitan | Interior | 33° 32′ | 71° 11′ | 436 | 16.7 | 16.3 | Semiarid |
| Maule | Coastal | 35° 14′ | 72° 18′ | 839 | 13.5 | 35.3 | Humid | |
| Biobío | Interior | 37° 09′ | 72° 32′ | 1122 | 13.4 | 47.9 | Very humid | |
| Nursery experiment | Maule | Interior | 35° 26′ | 71° 37′ | 757 | 14.6 | 30.7 | Humid |
| Field test site | Maule | Interior | 35° 34′ | 72° 06′ | 984 | 13.4 | 42.0 | Humid |
Nursery experiment
The study was conducted in a nursery under ambient conditions, located at the Universidad Católica del Maule (35° 26′ S, 71° 37′ W; 131 m a.s.l.), city of Talca, Chile. According to the De Martonne aridity index, the climate at the nursery is considered humid. Two container sizes were selected to produce planting stocks of different quality. A small container (typically used in Q. saponaria nurseries) was a tapered cylindrical pot (Termomatrices®, Santiago, Chile) with cell volumes of 140 mL (15 cm height × 4 cm top diameter × 3 cm bottom diameter) and 88 cavities per holding tray (cultivation density of 463 seedlings m-2). The large container (BASF®, Santiago, Chile) had rectangular cells of 280 mL (14 cm height × 5.5 cm top diameter × 4.5 cm bottom diameter) and each tray was made of 60 cells (cultivation density of 254 seedlings m-2). The substrate consisted of a mixture of composted bark of Pinus radiata D. Don and perlite (7:3 v/v), combined with the slow release fertilizer Basacote 9M® (16% N, 8% P2O5, 12% K2O, 12% SO3, 2% MgO, 0.02% B, 0.05% Cu, 4% Fe and 0.06% Mn - Compo Expert GmbH, Münster, Germany) at a dose of 4 g L-1. Seeds were sown on October 2016 and cultivated by 10 months. Seedlings were watered regularly at container capacity with an automatic irrigation system. Temperature and relative humidity averaged 19 °C and 70%, respectively for the cultivation period. Seedlings were arranged in a randomized complete block design, with a border row of extra seedlings around each tray to minimize edge effects. The total number of seedlings evaluated were 540 (i.e., 2 container sizes × 3 provenances × 3 replicates × 30 seedlings per replicate).
At field planting (August 2017), height (H) and root collar diameter (D) were measured for all the provenances in all seedlings using a metric tape and a digital caliper, respectively. Average H and D were 22.7 ± 0.8 cm and 3.5 ± 0.07 mm, respectively. In addition, a subsample of five seedlings per treatment and replication (i.e., 90 seedlings) were used to determine biomass components. Seedlings were harvested from the containers and oven-dried at 65 °C for 48 h. Roots, leaves, and stems were separately weighed (± 0.01 g), giving the root dry weight (RDW), the leaves dry weight (LDW), the stem dry weight (SDW), and the total dry weight (TDW). The root to shoot ratio was derived as RSR = RDW/(SDW+LDW). According to Niklas & Enquist ([30]) and Grossnickle ([22]), the root: shoot ratio reflects the balance between water acquisition of the root system (roots component) and water losses of the transpiring shoot system (leaves plus stems).
Field trial
A field trial was established in August 2017 in the locality Nirivilo (35° 34′ S, 72° 06′ W; 254 m a.s.l), San Javier, central Chile, on a flat site property of the Universidad de Chile. The site has a Mediterranean-type climate with no summer rainfall, 675 mm of rainfall from June to August, 396 mm of evaporation from December to February, and a maximum temperature of 25 °C during January. According to the De Martonne aridity index, the climate at the field test site is considered humid. The soil is neutral (6.1 pH), sandy clay (47% sand, 17% lime, 36% clay), with low electrical conductivity (0.03 dS m-1) and 1.5% organic matter content. The available soil nutrient composition was 4 mg kg-1 N, 8 mg kg-1 P, and 168 mg kg-1 K.
The site previously supported a 6-year-old P. radiata plantation and introduced pasture grasses that were both severely damaged by the fires that occurred during January 2017 in central Chile. The field experiment followed a randomized complete block design with two levels of container size, three provenances, and 3 replications of 7-seedlings per row (i.e., 126 seedlings in total). The site was fenced to avoid herbivore damage and seedlings were hand planted at a spacing of 1 × 1 m. No fertilizer was added to the planting hole. During the first summer season after outplanting, seedlings were watered with 4 L plant-1 month-1 from December 2017 to March 2018 in order to mitigate the negative effect of water stress on early performance. Furthermore, seedlings were protected with tree shelters (Surpack S.A., Lima, Peru) made up of green, polyethylene tubes (80 cm tall × 20 cm wide). Light transmittance into the shelters was measured with a LI-250 quantum sensor (LI-COR Inc., Lincoln, NE, USA) during January 2019, and it was 15% of full sunlight, i.e., photosynthetic active radiation (PAR) of 300 mmol m2 s1. Additionally, soil temperature, and water content at 10-cm depth were measured once a month during the spring-summer of the first growing season (from September 2017 to March 2018) in three points per replicate by using a soil temperature/moisture sensor (GS3® dielectric probe, Decagon Devices Inc., Pullman, WA, USA). Average values are shown in Fig. 1.
Fig. 1 - Average soil temperature and water content at the field test site during the first growing season (September 2017 to March 2018).

