
Recent insights in soil nutrient cycling: perspectives from Pinus and Eucalyptus forest studies around the world
iForest - Biogeosciences and Forestry, Volume 17, Issue 6, Pages 394-404 (2024)
doi: https://doi.org/10.3832/ifor4530-017
Published: Dec 20, 2024 - Copyright © 2024 SISEF
Review Papers
Abstract
Soil nutrient cycling in forest ecosystems is a dynamic process fundamentally influenced by climatic and environmental factors. This review synthesizes studies focusing on nutrient dynamics in forests of Pinus and Eucalyptus species, highlighting the sensitivity of these systems to current climatic extremes. We emphasize that most research has been conducted predominantly in natural forests and plantations of Pinus (77%), with an increasing trend of studies on Pinus in natural environments and Eucalyptus in planted forests. Noteworthy, soil sampling in these studies has been primarily concentrated on the upper 30 cm of soil, where nutrient interactions are most pronounced. The relationship between litter and plant organ nutrients as well as soil fertility has been a significant focus of these studies, along with the role of nitrogen and carbon in response to global change. Also, we noticed the importance of research on water availability in the broader context of nutrient cycling. Our review underscores the necessity for continued research in this field, particularly to support informed management and adaptation strategies for both plantations and natural forests in the face of environmental change.
Keywords
Carbon (C), Nitrogen (N), Phosphorus (P), Natural and Planted Forests, Litter, Plant Nutrient, Soil Solution
Introduction
The multifunctionality of forest ecosystems, defined as “the simultaneous provision of multiple services and functions by landscape to society” ([87], [18], [90], [23]), provides climate regulation, water cycle, waste decomposition, nutrient cycling, and wood production ([135], [4], [18], [23]). The multifunctionality of ecosystems is enhanced by the diversity of forest resources such as litter composition and root exudates, which are found in environments with a variety of plant species ([84], [23]). This diversity can also influence several enzymatic activities related to cycles of nitrogen, phosphorus, carbon, and sulfur ([9], [55]), as well as soil respiration and composition of microbial communities ([23]).
Ecologists have investigated the relationship between biodiversity-related factors and ecosystem functions under human-driven conditions, including land use changes, nitrogen additions, and climate changes ([88], [159], [10], [111], [146], [153]). These studies are important because the ecosystem services are current threatened by climate change, pests and diseases, wildfires, drought, soil compaction due to machinery use, and excessive exploitation due to forest harvesting ([23]).
Forest plantations cover 294 million ha worldwide, accounting for 7% of the global forest area ([39]). The selection of the tree species in plantations and the silvicultural practices aimed at maximum and constant wood production leads to a simplification and homogenization of the forest landscape. This results in a reduction in biodiversity ([110]) and in the complexity of forest ecosystems ([99], [6]), significantly impacting on the potential forest functions ([14], [101]).
Tree species belonging to the genera Pinus and Eucalyptus account for 30% of plantations worldwide. They play a pivotal role in meeting human demands while conserving native forests from logging ([104]). However, their fast growth, along with the canopy closure and the suppression of weeds, may deeply alter soil nutrient cycling and water availability ([127], [101]).
In recent decades, emphasis has been placed on designing forest treatments that mimic natural processes and disturbances to promote and preserve biodiversity, complex dynamics, and ecosystem functioning ([42], [128], [129], [6]). Nonetheless, monocultures are still prevalent in plantations and are often characterized by the introduction of non-native, fast-growing species, with short rotation periods and intensive harvesting, which can substantially alter the physical, chemical, and biological attributes of the soil. On the other hand, the establishment of Pinus and Eucalyptus plantations may act as a bulwark against soil erosion in many sites, also offering habitats for local wildlife, enhancing the aesthetic value of landscapes, and potentially restoring hydrological services on former denuded lands ([16], [8]). Evidence has been reported that Pinus and Eucalyptus plantations can locally provide more benefits than harms in terms of delivering ecosystems services ([149]). Further, the ability of such plantations to sequester significant amounts of carbon (C) due to rapid growth represents an important contribution to carbon dioxide mitigation efforts ([16]). Indeed, this forest C pool has been used in ecosystem models for developing climate mitigation policies ([154]).
Substantial evidence indicates that afforestation of previously arable land with planted forests has led to marked increases in total nitrogen (TN), total phosphorus (TP), and soil organic carbon (SOC), thus enhancing soil quality ([117]). While forest plantations may provide fewer ecosystem services than natural ecosystems, their benefits still outweigh those of leaving land unplanted ([25]). Such services, including soil aeration, water permeability, and fertility, are intimately linked to the integrity of soil physical properties ([15]).
This review aims to consolidate research findings on soil nutrient cycling in both natural and plantation forests of Pinus and Eucalyptus. It seeks to answer the following questions: (i) has there been an increase in studies on these topics in recent years? (ii) Has more research on nutrient cycling been conducted in natural forests or in plantations? (iii) Which elements (ions) have been primarily investigated? (iv) What factors have triggered the most significant changes or concerns regarding nutrient cycling?
We collected information to shed light on the significance and management implications of soil nutrients in these environments and summarize advances, trends, and gaps to guide future research in this field.
Material and methods
We searched the existing literature for studies focused on nutrient cycling in Pinus and Eucalyptus forests worldwide. The literature review (Fig. 1) was conducted according to the PRISMA methodology ([96]). Data were sourced from the Web of Science™ (WoS) and Scopus™ databases. The terms “Soil Nutrient Cycling”, “Forest”, “Soil water”, and “Pine” or “Pinus” or “Eucalypt” or “Eucalyptus” were searched in the paper title, abstract, author keywords, and Keywords Plus, focusing exclusively on research articles. This search yielded 152 articles from Scopus and 446 from WoS, including 132 duplicates that were removed, resulting in a corpus of 316 articles.
Fig. 1 - Workflow description of the PRISMA searching methodology, adapted from Moher et al. ([96]).
