
The complexity of mycobiota associated with chestnut galls induced by Dryocosmus kuriphilus in Galicia (Northwestern Spain)
iForest - Biogeosciences and Forestry, Volume 17, Issue 6, Pages 378-385 (2024)
doi: https://doi.org/10.3832/ifor4559-017
Published: Dec 14, 2024 - Copyright © 2024 SISEF
Research Articles
Abstract
The European chestnut tree (Castanea sativa) is a highly valued deciduous species in Galicia (Northwestern Spain), mainly due to economic, landscape, and social or cultural reasons. However, the Asian wasp Dryocosmus kuriphilus, one of the biggest threats to C. sativa, is severely affecting tree vigor and reducing chestnut yields. Some studies indicated that this wasp and the galls that it produces may play an important role in spreading fungal disease. The present work aimed to characterize the complex of fungi associated with galls induced by D. kuriphilus in Galician chestnut trees, focusing on the study of plant-pathogen diversity. For this purpose, branches with necrotic galls were collected from seventy-eight chestnut stands located in the four Galician provinces (A Coruña, Lugo, Ourense, and Pontevedra). In total, 1170 necrotic galls of the year of study were collected at the different sampling sites. In the laboratory, four galls were randomly selected from each point, thus analyzing 312 galls, which were surface disinfected and dissected, and tissue fragments placed on culture media. From the selected galls, 308 fungal colonies were isolated and then identified by the morphology of their fruiting bodies and the molecular analysis of the ITS, beta-tubulin, elongation factor, and histone regions. They were classified into 27 genera and 53 species, of which 5 genera and 29 species of fungi were reported for the first time associated with D. kuriphilus galls, including phytopathogenic fungi. Results show that further research is needed to study in detail the role of galls as entry points and reservoirs of pathogenic fungi.
Keywords
Asian Chestnut Gall Wasp, Castanea sativa, Endophytes, Phytopathogens
Introduction
The European chestnut tree (Castanea sativa Mill.) registered under the Protected Geographical Indication (PGI) regulation “Castaña de Galicia” is a highly valued deciduous species in Galicia (Northwestern Spain), mainly due to economic, landscape, and social or cultural reasons. Chestnut trees are of great economic importance in this area due to their nutritional value, high fruit yield, and excellent wood quality. Spain is the second largest producer in the world, where approximately 60% of the European nut production originates, and 66% of the Spanish stands are located in Galicia, representing 92% of the country’s total nut production ([20]).
The Asian chestnut gall wasp Dryocosmus kuriphilus Yasumatsu (Hymenoptera, Cynipidae) is one of the most important pests of C. sativa. It prevents fruit production by altering the growth and physiology of the host plant, interrupting photosynthetic organ development, and altering flower growth ([18]). Consequently, it affects tree growth and reproduction, resulting in decay, fewer fruits and reduced crop yields, and, in severe cases, tree death ([19]). According to various studies, chestnut crop yields can be reduced by 50%-80% depending on the variety and the biological control measures used ([18], [19]).
This insect is listed as a harmful organism in Annex III (List of protected zones and the respective protected zone quarantine pests and their respective codes) of the Commission Implementing Regulation (EU) 2019/2072 and has been included in the A2 list of the European and Mediterranean Plant Protection Organization (EPPO) since 2003 ([19]).
The wasp is native to China, although it is currently present in Southeast Asia, most of Europe and the east coast of the United States ([19]). In Europe, this wasp was first recorded in Italy in 2002, spreading and invading the rest of the continent. In Spain, it was detected for the first time in 2012 in Catalonia and, two years later, in Galicia and northern Portugal ([41]). According to data collected in annual samplings carried out by the Regional Government of Galicia (Xunta de Galicia), this pest is now widespread in Galicia and its incidence has greatly increased. This wasp can spread locally as an adult since it is capable of flying. Additionally, it can be carried by the wind, transported by humans, or travel long distances due to the movement of infested plant material. The wasps may be inconspicuous within the plant tissues, making them difficult to detect visually ([18]).
D. kuriphilus is a univoltine cynipid (produces one generation per year) and reproduces by thelytokous parthenogenesis; that is, only females develop from unfertilized eggs, thus no males of the species are known ([19]). During their life cycle, D. kuriphilus larvae feed for 20-30 days before pupating, which induces green to reddish gall formation on buds, leaves, and male catkins ([41]). Depending on the climatic conditions of the area and the chestnut cultivar, pupation takes place between mid-May and July. Adult females emerge from the galls between late May and July and can infect new shoots immediately ([41]). After emergence, galls dry, become wood-like, and remain on the tree for several years ([19]).
According to some studies, the galls produced by this wasp can act as entry points and sources of inoculum, thereby causing an increase in the incidence of chestnut diseases, such as chestnut blight, caused by Cryphonectria parasitica (Murrill) M.E. Barr ([35]), or brown rot, produced by Gnomoniopsis smithogilvyi L.A. Shuttleworth, E.C.Y. Liew & D.I. Guest (syn. G. castaneae Tamietti - [34]). On the other hand, several studies suggest that some endophytic species present in galls could be used as biological control agents against D. kuriphilus and other pests ([1], [49]).