Morphological and leaf-level physiological measurements at field test site
At the end of the second growing season (May 2019), H, D, and survival (as a categorical trait: 1 = alive; 0 = dead) were measured for each seedling. The H increment was calculated as the difference in growth between May 2019 and August 2017. Additionally, the last week of January 2019 we measured light-saturated photosynthetic rate (Asat, µmol CO2 m-2 s-1), transpiration (E, mmol H2O m-2 s-1), stomatal conductance (gs, mol H2O m-2 s-1), and the derived intrinsic water use efficiency (WUEint= Asat/gs), in a fully-developed leaf obtained on the upper third of the seedlings (5 seedlings per experimental unit). These measurements were taken between 11:00 and 15:00 (local time) using a portable photosynthesis system (Li-6400XT®, LI-COR Inc., Lincoln, NE, USA) provided with a light source (Li-6400-02B). In order to avoid sudden changes in leaf light level we opened the shelters and quickly put a leaf into the chamber. Temperature, air CO2 concentration, and light source were set at ambient conditions to 25 °C, 400 ppm, and 1800 mmol m2 s1, respectively. Chlorophyll density measurements (i.e., SPAD values) were also obtained using a chlorophyll meter (Minolta SPAD-502® meter, Minolta Camera CO., Osaka, Japan) on three individual leaves per tree. The SPAD-502 meter provides indirect reliable estimates of relative densities of leaf chlorophyll ([38]).
Data analysis
All variables were subjected to an analysis of variance, except survival which was tested using a Chi-square test. The statistical model applied to both the nursery and field experiment was (eqn. 1):
