
Gnomoniopsis castaneae associated with Dryocosmus kuriphilus galls in chestnut stands in Sardinia (Italy)
iForest - Biogeosciences and Forestry, Volume 10, Issue 2, Pages 440-445 (2017)
doi: https://doi.org/10.3832/ifor2064-009
Published: Mar 24, 2017 - Copyright © 2017 SISEF
Research Articles
Abstract
Invasive fungal pathogens and pests of sweet chestnut (Castanea sativa Mill.) forests drastically reduce their productivity. The recently described Gnomoniopsis castaneae is one of the main agents involved in the epidemic of brown rot of chestnut nuts worldwide. In 2014, during an investigation aimed at evaluating the health status of chestnut forests in Sardinia, a high incidence of necrotic galls induced by the Asian gall wasp Dryocosmus kuriphilus Yasumatsu (Hymenoptera, Cynipidae) was observed. Several fungal isolates were consistently isolated from necrotic gall tissues. Based on their morphological characters and analyses of both the ITS and EF1-α-coding gene sequences, all isolates were identified as Gnomoniopsis castaneae.
Keywords
Galls Necrosis, Brown Nut Rot, Invasive Pests, Forest Pathogens
Introduction
Over the last decades, insect and pathogen invasions have exponentially increased in Europe, damaging forests and other ecosystems ([33]). The introduction of a non-native microorganism may in fact negatively affect the existing trophic interactions and alter the equilibrium of natural ecosystems. Plant trade is one of the major pathways of introduction for many fungal pathogens and insect pests ([3]).
Sweet chestnut (Castanea sativa Mill.) is one of the best example of a forest tree species threatened by invasive pathogens and insect pests, with major impacts caused by Cryphonectria parasitica, Phytophthora spp. and Dryocosmus kuriphilus ([17], [32], [41], [30], [34], [39], [31]). Sweet chestnut is a typical nut crop of the Mediterranean basin and one of the most common forest trees in the hilly parts of Italy ([2]). In Sardinia, sweet chestnut grows between 500 and 1300 m a.s.l., cultivated for both fruit and timber production ([5]). The largest populations are located in the western area of the Gennargentu Mountain, with about 2239 hectares ([15]). Actions for sustainable territorial development of stakeholders allowed the revival of many recovery and promotional initiatives. The Sardinian government supported such actions by funding a regional program based on the principles of multi-functionality of this forest ecosystem (⇒ http://www.regione.sardegna.it/documenti/1_274_20121023092725.pdf). Several actions have been undertaken to improve the management of chestnut orchards and to tackle phytosanitary problems, including trunk injection of potassium phosphite to control ink disease ([12]) and spread of hypovirulence of C. parasitica to control chestnut blight ([39]).
Nonetheless, chestnut forests in Sardinia have been infested by the Asian gall wasp Dryocosmus kuriphilus Yasumatsu (Hymenoptera Cynipidae), as in many other parts of Italy ([4], [14]). The chestnut gall wasp is native to northern China and it is considered one of the most important pests of chestnut worldwide, causing significant damage to European chestnut, as well as to the susceptible eurojapanese hybrids ([9]). Dryocosmus kuriphilus is a quarantine pest and is included in the A2 list of the European and Mediterranean Plant Protection Organization ([10]). It is a univoltine species that reproduces by parthenogenesis. Adults, all females, emerge in the early summer and lay eggs in the buds of the host plant. The eggs hatch soon after, but larval growth begins at budbreak the following spring, when the larvae induce the rapid formation of stem, petiole, and leaf galls. Galling prevents normal shoot growth and fruiting ([17], [40]). In Europe, the insect was first observed in 2002 in Italy ([4]). In Sardinia it was reported in 2007, whereas on the basis of collected data its introduction probably occurred between 2003 and 2005 by seedlings from Piedmont nurseries ([27]). Since 2009, biological control of populations of the Asian chestnut gall wasp has been performed using the natural enemy Torymus sinensis Kamijo ([22]). On the basis of experiences carried out in Italy ([30], [28]), the use of natural enemies for the biological control of D. Kuriphilus, besides the prevention of chestnut stand degradation and abandonment during the long delay period before the parasitoid is effective, resulted in a successful reduction of pest population. Investigations on gall development induced by D. kuriphilus on Castanea sativa provide indications of the critical time periods for the management of the pest and its effective control ([31]).