This work aimed to characterize, for the first time in Galicia, the complex of fungi associated with galls induced by D. kuriphilus in chestnut trees, focusing on the study of plant-pathogen diversity.
Material and methods
Study area and sampling sites
Seventy-eight chestnut stands randomly selected in the four Galician provinces (A Coruña, Lugo, Ourense, and Pontevedra) were examined during dormancy. Of these stands, 70% belong to the area covered by the PGI Castaña de Galicia (Fig. 1).
Fig. 1 - The location of Galicia in Northwestern Spain and the geographical distribution of chestnut sampling points in the region.
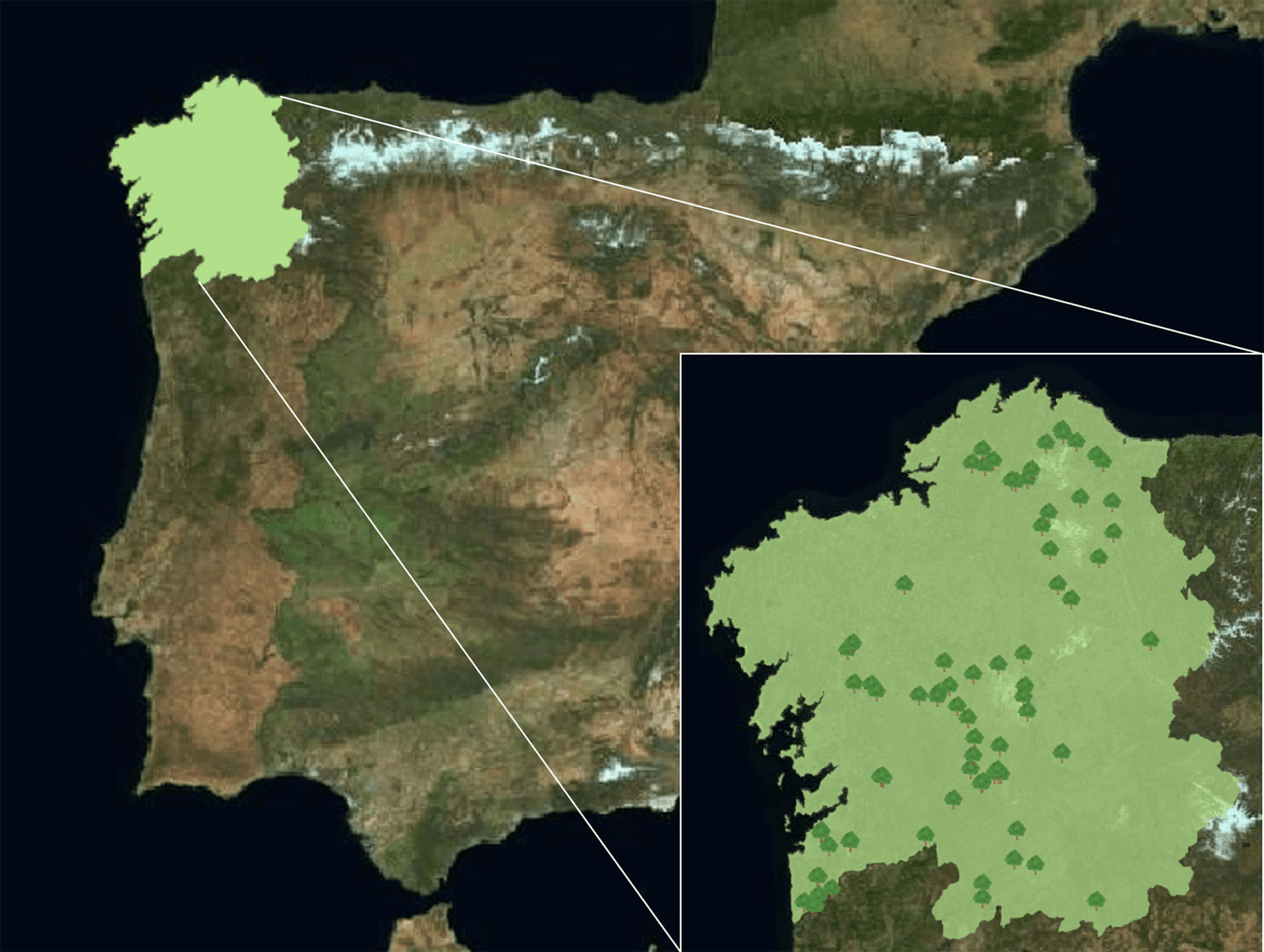
In each site, three specimens of C. sativa showing symptoms caused by D. kuriphilus were selected. Then, several branches with necrotic galls were collected (Fig. 2), totaling five galls per tree, thus obtaining 15 galls per site. In total, 1170 necrotic galls of the year of study were collected at the different sampling sites (120 from A Coruña, 375 from Lugo, 255 from Ourense, and 420 from Pontevedra). Samples were then coded and stored cold until analysis.