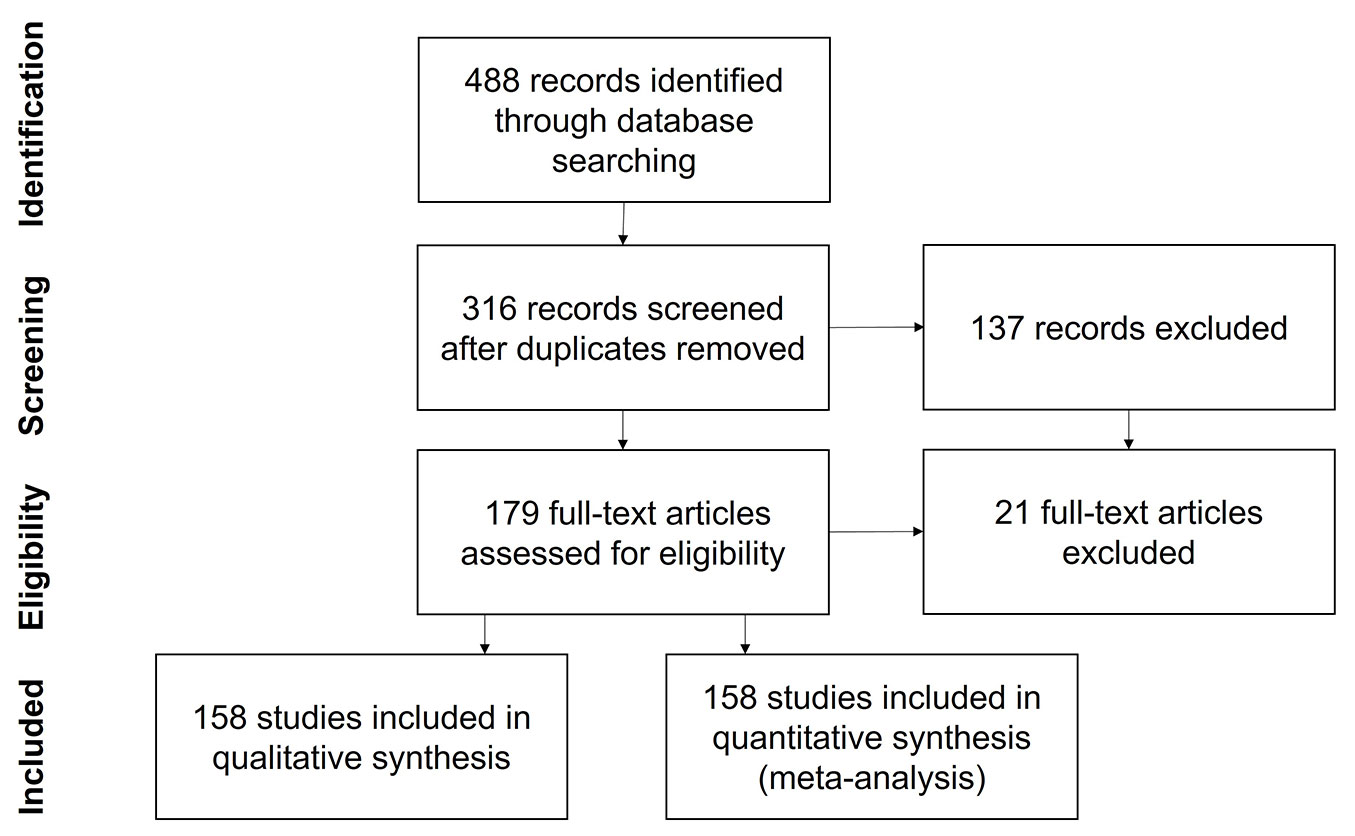
We carefully scrutinized these papers and discarded those that did not match the following criteria/topics: (i) review articles, solely analyzed; (ii) wood volume and production; (iii) production and mortality of fine roots; (iv) soil water use and physical properties; (v) rainfall partitioning and groundwater recharge; (vi) soil communities, including fungal and microbial presence; (vii) distribution and diversity of tree species. We retained 158 articles that specifically addressed various aspects of the nutrient cycle in forests of Eucalyptus or Pinus for further in-depth review.
We employed descriptive statistics using R v. 4.3.1 and RStudio v. 2024.04.2 to synthesize the data. The analysis provided an overview of trends in publication frequency and countries of study. Additionally, we examined the soil depth of the studies, the primary species of the genera investigated, and emerging topics of interest. Furthermore, we meticulously evaluated, extracted, and analyzed pertinent information from the selected 158 papers to comprehend the focal treatment applied (TR), the primary analyses undertaken (AN), and the main subject of analysis (SA). We established three categorical variables (TR, AN, SA) to facilitate the data organization according to the main genera studied - Pinus and Eucalyptus.
Results
Soil nutrient cycling studies in Pinus and Eucalyptus
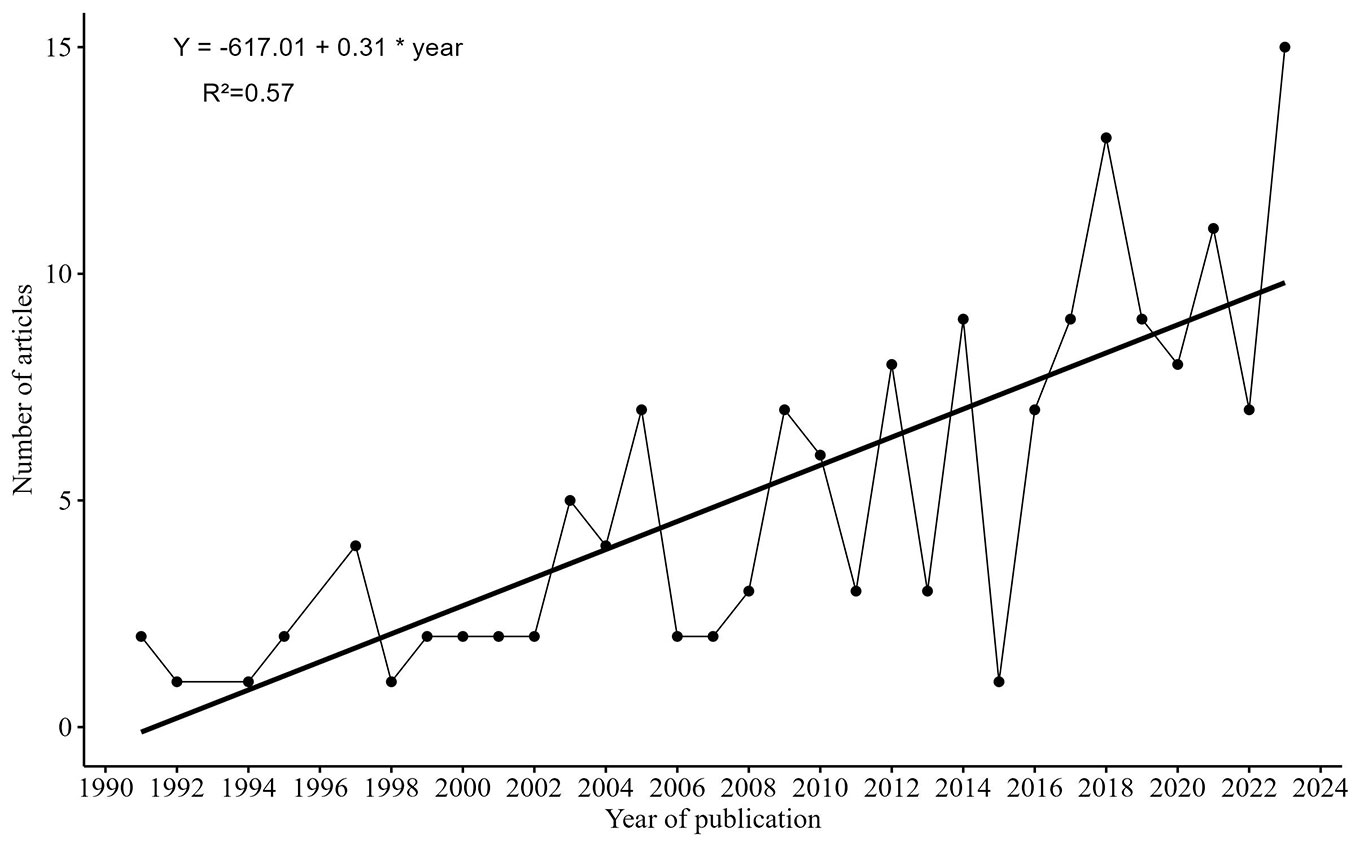
The first study addressing the nutrient cycle in Eucalyptus or Pinus forests was published in 1991. Since then up to 2023, 158 papers were published in this field, with an average of 4.8 publications per year, from one (1991) to 15 (2023) publications per year, showing an obvious growing trend (Fig. 2). Although 58% of the publications were concentrated in the last ten years (2013-2023), this upsurge underscores a growing interest in understanding nutrient and ion dynamics in soil, particularly in response to extreme weather events such as droughts and floods. Valduga et al. ([137]) reviewed articles about the ecological effects of non-natives trees in Brazil such as Pinus and Eucalyptus from 1992 to 2012, finding that more than 80% of papers were published between 2005 and 2012. This period saw a surge in studies about Pinus and Eucalyptus, which were followed by the increase observed in this review about the soil nutrient cycling of these genera.
Fig. 2 - Number of articles per year.
The retrieved studies span various environments: 44% took place in natural forests (8% in Eucalyptus and 35% in Pinus), 47% in plantations (12% in Eucalyptus and 34% in Pinus), and the remaining 8% covered both forest types. Further, 77% of the studies were conducted in Pinus forests and 23% in Eucalyptus, similar to the results found by Pinheiro et al. ([107]), with 92 papers on Pinus and 32 of Eucalyptus in their systematic review about proteomics involved in stress responses. However, it is notable that Eucalyptus-related studies predominantly occurred in plantation settings (64%), while Pinus studies were more common in natural environments (53%).
The United States of America is the leading country in terms of publications contributing 25% with 39 documents, followed by China with 18% (28 documents) and Australia with 11% (18 papers). These three countries accounted for 54% of the total publications, while the remaining 46% were distributed across 23 countries.