Several fungal species have been associated with necrotic galls of D. kuriphilus, including Fusarium and Alternaria species ([1], [38]). Species in the Colletotrichum genus have been reported causing extensive larval mortality of D. kuriphilus ([11], [13]). Recently, necrosis on galls of D. kuriphilus and insect’s death have been associated with a Gnomoniopsis sp. ([23], [42]).
The possibility of using these fungal species as biocontrol agents against D. kuriphilus and evaluating their impacts on plants and animals, is considered an alternative to natural enemies ([1], [38]). However, the genus Gnomoniopsis is found as endophyte in healthy plants, but also behaves as pathogen infecting branches, leaves and chestnut fruits ([44], [24], [35], [7], [29]).
In 2014, during an investigation aimed at evaluating the health status of chestnut forests in Sardinia, galls with black lesions on the exterior and often with a slight depression were observed, suggesting a possible involvement of the above mentioned fungal pathogens.
The aim of this study was to isolate and identify the fungi associated with necrosis of D. kuriphilus galls in chestnut areas of Sardinia. To this end, the fungal species isolated from these galls were characterized using both morphological and molecular methods. The fungus most consistently isolated, Gnomoniopsis castaneae, was identified as the most likely agent of the necrosis associated with D. kuriphilus galls.
Materials and methods
Study sites and sampling
Chestnut stands were selected in spring 2015 in two villages, Aritzo and Belvì (Nuoro province) located in the Sardinian region of “Barbagia di Belvì”, on the basis of the high presence of galls induced by D. kuriphilus. Two sites per village were selected (Tab. 1) and for each site sampling was undertaken on five chestnut trees, including one central tree and the four most adjacent trees radially. For each tree, 10 symptomatic and 5 asymptomatic galls were collected at the end of June 2015 from both twigs and leaves. The samples were collected from random locations in each tree, bagged and labeled with an identification code and collection date, and stored in the laboratory at 4 °C for subsequent analysis.
Tab. 1 - Main data station of survey sites.
| Site | Aritzo (A1) | Aritzo (A2) | Belvì (B1) | Belvì (B2) |
|---|---|---|---|---|
| Elevation (m a.s.l.) | 890 | 990 | 936 | 910 |
| Exposure | North | North-West | North-East | North-East |
| Morphology | Hill | Hill | Hill | Hill |
| Slope% | 20 | 15 | 10 | 5 |
| Substrate | Schists | Schists | Schists | Schists |
| Crop | Chestnut-Hazel | Chestnut-Hazel | Chestnut | Chestnut |
| Management | Coppice | High Forest | High Forest | High Forest |
| Geographic Coordinates | 39°56′ 57.87″ N 09°11′ 23.21″ E |
39°56′ 55.78″ N 09°11′ 45.12″ E |
39°58′ 20.32″ N 09°09′ 41.04″ E |
39°58′ 34.87″ N 09°09′ 32.64″ E |
Fungal isolation and DNA analysis
A total of 200 symptomatic galls and 100 asymptomatic galls were surface sterilised by immersing each gall in 70% ethanol for 1 min. For symptomatic galls, portions of tissues from the margin of each necrotic lesion were placed onto PDA (Potato Dextrose Agar) and incubated in a growth dark chamber at 21 °C. Portions of tissues taken just under exterior tissues of asymptomatic galls were placed and incubated using the same conditions.
Representative samples of each growing colony were subcultured onto PDA for morphological and molecular identification. Cultures were stored at the Cork and Silviculture Research Department - Agris Sardegna.
For morphological identification, three replicates of each isolate on PDA were incubated at 23 °C in the dark for 20 days. Colony morphology was characterized by observing its form, margin, surface, growth and colour. Conidia were measured under an illuminated Carl Zeiss light microscope (Carl Zeiss, Standard RA 38, Germany) at 400× magnification. Standard deviation and length ratio were determined for conidia.