Fig. 2 - Necrotic galls produced by D. kuriphilus on chestnut branches.

Isolation and morphological characterization
In the laboratory, four galls randomly selected from each site were surface sterilized with 1% sodium hypochlorite for one minute and rinsed twice with sterile water for at least five minutes. Subsequently, they were dried with blotting paper, labeled, and cut into small fragments (5 × 5 mm). Around 5-10 fragments were placed in 90 mm Petri dishes containing the culture media Malt Extract Agar with streptomycin 0.5 g L-1 (AMs) and kept in a laminar flow chamber under sterile conditions. Plates were sealed with Parafilm® and incubated in the dark at 24 °C. Fragments were checked daily under the microscope, and after 4-5 days, a portion of each mycelium grown from the seeded fragments was transferred to a new plate with Potato Dextrose Agar (PDA) to obtain pure cultures.
Fungal isolates were first identified according to their morphology at the genus level and then by molecular techniques at the species level.
For the morphological study of each isolate, macroscopic features of the mycelium, such as colony shape, color, and texture, and microscopic features (size and shape of the mycelium and size, shape, and color of the conidia) were recorded for each isolate using a Nikon Eclipse® E600 microscope and compared with different fungal taxonomic keys.
Molecular identification
Molecular analysis was performed for all morphologically identified isolates by amplification, sequencing, and phylogeny of different fungal regions. Genomic DNA was extracted from 7-day-old pure colonies grown in PDA culture medium using the commercial E.Z.N.A. Fungal DNA Mini Kit (Omega Bio-tek, Norcross, GA, USA) following the short protocol. According to the genus determined by morphological analysis, different molecular markers were amplified and sequenced for each isolate: ITS, beta-tubulin, elongation factor 1-alpha, and histone (Tab. 1).
Tab. 1 - Primers from the ITS, beta-tubulin, elongation factor and histone regions used in this study.
| Region | Primer name | Reference |
|---|---|---|
| ITS | ITS1F | Gardes & Bruns ([22]) |
| ITS4 | White et al. ([50]) | |
| Bot2R | García-Figueres et al. ([21]) | |
| Beta-tubulin | T1 | O’Donnell & Cigelnik ([38]) |
| BT2B | Glass & Donaldson ([24]) | |
| Elongation factor 1-alpha |
EF1 | Geiser et al. ([23]) |
| EF2 | Geiser et al. ([23]) | |
| EF1-728F | Carbone & Kohn ([12]) | |
| Histone | H3-1b | Glass & Donaldson ([24]) |
| CYLH3F | Crous et al. ([15]) |
Selected regions were amplified in a SureCycler® 8800 thermal cycler (Agilent Technologies, Santa Clara, CA, USA). PCR products were separated on a 2% (w/v) electrophoresis gel in 0.5× TBE, stained with Midori Green® (Nippon Genetics, Europe), and examined under ultraviolet light. PCR products were purified with the Ilusta ExoProStar® 1-Step kit (GE Healthcare Life Sciences, Chicago, IL, USA). Amplicons obtained were sequenced in both directions (forward and reverse) using the two primers with the Big Dye Terminator V3.1 Cycle Sequencing Kit (Applied Biosystems, Waltham, MA, USA) on the ABIPrism 3500 Genetic Analyzer.
Bioinformatic analysis
The consensus sequence was obtained with the Mega v. 7 software (⇒ https://www.megasoftware.net/) and compared with the homologous sequences using the BLASTn search application of the NCBI GenBank nucleotide database (⇒ https://blast.ncbi.nlm.nih.gov/). Subsequently, a mono- or multi-phylogenetic analysis was performed for each isolate of the more complex genera Fusarium and Trichoderma analyzed, according to the loci used in the molecular identification process.
The sequences of one representative isolate per species were deposited in GenBank, and the accession numbers are specified in Tab. S1 (Supplementary material).
