Local spread of an exotic invader: using remote sensing and spatial analysis to document proliferation of the invasive Asian chestnut gall wasp
iForest - Biogeosciences and Forestry, Volume 5, Issue 5, Pages 255-261 (2012)
doi: https://doi.org/10.3832/ifor0633-005
Published: Oct 24, 2012 - Copyright © 2012 SISEF
Short Communications
Abstract
Remote sensing and spatial analysis represent useful tools for modeling species’ dispersal, characterizing the spread of invasions and the invasability of a region, and thus allowing more accurate predictions for developing mitigation strategies. American chestnut, Castanea dentata, was historically a dominant forest species in North America, but occurs only sporadically today after its functional elimination by an exotic fungal pathogen in the early 1900’s. In recent decades Castanea resources have increased due to restoration efforts, commercial chestnut plantations, and horticultural uses. This resurgence is threatened by an additional exotic species, the globally invasive Asian chestnut gall wasp, Dryocosmus kuriphilus. The gall wasp was first discovered in Lexington, Kentucky (USA) in 2010. We used remotely sensed data and Geographic Information Systems to describe the local distribution of the Castanea hosts, and the occurrence and dispersal of the gall wasp. We tested the hypotheses that geomorphology, Castanea occurrence, and prevailing winds influence local proliferation. We found that gall wasp spread may be attributable to host plant distribution and to the effects of prevailing winds occurring during a brief period of adult insect emergence, and is influenced by topography. Our results suggest that weather data and topographic features can be used to delineate currently infested areas and predict future gall wasp infestations.
Keywords
Dryocosmus Kuriphilus, Cynipidae, Remote Sensing, GIS, Spatial Analysis
Introduction
Exotic species invasions pose a substantial threat to biological integrity and sustainability. In our increasingly global economy, the rate of species’ invasions is increasing exponentially, compromising biodiversity and altering ecosystem function. Non-native invasive species can affect plant, animal, and human health, and these invasions can have devastating economic impacts ([30]). A critical challenge for invasive species management is gaining a complete understanding of the invasion process. Predicting the spatial and temporal dynamics of newly established invaders is crucial to understanding their proliferation and mitigating their impacts.
Emerging technologies such as remote sensing improve our ability to better understand factors influencing an invasion, including the invasability of an area, the dynamics of an invasion, predictions about invasiveness ([30], [16]), and mitigation of these invasions. The combined use of Global Positioning Systems (GPS) and Geographic Information System (GIS) offers a powerful set of tools to record movement and describe the behavior of invasive organisms. In the last few decades GIS-based analyses have been used effectively to investigate the pattern of dispersal of many diverse organisms. Hyperspectral images, GPS collected data and GIS have been used to locate and map invasive plants in California ([31]), and combined with spatial regression analysis, to identify the parameters affecting their spread ([7]). Remotely sensed data and GIS have also been used to understand invasions by naturalized horticultural imports ([20]) and woody plants ([29]).
To predict the impacts of the invasive hemlock woolly adelgid, Adelges tsugae Annand (Hemiptera: Adelgidae), in eastern North America, remote sensing and spatial analysis has been used to map the occurrence of the highly susceptible eastern hemlock, Tsuga canadensis (L.) Carr. ([5]). These technologies have also proven effective in modeling the spread of windborne and flying insects ([28], [26]). Remote sensing and GIS have been used to locate bark beetle (Coleoptera: Curculionidae) populations, characterize the scope and magnitude of infestations, and predict their spread ([35]). Similarly, gypsy moth, Lymantria dispar L. (Lepidoptera: Lymantriidae), distribution in North America has been described and predicted using GIS and spatial analysis ([21]). These technologies have also been used to monitor pests in cropping systems. Carriere et al. ([4]) modeled the movement of the lygus bug, Lygus hesperus Knight (Heteroptera: Myridae), through diverse settings. They tracked lygus bug populations within different crops over time, and were able to distinguish between source and sink locations. Clearly GIS-based spatial analyses allow a greater understanding of the biology, behavior and ecology of insects and how they interact with biotic and abiotic factors, and represent a powerful tool to mitigate the impacts of species’ invasions. We use these tools here to address local proliferation of an exotic insect pest attacking a sporadically occurring tree genus containing both native and non-native members, and to understand how local conditions influence its spread.
American chestnut, Castanea dentata (Marshall) Borkh, was historically a dominant component of forests of eastern North America ([3]), but was functionally eliminated by the 1904 introduction of the exotic chestnut blight fungus, Cryphonectria parasitica (Murr.) Barr ([14]). Lingering American chestnuts persist across the landscape as woody shrubs that reach 1-2 m in height before dying back, and Chinese chestnut, C. mollissima Blume, is widely planted as a landscape and horticultural tree.
