
Stability analysis and selection of optimal Eucalyptus urophylla × E. grandis families
iForest - Biogeosciences and Forestry, Volume 18, Issue 5, Pages 293-300 (2025)
doi: https://doi.org/10.3832/ifor4476-018
Published: Oct 20, 2025 - Copyright © 2025 SISEF
Research Articles
Abstract
To analyze the stability of growth traits and the chemical properties of wood in 55 full-sib families of Eucalyptus urophylla × Eucalyptus grandis from the reciprocal recurrent selection scheme for Eucalyptus in the Congo, two trials were conducted to compare planting spacing (833 and 2500 stems ha-1). The effect of spacing at 55 months after planting was determined for height, circumference, lignin, cellulose, extractive contents, and the genetic variance component of the syringyl to guaiacyl ratio, using single and multisite linear mixed models. The families’ stability analysis was conducted by evaluating the Pi index of Lin. Results show that increased competition between trees raises the heritability of the chemical properties of wood, except for extractive content, and reduces the heritability of growth traits. Results also showed that the chemical properties of wood are relatively constant and that selection would involve a very slight drop in lignin, cellulose, and extractive content. In the case of a short rotation, plantation spacing alters the mean values of the chemical properties of wood in most families, although some remain stable in both plantation densities. For families with a low Pi index, planting spacing has very little influence on wood quality. This suggests that high-density plantations could be established with no negative impact on wood quality. The interaction between planting spacing and the effects of the main male and female primarily concerns growth traits. The interaction between planting spacing and the family effect tends to affect the chemical properties of wood.
Keywords
Planting Density, Wood Chemical Properties, Genetic Gain, Genotype-by-environment Interaction, Genetic Variance Component
Introduction
When grown in plantations, forest tree species are subject to high spatial and temporal variations, which are sources of genotype-by-environment interaction (G×E). Different environmental conditions can affect the performance of forest trees ([33], [35], [62], [63], [61], [29], [66]). Some genotypes can produce different phenotypes depending on the environment ([47], [56]). This phenomenon is known as phenotypic plasticity. It is particularly relevant today in the face of environmental change.
Global environmental changes, pollution, habitat destruction, and the introduction of invasive species are exerting intense selective pressure on forest populations. Phenotypic plasticity is a potential lifeline for many species, enabling them to adapt rapidly to these new conditions and increase their chances of survival ([39]). By modifying their physiology, morphology, or behavior, organisms can mitigate the adverse effects of a changing environment ([40]). A fundamental tool for studying phenotypic plasticity is the reaction norm, a graph that describes the phenotype of a given genotype as a function of the environment. When reaction norms are plotted for multiple genotypes, the slopes of the curves can be compared to visualize the genetic and environmental effects and the G×E. If the slopes are not parallel, the G×E is significant and the genotypes have different plasticity ([59]).
The establishment of forest plantations, which provide high-quality raw material for the pulp and paper industry, energy products, wood panels, and sawn timber, plays a crucial role in mitigating the deforestation of natural forests. Eucalyptus has been the subject of numerous genetic breeding studies, due to its rapid growth and adaptability to Congo’s climatic and soil conditions. This makes it an important species for reducing deforestation of native species. In general, forest plantations are established with a design that aims to provide optimal conditions for tree growth and maximum productivity, and to manage timber quality with a focus on the desired end products ([38]).
The immediate goal in genetic improvement is to predict the genotypic value of candidates for selection ([3]). This goal is feasible if we can accurately predict genetic value and maintain its stability under different environmental conditions. However, the G×E complicates selection in plant improvement programs by making the expected genetic gains inconsistent ([1]). Testing trees in different environments, therefore, improves our understanding of the plant material we are interested in harvesting.