Results
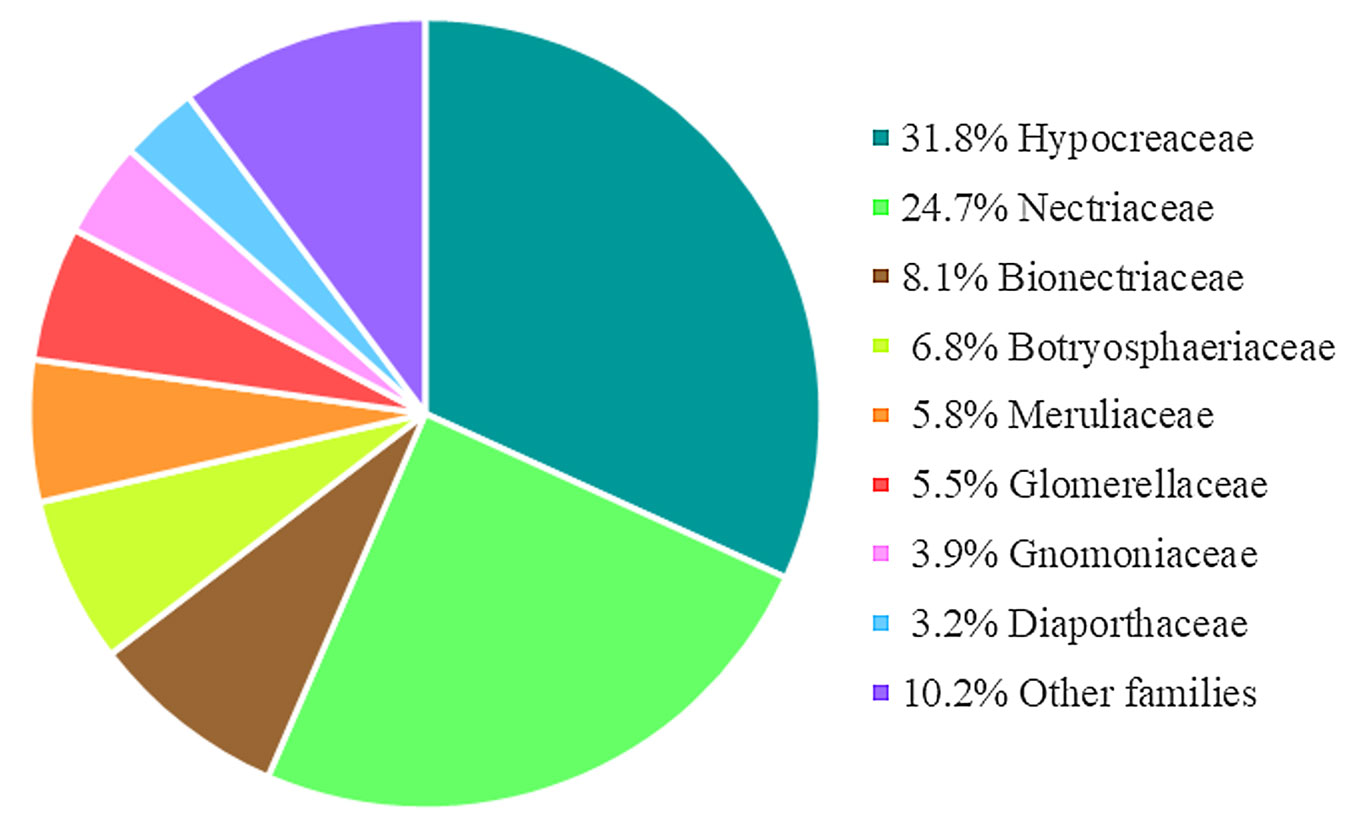
From the analysis of 312 D. kuriphilus galls we identified 308 fungal colonies. Of the isolates, 91.6% belong to the Ascomycota phylum, being the Hypocreaceae (31.8%) and Nectriaceae (24.7%) the most abundant families (Fig. 3). At the morphological level, the isolates were classified into 27 different genera (Tab. S1 in Supplementary material), of which five were reported for the first time: Abortiporus Murrill, Neurospora Shear & B.O. Dodge, Phlebia Fr., Rosellinia De Not., & D. Hawksw. and Schizophyllum Fr. The most abundant genera were Fusarium Link with 76 isolates (24.7%) and Trichoderma Persoon ex Gray, having 98 isolates (31.8%) (Tab. S1 in Supplementary material).
Fig. 3 - Relative abundance of the isolated fungi at the family level.
The morphological results combined with the molecular analysis of the ITS, beta-tubulin, elongation factor, and histone regions allowed the identification of 53 different species (Tab. S1 in Supplementary material), thus showing the great diversity of fungi in the analyzed galls. In fact, no references of presence in galls have been found for 29 species, accounting for 55% of the species detected (Tab. 2). A conclusive morphological and molecular identification at the species level was not obtained for 28 isolates, which is approximately 10% of all isolates. These unidentified isolates belong to 11 different genera, which can be consulted in Tab. S1 (Supplementary material). The presence of the genus Gnomoniopsis is highlighted since, in recent years, new species such as G. daii, G. castanopsidis, G. fagacearum, G. guangdongensis, G. hainanensis, G. rossmaniae and G. silvicola ([33]) have been described. The 5 isolates that could not be identified may belong to a new Gnomoniopsis species not yet described, as this genus is currently under study.