However, chestnut is again under threat from an additional exotic invader, the Asian chestnut gall wasp (ACGW), Dryocosmus kuriphilus Yasumatsu (Hymenoptera: Cynipidae). The gall wasp induces formation of spherical, 1-3 cm galls on new spring shoots, thus disrupting tree growth, leading to plant decline and drastic yield reductions ([25]). D. kuriphilus is univoltine, and reproduces by thelytokous parthenogenesis ([11]). Larvae develop within the multi-chambered galls throughout the spring, pupate, and adults emerge in summer (late May to late July). Adult flight occurs over 10-15 days, depending on latitude ([11]). Each adult produces 100-150 eggs, and will oviposit up to 20 eggs in a single developing chestnut bud ([11]), which hatch in 30-40 days. First instar larvae overwinter within dormant chestnut buds until the following spring; gall development occurs concurrently with budbreak and leaf expansion. Detection within dormant buds is impossible by external plant inspection.
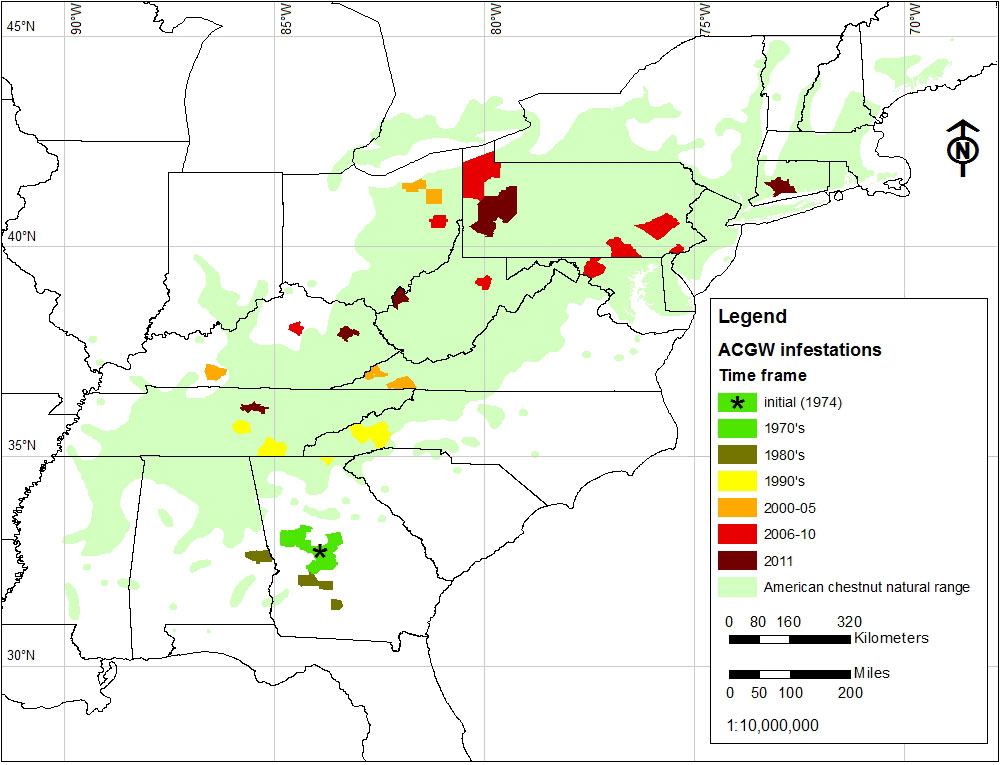
Since its initial introduction into North America in Peach County, Georgia, in 1974, the gall wasp has spread throughout the natural range of American chestnut (Fig. 1). Initially (1970s and 1980s) range expansion was fairly localized. However, beginning in the late 1990s there was a rapid range expansion in a north-northeasterly direction. Because of its cryptic nature gall wasp infestations spread easily due to movement of infested plant material, evidenced by the appearance of disjunct satellite populations in Ohio and Maryland ([27]). However dispersal by flight, influenced by winds, likely also plays a role ([1], [27], [12], [9]). Chestnut resources occur sporadically across the landscape, either as lingering American chestnut or as Chinese chestnut trees for horticultural purposes. Presumably D. kuriphilus utilized these resources, facilitated by wind, to spread through forested regions from northern Georgia to Virginia in the 1990’s and early 2000’s (Fig. 1 - [1], [27]). The Asian chestnut gall wasp is currently established in 11 states (Fig. 1), with infestations proliferating throughout the historic range of American chestnut.
Fig. 1 - Distribution and spread of the Asian chestnut gall wasp in North America since its introduction in 1974 (updated: December 2011).