DNA was extracted from mycelium of colonies ([25]). The universal primers ITS1 and ITS4 ([47]), were used to amplify the ITS-1, 5.8S and ITS-2 regions of the nuclear rDNA. Primers EF1-728F and EF1-1199R ([46]) were used to amplify the EF1-α-coding gene. Successful amplifications were verified by gel electrophoresis at 90 V for 40 min with 1.5% agarose gel and 4 μl of SYBR Safe. PCR products were purified by Eurogold® Gel Extraction Kit (Euroclone, Pero, Milan, Italy) according to the manufacturer’s instructions and sequenced by BMR Genomics Lab (⇒ http://www.bmr-genomics.it). All sequences were identified by querying the GenBank database, using the nucleotide BLAST (⇒ https://blast.ncbi.nlm.nih.gov) search option (blastn), available through the National Center for Biotechnology Information and the UNITE online database (⇒ http://unite.ut.ee/index.php). Sequence similarity was assessed by Multiple Sequence Alignments using the software Clustal Omega (⇒ http://www.clustal.org). Sequences were deposited in GenBank. Accession numbers for ITS sequences are: LT593843 for isolate B1P1RS0; LT593844 for isolate A1P1GS0; LT593845 as representative of isolates A2P4GS0, B1P2GS0, B2P5GS0, A2P4GS0; LT593847 for isolate B2P2GS0; LT593846 as representative of isolates B2P1GS0, A1P2GA0, A2P3GS0, A1P5GS0, A2P5GS0, B2P5RS0, A1P1RS0. Accession number for EF1-α-coding gene sequences is: LT593848 as representative of isolates A1P1GS1, B1P1RS1, B2P5GS1, A1P2GA1, B1P2GS1.
Phylogenetic analyses were carried out using the software MEGA version 7 (⇒ http://www.megasoftware.net). Neighbour-joining trees were constructed using Kimura’s two-parameter model ([18]) with 1000 bootstrap replicates, including sequences from other species retrieved from databases.
Results and discussions
We isolated mycelia cultures from 200 symptomatic galls and 100 asymptomatic galls in order to identify the fungi associated with necrosis. In 68% of symptomatic and 34% of asymptomatic galls collected, we observed all colonies with the same features: woolly and dense mycelium, colour variable from greyish to brown (Fig. 1). Only from 1% of symptomatic galls, two different fungal agents were isolated (two colonies with the same aspect in one of the two galls and two colonies with another aspect from the other gall). In 12% of symptomatic and 56% of asymptomatic galls, no fungus was isolated. The 19% of symptomatic galls and 10% of asymptomatic galls resulted contaminated from bacteria. The colonies with the same features isolated from symptomatic and asymptomatic galls were regular in outline and developed in concentric circles. Orange and creamy drops of conidial growth appeared in concentric rings or scattered on the surface of older colonies. Conidia were hyaline, oval, obovoid, pyriform, or curved with an average size (n = 120) of 6.5 ± 0.58 μm (SD) × 2.2 ± 0.26 μm (length × width). On the basis of these morphological features, the fungus was identified as a Gnomoniopsis sp. This was confirmed by sequencing the ITS region from 13 isolates and the EF1-α locus from five isolates originated from symptomatic and asymptomatic galls. In order to check the genetic variability of the strains, a multiple sequence alignment was made. Sequence alignments of the ITS region (523 bp) and the EF1-α-coding gene (578 bp) showed 99.62% and 100% similarity, respectively, across all isolates, suggesting that they belong to the same species. Interestingly, an adenine deletion was shared by four of the isolates, which originated from both village locations. BLAST comparison of the 13 ITS sequences gave very high similarity with G. smithogilvyi (from 99% to 100%, 100% query coverage) and G. castanea (99%, but 94% query coverage) and comparison of the five EF1-α-coding gene sequences gave very high similarity (100%, 100% query coverage) with G. smithogilvyi and an unknown Gnomoniopsis species. In order to assign our sequences to a taxonomic species, a phylogenetic tree including sequences of G. smithogilvyi and G. castanea, in addition to sequences of two other species of Gnomoniopsis was built (Fig. 2).
Fig. 1 - (a) Necrotic galls infested by Gnomoniopsis castaneae; (b) colony of Gnomoniopsis castaneae on PDA; (c) conidia drops; (d) conidia. Scale bars: 10 μm.

