
Respiratory costs of woody tissues in a Quercus pyrenaica coppice
iForest - Biogeosciences and Forestry, Volume 11, Issue 3, Pages 437-441 (2018)
doi: https://doi.org/10.3832/ifor2599-011
Published: Jun 18, 2018 - Copyright © 2018 SISEF
Short Communications
Abstract
Long-term coppicing leads to the development of massive root systems. A disproportionate carbon investment in root maintenance has been pointed as a cause of the widespread decline of abandoned coppices. We aimed at assessing how coppicing has influenced root and shoot development and related carbon loss ascribed to maintenance of woody tissues in Quercus pyrenaica. For this goal, results from published studies on root dynamics, woody biomass and respired CO2 fluxes in an abandoned Q. pyrenaica coppice were integrated and extended to quantify overall respiratory expenditures of above- and below-ground woody organs. Internal and external CO2 fluxes together with soil CO2 efflux were monitored in eight stems from one clone across a growing season. Stems and roots were later harvested to quantify the functional biomass and scale up root and stem respiration (RR and RS, respectively) to the clone and stand levels. Below- and above-ground biomass was roughly equal. However, the root-to-shoot ratio of respiration (RR/RS) was generally below one. Relatively higher RS suggests enhanced metabolic activity aboveground during the growing season, and highlights an unexpected but substantial contribution of RS to respiratory carbon losses. Moreover, soil and stem CO2 efflux to the atmosphere in Q. pyrenaica fell in the upper range of reported rates for various forest stands distributed worldwide. We conclude that both RS and RR represent an important carbon sink in this Q. pyrenaica abandoned coppice. Comparatively high energetic costs in maintaining multiple stems per tree and centennial root systems might constrain aboveground performance and contribute to coppice stagnation.
Keywords
Carbon Loss, CO2 Fluxes, Coppice Stagnation, Oak, Resprouting Species, Root Respiration, Stem Respiration
Introduction
Quercus pyrenaica Willd. is a vigorous root-resprouting species that has been intensively coppiced for firewood, charcoal and woody pastures for centuries ([20]). Due to the appearance of new energy sources and rural exodus that occurred in the 1970s, coppicing has mostly ceased, and symptoms of decline - slow stem growth, branch dieback, and scarce acorn production - are widely observed in abandoned stands ([26]). Coppice stagnation entails ecologic, economic and social problems, namely increased fire risk, stand over-aging, low productivity, absence of sexual regeneration, and consequently, hindered application of alternative management plans. Due to the wide distribution and significant ecological value of coppiced stands of Mediterranean oak species, silviculture faces the crucial challenge of finding new alternative uses for these abandoned coppices ([6], [4]). Attempts to conversion into high forests via thinning have not been successful to date, partly due to the lack of a comprehensive understanding of the physiological mechanisms underpinning tree stagnation ([25]).
Disproportionate respiratory costs of large root systems grown after centennial coppicing have been suggested as a potential driver of Q. pyrenaica decay ([4]), but assessments on carbon expenditures have not been essayed to date. Quantification of the relative weight of respiratory carbon sinks for the plant is crucial for a better understanding of tree carbon budgets ([32], [1], [17]). Notwithstanding, our comprehension of respiratory processes, particularly of woody organs, is limited compared to our knowledge of photosynthesis ([10], [17], [13]). In resprouting deciduous species, nonstructural carbohydrates are stored in large amounts in woody organs ([3]) that can contain a large portion of living parenchyma ([19]). The penalty in terms of respiratory carbon loss associated to the maintenance of these storage tissues ([14], [9]) could be of particular relevance in carbon budgets of root-resprouting Q. pyrenaica. Long lasting coppicing might lead to massive systems of living roots ([21], [31]) that store but also consume a large portion of carbohydrates assimilated aboveground.
To better understand the role of respiratory carbon loss in Q. pyrenaica decay, we gathered and extended previous published work on Q. pyrenaica biomass and respiration of woody tissues ([22], [21], [23], [24]) to scale up stem and root respiration (RS and RR, respectively) to the tree and stand levels. We aimed at comparing respiratory expenditures (i) between above- and below-ground woody organs across one growing season, and (ii) in relation to data gathered from various forests to provide an insight of the magnitude of carbon invested for woody tissue respiration in Q. pyrenaica coppices. We expect RS and particularly RR to be important carbon sinks for the plant (i.e., from the plant perspective), and therefore high RR/RS ratios as well as woody tissue respiration rates relative to other forest stands.