On a localized scale, gall wasps disperse by active flight. Winds play a crucial role in influencing adult cynipid flight, with the direction of dispersal being consistent with that of prevailing winds ([17]). Wind speed is critical in determining the mechanism of this dispersal ([23], [9]). Low wind speeds stimulate adult flight (0.15-0.45 m s-1), after which flying wasps are carried on prevailing winds. While higher wind speeds inhibit flight (≥ 0.73 m s-1), these will also result in wasps being passively carried with the prevailing winds.
In 2010 the Asian chestnut gall wasp was discovered on a single Chinese chestnut in Lexington, Kentucky (USA). Lexington lies in the heart of the Bluegrass Region ([3]), in an area where suitable host plants occur only very sporadically. Chestnut was historically absent from the Bluegrass ([33]), but Chinese or hybrid chestnut trees planted in farmland and urban settings occur. Because the gall wasp is highly invasive in eastern North America and Castanea host plants occur only infrequently in the Bluegrass Region, we sought to understand its establishment and proliferation through the area. We used GIS and spatial analysis to map the distribution and dispersal of the gall wasp on a local scale (Lexington, Kentucky USA), and to evaluate factors influencing that spread. We evaluated local distribution of the gall wasp in relation to: (1) chestnut distribution; (2) landscape features; and (3) prevailing winds. We hypothesized that the local gall wasp population originated from a single source tree, and dispersed locally from the source tree through active flight by adult wasps, facilitated by prevailing winds, since movement of infested plant material within this locale is unlikely.
Materials and methods
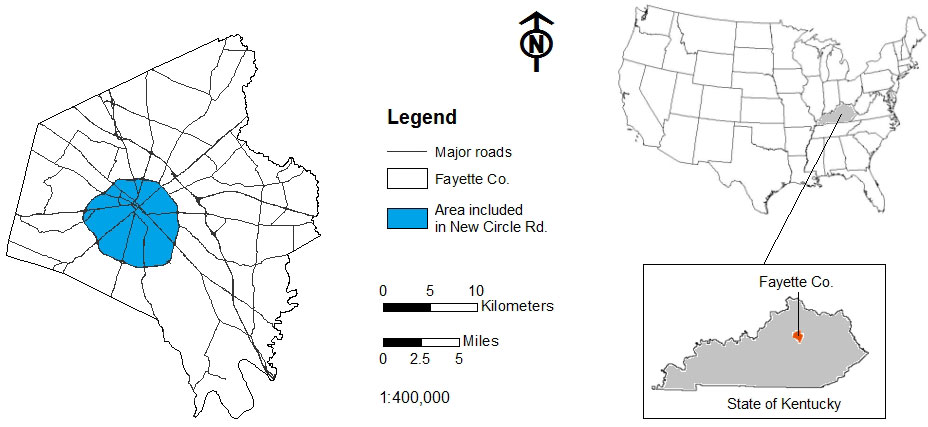
The city of Lexington, Kentucky was systematically surveyed for Castanea in 2010 and 2011 in the area encompassed within New Circle Rd. (Fig. 2). An additional area extending 1 km beyond New Circle Rd. was also surveyed. Each located chestnut was geo-positioned using a portable GPS device (Garmin Ltd, Olathe KS, USA), characterized (height, diameter, crown condition), and the presence or absence of the gall wasp was recorded (Tab. 1). These geopositioned chestnuts delineate our study area, and occur within an 8 × 5 km2 area (Fig. 3), with elevation ranging from 240 to 330 m a.s.l.
Fig. 2 - Area in Lexington, KY, surveyed for Castanea.
Tab. 1 - Location and characteristics of Castanea resources and status of infestation by the Asian chestnut gall wasp in Fayette County, KY (USA), determined during a systematic survey (2010-2011). L17 tree is the presumed infestation source. (a): diameter at 1.4 m above ground level; (b) crown condition - (1): live canopy 90-100%; (2) : live canopy 75-90%; (3): live canopy <75%; (c): ACGW infestation - low: <30% shoots galled, high: >30% shoots galled ([15], [9]). (*): estimated year of infestation.