As for the thematic focus, the keyword “water” was the most prevalent, appearing in 23% of the documents, followed by “dynamics” and “forest” cited in 22% of papers each, and “nitrogen” in 20%. The high frequency of these subjects suggests a research effort focused on understanding nitrogen cycles in forests, particularly in relation to water. This focus is likely influenced by the context of climate change and reflects a broader concern for the sustainability of forest ecosystems.
Soil sample depths
The analysis of the experimental data revealed the adoption of a common methodology concentrated on the upper layer of the soil for physicochemical analyses. Among the studies reviewed, 24% (38 documents) sampled only the top 10 cm of soil and 47% (74 documents) sampled the top 20 cm, and over half of investigations (58% - 92 studies) focused on the topmost 30 cm of the soil profile.
Studied species
A key aspect of this review concerns the genera Pinus and Eucalyptus. Notably, 77% of the assessed studies pertained to Pinus species. Of the 122 articles examining Pinus sp., 27 featured Pinus sylvestris (22%), which was the more studied species. Although Pinus sylvestris is not a typical planted commercial species, it is prevalent in natural boreal forests, particularly within the European boreal coniferous forests predominantly composed of Pinus sylvestris (Scots pine) and Picea abies (Norway spruce). Due to their adaptability, Pinus sylvestris stands are more widespread in less fertile soils ([19], [35], [103]).
Pinus taeda, native to Southeast America, was the second most studied pine species, comprising 15% of research. It plays a vital role in subtropical forestry worldwide ([62], [106]) and has been a cornerstone of the timber industry in the Southeastern USA for over a century ([5], [40], [75], [92], [120]). Its cultivation extends to China, where it is recognized as a fast-growing timber species ([157]), and to Brazil, where it stands out among the 1.7 million ha of Pinus spp. and the 8.61 million ha of planted forests in the country ([71]).
Regarding Eucalyptus, the species primarily used in afforestation include Eucalyptus urophylla S.T. Blake, Eucalyptus grandis Hill, and Eucalyptus dunnii Maiden ([78], [156], [164]). The research corpus reveals a focus on the Eucalyptus urophylla-grandis hybrid - resulting from the cross-breeding of Eucalyptus urophylla and Eucalyptus grandis - as the most recent subject of nutrient cycling studies. Indeed, it is present in 7 out of 38 papers that studied Eucalyptus spp. (18%). This hybrid is extensively utilized in commercial forestry, reflecting its significance and relationship with the type of environment (planted forests).
Research subjects
We evaluated the subjects addressed in each study regarding the treatment (TR) applied (Fig. 3), and found that a primary focus was on comparing forest types and soil/land uses, which appeared in 34% of the papers (53 out of 158). These comparisons often involved alien vs. native species or contrasted species within the same genus or habitat. The second most prevalent subject was management (19%), which involves silvicultural activities of site preparation, including herbicide use and scalping, stand densities, planting or forest arrangement techniques such as agroforestry ([100]), harvest residue management ([24]), thinning, litter fall removal ([51], [130]), and prescribed burning ([51]).
Fig. 3 - Number of papers by treatment (TR) applied on research of the paper.