Materials and methods
To estimate respiratory carbon loss of woody organs in a coppice system of Q. pyrenaica, we reviewed our previous work on Q. pyrenaica root development and biomass ([21]), and internal and external stem CO2 fluxes ([22], [23], [24]), together with unpublished data of soil CO2 efflux. These studies were performed in a one-hectare experimental plot located in the Monte Matas de Valsaín (Segovia, Spain) at an altitude of 1140 m a.s.l. Climate is sub-Mediterranean with an average annual rainfall and temperature of 885 mm and 10 °C, respectively. It consists on a monospecific one-storied regular coppice of Q. pyrenaica with a stand density of 781 stems ha-1. The forest has been subjected to coppicing since at least the XII century, and traditional management was abandoned in the 1970s. Stems within the plot were geo-referenced and leaves collected for genetic analyses to delineate the commonly inconspicuous clonal structure of Q. pyrenaica coppiced stands. Note that Q. pyrenaica is a root resprouting species, hence one clone can be constituted by several stems located far away (dozens of meters) from each other ([30]). The clonal assignment of stems was based on nuclear microsatellite molecular markers ([29] - see Fig. 1 in [22]). Data on woody tissue respiration was collected from the eight stems belonging to a single clonal genotype (clone). Four 24-h measurement campaigns were conducted across the growing season of 2013, at the end of which stems and roots were harvested for biomass quantification and scaling up of RS and RR to the clone and stand levels. The root system was hydraulically excavated with a high-pressure water pump down to 1 m depth over an area of 81 m2 (Fig. 1). Biomass was partitioned into leaves, branches, stems, taproots, coarse roots and fine roots. Woody biomass was further partitioned into bark, sapwood and heartwood tissues from allometric equations adjusted by means of exhaustive sampling of branches, stems, taproots and coarse roots. Leaf area index (LAI) was estimated from measurements of specific leaf area of sampled leaves and total leaf biomass. Further details on stand characteristics, excavation methodology, and above- and below- ground biomass measurements can be seen in Salomón et al. ([21]).
Fig. 1 - Root systems of two hydraulically excavated clones of Quercus pyrenaica located in the Monte Matas de Valsaín (Segovia, Spain). The root system of the large clone surveyed in this study (lower part of the photograph) covered at least 81 m2. Before harvesting, the clone was composed by eight stems connected through root grafts and parental roots.

Stem CO2 efflux to the atmosphere (EA) was measured in every stem with a portable infrared gas analyzer (LI-6400®, Li-Cor Inc., Lincoln, NE, USA) and a soil chamber (LI-6400-09®) using PVC collars attached to the stems. Stem EA measured on a surface area basis (EA(S)) was expressed on a volume basis (EA(V) - eqn. 1):
where S and V are the axial surface area and the volume of living tissues (bark and sapwood) of the stem segment, respectively; rh and rh+l denote the radius of heartwood and heartwood plus living tissues, respectively, and L is the vertical length of the stem segment ([24]). Stem respiration (RS) was estimated as the sum of EA(V) and the internal CO2 flux through xylem (FT) as RS = EA + FT (adapted from [16]). FT was calculated as a function of the sap flux and the vertical gradient of CO2 dissolved in sap solution (sap [CO2*]). Sap flux density was measured using Granier-type thermal dissipation probes, and sap [CO2*] was estimated from measurements of xylem [CO2] in the gas phase, sap temperature, and sap pH in each stem applying Henry’s law. Briefly, xylem [CO2] was measured with solid non-dispersive infrared (NDIR) CO2 sensors (model GMM221®, Vaisala, Helsinki, Finland) inserted into the stem above and below the stem collar. Stem temperature was measured with type-T thermocouples inserted 5 cm away from the NDIR probe. Sap pH was measured with a micro-pH electrode and a portable pH meter (Crison, Barcelona, Spain) on sap samples expressed from detached twigs using a pressure chamber (see [23] for further details). Overall EA, FT and RS at the clone level were estimated by aggregating EA, FT and RS scaled up for each stem (and their branches) according to their corresponding volume of living woody biomass. Aboveground clonal respiratory fluxes were eventually expressed on a soil surface area basis taking into account the clonal surface extension.