| No. | Chestnut species |
Location | Setting | Elev (m) |
Slope (%) |
Aspect | Height (m) |
Diametera (cm) |
Crown conditionb |
ACGW infestationc |
Initial report (yr) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| L01 | C. dentata | Arboretum | park | 317.6 | 5.8 | S | 2 | 5 | 1 | none | - |
| L02 | Hybrid | Arboretum | park | 318.3 | 3.7 | S | 4.5 | 14 | 1 | none | - |
| L03 | Hybrid | Arboretum | park | 318.3 | 3.4 | S | 3 | 7 | 1 | none | - |
| L04 | C. mollissima | Arboretum | park | 318 | 1.5 | W | 5 | 12 | 1 | low | 2011 |
| L05 | C. mollissima | Arboretum | park | 318.2 | 1.4 | W | 4 | 9 | 1 | low | 2011 |
| L06 | C. mollissima | Arboretum | park | 317.9 | 1.1 | W | 8.5 | 26 | 2 | low | 2011 |
| L07 | C. mollissima | Cooper | yard | 313.2 | 4.5 | NE | 10 | 46 | 1 | low | 2011 |
| L08 | C. mollissima | Cooper | yard | 318.3 | 2.9 | NW | 5.5 | 25 | 1 | none | - |
| L09 | C. mollissima | Cooper | yard | 319.1 | 1.3 | W | 5.5 | 26 | 1 | none | - |
| L10 | C. mollissima | Cassidy | street | 316.6 | 0.2 | NE | 8 | 32 | 1 | low | 2011 |
| L11 | C. mollissima | Cassidy | street | 316.4 | 0.6 | SE | 8 | 38 | 2 | low | 2011 |
| L12 | C. mollissima | Cassidy | street | 316.4 | 0.6 | SE | 8.5 | 42 | 2 | low | 2011 |
| L13 | C. mollissima | Cassidy | street | 316.2 | 1.3 | NE | 9 | 38 | 2 | low | 2011 |
| L14 | C. mollissima | Cassidy | street | 316.1 | 2.2 | NW | 9.5 | 44 | 2 | low | 2011 |
| L15 | C. mollissima | Cassidy | street | 316.1 | 2.4 | NW | 9.5 | 37 | 2 | low | 2011 |
| L16 | C. mollissima | Fontaine | yard | 315.7 | 3.5 | NE | 8.5 | 32 | 1 | low | 2011 |
| L17 | C. mollissima | Fincastle | yard | 310.3 | 3 | SW | 8.5 | 31 | 2 | high | 2010 |
| L18 | C. mollissima | Oldham | yard | 303.7 | 0.5 | NW | 13.5 | 38 | 2 | low | 2011 |
| L19 | C. mollissima | Oldham | yard | 304.4 | 0.6 | W | 8.5 | 22 | 1 | low | 2011 |
| L20 | C. pumila | Arboretum | park | 316.7 | 6.4 | SW | 1.5 | 4 | 1 | none | - |
| L21 | C. mollissima | Tateswood | yard | 308.5 | 2.9 | S | 11 | 39 | 1 | low | 2011* |
| L22 | C. mollissima | Garden | yard | 314.1 | 2 | SW | 3.5 | 6 | 1 | low | 2011* |