Fig. 2 - Neighbour-joining trees based on a multiple sequence alignment of the ITS region of rDNA (A) and EF1-α locus (B) from fungi isolated in this work and from other fungal species retrieved from GenBank. Apiognomonia veneta was used as outgroup as in Visentin et al. ([44]). Soil sample codes: A1 and A2 indicate Aritzo (site 1 and site 2), B1 and B2 indicate Belvì (site 1 and site 2), P1-P5 indicates plant number, GS and GA identify symptomatic and asymptomatic galls, RS identify symptomatic twigs.
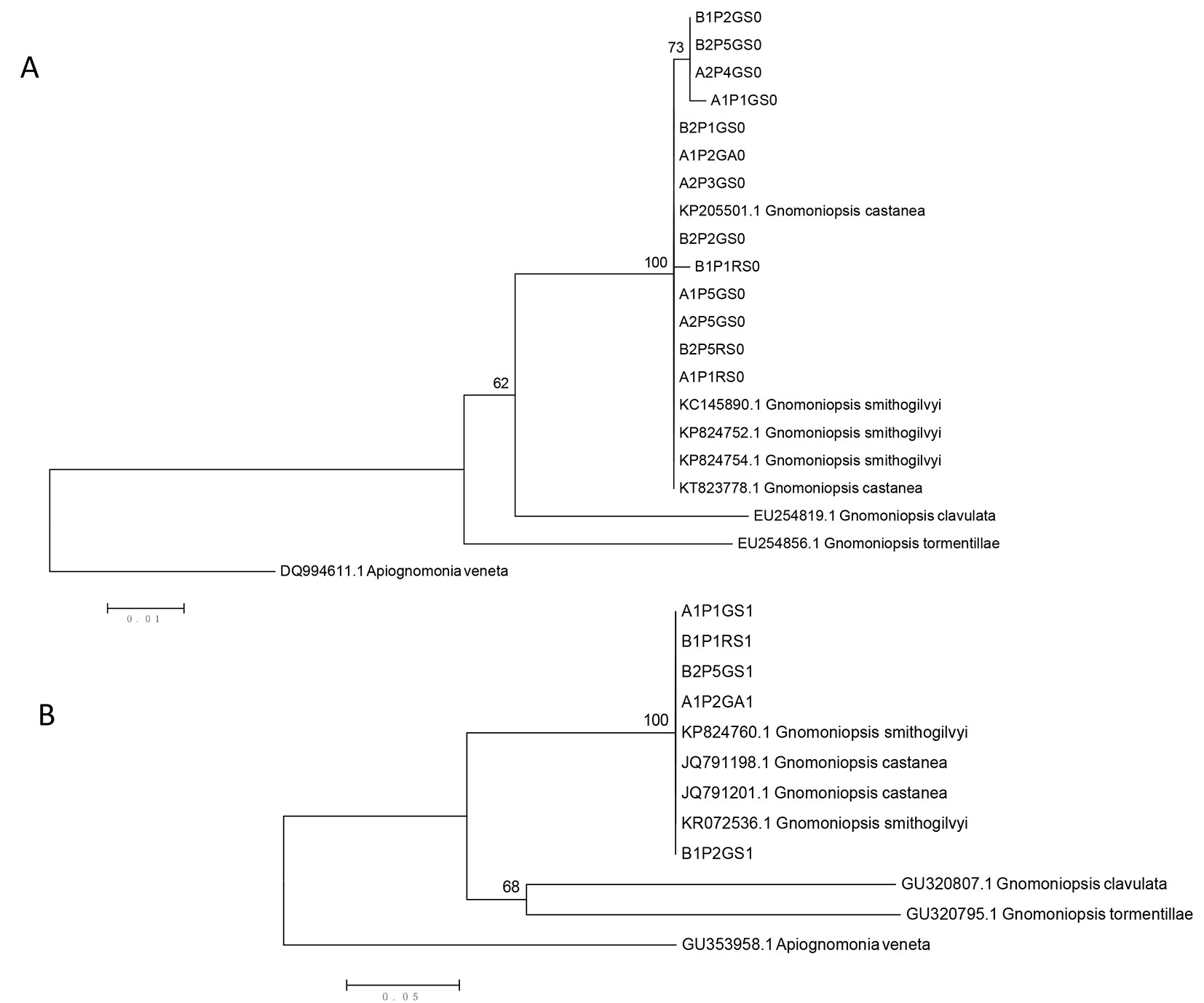
Results for both ITS and EF1-α showed that the fungus consistently isolated in this study clustered together with G. smithogilvyi and G. castanea, and separately from the other Gnomoniopsis species G. clavulata and G. tormentillae. In the ITS tree the four isolates sharing the same deletion clustered together with G. smithogilvyi and G. castanea. Since G. smithogilvyi and G. castanea are conspecific, as clearly stated by Meyer et al. ([26]) and Shuttleworth et al. ([36]), and given that Pasche et al. ([29]) suggested using the name G. smithogilvyi as the agent of the chestnut shoots canker symptoms in Castanea sativa, we can conclude that the causal agent of the necrosis associated with D. kuriphilus galls in the chestnut areas located in the “Barbagia di Belvì” region of Sardinia is G. smithogilvyi. However, according to Tamietti ([37]), since G. castaneae (“castanea”) was described and published earlier than G. smithogilvyi, the legitimate name of the fungus is G. castaneae (“castanea”). For this reason hereafter we follow this rule to indicate the agent of the chestnut shoots canker symptoms in Castanea sativa.
The finding of G. castaneae on both symptomatic and asymptomatic galls suggests that this fungus may behave as both endophyte and pathogen. Endophytes are known to produce pathogenic lesions on cynipid galls and interact with gall-forming insects ([48]).
In this study, we started to evaluate whether chestnut gall necrosis can affect the vitality and emergence of D. kuriphilus and parasitoids, in Sardinia. The rates of living larvae (unidentified species) as well as pupae and adults of D. kuriphilus, recovered inside necrotic and asymptomatic galls, were statistically compared. In addition, some observations on adult emergence of D. kuriphilus and parasitoids from necrotic galls were conducted. Preliminary data showed no adverse effects of gall necrosis on insect vitality. However, the number of chestnut gall samples used so far for vitality test was too small and further more accurate surveys are required to obtain reliable results. In this context, Lione et al. ([20]) reported a large number of living adults inhabiting galls colonized by G. castaneae and a