Soil CO2 efflux (ES) was measured with a portable infrared gas analyzer and a soil chamber using soil PVC collars (see [22] for detailed methodology). Unpublished data from four soil collars located below the canopy of the eight monitored stems were averaged to obtain clonal ES on a soil area basis (m2). Since roots barely extended beyond the excavated area (81 m2), a buffer of 0.63 m was added to estimate the clone extension (102 m2). This buffer distance was chosen to meet actual stand density (8 stems in 102 m2 = 784 stems ha-1) and scale up results to the stand level. Root-respired CO2 diffusing to the atmosphere through soil (ES-ROOT) was estimated from ES measurements and the relative contribution of autotrophic respiration to ES. Seasonality of root autotrophic contribution to ES was obtained from two studies in a Mediterranean Quercus cerris coppice cut one ([18]) and 17 ([27]) years before ES measurements. Spring contributions reported in these studies were attributed to the first measurement campaign (DOY 143-144), summer contributions to the second (DOY 183-184) and third (DOY 218-219) campaigns, and autumn contributions to the fourth campaign (DOY 266-267). To account for FT on RR estimates (RR = ES-ROOT + FT - [2]), internal and external CO2 fluxes were measured at the base of the stem (0.1 m).
To compare ES, RR and RS averaged over the growing season within the monitored clone, ANOVA and pairwise comparisons were performed in R software (version 3.4.0). Respiratory fluxes at the stand level from 15 sites were gathered (Tab. S1 in Supplementary material) to evaluate the relative magnitude of respiratory costs of the surveyed Q. pyrenaica coppice. Inter-site statistical comparisons were not performed because only one site was surveyed in this study.
Results and discussion
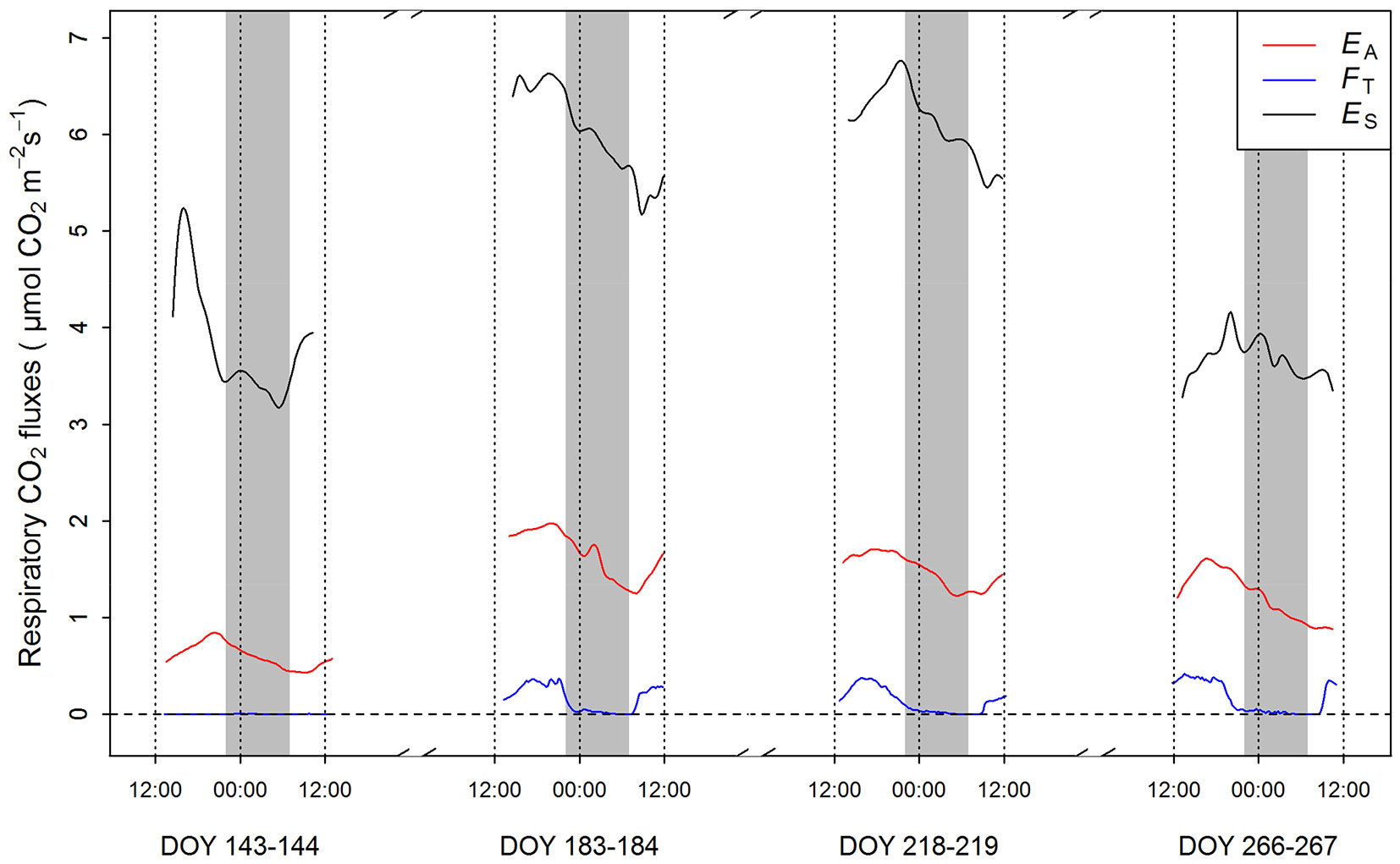
Seasonal and diel variation in EA, FT and ES on a soil area basis are shown in Fig. 2. The contribution of FT to aboveground RS was less than 10% ([24]), whereas FT belowground was less than 2% of ES-ROOT ([22]). The modest contribution of axial CO2 transport to total respiration rates is explained by the low xylem [CO2] observed in Q. pyrenaica, generally lower than 1%. This concentration is about one order of magnitude lower than that reported for other tree species using this methodology ([28]). The limited build-up of CO2 in the xylem was partly ascribed to the low resistance to radial CO2 diffusion, likely