Tab. 2 - Taxonomic classification, number of isolates and relative abundance of the species reported for the first time associated with D. kuriphilus galls.
| Species | Phylum | Family | No. isolates |
Relative abundance (%) |
|---|---|---|---|---|
| Abortiporus biennis | Basidiomycota | Podoscyphaceae | 1 | 0.32 |
| Colletotrichum fioriniae | Ascomycota | Glomerellaceae | 17 | 5.52 |
| Diaporthe eres | Ascomycota | Diaporthaceae | 4 | 1.30 |
| Diaporthe phaseolorum | Ascomycota | Diaporthaceae | 1 | 0.32 |
| Diaporthe phillipsii | Ascomycota | Diaporthaceae | 1 | 0.32 |
| Diplodia corticola | Ascomycota | Botryosphaeriaceae | 2 | 0.65 |
| Diplodia mutila | Ascomycota | Botryosphaeriaceae | 3 | 0.97 |
| Diplodia sapinea | Ascomycota | Botryosphaeriaceae | 5 | 1.62 |
| Dothiorella iberica | Ascomycota | Botryosphaeriaceae | 1 | 0.32 |
| Fusarium acuminatum | Ascomycota | Nectriaceae | 2 | 0.65 |
| Fusarium culmorum | Ascomycota | Nectriaceae | 2 | 0.65 |
| Fusarium foetens | Ascomycota | Nectriaceae | 3 | 0.97 |
| Fusarium graminearum | Ascomycota | Nectriaceae | 7 | 2.27 |
| Fusarium solani | Ascomycota | Nectriaceae | 3 | 0.97 |
| Fusarium sporotrichioides | Ascomycota | Nectriaceae | 18 | 5.84 |
| Fusarium temperatum | Ascomycota | Nectriaceae | 1 | 0.32 |
| Fusarium torulosum | Ascomycota | Nectriaceae | 1 | 0.32 |
| Neofusicoccum eucalyptorum | Ascomycota | Botryosphaeriaceae | 1 | 0.32 |
| Neurospora crassa | Ascomycota | Sordariaceae | 1 | 0.32 |
| Pestalotiopsis brachiata | Ascomycota | Sporocadaceae | 1 | 0.32 |
| Pestalotiopsis paeoniicola | Ascomycota | Sporocadaceae | 1 | 0.32 |
| Phlebia acerina | Basidiomycota | Meruliaceae | 16 | 5.19 |
| Phlebia radiata | Basidiomycota | Meruliaceae | 1 | 0.32 |
| Rhizopus microsporus | Mucoromycota | Rhizopodaceae | 2 | 0.65 |
| Rosellinia corticium | Ascomycota | Xylariaceae | 2 | 0.65 |
| Schizophyllum commune | Basidiomycota | Schizophyllaceae | 2 | 0.65 |
| Talaromyces cecidicola | Ascomycota | Trichocomaceae | 2 | 0.65 |
| Trichoderma gamsii | Ascomycota | Hypocreaceae | 10 | 3.25 |
| Trichoderma koningiopsis | Ascomycota | Hypocreaceae | 2 | 0.65 |
Regarding life strategies, two large trophic groups stand out: plant pathogens and saprotrophs. Of the 53 species identified, according to the literature review, 37 species proved to be pathogenic. Of them, 21 were reported to cause damage to chestnut trees (Tab. 3), such as Gnomoniopsis smithogilvyi ([34]), with 2 isolates in A Coruña, 2 in Lugo, 2 in Pontevedra, and 1 in Ourense) and Cryphonectria parasitica ([35]), although in this case, it was isolated from a single gall (see Tab. S1 in Supplementary material).
Tab. 3 - List of phytopathogenic fungal species isolated in this work and their presence in symptomatic chestnut samples.
| Pathogen | Distribution | Host plants | Reported on Castanea |
Plant organs | Reference |
|---|---|---|---|---|---|
| Alternaria alternata | Cosmopolitan | Broad host range | Yes | Leaves, fruits | Ren et al. ([44]), Cakar & Akilli ([10]) |
| Botryosphaeria dothidea | Cosmopolitan | Broad host range | Yes | Bark, fruits | Akilli et al. ([5]), Hamasaki et al. ([25]) |
| Botrytis cinerea | Cosmopolitan | Broad host range | Yes | Fruits | Donis-González et al. ([17]), Rodrigues et al. ([46]) |
| Colletotrichum fioriniae | Cosmopolitan | Broad host range | Yes | Fruits | Cakar ([11]) |
| Cryphonectria parasitica | Cosmopolitan | Castanea spp., Quercus spp., Acer spp., Carpinus betulus | Yes | Bark of stems and branches, fruits | Rigling & Prospero ([45]), Cakar ([11]) |
| Diaporthe amygdali | Cosmopolitan | Broad host range | Yes | Bark | Aghayeva et al. ([2]) |
| Diaporthe eres | Cosmopolitan | Broad host range | Yes | Fruits, leaves | Ivić & Novak ([29]), Jiang et al. ([32]) |
| Diaporthe phaseolorum | Cosmopolitan | Broad host range | No | - | - |
| Diaporthe phillipsii | Europe | Vaccinium corymbosum | No | - | - |
| Diplodia corticola | Africa, America, Europe | Quercus spp., Vitis vinifera | No | - | - |
| Diplodia mutila | America, Europe | Broad host range | No | - | - |
| Diplodia sapinea | America, Asia, Europe | Broad host range | No | - | - |
| Diplodia seriata | Cosmopolitan | Broad host range | Yes | Branches and stems | Dar & Rai ([16]) |