large number of emerged D. kuriphilus, suggesting that an interaction between insect and fungus can occur during the insect development. By contrast, Vannini et al. ([43]) reported a relevant mortality of gall wasp before the adult flight period, suggesting G. castaneae as one of the most efficient sources of natural biological control of D. kuriphilus in Europe. It should be considered that, the impact of endophytic fungi on gall wasp vitality is correlated with structural gall traits that vary significantly according to geographic location and development phase ([6]).
The two additional fungal agents we isolated were identified as Fusarium lateritium and Cladosporium sp. Fusarium lateritium is a plant pathogen, reported as the causal agent of nut gray necrosis on hazelnut ([45]). Addario & Turchetti ([1]) also isolated Fusarium from necrotic galls.
Gnomoniopsis castaneae has recently been reported in Sardinia as a weak pathogen on hazelnuts that grows in the same area of chestnut forests ([21]). According to Lione et al. ([20]) the infestations of D. kuriphilus could have increased the pathogen spreading ability. On the basis of these works we propose two possible scenarios on the diffusion of G. castanea in Sardinia after its introduction: (i) it may be widespread from chestnut on the hazelnut without producing disease on this host; (ii) it was already present on hazelnut as weak pathogen and then has infested chestnut trees after gall wasp invasion. Considering that Sardinia chestnut forests are totally isolated from the Italian chestnut range, the relationship between host and pathogen could have different performances compared to the continental case. However, often the lack of data on host or pathogen diversity and biogeography makes it difficult to say whether a disease has been favored by changing environmental conditions and does not yield reliable conclusions on the host-pathogen relationship ([8]). Recently, problems never observed before on the quality of chestnut productions have been reported by the local population. Chestnut forests are very important for local economy because of both fruits and timber productions in rural areas of Sardinia. G. castanea is currently considered the main agent of nut rot ([44], [7], [19]). The presence of G. castaneae in Gennargentu Mountain ecosystems could, therefore, represent a danger in chestnut productions. Furthermore, since Gnomoniopsis spp. exist as endophytes in healthy host plants ([24], [7]) and the European system of plant biosecurity is based principally on visual inspections for symptoms, it is difficult to detect pathogens in asymptomatic plants ([16]) and therefore, the risk of G. castaneae spreading with visually healthy host plants is very high.