Wood characteristics can vary in properties and performance for specific applications, influenced by both genetic and environmental factors. However, the precise mechanisms by which these factors contribute to wood variation remain unclear. As a result, studies are being conducted to improve our understanding of how environmental variables, such as planting density, affect wood quality. Plantation spacing is one of the most important factors to consider when planning and implementing forestry projects for timber production. The distance between trees, which represents the useful area, has economic, silvicultural, and technological implications, affecting forest production costs, growth rate, forest management practices, and wood quality ([9], [10], [50], [58]).
Plantation spacing is a limiting factor for tree growth, as it influences the resource consumption per unit area ([21]). Changes in environmental resource availability are known to affect productivity, as reported by Resende et al. ([49]). Each species, clone, or cultivar responds differently to the growing conditions imposed by the environment, such as plantation spacing. For fast-growing species with short rotation cycles, such as those in the genus Eucalyptus, studying wood chemical properties is particularly important due to the higher proportion of juvenile wood in the stem and its various uses ([26], [17], [44]).
Planting density can alter wood chemical properties by affecting tree growth and morphology at different stages ([52], [11]). Studies reporting the influence of spacing on eucalyptus wood chemical properties mainly focus on wood basic density ([2], [23], [27], [12], [46], [42]). Morphological characteristics, rather than wood chemical properties, are most commonly used to assess how planting density affects plant growth. There has been little research on the effect of planting density on wood anatomy and chemical composition ([52], [11], [10], [55]). Because of the significance of forest plantations in the world and the possible use of planting density as a management tool, it is crucial to investigate the response of plantation species to various densities at both the individual and stand levels.
This study aims to select the most stable and productive Eucalyptus urophylla × Eucalyptus grandis families from the Eucalyptus breeding program in Congo. We asked three research questions: (i) What is the effect of planting spacing on key wood chemical properties? (ii) What are the heritability and variance-causal components of growth traits and wood quality stability at the family level? (iii) What is the genetic gain in growth traits and wood quality when the most stable and productive families are selected?
Material and methods
Field experimental data
The study was conducted in the Atlantic coastal zone of the Republic of the Congo (11° 59′ 21″ E, 04° 45′ 51″ S - Fig. 1). Mean annual rainfall is approximately 1200 mm (between the years 2012 and 2022); daily temperature is 25-26 °C in the rainy season (October to May) and 22-23 °C in the dry season (June to September). The average monthly rainfall during the rainy season ranges from 83 to 92 mm, and during the dry season from 1 to 20 mm. Relative humidity is high (85%) with low seasonal variability (2%). The soil is ferralitic and highly desaturated in bases ([37]). We used a 13-female E. urophylla × 8-male E. grandis incomplete factorial mating design to produce 55 full-sib families. The progeny test was installed according to two contrasted stocking densities of 883 and 2500 trees ha-1. For each density, the field experiment was a complete block design with two replications. The total number of trees in this study was 2503 (1002 at a density of 833 trees/ha and 1501 at a density of 2500 trees/ha). All families were fully represented at both stocking densities, with each family having an average of 46 progenies (see Tab. S1, Tab. S2, and Tab. S3 in Supplementary material).
Fig. 1 - Location of the experimental station.
Measured traits
Total tree stem growth (HT) and circumference at breast height (C) were measured 55 months after planting using Vertex V (Haglöf, Långsele, Sweden) and a tape measure, respectively. Wood chemical property traits, including Klason lignin content (LIG), α-cellulose content (CEL), syringil/guaiacyl ratio (SG), and extractive content (EXT), were predicted using NIRS (Near-Infrared Spectroscopy) in diffuse reflectance from spectra, measured on sawdust samples collected at 55 months. The sawdust samples were collected from the tree trunk at 1.30 m above the ground using a drill.
NIRS is an analysis method that uses the near-infrared region of the electromagnetic spectrum (from 780 to 2500 nm). It is an absorption spectroscopy whose principle is based on the absorption of near-infrared radiation by organic matter. Depending on their nature, chemical bonds behave like oscillators constantly vibrating at different frequencies ([6]). The NIRS method is both qualitative, as it allows for identifying a product by its spectral fingerprint, and quantitative, as it enables predicting the content of chemical constituents or the value of physical properties under chemical control associated with a sample.