| L23 | C. mollissima | Garden | yard | 313.9 | 2 | SW | 6.5 | 22 | 2 | low | 2011* |
| L24 | C. mollissima | Garden | yard | 314 | 2 | SW | 7.5 | 22 | 2 | low | 2011* |
| L25 | C. mollissima | Wilson | yard | 307 | 5.5 | NE | 8.5 | 30 | 2 | none | - |
| L26 | C. mollissima | Wilson | yard | 306.5 | 4 | NE | 7.5 | 30 | 3 | none | - |
| L27 | C. dentata | McConnell | woods | 282.4 | 13.2 | N | 4 | 7 | 1 | none | - |
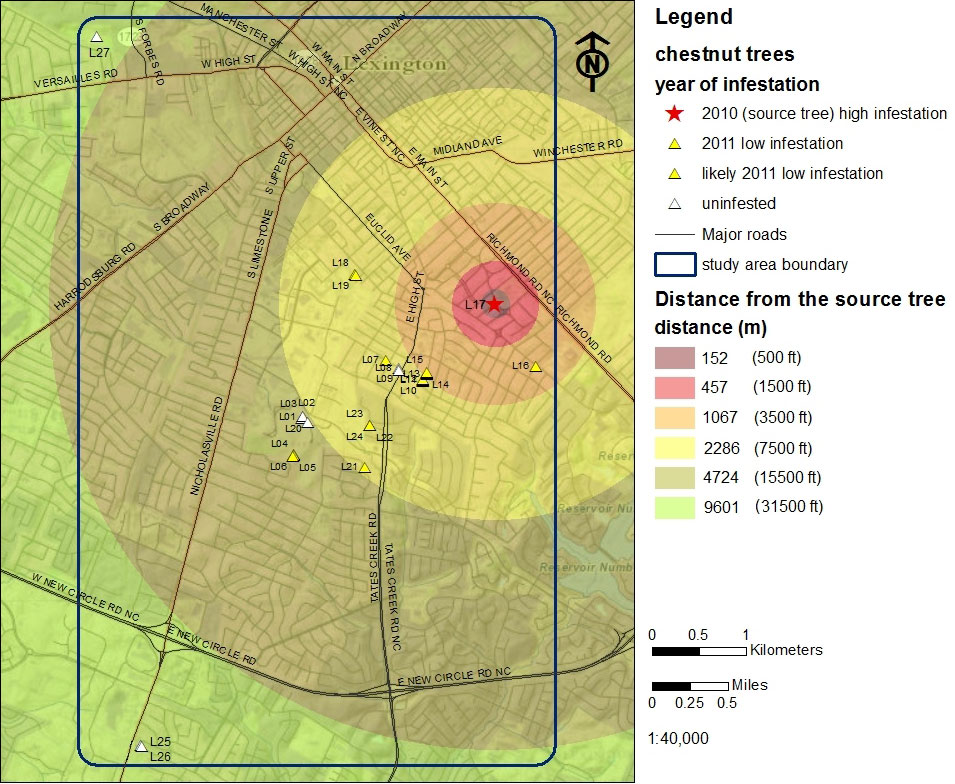
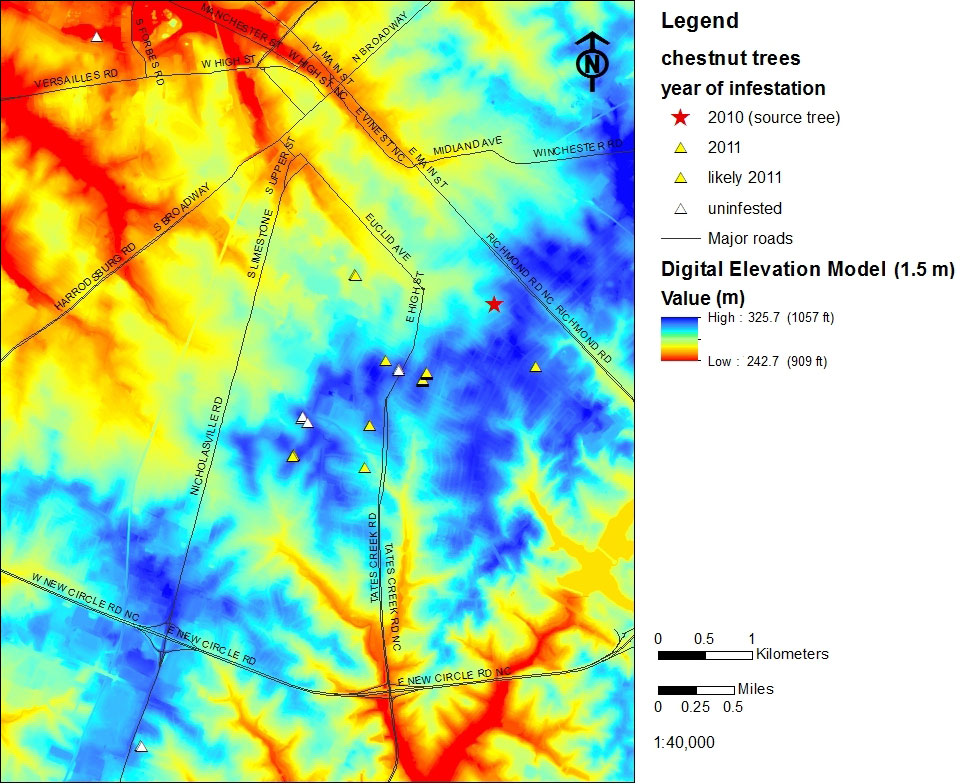
Fig. 3 - Location of Castanea trees and the relative incidence and location of gall wasp infested trees within the study area of Fayette Co., KY (USA).
Data on chestnut location and condition were georeferenced and included in a GIS using ArcGIS10 (ESRI, Redlands, CA, USA). Additional data obtained for spatial analysis included Fayette County boundaries, major roads, topographic information and digital orthoimages from the Kentucky Geographic Network website ([6]). We also obtained a digital elevation model (DEM) with 9.1 m (30 ft) resolution from the US Geological Survey National Elevation Dataset ([32]) and a second digital elevation model with 1.5 m (5 ft) resolution from the Geographic Network ([6]).
These additional data were combined with the results of our survey using ArcGIS10 to produce maps. Digital elevation models were used to visualize slope and aspect (9.1 m = 30 ft DEM) and land shape (high resolution 1.5 m = 5 ft DEM). Land shape was visualized by applying a dynamic range adjustment to the DEM ([2]). The slope was represented by using natural breaks (Jenks optimization method - [18]).
Wind direction and speed were extracted from the monthly climatological data of the Kentucky Climate Center ([34]) for the time period corresponding to the emergence and adult flight period for those wasps causing the infestation observed in 2011 (June 23 to July 4, 2010). These measurements were generated at the University of Kentucky South Farm (Lat. 37.98°, Lon. -84.53°), located 13.5 km south-southwest of the initial gall wasp find.