where Yijkl is the measurement, µ is the overall mean, Bi is the fixed effect of the ith block, Pj is the fixed effect of the jth provenance (j = Metropolitan, Maule, and Biobío provenances), Ck is the fixed effect of the kth container size (k = 140 vs. 280 mL), P×Cjk is the interaction between provenance and container size, and eijkl is the experimental random error. Mean comparisons were made by the Tukey multiple range test (α = 0.05). Additionally, the relationship between seedlings initial characteristics at the nursery stage with field performance was investigated separately for each provenance through regression analysis. All the statistical analyses were performed with the software SPSS® ver. 18.0 (SPSS Inc., Chicago, IL, USA).
Results
Growth and biomass at nursery
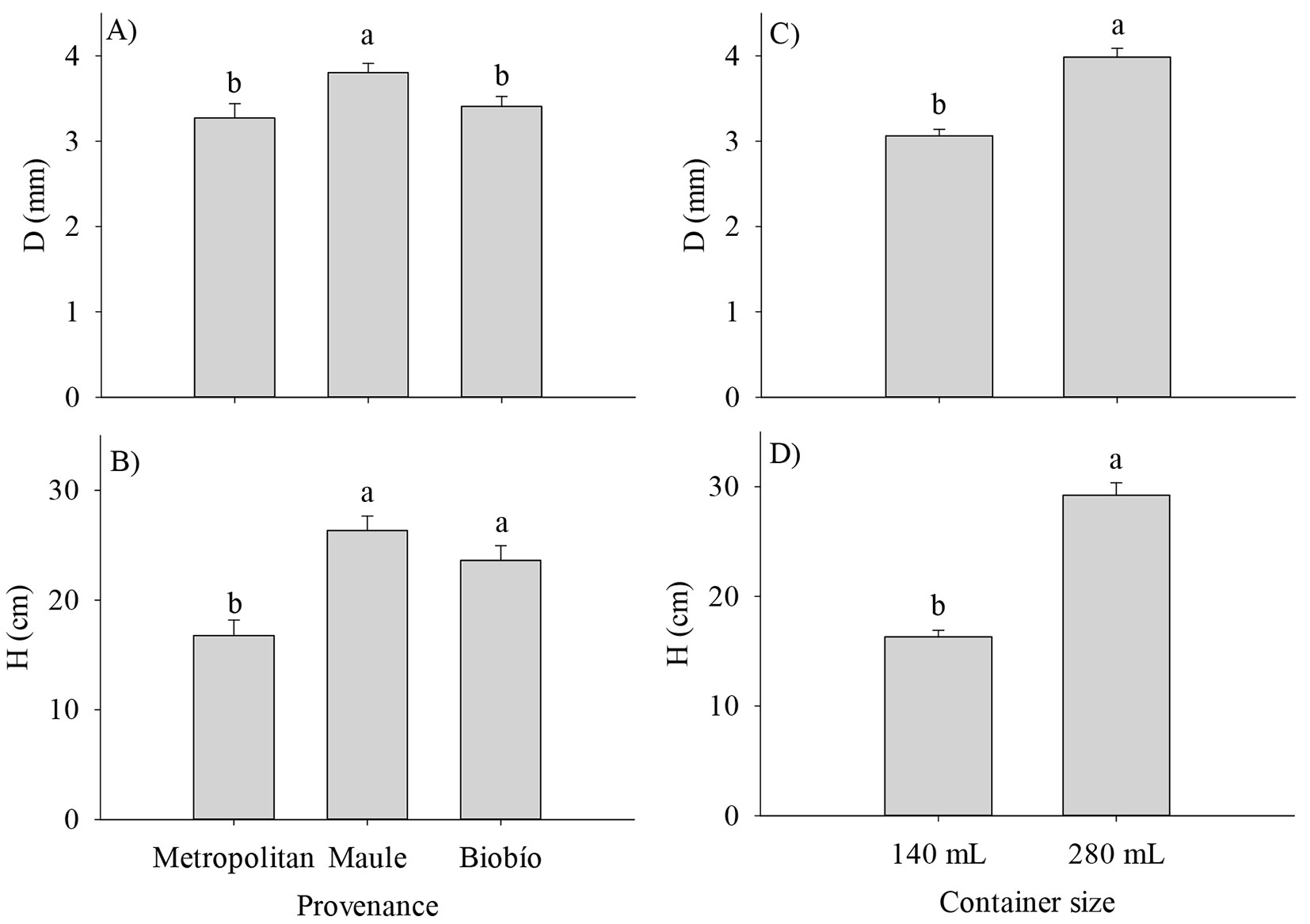
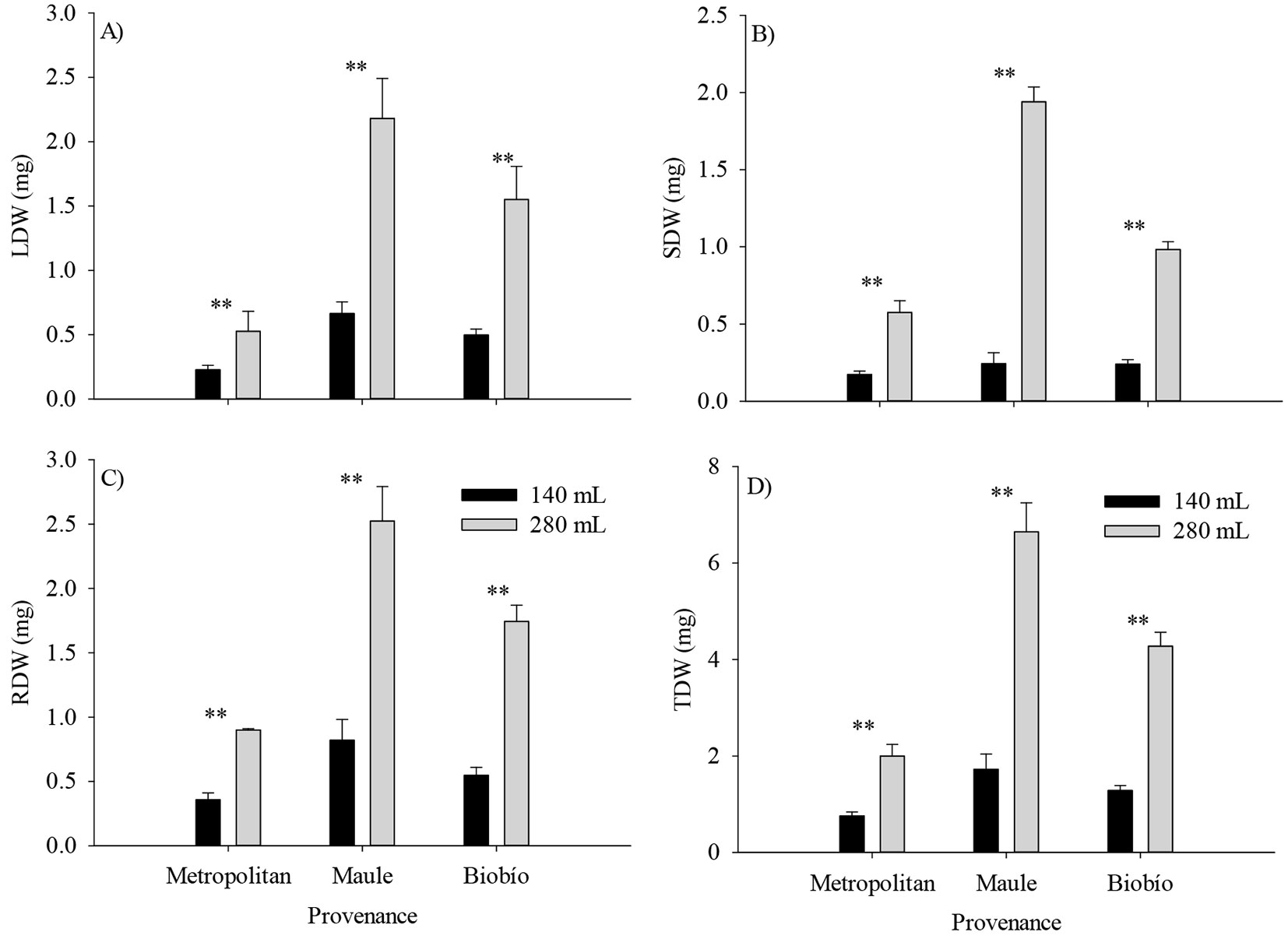
At the end of the nursery phase, the analysis on the main effects showed that the Maule and Metropolitan provenances had the highest and lowest values for D and H (Fig. 2A, B). Similarly, larger containers produced larger seedlings (P < 0.05 - Fig. 2C, D). Interestingly, H was 2-fold higher in the larger containers. The effect of the provenance by container size interaction was significant for most of the biomass components but not for H, D, and RSR (Tab. 2). The highest responses in TDW, RDW, SDW, and LDW to the container size of the Maule provenance relative to the Metropolitan provenance explained the provenance by container size interaction (Fig. 3).
Fig. 2 - Growth traits of Q. saponaria seedlings at the end of the nursery stage. (D): root collar diameter; (H): height. Different letters indicate significant (p<0.05) differences among means. The error bars indicate the standard error.