Before NIRS measurements, the sawdust samples were dried in an air-conditioned room (23 °C) to stabilize the sample moisture content at 12%. Spectral acquisition data were processed using a Tango FT-NIR spectrometer (Bruker, Ettlingen, Germany) and Opus software v. 7.0. To predict the chemical properties, we used existing NIRS models of multispecies of eucalypts for samples from this study ([15], [31], [54]).
Data analysis
To test the effect (α = 0.05) of planting spacing on the growth traits and wood chemical properties, estimates for variance components and genetic parameters were obtained using the Restricted Maximum Likelihood and Best Linear Unbiased Prediction (REML/BLUP). The following linear mixed model analysis was used to analyze families at a single site and across environments. For individual analysis by trial site (eqn. 1):
and for joint analysis (eqn. 2):
where the vectors are: y = phenotypic observation; B = replication effect (assumed as fixed); P = plot effect (assumed as fixed); d = density effect (assumed as fixed); B(d) = replication within density effect (assumed as fixed); other vectors were assumed to be random, M = male effect; F = female effect; M×F = male-by-female interaction effect; d×M = density-by-male interaction effect; d×F = density-by-female interaction effect; d×M×F = density-by-family interaction effect; ε = residuals. X and Z are the incidence matrices of fixed and random effects, respectively.
To assess the effect of planting spacing on heritability, the genetic parameters were calculated using the following classical quantitative genetic formulas. For additive variance (eqn. 3):
with σ²M and σ²F denoting male and female variance; for the dominance variance (eqn. 4):
while for the strict sense heritability (eqn. 5):
and for the dominance proportion (eqn. 6):
The analysis of family stability was performed by calculating the Pi index of Lin ([28]) with the following formula (eqn. 7):
where Yij is the performance of family i in environment j, Mj is the performance of the best performing family in environment j, and n is the number of environments tested.
To determine the genetic gain in growth traits and wood chemical properties when the most stable families are selected, the best linear unbiased predictors (BLUP) related to the total genetic effects were computed by solving the mixed model 3 equations. To estimate the relative genetic gain in each trait in a density trial, the following formula was used ([64] - eqn. 8):
BLUPsel being the BLUP of the selected families using three selection intensities (the selection proportions equal to 5%, 10% and 20%), n is the number of selected families, and Ó¯ is the overall trait mean.
Results
Variability of single-site growth and wood quality traits
Tree height and circumference are greater at density 833 than at 2500 stems ha-1. The difference in average circumference was significant (>8 cm), and the height difference was negligible (≈0.2 m) and not significant. The coefficients of variation for both traits were higher in the high-density plots (Tab. 1). Only the male and male-by-female interaction variances contributed to the genetic variability. However, with a density of 2500 stems ha-1, the main male effect was not apparent. At the same time, we noted a slight decrease in the participation of male-by-female interaction in the genetic variance (Tab. 1). Regarding the chemical properties of wood, the cellulose content, lignin content, and SG ratio were higher in high-density planting, while the extractive content was greater in low-density planting. Cellulose and lignin showed stable variability in both planting densities. The variability of extractive content increased with increasing planting density, while that of SG decreased. The cellulose and lignin content of trees varied very little between families (CV < 10%). The genetic variance components had highly variable trends depending on the wood quality traits considered and planting density (Tab. 1).