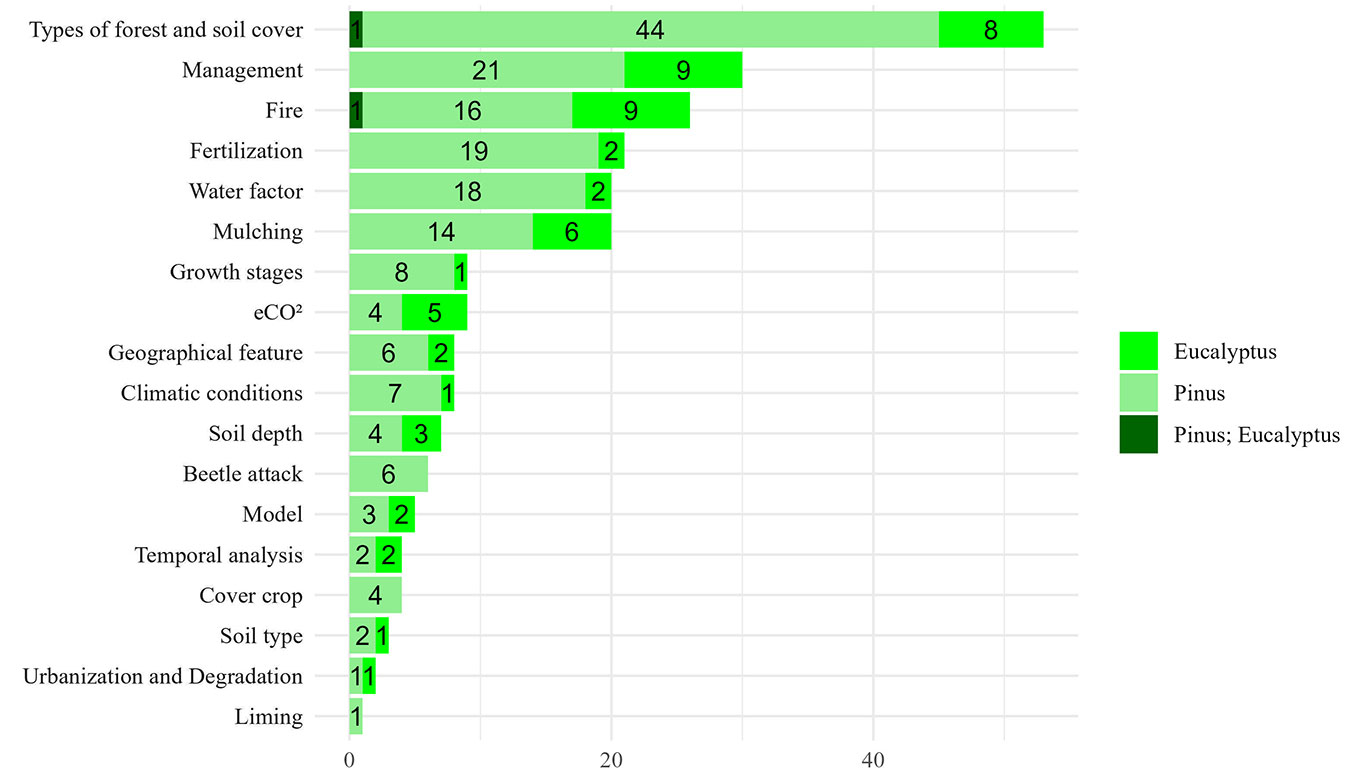
Fire-related studies constituted the third most prevalent subject, accounting for 16% of the research and divided between prescribed burns ([70], [86], [17]) and wildfires ([48], [23], [111]), which have been more explored in recent years.
The impact of climate change on fire incidence has been significant, as fires act as agents that rapidly mineralize and mobilize nutrients ([116]). They are pivotal in the carbon cycle ([21], [112], [126]) by affecting carbon release through soil and microbial respiration ([56], [98], [133]). Additionally, fires alter the composition of soil organic matter ([69], [118]) and disturb the soil litter, upper humus horizons, plant roots and detritus ([139], [125]).
Another significant topic is “Fertilization” (13%), which can refer to synthetic products such as chemical fertilizers ([105], [107]), organic fertilizer such as poultry litter ([76]), or even about natural fertilizers such as the deposition of atmospheric nitrogen ([61], [124]), which have been explored due to ecosystem changes.
Water factor is also one of the most explored subjects (13%). Several studies aimed to investigate the nutrients’ behavior in various environments with different water availability, mostly focusing on drought changes and water limited ecosystems ([91], [53], [63]).
Mulching is another significant subject, which was addressed in 13% of the articles. These studies encompassed a range of activities, from logging and harvest residues ([132]) to the application of biochar ([34]) and manure ([167]), and the comparison of different types of litter and organic residue management ([140]).