Results and Discussion
Chestnut distribution
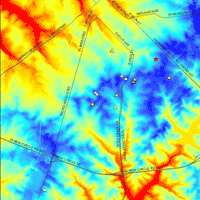
Our survey revealed the presence of 27 Castanea trees in a 40 km2 area (Fig. 3) within the greater surveyed area. Twenty trees were discovered in 2010 and seven additional trees were located in 2011 (Tab. 1). The majority (81%) are Chinese chestnut planted in residential yards or as street trees. Two (7%) are American chestnuts, two are C. dentata × C. mollissima hybrids, and one (4%) is a chinquapin (C. pumila). Chestnut resources in Fayette County are clustered (Fig. 3); nine (33%) C. mollissima are growing in the Cassidy - Cooper area. Seven (26%) are growing in the University of Kentucky Arboretum and State Botanical Garden, including three Chinese chestnuts, both hybrids, a single American and a chinquapin. Land shape clearly influences chestnut occurrence in our study area, but chestnut occurrence is confounded by human influences. All are growing at elevations of 303-319 m a.s.l. (Tab. 1, Fig. 4). The majority are located on low (3-15%) slopes (Tab. 1, Appendix 1), but their aspect is variable (Tab. 1, Appendix 2).
Fig. 4 - Digital elevation modeling (1.5 m resolution) demonstrates that chestnut occurs at relatively consistent elevations in the study area. See also Tab. 1.
Gall wasp presence and landscape features
In 2010 the gall wasp was found only on a single tree (L17) facing southeast (Tab. 1, Fig. 3). The initial gall wasp discovery was well established and the infestation level was high; this tree is considered the source tree for subsequent gall wasp infestations. After the initial gall wasp find in 2010, 17 additional hosts were found to be infested in 2011, primarily located in a south southwest direction. Thirteen of those trees were gall free during the initial 2010 survey, thus indicating that the 2011 infestations were due to dispersal of the wasp during the adult flight in summer 2010. The remaining four trees were surveyed for the first time in 2011 and found to be already infested. However, their very low galling rate suggests those trees were initially infested in summer 2010 as well. Seven trees remain gall wasp free. Thus 67% of the chestnut in our study area (18 trees) harbor confirmed gall wasp populations (Tab. 1, Fig. 3).
The trees newly infested in 2011 had very low gall wasp populations, and were 700 - 2700 m from the source tree, in a primarily south-southwest direction (Fig. 3). Aspect plays a role in gall wasp occurrence, since many of these newly infested trees were oriented northeast or east, essentially facing the source tree. Collectively this information suggests that the source tree and the infested chestnut are situated in positions exposed to the same wind flows: higher elevation (ridges), source tree facing newly infested trees, and low slopes.
Prevailing winds
There was a 12 days period of adult gall wasp flight in 2010 (June 23 - July 4) during which ambient conditions were relevant. The average daily temperature during this period was 24.3 °C, well within the acceptable temperature range for adult wasp flight (Tab. 2). Measurable precipitation, which clearly could impede flight, was recorded only on two days (24 and 28 June), during which solar radiation was less than 20 MJ m-2 and below the mean for the 12 days flight period (19.9 MJ m-2). The average relative humidity, 66%, did not deviate from normal (Tab. 2). On four days (June 25, June 30, July 1, and July 2) prevailing winds were south westerly. On one day (June 29) prevailing winds were directed easterly (Tab. 2). During the remaining days winds were directed mainly northeasterly at 1.8-4 m s-1 (mean = 3 m s-1 - Tab. 2), appropriate for passive adult transport (> 2.1 m s-1) rather than active flight.
Tab. 2 - Ambient conditions in Fayette Co., KY (USA), during the adult flight period of Dryocosmus kuriphilus (23 Jun - 4 Jul 2010)
| Date | Mean T (°C) |
Precipitation (mm) |
Solar radiation (MJ/m2) |
Humidity | Wind direction |
|
|---|---|---|---|---|---|---|
| minimum (%) | maximum (%) | |||||
| 23-Jun | 27.3 | 0 | 27.5 | 47 | 90 | SW |
| 24-Jun | 25.8 | 4.6 | 17.5 | 48 | 88 | W |
| 25-Jun | 24.8 | 0 | 29.5 | 40 | 86 | NNE |
| 26-Jun | 25.4 | 0 | 27.4 | 50 | 95 | SW |
| 27-Jun | 27.6 | 0 | 25.7 | 57 | 89 | SW |
| 28-Jun | 24.2 | 3.8 | 15.9 | 63 | 96 | WSW |
| 29-Jun | 24.9 | 0 | 24.6 | 39 | 98 | NW |
| 30-Jun | 21.3 | 0 | 30 | 38 | 83 | NE |
| 1-Jul | 20.1 | 0 | 31.2 | 32 | 77 | NE |
| 2-Jul | 20.8 | 0 | 31.5 | 31 | 78 | ENE |
| 3-Jul | 23.7 | 0 | 27.8 | 37 | 75 | SSW |
| 4-Jul | 25.7 | 0 | 24.8 | 47 | 93 | SSW |
In 2010 we located a single heavily gall-infested source tree. In 2011, sixteen of the 21 trees (76%) located southwest and east of the source tree were infested. Prevailing winds during adult flight (June 23 - July 4, 2010) were sufficient to account for the pattern of gall wasp-infested chestnuts documented in our surveys (Fig. 5). Nevertheless we must use caution in interpreting our result. The majority of the chestnut we located were southwest of the source tree, so it’s conceivable that the geographic distribution of these hosts could in and of itself be the cause of the infestation pattern we observed. However, to fully test our hypothesis would require a more evenly distributed host plant base than what is available in Lexington.