This is the first investigation on presence of G. castaneae infecting chestnut cynipid galls in Sardinia. Future research is needed to deeper analyse the interaction between D. kuriphilus and G. castaneae and to understand how this fungus can affect the health status of Mediterranean chestnut ecosystems and then, the quality of their products. The availability of specific primers ([19]) will help to fast monitor the presence of G. Castaneae in these ecosystems.
Acknowledgements
The authors would like to thank Sarah Green (Forest Research, Scotland) for her kindly help to English language editing. We thank Giuseppe Manconi, Giovanni Maria Mureddu and Giovanni Antonio Puliga for technical assistance. This research was funded by the Sardinian Regional Government’s Department of Environmental Protection.
References
Gscholar
Gscholar
Online | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Anna Cerboneschi
Clizia Sechi
Cork and Silviculture Research Service, Agris Sardegna, v. Limbara 9, I-07029 Tempio Pausania (Italy)
Institute for Sustainable Plant Protection, SS Torino - CNR, v.le Mattioli 25, I-10125 Torino (Italy)
Corresponding author
Paper Info
Citation
Seddaiu S, Cerboneschi A, Sechi C, Mello A (2017). Gnomoniopsis castaneae associated with Dryocosmus kuriphilus galls in chestnut stands in Sardinia (Italy). iForest 10: 440-445. - doi: 10.3832/ifor2064-009
Academic Editor
Alberto Santini
Paper history
Received: Mar 21, 2016
Accepted: Nov 27, 2016
First online: Mar 24, 2017
Publication Date: Apr 30, 2017
Publication Time: 3.90 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2017
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 53302
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 42709
Abstract Page Views: 4571
PDF Downloads: 4243
Citation/Reference Downloads: 183
XML Downloads: 1596
Web Metrics
Days since publication: 3393
Overall contacts: 53302
Avg. contacts per week: 109.97
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2017): 17
Average cites per year: 1.89
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
The complexity of mycobiota associated with chestnut galls induced by Dryocosmus kuriphilus in Galicia (Northwestern Spain)
vol. 17, pp. 378-385 (online: 14 December 2024)
Research Articles
Emerging pests and diseases threaten Eucalyptus camaldulensis plantations in Sardinia, Italy
vol. 9, pp. 883-891 (online: 29 June 2016)
Research Articles
Brown rot on nuts of Castanea sativa Mill: an emerging disease and its causal agent
vol. 6, pp. 294-301 (online: 16 July 2013)
Research Articles
Fungal community of necrotic and healthy galls in chestnut trees colonized by Dryocosmus kuriphilus (Hymenoptera, Cynipidae)
vol. 12, pp. 411-417 (online: 13 August 2019)
Research Articles
Temporal development of collar necroses and butt rot in association with ash dieback
vol. 10, pp. 529-536 (online: 05 May 2017)
Research Articles
Lenticel infection in Fraxinus excelsior shoots in the context of ash dieback
vol. 12, pp. 160-165 (online: 04 March 2019)
Research Articles
Natural spread of Verticillium wilt as effective constraint on Ailanthus altissima invasion
vol. 18, pp. 391-398 (online: 22 December 2025)
Review Papers
Increasing resistance and resilience of forests, a case study of Great Britain
vol. 17, pp. 69-79 (online: 21 March 2024)
Short Communications
Local spread of an exotic invader: using remote sensing and spatial analysis to document proliferation of the invasive Asian chestnut gall wasp
vol. 5, pp. 255-261 (online: 24 October 2012)
Research Articles
Essential environmental variables to include in a stratified sampling design for a national-level invasive alien tree survey
vol. 12, pp. 418-426 (online: 01 September 2019)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