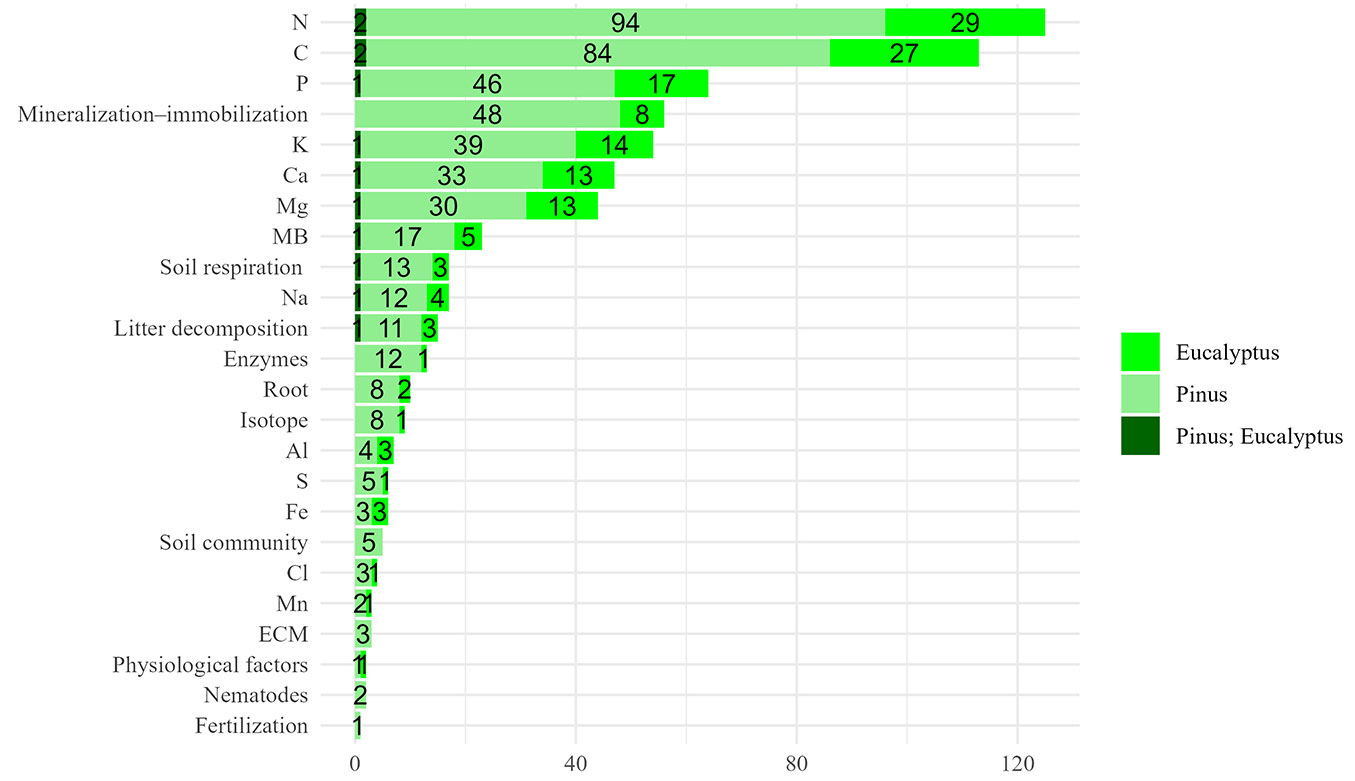
In terms of the main analyses (AN), nitrogen (N) emerged as the most examined variable (Fig. 4), featured in 79% of the studies (125 out of 158). Nitrogen in these ecosystems is primarily derived from biological fixation and atmospheric deposition, predominantly in organic forms ([154]). Carbon was the second most studied variable, present in 72% of the papers (113 out of 158). It is susceptible to climate shifts and essential to plant photosynthesis and respiration processes ([7], [154]). Phosphorus (P), primarily sourced from the weathering of parent material and atmospheric deposition and often present in forms inaccessible to plants, was examined in 41% of the articles (64 out of 158 - [141], [150], [49], [154]). Other significant analyses were about the shift of some nutrients, mostly due to the mineralization process and, to a lesser extent, immobilization as well. This analysis is important because the presence of nutrients in the soil does not necessarily mean that plants can utilize them. Most analyses have focused on inorganic nitrogen, specifically in the forms of ammonium (NH4+) and nitrate (NO3-) ([132], [54], [34], [91]). These studies highlighted complex interactions between C and N cycling, which are associated with litter production and decomposition ([121], [22]).
Fig. 4 - Number of papers by main analyses (AN) of research.
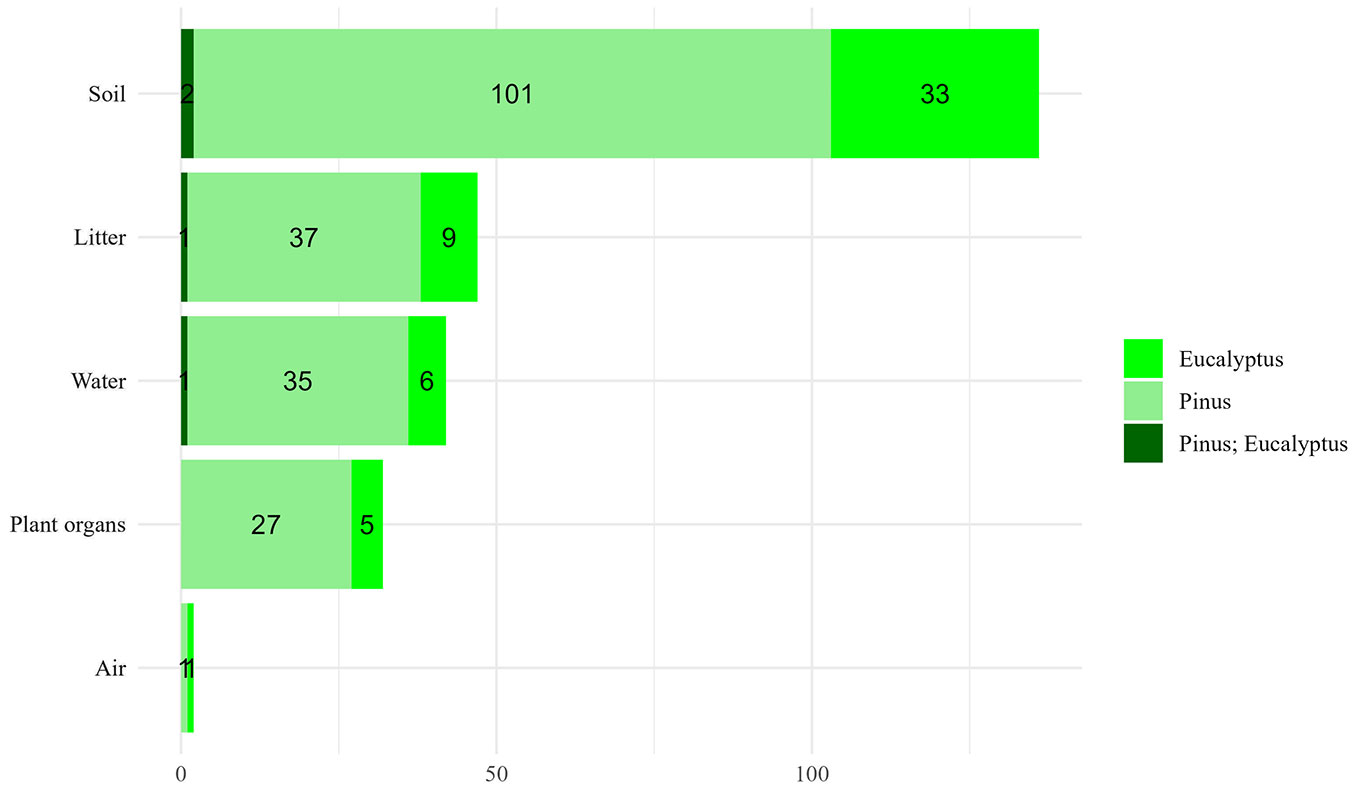
A vast majority of studies (86%) investigated biogeochemical cycle variables in the soil (Fig. 5). Nutrient measurements in forest litter were conducted in 30% of the articles ([47], [108], [136], [2]), with a predominant focus on leaf or needle components. Approximately 27% of the studies examined nutrients in water in the forest ecosystem, such as stemflow, rainfall, throughfall, and outflow ([97], [131]), with most studies concentrating on rainfall ([60]). Additionally, 20% of this research analyzed plant organs such as wood, bark, branches, and roots ([79]). Distinctions between absorptive and transport roots were made to elucidate nutrient pathways ([52], [151], [43]). Only one study assessed nutrients in the atmosphere ([123]).