related to the poor plant water status of species distributed across drought-prone regions ([23]). Averaged over the growing season, ES (38.9 mol CO2 clone-1 day-1) was greater than RS (11.7 mol CO2 clone-1 day-1) and RR (8.5 mol CO2 clone-1 day-1 - P < 0.001; Tab. 1, Fig. 2), being RS and RR not significantly different (P > 0.10). Due to the magnitude of ES, the contribution of autotrophic respiration to ES substantially determined the root-to-shoot ratio of respiration (RR/RS). To illustrate this, a contribution of RR to ES ranging between 14 and 27% ([18], [27] - data reported for a Mediterranean oak coppice at different states of maturity) yielded RR/RS ratios ranging between 0.42 and 1.18 across the growing season (Tab. 1). Alternatively, if heterotrophic and autotrophic contributions to ES were considered equal, as generally assumed for different forest biomes ([11], [2]), RR/RS would reach values ranging from 1.36 to 2.18.
Fig. 2 - Diel variations in stem CO2 efflux to the atmosphere (EA), stem internal CO2 transport through xylem (FT) and soil CO2 efflux (ES) on four dates over the growing season in an abandoned coppice of Quercus pyrenaica. Fluxes registered from one stem segment and one soil collar intensively monitored (18 times day-1) are shown. Additional stem segments and collars used to average EA, FT and ES are not displayed in this figure as they were monitored less intensively (4 times day-1). EA and FT on a volume basis was obtained from previous work ([24]) and expressed on a soil area basis for comparison with ES. Shaded areas indicate night-time.
Tab. 1 - Above- and below-ground respiratory fluxes in an eight-stemmed clone of Quercus pyrenaica. Stem CO2 efflux to the atmosphere (EA), soil CO2 efflux (ES), and stem and root respiration (RS and RR, respectively) were measured during four 24-h campaigns throughout 2013 growing season. (a): RS was estimated as the sum of EA and the internal CO2 flux through xylem (FT). (b): RR was estimated from ES measurements and the contribution of autotrophic respiration to ES. Autotrophic contribution to ES was obtained from two reports of a Mediterranean coppice of Quercus cerris cut one ([18]) and 17 years ([27]) before measurements. Estimations of RR and RR/RS ratios from contributions reported in both studies (recently coppiced vs mature stand) are shown in left and right sub-columns, respectively. FT was neglected in RR due to its low contribution (< 2%).