Fig. 5 - Hypothesized movement of the chestnut gall wasp in Lexington in the period June 23 - July 4 2010, resulting in newly infested trees in 2011.
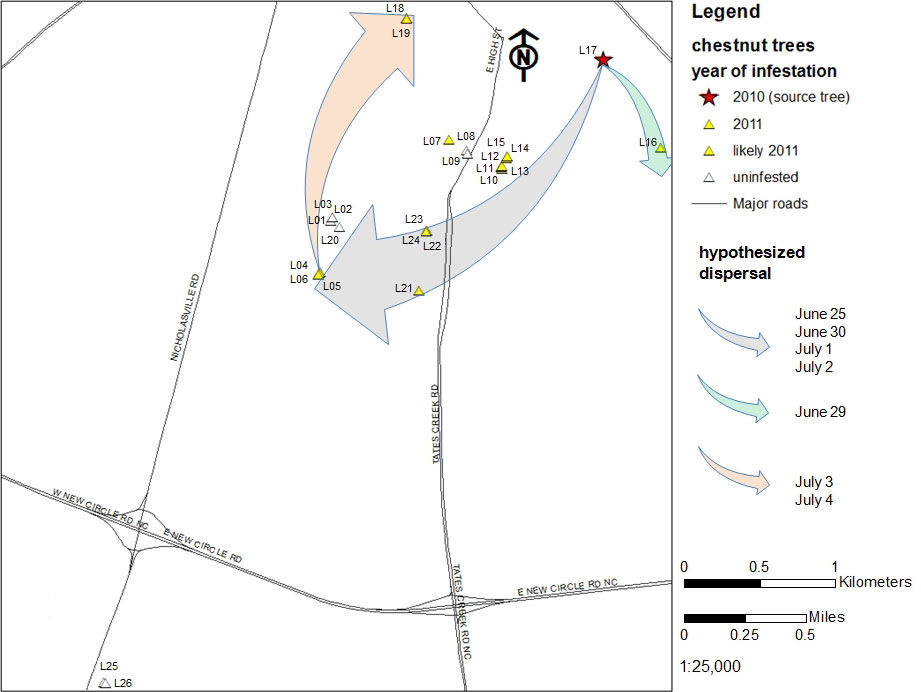
Our data suggest that proliferation of the gall wasp through the city of Lexington was determined by host plant distribution and was strongly affected by prevailing winds. Wind is a well-documented factor influencing insect dispersal ([19], [13], [22], [8]), including that of a cynipid gallmaker on oak (Quercus sp. - [17]), and is an important aspect of pest dispersal in agro forest settings ([10]). Our data suggests that adult Asian chestnut gall wasps were transported by wind in a southwesterly direction from the source tree to suitable hosts on four dates (Fig. 5): June 25, June 30, July 1 and July 2. Evidence also suggests that winds blowing southeasterly on June 29 facilitated movement of wasps from the source tree to the tree L16, located south southeast of the source tree (Fig. 5). Furthermore, winds on July 3 and 4 were of suitable speed and an appropriate direction to passively transport adults to two trees in the northeastern edge of the study area (L18 and L19 - Fig. 5).
Not all hosts southwest of the source tree were infested. The gall-free trees L08 and L09 were heavily pruned in winter 2010, potentially removing infested buds and locally extinguishing the gall wasp population. Trees L1, L2, L3 and L20 are surrounded by vegetation that may have perturbed wind flow, disrupting exposure to prevailing winds that would carry adult wasps and lead to gall wasp infestations. Vegetation, buildings, and other windbreaks do impact local insect movement ([17], [24]), and likely play a role in the patterns of gall wasp proliferation we observed.