Tab. 1 - Trait’s statistical description and estimation of components of the genetic variance. (HT): height; (C): circumference; (CEL): cellulose content; (LIG): lignin content; (EXT): extractives content; (SG): syringil/guaiacyl ratio; (h2): strict sense heritability; (D2): dominance proportion; (833, 2500): stand density (n ha-1).
| Parameters | HT (m) | C (cm) | CEL (%) | LIG (%) | EXT (%) | SG | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 833 | 2500 | 833 | 2500 | 833 | 2500 | 833 | 2500 | 833 | 2500 | 833 | 2500 | |
| Mean | 18.3 | 18.0 | 39.4 | 31.1 | 37.81 | 38.05 | 27.75 | 28.47 | 5.51 | 5.19 | 2.78 | 3.10 |
| SD | 4.4 | 4.7 | 12.1 | 9.9 | 2.16 | 2.40 | 1.84 | 1.80 | 1.57 | 1.75 | 0.56 | 0.49 |
| Min | 2.6 | 7.2 | 5.4 | 11.1 | 29.30 | 24.60 | 21.99 | 22.60 | 1.31 | 0.30 | 0.97 | 0.30 |
| Max | 28.4 | 27.6 | 73.0 | 64.5 | 46.15 | 49.20 | 34.74 | 38.50 | 11.34 | 11.20 | 4.45 | 4.80 |
| CV(%) | 24 | 26 | 31 | 32 | 6 | 6 | 7 | 6 | 28 | 34 | 20 | 16 |
| p-value(family) | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | ||||||
| p-value(family × density) | 0.079 | 0.657 | <0.001 | <0.001 | 5.00e-11 | <0.001 | ||||||
| σ2F | 0.00 | 0.00 | 0.00 | 0.00 | 0.04 | 0.26 | 0.36 | 0.57 | 1.23e-08 | 0.00 | 1.21e-11 | 6.89e-03 |
| σ2M | 2.89 | 0.00 | 16.98 | 0.00 | 0.00 | 0.00 | 1.38 | 0.00 | 0.00 | 0.00 | 2.86e-02 | 0.00 |
| σ2M×F | 0.35 | 1.64 | 7.10 | 6.94 | 0.35 | 0.00 | 1.11 | 5.87e-10 | 0.00 | 0.15 | 5.87e-10 | 1.83e-04 |
| σ2ε | 15.74 | 21.46 | 136.61 | 97.78 | 2.51 | 3.54 | 1.53 | 1.65 | 1.61 | 2.43 | 9.94e-02 | 1.04e-01 |
| h² | 0.25 | 0.00 | 0.17 | 0.00 | 0.02 | 0.13 | 0.36 | 0.40 | 1.53e-08 | 0.00 | 0.16 | 0.11 |
| D² | 0.06 | 0.23 | 0.14 | 0.22 | 0.35 | 0.00 | 0.46 | 8.41e-10 | 0.00 | 0.19 | 0.55 | 6.15e-03 |
The heritability for height and circumference was 0.25 and 0.17, respectively, in the plot with a density of 833 stems ha-1. It was non-existent in the plot with 2500 stems ha-1. At the same time, we noted an apparent increase in the dominance proportion, which ranged from 0.06 to 0.23 and from 0.14 to 0.22, respectively, for height and circumference. For lignin and cellulose content, heritability increased with planting density, while the opposite was observed for extractive content and SG ratio (Tab. 1).