| Dothiorella iberica | Cosmopolitan | Broad host range | Yes | Branches | Batista et al. ([8]) |
| Epicoccum nigrum | Cosmopolitan | Broad host range | Yes | Bark | Aghayeva et al. ([2]) |
| Fusarium acuminatum | Cosmopolitan | Broad host range | Yes | Fruits | Rodrigues et al. ([46]) |
| Fusarium avenaceum | Cosmopolitan | Broad host range | No | - | - |
| Fusarium culmorum | Cosmopolitan | Broad host range | Yes | Fruits | Donis-González et al. ([17]) |
| Fusarium foetens | Cosmopolitan | Mainly Begonia × Hiemalis | No | - | - |
| Fusarium graminearum | Cosmopolitan | Broad host range | Yes | Fruits | Donis-González et al. ([17]) |
| Fusarium lateritium | Cosmopolitan | Broad host range | No | - | - |
| Fusarium oxysporum | Cosmopolitan | Broad host range | Yes | Fruits | Rodrigues et al. ([46]) |
| Fusarium proliferatum | Cosmopolitan | Broad host range | Yes | Fruits | Ivić & Novak ([29]) |
| Fusarium sambucinum | Cosmopolitan | Broad host range | No | - | - |
| Fusarium solani | Cosmopolitan | Broad host range | Yes | Fruits | He et al. ([27]) |
| Fusarium sporotrichioides | Cosmopolitan | Broad host range | No | - | - |
| Fusarium temperatum | Cosmopolitan | Zea mays | No | - | - |
| Fusarium torulosum | Cosmopolitan | Broad host range | No | - | - |
| Gnomoniopsis smithogilvyi | Cosmopolitan | Castanea spp., Buxus sempervirens, Corylus avellana, Fraxinus ornus, Pinus pinaster, Quercus cerris, Quercus ilex | Yes | Leaves, flowers, fruits, shoots, stems, branches |
Lema et al. ([34]) |
| Mucor hiemalis | Cosmopolitan | Broad host range | Yes | Fruits | Jermini et al. ([31]) |
| Neofusicoccum eucalyptorum | Cosmopolitan | Eucalyptus spp. and other Myrtaceae | No | - | - |
| Neofusicoccum parvum | Cosmopolitan | Broad host range | Yes | Bark, stems, branches, fruits | Ciordia et al. ([13]), Seddaiu et al. ([47]) |
| Penicillium glabrum | Cosmopolitan | Broad host range | Yes | Fruits | Overy et al. ([39]) |
| Pestalotiopsis paeoniicola | America, Asia | Paeonia suffruticosa, Pouteria sapota | No | - | - |
| Rhizopus microsporus | Cosmopolitan | Oryza sativa, Zea mays, Helianthus annuus, Brassica juncea | No | - | - |
| Rhizopus arhizus | Cosmpolitan | Broad host range | No | - | - |
| Schizophyllum commune | Cosmopolitan | Broad host range | Yes | Bark, stems, branches | Takemoto et al. ([48]) |
As mentioned above, the genera Fusarium and Trichoderma were the most abundant. The multilocus phylogenetic analysis allowed the identification of isolates at the species level. The galls’ condition at the time they were collected could explain the presence of a great diversity of species belonging to these genera, since they were necrotic and thus a great diversity of saprophytic fungi was expected to be found. In the case of the genus Fusarium, isolates belonging to 13 different species were detected, being F. avenaceum (Fr.) Sacc., F. graminearum Schwabe, F. oxysporum Schltdl. and F. sporotrichioides Sherbakoff those with the highest incidence (Tab. S1 in Supplementary material). For the first time, 8 species were reported in association with D. kuriphilus galls: F. acuminatum Ellis & Everhart, F. culmorum (Wm. G. Sm.) Sacc., F. foetens Schroers, O’Donnell, Baayen & Hooftman, F. graminearum, F. solani (Mart.) Sacc., F. sporotrichioides, F. temperatum Scaufl. & Munaut and F. torulosum (Berk. & M.A. Curtis) Nirenberg (Tab. 2). The isolates obtained from the genus Trichoderma corresponded to 7 different species, mainly T. atroviride Bissett and T. harzianum Rifai (Tab. S1 in Supplementary material), being T. gamsii Samuels & Druzhinina and T. koningiopsis Samuels, Suarez & Evans first reported associated with D. kuriphilus galls (Tab. 2).
Discussion
The presence of D. kuriphilus is one of the greatest threats to chestnut trees, as it limits tree vigor and nut yields. Moreover, this wasp and its galls may play an important role in spreading fungal diseases. Both in our work and in previous studies, it has been observed that most isolated fungal species are phytopathogenic, thus providing evidence that galls can serve as a reservoir for plant pathogens as well as other tissues, mainly leaves, stems, shoots, or bark ([37]).