The maximum distance the gall wasp dispersed in Lexington during the 2010 flight period was the distance between the source tree L17 and the L4 L5 L6 cluster, approximately 3 km (Fig. 3, Fig. 5). The gall wasp did not colonize trees L25 and L26, located 6 km southwest of the source, but those are very isolated hosts at twice the distance. Our findings on local gall wasp proliferation are comparable with data from Europe, where gall wasp dispersal by flight has been reported at 8 km per year ([9]). Dispersal rates of 15-25 km per year were reported in the USA in the years immediately following the gall wasp introduction in Georgia in 1974, but the movement of infested plant material was likely involved ([27]).
Conclusions
We used remotely sensed data and GIS technology to characterize local proliferation of a newly arrived exotic insect, the Asian chestnut gall wasp, attacking a sporadically occurring tree in the Bluegrass Region of Kentucky. We sought to understand how host plant distribution, landscape characters, and prevailing winds might influence the gall wasp’s spread in the formative stages of the invasion and barring movement of infested plant material by humans.
This is the first use of remotely sensed data and GIS analysis to document movement of the invasive Asian chestnut gall wasp. Our analysis provides insight into factors that affect the local spread and proliferation of the gall wasp in a newly invaded area. The gall wasp colonized 17 sporadically distributed hosts found at a very low density within a 40 km2 area. Our analysis demonstrates how the invasiveness of the gall wasp is enhanced by abiotic factors. This work will provide a useful tool and model system to test hypotheses and to make predictions about the spread of this invasive pest on local scale, leading to appropriate mitigation efforts.
Acknowledgements
The authors thank Demetrio Zourarakis for assistance with experimental protocol, Melanie Sprinkle for field assistance, and Josh Adkins, Luke Dodd, and two anonymous referees for reviewing early versions of this manuscript. This work was funded in part by the Italian Fulbright Commission (to IG), the Northern Nut Grower’s Association, the American Chestnut Foundation, the USDA Forest Service, and McIntire Stennis Funds from the Kentucky Agricultural Experiment Station, and is published as Experiment Station Project 12-08-032.
References
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
LK Rieske
Department of Entomology, University of Kentucky, S225 Agricultural Science North, 40546-0091 Lexington, KY (USA)
Corresponding author
Paper Info
Citation
Graziosi I, Rieske LK (2012). Local spread of an exotic invader: using remote sensing and spatial analysis to document proliferation of the invasive Asian chestnut gall wasp. iForest 5: 255-261. - doi: 10.3832/ifor0633-005
Academic Editor
Andrea Piotti
Paper history
Received: Apr 25, 2012
Accepted: Oct 06, 2012
First online: Oct 24, 2012
Publication Date: Oct 30, 2012
Publication Time: 0.60 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2012
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 66425
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 54861
Abstract Page Views: 3783
PDF Downloads: 5944
Citation/Reference Downloads: 26
XML Downloads: 1811
Web Metrics
Days since publication: 5037
Overall contacts: 66425
Avg. contacts per week: 92.31
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Jul 2026)
Total number of cites (since 2012): 12
Average cites per year: 0.80
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Fungal community of necrotic and healthy galls in chestnut trees colonized by Dryocosmus kuriphilus (Hymenoptera, Cynipidae)
vol. 12, pp. 411-417 (online: 13 August 2019)
Research Articles
Gnomoniopsis castaneae associated with Dryocosmus kuriphilus galls in chestnut stands in Sardinia (Italy)
vol. 10, pp. 440-445 (online: 24 March 2017)
Research Articles
The complexity of mycobiota associated with chestnut galls induced by Dryocosmus kuriphilus in Galicia (Northwestern Spain)
vol. 17, pp. 378-385 (online: 14 December 2024)
Research Articles
Case study of a new method for the classification and analysis of Dryocosmus kuriphilus Yasumatsu damage to young chestnut sprouts
vol. 5, pp. 50-59 (online: 10 April 2012)
Review Papers
Remote sensing-supported vegetation parameters for regional climate models: a brief review
vol. 3, pp. 98-101 (online: 15 July 2010)
Review Papers
Accuracy of determining specific parameters of the urban forest using remote sensing
vol. 12, pp. 498-510 (online: 02 December 2019)
Review Papers
Remote sensing of selective logging in tropical forests: current state and future directions
vol. 13, pp. 286-300 (online: 10 July 2020)
Research Articles
Estimating the potential threat of increasing temperature to the forests of Turkey: a focus on two invasive alien insect pests
vol. 15, pp. 444-450 (online: 03 November 2022)
Technical Reports
Detecting tree water deficit by very low altitude remote sensing
vol. 10, pp. 215-219 (online: 11 February 2017)
Research Articles
Geostatistical techniques for estimating aboveground biomass in eastern Amazonia
vol. 19, pp. 85-93 (online: 13 March 2026)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