Fig. 5 - Number of papers by the main subject of analysis (SA) of each paper.
Discussion
In this systematic review, we found a rising number of papers published each year, which may reflect an increasing concern among researchers regarding nutrient cycling. When analyzing the number of publications per country, per genera and per type of forest (natural or planted), a distinct trend emerges. The USA researchers tends to investigate Pinus in natural environments, while Australian scientists are primarily concerned with Eucalyptus in natural forests. Interestingly, China, which is not a primary natural habitat for either Pinus or Eucalyptus, has emerged as the second country with more publications, focused mainly on comparing natural environments and planted forests with these genera.
Research commonly focuses on keywords such as water, dynamics, forest, and nitrogen, reflecting an effort by scientists to understand the nitrogen cycle in forests, particularly in relation to water, which is often affected by extreme climate events. Additionally, the papers analyzed in this review indicate that the upper 30 cm of soil is the preferred layer for sampling, as it provides a more detailed response.
The species more studied were P. sylvestris, P. taeda and E. urophylla-grandis. It is worth to note that the first one is a species frequently found in natural boreal forests, but the second and third species are frequently used in planted commercial forests, which can also provide some ecological services such as soil nutrient cycling.
The main subjects of the reviewed papers show that the majority of studies are aimed to compare different types of forest, especially between natural and monospecific forests. Regarding this aspect, it is well known that the soil nutrient cycling is richer in heterospecific forests compared to plantations due the composition and decomposition of litter. It is desirable that future research in this field should not only focus on management practices like site preparation, harvest residue management, and thinning - since these areas are vital for guiding commercial crops - but also explore less common and less intense techniques such as intercropping and mosaic planting. Investigating these methods could reveal their effectiveness in enhancing plant activities related to nutrient uptake and improving nutrient cycling overall.
Our results stress that climate change, particularly concerning water availability, is a critical issue for future research and the well-being of both natural and commercial forests. Indeed, many studies focused on soil nutrients - specifically nitrogen, carbon, phosphorus, and mineralization - in environments with varying water availability. These studies are essential for understanding how extreme weather events, such as heavy precipitation and droughts, can impact these forests. However, we recommend further research due aimed at disentagling the complex role of water in plant development, physiology, morphology, and anatomy. Additionally, we highlight the need of further investigations on plant plasticity in relation to nutrient availability, uptake, and use, especially given the rapid changes occurring in the environment.
Soil sample depths
In a Masson pine plantation, Justine et al. ([66]) observed a decline in total organic carbon (TOC) from 4.48% to 1.95% and total nitrogen (TN) from 0.32% to 0.14% when comparing the upper (0-30 cm) to the lower (30-60 cm) layers. Similarly, levels of dissolved organic carbon (DOC) and dissolved organic nitrogen (DON) predominated in the upper layer, underscoring its role as a significant reservoir of dissolved organic matter (DOM).
Consistent with these findings, a general decrease in soil carbon, TN, and total phosphorous (TP) with increasing depth was observed in a different-aged Masson pine forest plantation in Southwest China ([158]). In a temperate coniferous rain forest at the H.J. Andrews Experimental Forest in the Cascade Mountains of west-central Oregon, USA, the mineral topsoil (0-2 cm and 0-10 cm, respectively) is the primary source of DOC and DON ([155], [140]).
In a detailed study by Versini et al. ([140]), six soil depths (2, 15, 50, 100, 200, and 400 cm) have been studied, finding that nitrate (NO3) concentrations were anomalously higher at 15 cm compared to 2 cm depth, diverging from the trend seen with other elements where calcium (Ca) and NH4+ fluxes decreased within the 0-15 cm layer.
Maxwell et al. ([91]) examined soil microorganism activity and nutrient availability down to a 90 cm depth. Their findings highlighted a link between soil microorganism activity and carbon and nitrogen levels, gauged by extracellular enzyme activities (EEA), which were notable at an intermediate depth of 15-30 cm. Additionally, they found that increased water availability amplified phosphorus-related EEA in the top 30 cm of soil.
Caldeira et al. ([20]) examined the chemical composition of soil solutions across a profile extending from 0 to 300 cm. They observed a general decline in cation and anion concentrations from the forest floor to the 15 cm depth, except for Mg2+ and NO3 in clay soils and SO2-4 in sandy soils, which peaked at 15 cm depth.
It has been commonly observed that the concentration of organic C, TN, TP, available N, and P tends to decrease with increasing soil depth ([32], [37]). This trend is likely due to the contributions from surface litter decomposition, animal remains, and feces ([154]).
Our analysis indicates that sampling within the upper 30 cm of soil is generally adequate for studies focused on soil nutrient composition and microbial activity, particularly in the top 15 cm, which is more affected by water, biological activity, and litter. However, for investigations into nutrient leaching and soil flux drainage, it is recommended to sample deeper, potentially reaching up to 100 cm.
Soil nutrients
The soil carbon, nitrogen, and phosphorus stoichiometry is a critical indicator of the dynamic changes in soil mineral elements and has been widely used in studies to ascertain the interactions and balance among these elements ([38], [166], [163]).
Soil N originates primarily from biological N fixation and atmospheric deposition, and it is found mainly in organic form ([154]). However, it became available to plants in the inorganic form, which is available after decomposition of the organic compounds by microorganisms ([93], [13], [44], [154]). Soil phosphorus (P) primarily originates from mineral weathering and atmospheric deposition, and it exists in both organic and inorganic forms. However, most of these forms are unavailable for plant uptake due to their organic nature ([154]). As a result, the availability of certain nutrients is influenced by soil microbial activity.