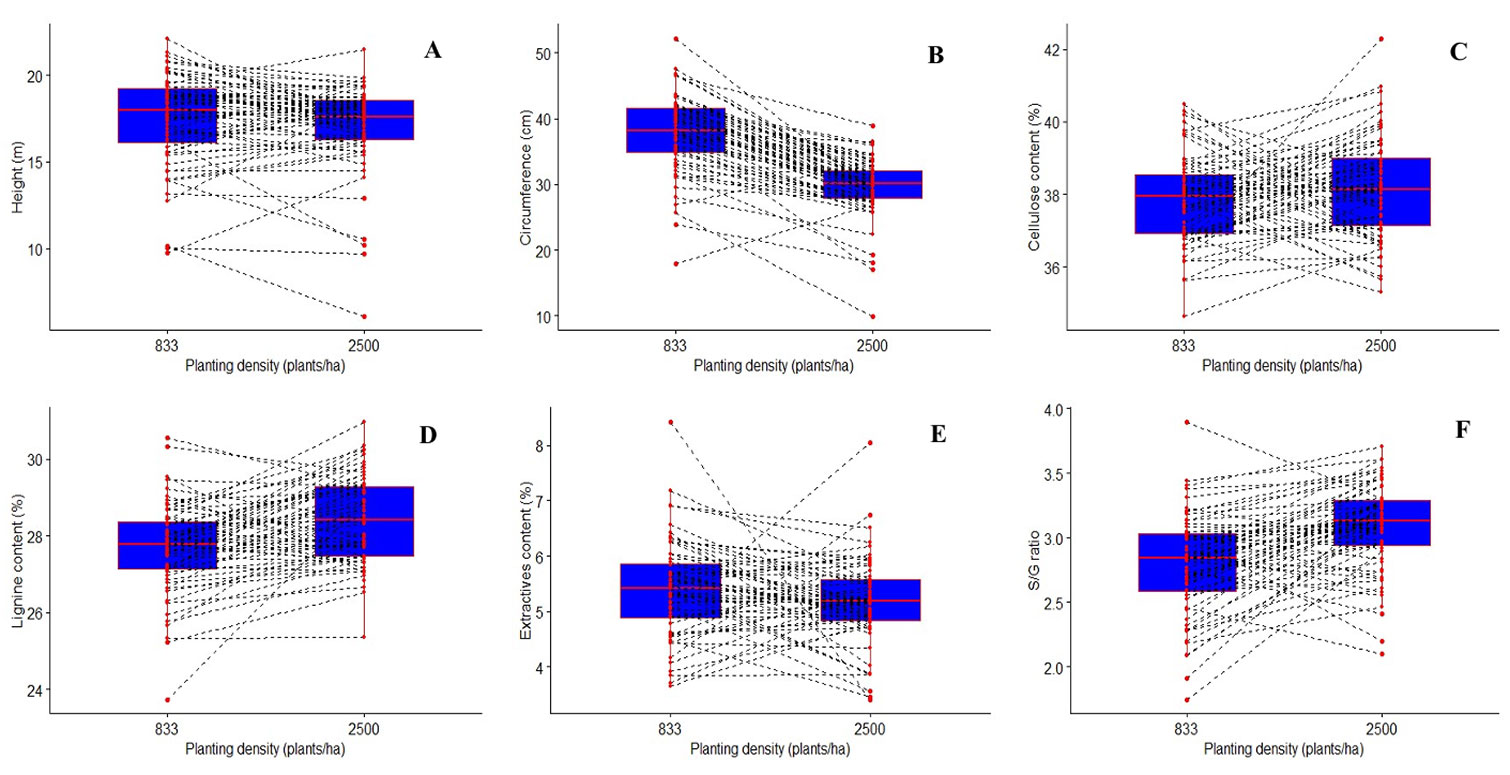
The reaction norms of all the traits studied showed that the performance of most families differs with planting densities. The performance of some families was stable at both plantation densities. Some families performed well, but were unstable at both densities; some performed well and were stable, and some performed poorly, but were stable at both densities (Fig. 2). In other words, some families perform very well at a density of 833 stems ha-1 and not at 2500 stems ha-1 and vice versa; some performed well at both densities, some performed very poorly at both densities, and some families had a mean performance at both densities.
Fig. 2 - Paired boxplots showing reaction norms for growth traits (A, B) and wood chemical properties (C, D, E, F).
Variability in growth and wood quality traits at both densities
Regarding growth, we observed a significant effect of density on circumference (p <0.001), but not on tree height (p = 0.5). For wood quality, the impact of density was significant for all traits. Statistically significant differences were observed for the family effect for growth traits and wood quality (p < 0.001 - Tab. 1). Overall, the density-by-female interaction was apparent in growth traits. For wood chemical properties, it was only evident for extractive content and cellulose content. It did not appear to affect lignin content and SG ratio. This interaction was very pronounced for circumference growth (σ2d×F = 1.41), while it was moderate for height growth (σ2d×F = 0.4) and weak for cellulose content (σ2d×F = 0.14). The interaction was negligible for extractive content (σ2d×F = 8.8e-02) and zero for lignin content and SG ratio.