Tab. 2 - Significance of the main factors and interactions on growth and biomass traits of Q. saponaria seedlings as derived from the analysis of variance (ANOVA). (P): provenance; (C): container size; (D): root collar diameter; (H): height; (RDW): roots dry weight; (LDW): leaves dry weight; (SDW): shoot dry weight; (TDW): total dry weight; (RSR): root to shoot ratio.
| Trait | P | C | P × C |
|---|---|---|---|
| D | 0.02 | <0.01 | 0.13 |
| H | <0.01 | <0.01 | 0.30 |
| RDW | <0.01 | <0.01 | 0.02 |
| LDW | <0.01 | <0.01 | <0.01 |
| SDW | <0.01 | <0.01 | 0.01 |
| TDW | <0.01 | <0.01 | 0.00 |
| RSR | 0.38 | 0.16 | 0.38 |
Fig. 3 - Effect of the provenance by container size interaction on biomass traits of Q. saponaria seedlings at the end of the nursery stage. (LDW): leaves dry weight; (SDW): shoot dry weight; (RDW): roots dry weight; (TDW): total dry weight; (**): p<0.01; (*): p<0.05. The error bars indicate the standard error.
Survival, growth, and leaf-level physiology at field- test site
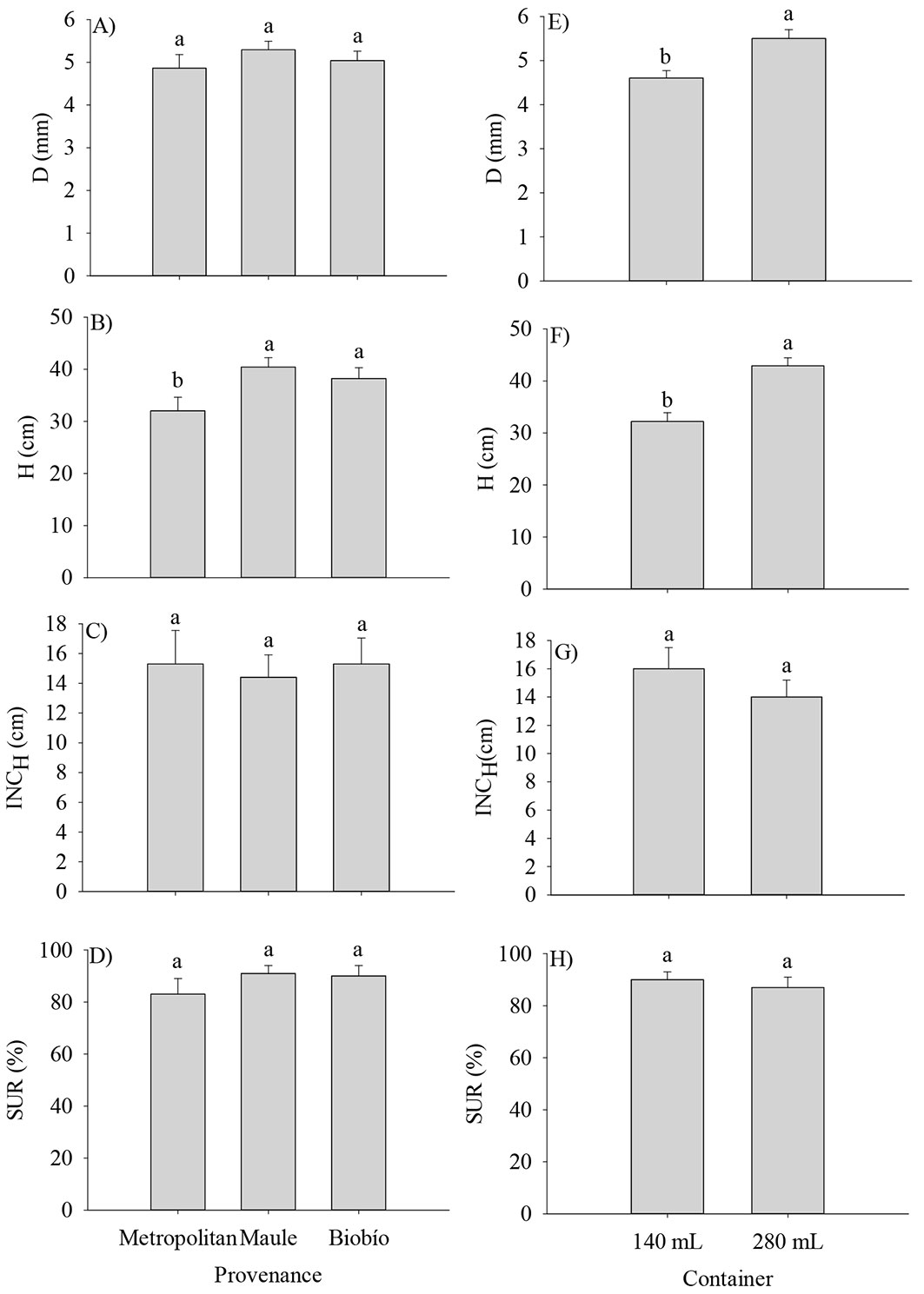
Two growing seasons after outplanting, the analyses of main effects revealed that D and H were higher in large containers, and H was superior in the Maule and Biobío provenances (Fig. 4). There were no effects of the provenance, the container size, or their interaction on D, survival, and H increment (P > 0.05 - Fig. 4A, C, D, G, H).
Fig. 4 - Growth and survival of Q. saponaria seedlings after two growing seasons in the field test site. (D): root collar diameter; (H): height; (INCH): increment for height; (SUR): survival. Different letters indicate significant (p<0.05) differences among means. The error bars indicate the standard error.