Soil stoichiometry is affected not only by soil fertility but also by the ecological succession. Zhang et al. ([163]) noted that shifts in the C:N:P ratio and microbial biomass correlate with the age of restoration efforts. This correlation highlights the importance of monitoring these ratios as potential indicators of soil P limitations and N supply adequacy ([130], [115], [114]). Temporal changes in soil stoichiometry can significantly alter soil organic matter quality, thereby affecting the rates of crucial ecosystem functions ([84]). For example, a low C:N ratio can stimulate organic matter mineralization, whereas a high C:N ratio could lead to nutrient immobilization ([68], [84]). Baruch et al. ([8]) found that native forest soils exhibit significantly higher carbon and mineral content compared to Pinus caribaea plantations, yet interestingly, there was no discernible difference in soil C:N and N:P ratios between the two vegetation types.
A comparative analysis of soil use indicates that pine plantations may possess superior soil properties over farmlands, with notable improvements in soil organic carbon, total nitrogen, available phosphorus, and microbial biomass, which tend to increase with forest age ([163]). However, Yin et al. ([158]) reported a fluctuating trend in soil carbon, total nitrogen, total potassium, and total phosphorus in P. massoniana plantations, with available phosphorus increasing and available nitrogen initially decreasing but later increasing with plantation age. This can be explained by the mineralization process (i.e., the transformation of total to available N) due to forest ageing. This study also highlighted a positive correlation between total nitrogen and available potassium and a negative one between available phosphorus and soil water content, indicating the complex interplay between these elements.
While forest age is a significant factor influencing nutrient availability and mineralization, soil biological activity is equally crucial in organic matter metabolism, thereby impacting nutrient cycling ([46], [161]). Maxwell et al. ([91]) demonstrated that increased water availability could enhance phosphorus-related extracellular enzyme activity, which plays a pivotal role in the soil nutrient mineralization.
According to Liu et al. ([81]), soil available phosphorus in Pinus sylvestris plantations was substantially higher in wetter years compared to drier ones. This finding aligns with other studies indicating that both soil net nitrogen mineralization rates and phosphorus availability tend to be higher in wetter conditions ([168], [143]).
Coniferous forests have significantly higher concentrations of nitrate and sulfate compared to beech forests, likely due to higher throughfall N and S fluxes. The higher concentrations may affect the mineral soil cation exchange complex and could potentially lead to greater leaching of macronutrients ([31], [82], [12]). The study by Mareschal et al. ([89]) indicated that net nitrification rates in Eucalyptus stands were highest in younger forests and reduced in older ones, with rates significantly higher than those in local natural forests (Savannas).
Lastly, nitrogen fertilization has been shown to increase leaf nitrogen concentrations, reduce nitrogen resorption efficiency, and significantly increase soil NO3-, NH4+, inorganic N, and available P levels. However, these changes in soil chemistry do not appear to have a direct relationship with growth, as measured by stem diameter increment, of Pinus sylvestris plantations ([160], [81]).
Litter
Litterfall can affect soil fertility, soil community composition and nutrient uptake of succeeding plants, and plays a significant role in mediating the relationships between biodiversity and ecosystem functions ([111], [165]).
The process of litter decomposition, which plays a crucial role in nutrient resorption, is primarily influenced by litter chemistry ([81]). A comparative analysis of litter from pine plantations and native forests by Baruch et al. ([8]) revealed a significantly higher litter mass in the pine (Pinus spp.) plantations. However, the nutrient content (N, P, Ca, K) was found to be 3-10 times lower than that in the native forest soil. Despite lower nutrient content, the carbon storage in pine plantations can exceed that of other forest types due to the high biomass yield from fast pine growth and the slow decomposition of pine litter. In their study, Baruch et al. ([8]) recorded a 39% higher carbon storage in pine plantations compared to a lower montane cloud forest.
Achilles et al. ([1]) found that foliar litter fall was the predominant mechanism for the return of base cations to the topsoil. In a European beech forest, the return of base cations through litter fall was nearly twice that of the base cation deposition from throughfall and stemflow. However, in coniferous forests, these inputs were of the same magnitude. This indicates that native forests, with faster litter decomposition, may provide superior soil nutrient enrichment compared to pine plantations, where low litter quality and slower decomposition cause difficulty in nutrient cycling ([8]). However, this process, even when slow, is important because they are potentially the most important in mediating soil functioning during ecosystem recovery ([80], [111]).
Environment factors
The analysis of the reviewed papers allowed us to identify two main groups of subjects: the first is related to anthropogenic activities, such as management, fire (prescribed and due to illegal activity), fertilization, and mulching, whereas the second group of subjects is related to “natural events”, which are intensified and/or accelerated due anthropogenic activities such as climatic conditions, the growth stage of plants, fire (wildfire), and water factors.
We noted a major concern about the future of plant behavior and the characteristics of the environment due to changes caused by these accelerated events, such as the forest C cycle, which is sensitive to a higher soil temperature and changing of climate ([45], [162], [154]), and the nutrient cycling process, which is sensitive to changes in temperature and precipitation ([122], [3], [11], [154]), affecting the amount of soil available N and P ([58], [154]).
Wildfires became more extreme and uncontrollable with regime shifts, and are regarded as global drivers, pumping large amounts of greenhouse gases into the atmosphere ([142]). These events are expected to devastate ecosystems and communities ([28], [29], [111]). Soil water content, which is also affected by the extreme events, influences nutrient concentration, availability, migration, and the uptake ability of plant roots through soil microbial activity, aeration, and temperature ([77], [94], [64]).
Plant organs
Nitrogen is one of the nutrients more evaluated in green leaves or needles. The foliar total nitrogen mass is positively correlated with foliar N isotope composition (δ15N) and is an indicator of plant productivity ([152], [148], [67], [59], [86]). N availability influences the photosynthetic capacity of leaves because the Calvin cycle proteins and thylakoids contain most of the leaf N ([152], [148], [57]). N is also the main component of the Rubisco enzyme involved in photosynthesis ([36], [86]).
Liu et al. ([81]) hypothesized that decreased precipitation would decrease needle nutrient concentrations. Lu & Han ([83]) demonstrated that water addition increased N concentration in green leaves in an experiment with simulated precipitation. Many other studies corroborated the positive correlation between soil moisture and plants N and P uptake ([30], [147], [119], [81]), and nutrient concentration ([113], [102]). Indeed, low precipitation can lead to the reduction of mass flow and nutrient diffusion in soil ([26], [73]), reducing the activity of fine roots ([74]) and the nutrient uptake ([81]).