The density-by-male interaction appears to affect growth traits above all. For wood chemical properties, it was only evident for extractive content, cellulose, and S/G ratio, while it was null for lignin content. Male variance was found to be the component of variance that interacted least with planting density. This interaction was very pronounced for growth traits, with σ2d×M = 2.16 and 0.62 for circumference and height growth, respectively. It was almost null for cellulose content (σ2d×M = 1.48e-08), extractive content (σ2d×M = 8.74e-10), and S/G ratio (σ2d×M = 2.07e-11), and null for lignin content.
The density-by-family interaction is highlighted for all wood chemical properties. It was found to be close to zero for circumference and height.
Family variance is the variance component that interacts the most with planting density. This interaction was very pronounced for lignin (σ2d×F×M = 0.93) and cellulose (σ2d×F×M = 0.82), weak for extractive content (σ2d×F×M = 0.29) and S/G ratio (σ2d×F×M = 0.12), while it was negligible for circumference (σ2d×F×M = 1.54e-06) and zero for height.
Growth and wood quality stability analysis of families using Pi index and selection of optimal families
Tab. S4 (Supplementary material) shows the Lin indexes for the 55 families studied for each trait. We found that families with a low Pi index reflect the stability and good performance of their individuals. However, several families showed a high Pi index, indicating instability and poor performance of the resulting individuals.
The rank held by each family, considering the Pi index for each trait, and the sum of the ranks considering all traits, are presented in Tab. S5 (Supplementary material). It should be noted that some families performed well and were stable across all characteristics. However, some families with the worst performance were also less stable for all the traits.
Expected genetic gains by direct selection for growth and wood property traits were determined according to three selection rates. We noted that genetic gain increased in line with selection intensity. Selecting the best families for growth and SG ratio generated considerable estimated gains. However, selecting the best families for cellulose, lignin, and extractive content generated little gain (Tab. 2).
Tab. 2 - Expected genetic gain (ΔG) of growth and wood chemical properties traits.
| Traits | Proportion of selection | ||
|---|---|---|---|
| 5% | 10% | 20% | |
| Height | 0.14 | 0.13 | 0.11 |
| Circumference | 0.13 | 0.11 | 0.10 |
| Cellulose | 0.005 | 0.004 | 0.003 |
| Lignin | 0.03 | 0.03 | 0.02 |
| Extract | 0.02 | 0.02 | 0.01 |
| SG ratio | 0.10 | 0.08 | 0.07 |
Discussion
Effect of plantation density on genetic variance components
Among growth traits, height was less affected by planting density than circumference. Research has already shown in the Pointe-Noire region that E. urophylla outperforms E. grandis ([8], [35]). In our study, the male additive variance (E. grandis) is the only significant effect expressed for height and circumference at a density of 833 stems ha-1. No specific effect was observed for the same traits with the 2500 planting spacing. On the contrary, a preponderance of female variance was observed by Makouanzi et al. ([33]). The difference between our crossover designs could explain the discrepancy between our results.
Planting density significantly affects the chemical properties of wood, with female variance (E. urophylla) playing a key role. The extractive content is determined by residual variance at both densities. At 833 stems ha-1, female additive variance expresses itself minimally. Environmental factors may explain the production of certain extracts as a defense mechanism, as noted by Favreau ([20]). At 833 stems ha-1, lignin content showed greater male variance, high family variance, and average female variance.
In contrast, at 2500 stems ha-1, female variance significantly increased while family variance decreased, overshadowing male variance. This highlights E. urophylla’s superiority over E. grandis in Pointe-Noire, attributed to E. urophylla’s higher variability upon its introduction to the Republic of the Congo ([7]). This advantage may also stem from the diverse and suitable seedlots initially introduced, as both species exhibit considerable genetic variation across their extensive natural ranges ([19]).