| Campaign DOY | E A | RS (a) | E S | RR (b) | RR/RS |
|---|---|---|---|---|---|
| (mol CO2 clone-1 day-1) | |||||
| 143-144 | 6.16 | 6.16 | 26.83 | 5.03 vs 7.25 | 0.82 vs 1.18 |
| 183-184 | 12.28 | 13.13 | 52.14 | 11.32 vs 11.70 | 0.86 vs 0.89 |
| 218-219 | 15.63 | 16.45 | 43.72 | 9.49 vs 9.82 | 0.58 vs 0.60 |
| 266-267 | 10.54 | 11.23 | 32.76 | 8.82 vs 4.68 | 0.79 vs 0.42 |
| Mean | 11.15 | 11.74 | 38.86 | 8.66 vs. 8.36 | 0.76 vs. 0.77 |
Above- and below-ground functional woody biomass (bark and sapwood) was similar in the surveyed clone: 1026 Kg aboveground and 972 Kg belowground ([21]). Consequently, seasonal deviations of RR/RS from unity reflected differences in the metabolic activity between below- and above-ground organs over time. RR/RS ratio above one was uniquely observed during spring (Tab. 1), likely explained by an earlier growth of roots relative to stems ([15]), particularly by intense fine root growth and belowground cambial activity at this time of the year ([8]). The decrease in RR/RS observed onward (ratios below one) resulted from the moderate root activity relative to the intensification of aboveground metabolism, namely stem growth, leaf development and phloem transport. Predominant RR/RS ratios below one along the growing season evidenced an unexpected large weight of aboveground woody tissue respiration as a carbon sink for the plant. The accumulation of woody biomass in stems and branches in this over-aged coppice (cut for the last time around 1967), together with the remarkably high portion of living parenchyma observed in Q. pyrenaica stems ([19]) may contribute to the high respiratory costs of woody organs aboveground.
Respiratory fluxes at the stand level, and extrapolated to the whole year, were compared with those reported for several forest sites (see Tab. s1 in Supplementary material for details on the extrapolation). Average ES and EA across 15 stands were 776 and 162 g C m-2 year-1, respectively, whereas these values increased up to 1164.2 and 297.0 g C m-2 year-1 in the surveyed Q. pyrenaica coppice (Tab. S1). That is, ES and EA were 1.5 and 1.8 times higher in this study, suggesting greater carbon losses from soil and stem respiration. These ratios could be substantially reduced, however, if xylem transport of respired CO2 was accounted for ([2]) in the 15 forest stands. Leaf area index, a key ecophysiological determinant of carbon gas exchange at the canopy ([5]), was additionally considered for comparison. LAI in our site was 3.8, a value that falls within the reported range for the species ([12]) and the genus Quercus ([5]). The slightly low LAI relative to that averaged across different forest types (4.2 - Tab. S1 in Supplementary material) does not suggest a greater potential for carbon assimilation. Taken together, these rough comparisons suggest a strong carbon sink for the plant associated to respiratory processes that do not scale with the carbon input within the surveyed Q. pyrenaica coppice.
To summarize, the relative importance of RR and RS as carbon sinks for the plant shifted along the growing season in accordance to the root and stem phenology and metabolic activity. RR/RS ratios lower than one point to an unexpected importance of aboveground woody tissue respiration in carbon budgets of Q. pyrenaica coppices. Large carbon losses via soil and stem respiration relative to those observed in several forest types supports the hypothesis of an imbalance between carbon sources and sinks, contributing to the decline of abandoned coppices ([7], [22], [21]). Nonetheless, these conclusions should be handled with caution due to the lack of data on carbon assimilation, and the limited sample size of the study, largely constrained by the laborious task of root excavation. Firstly, temporal patterns of leaf carbon exchange should be analyzed in detail to gain a comprehensive perspective of tree carbon budgets and to better address the role of a potential root-to-shoot physiological imbalance in Q. pyrenaica coppice decline. Secondly, a greater number of monitored individuals as well as soil collars within the plot and across longer periods would improve spatial and temporal upscaling of respiratory fluxes. In this line, comparisons between coppiced and non-coppiced sites would provide stronger empirical support to the hypothesis that historical coppicing leads to massive root development and constrained aboveground performance.
List of abbreviations
EA, stem CO2 efflux to the atmosphere; EA-LEAF, foliage CO2 efflux to the atmosphere; ES, soil CO2 efflux; ES-ROOT, root-respired CO2 that diffused to the atmosphere through soil; FT, internal CO2 flux through xylem; RR, root respiration; RS, stem respiration
Acknowledgements
We are grateful to Javier Donés (Centro de Montes y Aserradero de Valsaín) for economic and logistical support. We also thank Elena Zafra, Guillermo González, César Otero, Manuel Iglesias, Paula Guzmán, Aida Rodríguez, Jose Carlos Miranda, Rosa Ana López, Eva Miranda and Matías Millerón for their enthusiastic help with field work. This work was funded by the Comunidad de Madrid through CAM P2009/AMB-1668 and S2013/MAE-2760 projects and by the Organismo Autónomo de Parques Nacionales through the PPNN 1148/2014 project. Roberto L. Salomón was supported by a Ph.D. scholarship from the Universidad Politécnica de Madrid.
References
Gscholar
CrossRef | Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Jesús Rodríguez-Calcerrada
Luis Gil
María Valbuena-Carabaña
Forest Genetics and Ecophysiology Research Group, E.T.S. Forestry Engineering, Technical University of Madrid, Ciudad Universitaria s/n, 28040, Madrid (Spain)
Laboratory of Plant Ecology, Department of Applied Ecology and Environmental Biology, Faculty of Bioscience Engineering, Ghent University, Coupure links 653-9000 Ghent (Belgium)
Corresponding author
Paper Info
Citation
Salomón RL, Rodríguez-Calcerrada J, Gil L, Valbuena-Carabaña M (2018). Respiratory costs of woody tissues in a Quercus pyrenaica coppice. iForest 11: 437-441. - doi: 10.3832/ifor2599-011
Academic Editor
Giorgio Matteucci
Paper history
Received: Aug 11, 2017
Accepted: Apr 09, 2018
First online: Jun 18, 2018
Publication Date: Jun 30, 2018
Publication Time: 2.33 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2018
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 46349
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 39345
Abstract Page Views: 2949
PDF Downloads: 3054
Citation/Reference Downloads: 8
XML Downloads: 993
Web Metrics
Days since publication: 2929
Overall contacts: 46349
Avg. contacts per week: 110.77
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2018): 3
Average cites per year: 0.38
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Spatial heterogeneity of soil respiration in a seasonal rainforest with complex terrain
vol. 6, pp. 65-72 (online: 07 February 2013)
Review Papers
Separating soil respiration components with stable isotopes: natural abundance and labelling approaches
vol. 3, pp. 92-94 (online: 15 July 2010)
Research Articles
Soil respiration along an altitudinal gradient in a subalpine secondary forest in China
vol. 8, pp. 526-532 (online: 01 December 2014)
Research Articles
Soil respiration and carbon balance in a Moso bamboo (Phyllostachys heterocycla (Carr.) Mitford cv. Pubescens) forest in subtropical China
vol. 8, pp. 606-614 (online: 02 February 2015)
Research Articles
Thinning effects on soil and microbial respiration in a coppice-originated Carpinus betulus L. stand in Turkey
vol. 9, pp. 783-790 (online: 29 May 2016)
Research Articles
Short-time effect of harvesting methods on soil respiration dynamics in a beech forest in southern Mediterranean Italy
vol. 10, pp. 645-651 (online: 20 June 2017)
Research Articles
Effect of different dolomitic limestone dosages on soil respiration in a mid-altitudinal Norway spruce stand
vol. 12, pp. 357-365 (online: 05 July 2019)
Research Articles
Seasonal dynamics of soil respiration and nitrification in three subtropical plantations in southern China
vol. 9, pp. 813-821 (online: 29 May 2016)
Research Articles
Comparison of soil CO2 emissions between short-rotation coppice poplar stands and arable lands
vol. 11, pp. 199-205 (online: 01 March 2018)
Research Articles
Day and night respiration of three tree species in a temperate forest of northeastern China
vol. 8, pp. 25-32 (online: 26 May 2014)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