On the other hand, Liu et al. ([81]) found no correlations between needle N and P concentrations and soil available water in a four-year study, and similar results were obtained by Luo et al. ([85]) and Minoletti & Boerner ([95]). This inconsistency can be attributed to a decrease in precipitation, which limits mass flow of nutrients and their diffusivity in the soil. Such reduction can lead to low concentrations of nutrients in green needle or leaves, regardless of nutrient availability in the soil ([33], [134]). Additionally, it can be explained by the fact that foliar nitrogen (N) and phosphorus (P) concentrations are primarily driven by the plant demands rather than the availability of nutrients in the soil ([109]).
Fine roots play a key role in nutrient uptake and are essential for absorbing soil resources ([52], [151]). Studies show that higher concentrations of nitrogen (N) and phosphorus (P) in the roots can limit plant growth ([41]). Additionally, fine roots may exhibit increased respiration rates and mycorrhizal colonization, which enhances their ability to uptake nutrients ([50], [138], [144], [41], [43]).
Several studies indicate that nitrogen (N) addition enhances the availability of inorganic soil nitrogen and increases root nitrogen concentration (RNC) and the nitrogen-to-phosphorus (N:P) ratio ([65], [72]). Additionally, Geng & Jin ([43]) found a positive correlation between N addition and root phosphorus (P) concentration, as well as available P, similar to the findings of Wang et al. ([145]). N addition also stimulates P activity, increases P absorption, and helps maintain the balance of N and P in plants. However, the study by Chen et al. ([27]) reported that while N addition increased the carbon-to-nitrogen (C:N) ratio, it resulted in decreased root N and P concentrations.
Conclusion
Nutrient cycling in forest ecosystems emerges as a highly sensitive and complex process, markedly affected by climatic and environmental variables. In the face of current climatic change and extreme weather events, it becomes increasingly essential to elucidate the specific roles, contributions, and limitations of each nutrient across diverse environmental contexts.
This review identified a significant body of research and an increasing trend of published papers over the years. These studies are primarily focused on pine forests, both natural and cultivated, providing useful insights into nutrient dynamics in natural forest and plantations. These studies have frequently focused on the upper soil layer (less than 30 cm), which is often rich in information due to higher biological activity.
We found a trend of Eucalyptus-related studies in plantation settings (64%), while Pinus studies were more common in natural environments (53%). The reviewed papers often compare different types of forests, especially monospecific (plantations) vs. heterospecific (natural), the latter having a richer litter and a better nutrient cycling.
Most research analyzed N, C, P and the mineralization process of nutrients, especially N, with an effort in water dynamics. These investigation are crucial in light of the climate change scenario, particularly changes in water regime, wildfires, and temperature.
The complex role of nutrient availability/deficiency in plant development, physiology, morphology and anatomy, calls for further research on the dynamics of nutrient cycling in forests and on the plasticity of plants to face the extreme events, which can dramatically change the dynamic of nutrient cycling.
The collective findings of these studies are invaluable, particularly in informing the adaptive management of both plantation forests and natural ecosystems. Understanding these complex interactions is crucial for fostering resilience and sustainability in forest management practices in view of the challenges posed by a changing climate.
Acknowledgements
The authors are grateful to the Coordination for the Improvement of Higher Personnel Education (Capes, Brazil) for the financial support (scholarships).
References
CrossRef | Gscholar
CrossRef | Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
Online | Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Renato Marques 0000-0003-3011-6672
Álvaro Luis Pasquetti Berguetti 0000-0001-8020-3200
Janaina Gabriela Larsen 0000-0002-5603-5208
UFPR - Universidade Federal do Paraná, Curitiba, Paraná (Brazil)
UFSCar - Universidade Federal de São Carlos, Sorocaba, São Paulo (Brazil)
Corresponding author
Paper Info
Citation
Castilho Balbinot L, Marques R, Tonello KC, Pasquetti Berguetti ÁL, Larsen JG (2024). Recent insights in soil nutrient cycling: perspectives from Pinus and Eucalyptus forest studies around the world. iForest 17: 394-404. - doi: 10.3832/ifor4530-017
Academic Editor
Daniela Baldantoni
Paper history
Received: Nov 27, 2023
Accepted: Aug 21, 2024
First online: Dec 20, 2024
Publication Date: Dec 31, 2024
Publication Time: 4.03 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2024
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 20154
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 13814
Abstract Page Views: 4070
PDF Downloads: 1924
Citation/Reference Downloads: 13
XML Downloads: 333
Web Metrics
Days since publication: 558
Overall contacts: 20154
Avg. contacts per week: 252.83
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
(No citations were found up to date. Please come back later)
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Soil stoichiometry modulates effects of shrub encroachment on soil carbon concentration and stock in a subalpine grassland
vol. 13, pp. 65-72 (online: 07 February 2020)
Research Articles
Response of soil bacterial communities to nitrogen and phosphorus additions in an age-sequence of subtropical forests
vol. 14, pp. 71-79 (online: 11 February 2021)
Research Articles
Effect of plant species on P cycle-related microorganisms associated with litter decomposition and P soil availability: implications for agroforestry management
vol. 9, pp. 294-302 (online: 05 October 2015)
Research Articles
The manipulation of aboveground litter input affects soil CO2 efflux in a subtropical liquidambar forest in China
vol. 12, pp. 181-186 (online: 10 April 2019)
Research Articles
Effect of exogenous nitrogen and phosphorus inputs on the microbe-soil interaction in the secondary Castanopsis sclerophylla forest in east China
vol. 11, pp. 794-801 (online: 14 December 2018)
Research Articles
The effect of clear-cut age on soil organic carbon and nitrogen indices in Scots pine (Pinus sylvestris L.) stands
vol. 18, pp. 146-153 (online: 09 June 2025)
Research Articles
Effects of tree species, stand age and land-use change on soil carbon and nitrogen stock rates in northwestern Turkey
vol. 9, pp. 165-170 (online: 18 June 2015)
Research Articles
Soil fauna communities and microbial activities response to litter and soil properties under degraded and restored forests of Hyrcania
vol. 14, pp. 490-498 (online: 11 November 2021)
Research Articles
Effects of arbuscular mycorrhizal fungi on microbial activity and nutrient release are sensitive to acid deposition during litter decomposition in a subtropical Cinnamomum camphora forest
vol. 16, pp. 314-324 (online: 13 November 2023)
Research Articles
No home-field advantage in upper Andean tropical forests despite strong differences in site environmental characteristics
vol. 17, pp. 286-294 (online: 27 September 2024)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword