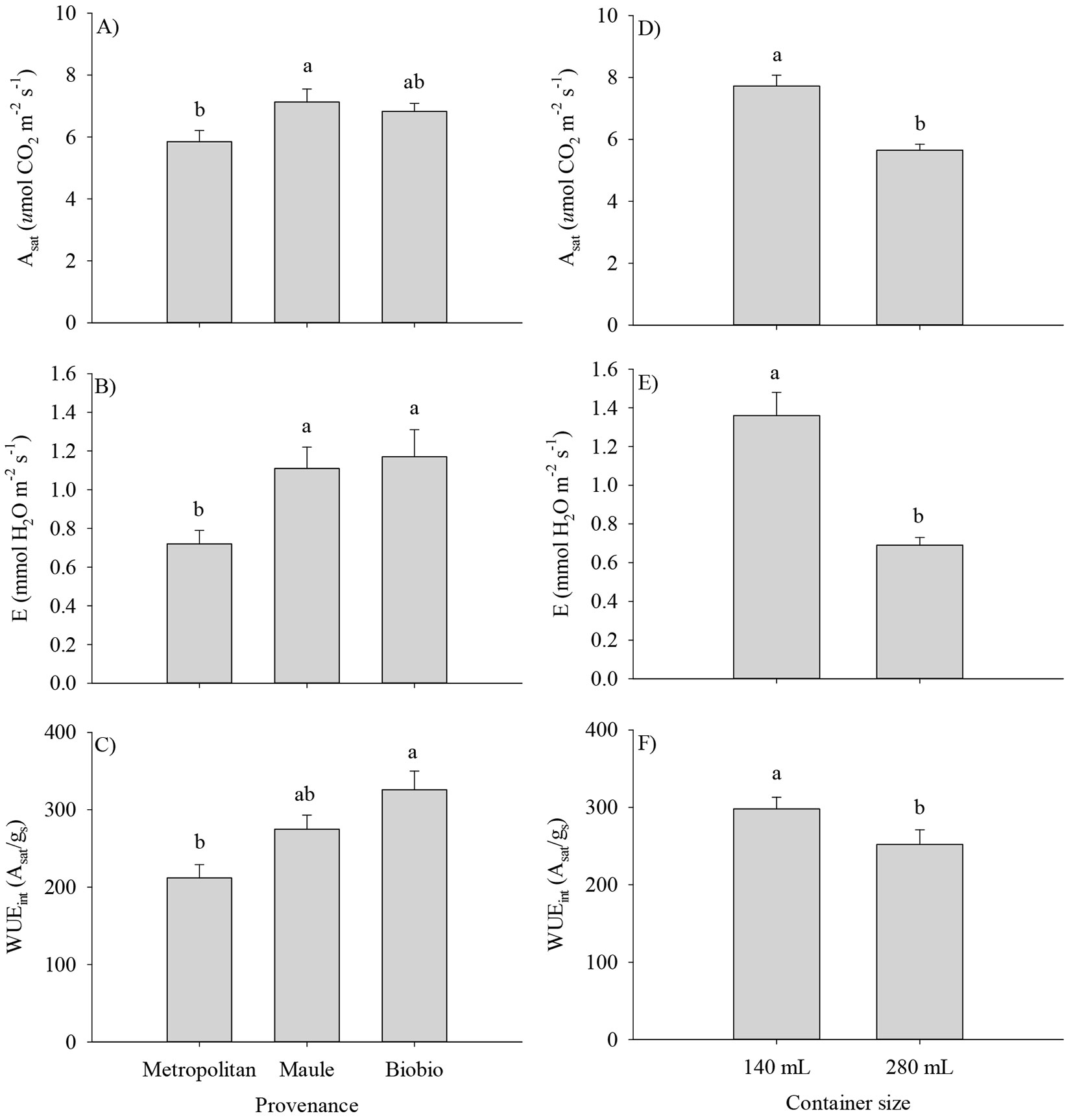
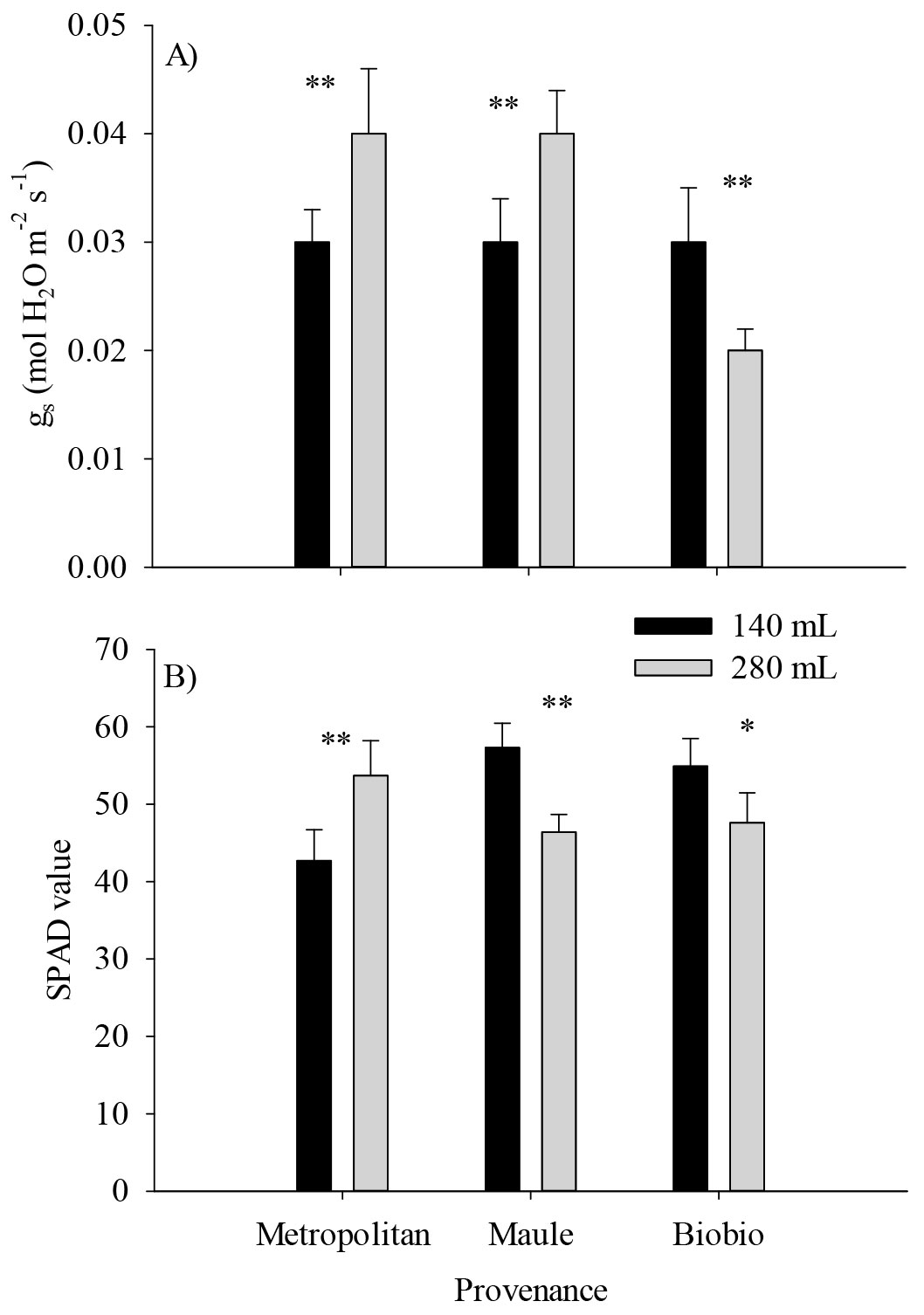
For gas exchange traits, means for Asat, E, and WUEint were lower for the Metropolitan provenance (Fig. 5A, B, C), and in the case of the container size, seedlings cultivated in small containers exhibited the highest Asat, E and WUEint (Fig. 5D, E, F). There was a significant effect of the provenance by container size interaction on gs and chlorophyll density (i.e., SPAD value - Tab. 3). Provenances differed in gs only in the larger containers, with the Maule and Metropolitan provenances doubling the values of the Biobío provenance (Fig. 6A). Unlike, the chlorophyll density in the Maule and Biobío provenances was higher in small but lower in the large containers when compared with the Metropolitan provenance (Fig. 6B).
Fig. 5 - Gas exchange traits of Q. saponaria seedlings after two growing seasons at the field test site, according to the provenance (panels A, B, C) and container size (panels D, E, F). (Asat): light-saturated photosynthetic rate; (E): transpiration; (WUEint): intrinsic water use efficiency. Different letters indicate significant (p<0.05) differences among means. The error bars indicate the standard error.
Tab. 3 - Significance of main factors and interactions on gas exchange traits of Q. saponaria seedlings derived from the analysis of variance (ANOVA). (P): provenance; (C): container size; (Asat): light-saturated photosynthetic rate (umol CO2 m-2 s-1); (E): transpiration (mmol H2O m-2 s-1); (gs): stomatal conductance (mol H2O m-2 s-1); (WUEint): Intrinsic water use efficiency (Asat/gs); (SPAD value): chlorophyll density.
| Trait | P | C | P × C |
|---|---|---|---|
| Asat | <0.01 | <0.01 | 0.64 |
| gs | 0.20 | 0.52 | <0.01 |
| E | <0.01 | <0.01 | 0.13 |
| WUEint | <0.01 | 0.03 | 0.06 |
| SPAD value | 0.56 | 0.40 | 0.01 |
Fig. 6 - Effect of the provenance by container size interaction on stomatal conductance (A) and chlorophyll density (B) of Q. saponaria seedlings after two growing seasons at the field test site. (**): p<0.01; (*): p<0.05. The error bars indicate the standard error.
Relationship between nursery stock and field performance
As expected, when comparing the correlations of D and H between the nursery and field test site, these were high and positive. However, with the exception of Biobío provenance, the relationship between seedlings characteristics measured at the nursery with survival and height increments in the field, showed low and poor correlations (Tab. 4).
Tab. 4 - Pearson’s correlation coefficients between the initial quality characteristics of seedlings and field performance two years after planting. (D): root collar diameter; (H): height; (RDW): roots dry weight; (LDW): leaves dry weight; (SDW): shoot dry weight; (TDW): total dry weight; (RSR): root to shoot ratio; (SUR): survival; (INCH): increment for height; (**): p<0.01; (*): p<0.05.
| Provenance | Seedling traits at nursery |
Seedling traits at field | |||
|---|---|---|---|---|---|
| D | H | SUR | INCH | ||
| Metropolitan | D | 0.88** | 0.70** | 0.47 | 0.12 |
| H | 0.69** | 0.94** | 0.29 | 0.09 | |
| Maule | D | 0.74** | 0.54** | 0.09 | 0.02 |
| H | 0.46** | 0.92** | 0.07 | 0.09 | |
| Biobío | D | 0.73** | 0.60** | 0.78* | 0.12 |
| H | 0.58** | 0.89** | 0.86* | 0.09 | |
Discussion
Growth and biomass at nursery level
The three provenances under study showed differences in growth traits at the nursery level, but some of such differences tend to disappear in the field. Seedlings from the Maule provenance exhibited superior H, aboveground (LDW, and SDW) and belowground (RDW) biomass. This superior performances may be probably related to the similarity of climatic conditions between seed collection site and nursery. Indeed, seeds were collected at just 12 minutes of latitude away from the nursery and both environments exhibited similar aridity index. On the contrary, the Metropolitan provenance seedlings, whose seeds were collected at latitude 33° and cultivated in a nursery located at latitude 35°, had the lowest H and biomass. In other sclerophyllous species, Bonito et al. ([5]) found the lowest seedling height for a provenance of Q. ilex whose seeds were collected in the north of Italy (latitude 45° N) and cultivated in a nursery located in the centre of the country (latitude 41° N). The authors attributed their results to the low relative growth rate of this provenance. The effect of the container size on biomass components of all of the provenances corroborates that small containers imposes physical restrictions on root growth ([2]). RDW was the lowest for the Metropolitan provenance seedlings cultivated in small containers. This provenance also exhibited the lowest biomass components, but this was especially true for small containers which had half of the volume in comparison to the large containers. Poorter et al. ([36]) pointed out that seedlings increase in weight using larger pots, i.e., the larger the container the larger the seedling that can be produced. In general, larger containers provide more water per gram of plant, and this holds especially for the Maule provenance cultivated in larger containers. In a nursery study with Q. saponaria, Espinoza et al. ([16]) reported a superior morphological and physiological performance for seedlings cultivated in large containers (280 mL). Similarly, Tsakaldimi et al. ([41]) in a nursery experiment with Q. ilex and Quercus coccifera L. found that seedlings exhibited superior diameter and root biomass when cultivated in the rigid containers of higher volume.
Survival and growth at field- test site
Our results showed that survival (90% on average) was not affected by provenance and container, though there was not enough evidence supporting the effect of the initial seedling size on the outplanting growth and survival in all provenances. Similarly, in the sclerophyllous species Q. ilex and Q. coccifera, Tsakaldimi et al. ([42]) and Del Campo et al. ([14]) found no significant correlations between field survival and seedling traits at nursery. This can be attributed to the fact that in water-limited environments (such as our study site) it is not clear which seedling attributes determines establishment success ([40]). Interestingly, seedlings from the Metropolitan provenance exhibited a high survival after outplanting, despite their low H and biomass at the nursery, and the environmental differences between their site of origin and the field test site. This provenance is likely to benefit from the wetter conditions experienced in the field test site. The deep and extensive root system of Q. saponaria ([19]) allows the species to tolerate water potentials up to -5.6 MPa in sites with Mediterranean-type climate ([31]). This could have avoided the competition for water with herbaceous plants in the top soil horizons ([21]). Padilla & Pugnaire ([33]) found that rooting depth had a strong positive effect on seedling development in arid environments. On the other hand, tree shelter protection used in the field trial may have also accounted for the high and consistent survival among provenances. Shelters are known to increase humidity, decrease excessive irradiance, and buffer extreme temperatures ([34]). Padilla et al. ([34]) and Rey-Benayas ([37]) found that shading exerted a positive effect on Q. ilex and Q. coccifera survival in arid landscapes.
At the nursery level, we observed a clear effect of the provenance on seedling growth; however, contrary to our expectations, there was no clear evidence of home site advantage for outplanting survival and increment for H in the Maule provenance. Indeed, theory predicts that local genotypes should perform better than non-local genotypes when grown at their site of origin ([45]), but there is also evidence suggesting that this is less frequent than commonly assumed ([26]). In our study the Biobío provenance, which was planted at a lower latitude as compared to the seed collection site (37° 09′ vs. 35° 34′, respectively), exhibited similar survival to the Maule provenance, whose seed was collected near the field test site (35° 14′). Ovalle et al. ([31], [32]) by using a very strict local provenancing in Q. saponaria (i.e., the field test site was located close to the seed collection site), found high levels of outplanting survival; however, these authors evaluated only one provenance and attributed their results to the nursery fertilization, watering, and control of herbivory in the field test site. It was expected that the translocation of Metropolitan provenance could result in poor adaptation; however, survival and height increment were homogeneous, despite that seedlings from the Metropolitan provenance were the shortest at the nursery. This reaffirms the contention of Broadhurst & Boshier ([7]) who indicated that “local” should not be based on administrative or geographic boundaries, but on an adaptive scale. Accordingly, our results suggest that plant material of Q. saponaria could be conveniently used for restoration purposes outside its area of origin with no negative effects on seedling survival. This could overcome the scarce availiability of seeds from local sources for the restoration of areas severely burned by fire in central Chile in 2017 ([13]). However, the possible use of non-local seed sources for other Mediterranean sclerophyllous species in highly disturbed ecosystems should be tested in advance through the implementation of common garden and reciprocal transplant experiments.
Leaf-level physiology at field test site
The photosynthetic rate of Q. saponaria seedlings in this study varied from 5.8 to 7.1 μmol CO2 m-2 s-1. Likewise, Ovalle et al. ([32]) found Asat values of 7.2 μmol CO2 m-2 s-1 for seedlings of Q. saponaria planted in a site with Mediterranean-type climate with precipitation of 300 mm year-1. They also found high increments in height after one season in the field (average height of 54.7 cm) using short seedlings at the planting date (average height of 6-8 cm). However, seedlings were cultivated in 400 mL containers and controlled-release nitrogen fertilization was applied at the planting hole. Moreover, they collected the seeds a few minutes of latitude away from the field test site. Thus, container size, fertilization, and local seed provenance should have a strong effect in their positive results.
Plasticity for physiological parameters has been reported to be greater than that for morphological traits ([43], [44], [29]). Unlike outplanting survival, our study showed that Q. saponaria exhibits differences among provenances in gas exchange parameters. This result agrees with several reports indicating that patterns of leaf-level gas exchange correlate with the climate of the seed origin, particularly with rainfall. Abrams et al. ([1]) and Seiler & Johnson ([39]) observed that Fraxinus pennsylvanica Marsh and Pinus taeda L. seedlings from xeric sources had lower rates of photosynthesis, leaf conductance, and transpiration than seedlings from mesic sources. This agrees with our study as we found lower values for Asat, E, and WUEint in Metropolitan provenance, which also had the lowest De Martonne aridity index, thus suggesting local physiological adaptations. As nitrogen plays an important role in photosynthesis, the low chlorophyll density (which reflects the nitrogen status) observed in the Metropolitan provenance cultivated in small containers could imply a low light capture capacity of this provenance, but this hypothesis needs to be tested by further research.
The superior WUEint of Biobío and Maule provenances compared to the Metropolitan provenance may be attributed to an increase of Asat rather than a reduction in gs. Both provenances showed superior levels of Asat compared with Metropolitan provenance, while stomatal conductance was similar across all provenances, even though a small decrease was only observed in the Biobío seedlings cultivated in large containers. As larger seedlings generally display a greater photosynthetic active surface in terms of leaf biomass ([27]), it seems that the better planting stock of Maule and Biobío provenances (i.e., high H and LDW in the nursery) was also reflected in a better photosynthesis in the field.
Conclusions
Testing seedlings from different provenances of Q. saponaria in a common field experiment revealed that plant material of this species could be used for restoration purposes outside its area of origin with no detrimental effects on seedling survival after outplanting. Provenance and container size had significant effects on seedling traits at the nursery stage, but these effects tend to disappear at the outplanting site. This suggests that the site conditions at the field test site (i.e., a degraded ecosystem with presence of summer drought and severe fires) are important drivers in affecting seedling performance; however, this hypothesis needs further research though reciprocal transplant experiments.
Acknowledgments
Seeds for the nursery experiment and field trial were kindly provided by the Centro de Semillas y Árboles Forestales (CESAF) from the Universidad de Chile. We are indebted with those who provided support in the field and laboratory work, especially Cristian Hernández, Gustavo Hernández, Francisco Méndez, Suraj Vaswani, Betsabé Abarca, and Nicole Toro from the Universidad de Chile, and César Gallardo and Pilar Ríos, from the Universidad Católica del Maule. SEE was supported by the Comisión Nacional de Ciencia y Tecnología (CONICYT - grant no. 79150013), AMC was supported by a postdoctoral fellowship from the Universidad Católica del Maule.
References
Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Authors’ Info
Authors’ Affiliation
Rómulo E Santelices 0000-0002-5790-8701
Universidad Católica del Maule, Facultad de Ciencias Agrarias y Forestales, Avenida San Miguel 3605, Talca (Chile)
Núcleo Científico Multidisciplinario, Universidad de Talca, 2 Norte 685, P.O. Box 747, Talca (Chile)
Universidad de Chile, Facultad de Ciencias Forestales y de la Conservación de la Naturaleza, Avenida Santa Rosa 11365, La Pintana (Chile)
Universidad Católica del Maule, Vicerrectoría de Investigación y Posgrado, Avenida San Miguel 3605, Talca (Chile)
Corresponding author
Paper Info
Citation
Espinoza SE, Yañez MA, Magni CR, Santelices RE, Cabrera AM (2020). Outplanting performance of three provenances of Quillaja saponaria Mol. established in a Mediterranean drought-prone site and grown in different container size. iForest 13: 33-40. - doi: 10.3832/ifor3142-012
Academic Editor
Gianfranco Minotta
Paper history
Received: May 03, 2019
Accepted: Nov 05, 2019
First online: Jan 21, 2020
Publication Date: Feb 29, 2020
Publication Time: 2.57 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2020
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 44853
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 37021
Abstract Page Views: 3787
PDF Downloads: 3157
Citation/Reference Downloads: 4
XML Downloads: 884
Web Metrics
Days since publication: 2364
Overall contacts: 44853
Avg. contacts per week: 132.81
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2020): 5
Average cites per year: 0.83
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Review Papers
The role of spatial criteria in enhancing forest restoration actions: a systematic review
vol. 19, pp. 157-167 (online: 11 May 2026)
Research Articles
Effect of restoration methods on natural regeneration in the Brazilian Atlantic Forest
vol. 18, pp. 23-29 (online: 15 February 2025)
Research Articles
Can the dynamics of forest restoration reduce landscape fragmentation in the Atlantic forest?
vol. 18, pp. 61-68 (online: 04 April 2025)
Research Articles
The effect of seed size on seed fate in a subtropical forest, southwest of China
vol. 9, pp. 652-657 (online: 04 April 2016)
Research Articles
Tropical seedling performance under drought: a functional trait approach for species selection in restoration
vol. 19, pp. 9-17 (online: 10 January 2026)
Research Articles
Nutrient cycling indicators in different models of successional groups and spacing for forest restoration
vol. 19, pp. 61-68 (online: 23 February 2026)
Review Papers
Shaping the multifunctional tree: the use of Salicaceae in environmental restoration
vol. 6, pp. 37-47 (online: 21 January 2013)
Short Communications
Differential adaptations in nursery seedlings from diverse Chilean provenances of Peumus boldus Mol.
vol. 9, pp. 409-413 (online: 29 January 2016)
Research Articles
Seed trait and rodent species determine seed dispersal and predation: evidences from semi-natural enclosures
vol. 8, pp. 207-213 (online: 28 August 2014)
Research Articles
Post-fire recovery of the plant community in Pinus brutia forests: active vs. indirect restoration techniques after salvage logging
vol. 11, pp. 635-642 (online: 04 October 2018)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