As previously reported, according to the literature review, of the 53 species identified, 37 were found to be pathogenic (Tab. 3) and can be transmitted horizontally to other plant species by means of free-living or gall-inducing insects. Of these pathogenic fungi, 21 species were found damaging chestnut trees, mostly fruits (Tab. 3).
Among the pathogenic fungi causing significant damage to chestnut trees, C. parasitica and G. smithogilvyi are the most important. C. parasitica causes chestnut blight, a serious disease that forms perennial necrotic lesions on the bark of stems and branches, leading to their death ([45]). This pathogen requires natural openings or wounds to penetrate the host tissue, so the galls produced by D. kuriphilus represent an entry point that could be related to the increased incidence of the disease in areas where both species are present ([35]). However, in the present work, it has only been detected in one gall, probably due to the time of the year and the randomness of the sampling. This opens the possibility for conducting a detailed study of gall-associated fungi in trees with canker and proving whether the presence of C. parasitica in the galls could influence the incidence of the disease, as reported in previous studies. G. smithogilvyi, first described in 2012, is an emerging pathogen recently found in chestnuts trees in Galicia that significantly affects crop yield ([3]). The most characteristic symptoms of this disease are the formation of cankers, leaf, flower, and gall necrosis, fruit mummification, and rot, both pre- and post-harvest, damaging nuts still present on the tree ([34] - Fig. 4). In several studies, this fungus was found as an endophyte, being isolated from galls, shoots, leaves, stems, flowers and asymptomatic immature fruits, and producing symptoms in them once they mature ([3], [34]).
Fig. 4 - Gnomoniopsis smithogilvyi symptoms and morphology: (a) premature fall of burrs with mummified nuts; (b) mummified nuts and premature opening of burrs affected by the pathogen; (c) characteristic brown rot of nuts; (d) colony morphology on PDA; (e) conidiomata formed on PDA; (f) conidia (scale bar: 10 µm).

Among other phytopathogenic fungi, the genus Fusarium stands out due to the diversity of species present and the large number of isolates obtained. These fungi produce different types of degradative enzymes and entomopathogenic mycotoxins such as fumonisins ([36]) and, in several studies, strains belonging to the Fusarium incarnatum-equiseti complex and the species Fusarium proliferatum (Matsush.) Nirenberg ex Gerlach & Nirenberg were isolated in necrotic galls produced by D. kuriphilus, which showed a high wasp mortality rate in laboratory tests ([1], [49]). However, according to Cooper & Rieske ([14]), the mortality of D. kuriphilus inside the galls could be related to the dryness and hardness of their necrotic tissues, which would make it difficult for adults to emerge and rule out a direct entomopathogenic effect. The genus Fusarium is not only associated with galls but it has also been detected as an endophyte in other chestnut tissues such as leaves, buds, stems, and fruits ([37]). In terms of pathogenicity, according to previous studies, there is evidence of chestnut fruit damage on F. acuminatum, F. culmorum, F. graminearum, F. oxysporum, F. proliferatum, and F. solani, species identified in this study (Tab. 3).
Furthermore, eight species of the Botryosphaeriaceae family were identified, known to be important pathogens in Europe and worldwide, namely Botryosphaeria dothidea, Diplodia corticola, D. mutila, D. sapinea, D. seriata, Dothiorella iberica, Neofusicoccum eucalyptorum and N. parvum (Tab. 3). This family includes some of the most important emerging and invasive plant pathogens worldwide ([4], [9]). Many of these species were reported for the first time on chestnut and on the D. kuriphilus galls in the present work, but only 4 of them were found causing symptoms in chestnuts: B. dothidea in bark ([5]) and fruits ([25]), D. seriata in branches and stems ([16]), D. iberica in branches ([8]), and N. parvum in bark, stems, branches ([13]) and fruits ([47]).
On the other hand, fungal spoilage of commercially purchased chestnuts is a cause for concern as the fungal colonization may have occurred at any stage of flowering, harvesting, storage, sorting, or transport. It can cause great economic losses and poses a serious health risk to consumers due to their mycotoxins ([39], [46]). Among these fungi, those belonging to the genera Aspergillus and Penicillium stand out as mycotoxin producers, secondary metabolites showing toxic, mutagenic, and teratogenic effects, including potential immunosuppressive activity and carcinogenic effects, having a long-term chronic or cumulative effect on human health ([42], [43]).
Beside the pathogenic species, it is worth highlighting the presence of fungi that can be used as biological control agents, such as Clonostachys rosea (Link) Schroers, Samuels, Seifert & W. Gams ([30]), and Trichoderma spp. ([28]). The latter is an important genus due to its diversity and abundance, including T. atroviride and T. harzianum, which are the main species found in this study. Trichoderma is a genus of fungi frequently associated with soils and the rhizosphere, though they also appear to colonize different parts of plants, such as leaves; that is, they are opportunistic symbionts ([26]). These fungi exhibit an antagonistic behavior against various pathogens, especially fungi, by inhibiting their growth through direct interactions, namely by antibiosis, competition, or mycoparasitism, or indirect ones, by increasing plant vigor and tolerance to stress ([28], [26]). Various studies evaluated different species against important pathogens of C. sativa, such as T. harzianum, T. parceramosum Bissett and T. viride Persoon, which showed antagonistic and protective effects against C. parasitica ([6], [7]) and T. atroviride against G. smithogilvyi ([40]). Therefore, further studies need to be conducted with Trichoderma strains isolated from Galician chestnut trees and evaluate their antagonism against new threats, such as G. smithogilvyi, which is one of the most important diseases found in chestnut stands in Galicia.