Effect of plantation density on heritability and dominance proportion
Heritability of growth traits and wood chemical properties varies with age ([67], [34]) and environment. We found that, in a less constrained environment, height heritability was low and circumference even lower. Several studies indicate that height heritability generally exceeds that of circumference ([25], [31], [34]). Makouanzi et al. ([35]) studied 64 full-sib families and found a decreasing trend in heritability from 833 stems ha-1 to 2500 stems ha-1. In this study, heritability was negligible in high-density plantations, indicating that these traits, being polygenic, are sensitive to environmental factors ([25]). In constrained environments, genetic interactions increase, with dominance playing a significant role. Dominance effects, especially in hybrid populations, are amplified in marginal areas like Pointe-Noire ([7], [34]).
At a density of 833 stems ha-1, the genetic control of wood chemical properties is low to medium. Mandrou et al. ([36]) found a medium heritability for lignin content and high heritability for the S/G ratio in 33 full-sib families of E. urophylla × E. grandis at 667 stems ha-1. In this study, extractive content was entirely influenced by environmental factors at both densities, a trend also noted in Eucalyptus dunii ([51]).
Interaction between density and genetic variance components
Overall, the interaction between planting density and the principal male and female effects involves growth traits. In contrast, the interaction between planting density and the family effect generally concerns the chemical properties of wood. These results contrast with those obtained by Makouanzi et al. ([35]), who found that the interaction between planting density and the main male and female effects significantly contributed to the chemical properties of wood. In our view, these results largely depend on the structure of the crossover design.
The reaction norms for growth and the chemical properties of wood
The G×E varies among families, with some showing stable family rankings between 833 and 2500 stems ha-1, while others exhibit unstable performance at higher densities. Increased plantation density impacts circumference more than height, and wood chemical properties remain relatively constant. Eucalyptus’s rapid growth heightens competition between individuals ([33]). Previous studies confirm that density primarily affects circumference over height ([16], [8], [45]). Brito et al. ([10]) indicated that 833 stems ha-1 is optimal for eucalyptus growth in terms of height and circumference, consistent with our findings.
Although wood chemical properties appear constant, individual families respond differently to environmental changes, exhibiting non-parallel reaction norms at varying plantation densities, indicating G×E ([35]). In short rotations (55 months), plantation density significantly alters the chemical properties of wood for most families by affecting extractive content. This contrasts with findings from Elerati ([18]) and Brito et al. ([10]), who reported non-significant variations in lignin, cellulose, and extract content with changing density, likely due to their smaller sample sizes (five and three clones, respectively) compared to our 55 families. Additionally, chemical composition in wood is influenced by genetic effects, tree age, and environmental interactions ([41], [52], [32], [33]). Competition and environmental stresses affect the chemical composition of broadleaved trees like eucalyptus. In high-density plantations, trees compete for light, water, and nutrients, which can lead to increased lignin content as a response to strengthen cell structures and enhance mechanical strength ([30]). Consequently, high-density plantations may exhibit higher lignin levels.
Families’ performance and stability
Our results indicate that some families exhibit stable performance in growth and chemical properties, characterized by the lowest sum of Pi ranks. Notably, several stable families share common parents, 9-21 (E. grandis) and 14-142 (E. urophylla), suggesting that the ability to adapt to different plantation spacings may be inherited. Stability might arise from adaptive genes ([22]), and research showed that heterogeneous genetic structures tend to be more stable than homogeneous ones ([24], [57]). Given our study’s single hybrid formula (E. urophylla × E. grandis) and a significant number of families (55), the potential for flexibly adapting to density changes is notable.
Stability in growth and wood quality is crucial for selecting plant material amidst global environmental changes. Lin’s Pi index helps assess performance and stability, with low values indicating stable, high-performing genotypes ([13], [5], [53]). Families with good performance under varying conditions are likely to provide valuable genetic material for developing elite hybrids ([48], [14]). A focused hybridization program can lead to new genetic combinations that enhance biotic tolerance and productivity ([4]). Our findings suggest that selecting high-performing, stable families is beneficial, as phenotypic plasticity allows for adaptation to new conditions and fosters adaptive genetic changes crucial for long-term evolutionary success ([60], [43]).