Conclusions
This work is the first study of fungal communities in D. kuriphilus galls in Galicia. The presence of a wide diversity of fungi, both phytopathogenic and saprophytic, has been identified, and some genera and species were reported for the first time. Therefore, this study provides interesting results for managing D. kuriphilus at the local scale and comparing it with fungal communities in galls found in other regions. However, further research is needed to determine the role of D. kuriphilus in dispersing these fungi.
Acknowledgments
The authors would like to thank Carmela Menduíña, Estación Fitolpatolóxica Areeiro, Pontevedra, Spain, for her contribution to identifying the isolates, and Marisol Guerreiro Lemos, Fauna Útil S.L., Pontevedra, Spain, for taking part in the samplings.
The authors declare that part of the data included in this paper was communicated as a poster presented at the 8th Spanish Forestry Congress held in Lleida (Catalonia, Spain) from 27 June to 1 July 2022 and included in the proceedings (in Spanish) under the title “Hongos asociados a agallas producidas por Dryocosmus kuriphilus en castaños de Galicia” (⇒ https://8cfe.congresoforestal.es/sites/default/files/actas/8CFE-1077.pdf).
References
CrossRef | Gscholar
Gscholar
CrossRef | Gscholar
Online | Gscholar
Authors’ Info
Authors’ Affiliation
María López-Burgos
Fauna Útil SL, Ctra. A Guarda-Tui km 194, Tomiño, Pontevedra (Spain)
Carmen Salinero 0000-0002-2061-0215
J Pedro Mansilla 0000-0003-3232-3712
Estación Fitopatolóxica Areeiro, Deputación de Pontevedra, Subida a la Robleda s/n, Pontevedra (Spain)
Corresponding author
Paper Info
Citation
Fraga Meizoso M, Aguín O, López-Burgos M, Salinero C, Mansilla JP (2024). The complexity of mycobiota associated with chestnut galls induced by Dryocosmus kuriphilus in Galicia (Northwestern Spain). iForest 17: 378-385. - doi: 10.3832/ifor4559-017
Academic Editor
Matteo Marchioro
Paper history
Received: Jan 11, 2024
Accepted: Aug 27, 2024
First online: Dec 14, 2024
Publication Date: Dec 31, 2024
Publication Time: 3.63 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2024
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 7234
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 3392
Abstract Page Views: 2146
PDF Downloads: 1368
Citation/Reference Downloads: 1
XML Downloads: 327
Web Metrics
Days since publication: 603
Overall contacts: 7234
Avg. contacts per week: 83.98
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Jul 2026)
Total number of cites (since 2024): 1
Average cites per year: 0.33
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Short Communications
Local spread of an exotic invader: using remote sensing and spatial analysis to document proliferation of the invasive Asian chestnut gall wasp
vol. 5, pp. 255-261 (online: 24 October 2012)
Research Articles
Case study of a new method for the classification and analysis of Dryocosmus kuriphilus Yasumatsu damage to young chestnut sprouts
vol. 5, pp. 50-59 (online: 10 April 2012)
Research Articles
Investigations on yellowing of chestnut crowns in Trentino (Alps, Northern Italy)
vol. 13, pp. 466-472 (online: 07 October 2020)
Research Articles
Fungal community of necrotic and healthy galls in chestnut trees colonized by Dryocosmus kuriphilus (Hymenoptera, Cynipidae)
vol. 12, pp. 411-417 (online: 13 August 2019)
Research Articles
Gnomoniopsis castaneae associated with Dryocosmus kuriphilus galls in chestnut stands in Sardinia (Italy)
vol. 10, pp. 440-445 (online: 24 March 2017)
Research Articles
Does management improve the state of chestnut (Castanea sativa L.) on Belasitsa Mountain, southwest Bulgaria?
vol. 8, pp. 860-865 (online: 27 April 2015)
Research Articles
Effects of drought and nutrient deficiency on grafts originating from sound and shaken sweet chestnut trees (Castanea sativa Mill.)
vol. 9, pp. 109-114 (online: 19 July 2015)
Research Articles
Brown rot on nuts of Castanea sativa Mill: an emerging disease and its causal agent
vol. 6, pp. 294-301 (online: 16 July 2013)
Short Communications
Endophytes in changing environments - do we need new concepts in forest management?
vol. 6, pp. 109-112 (online: 05 March 2013)
Research Articles
Density management diagrams for sweet chestnut high-forest stands in Portugal
vol. 10, pp. 865-870 (online: 06 November 2017)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