In short, for families with a low Pi index, planting density has very little influence on wood quality. Therefore, high-density plantations can be established without reducing wood quality. Lasserre et al. ([27]) and Brito et al. ([10]) have reached similar conclusions. This result suggests that wood with good paper quality can be produced with a higher plantation density.
Genetic gains
Our results show that selection might involve a very slight drop in lignin, cellulose, and extractive content. This suggests that the selection process in the E. urophylla × E. grandis improvement program in the Congo is unlikely to have any major effects on lignin, cellulose, and extractive content ([31]). As in hybrid poplar clones ([65]), our results indicate that selecting for growth traits will not significantly reduce wood quality traits. Based on our results, it would be possible to independently choose very productive families, as well as those with a low lignin content (a trait sought by paper makers) or a high lignin content (a trait sought by charcoal makers). However, selection should be carried out in a large population. Numerous parents and families are required to maximize the potential genetic gain. Despite the slight variation in lignin content, this chemical trait displays strong genetic control, suggesting that parental selection plays an important role.
Conclusions
This study shows that the chemical properties of wood are less variable than growth traits. Increased competition increases the heritability of the chemical properties of wood, but decreases the dominance proportion, except for extractive content. Increased competition decreases the heritability of growth traits and increases the proportion of dominant growth traits. Based on these results, we can recommend striking a balance between increased competition, which increases the heritability of wood chemical properties, and reduced heritability of growth traits. By adjusting the plantation spacing, it would be possible to favor a certain level of competition that would allow better expression of the chemical properties of the wood without significantly compromising the heritability of the trees’ growth traits. Family variance is the component that interacts most with planting density. The interaction between planting density and the main male effect, as well as between planting density and the main female effect, primarily concerns the development of growth traits.
In contrast, the interaction between planting density and the family effect concerns the development of the chemical properties of the wood. The plantation density affects growth and wood quality differently. We also found that the families studied respond differently to plantation densification in terms of growth and wood quality.
Acknowledgements
The authors would like to thank the CRDPI engineers for their assistance with field measurements. The authors would also like to thank Gilles Chaix from CIRAD, Montpellier, for the NIRS analysis.
References
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Ursula Armeline Mondzeki Bomouta
Henok-Jubref Baniakissa-Ndelo
ENSAF, École Nationale Supérieure d’Agronomie et de Foresterie, Université Marien Ngouabi, Brazzaville (République du Congo)
École Normale Supérieure, UMNG, BP 69 Brazzaville (République du Congo)
Corresponding author
Paper Info
Citation
Makouanzi Ekomono CG, Gomat H-Y, Mondzeki Bomouta UA, Baniakissa-Ndelo H-J (2025). Stability analysis and selection of optimal Eucalyptus urophylla × E. grandis families. iForest 18: 293-300. - doi: 10.3832/ifor4476-018
Academic Editor
Angelo Rita
Paper history
Received: Sep 23, 2023
Accepted: May 13, 2025
First online: Oct 20, 2025
Publication Date: Oct 31, 2025
Publication Time: 5.33 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2025
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 5105
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 2669
Abstract Page Views: 1466
PDF Downloads: 794
Citation/Reference Downloads: 0
XML Downloads: 176
Web Metrics
Days since publication: 279
Overall contacts: 5105
Avg. contacts per week: 128.08
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
(No citations were found up to date. Please come back later)
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Age trends in genetic parameters for growth and quality traits in Abies alba
vol. 9, pp. 954-959 (online: 07 July 2016)
Research Articles
Genetic variation and heritability estimates of Ulmus minor and Ulmus pumila hybrids for budburst, growth and tolerance to Ophiostoma novo-ulmi
vol. 8, pp. 422-430 (online: 15 December 2014)
Research Articles
Seedling emergence capacity and morphological traits are under strong genetic control in the resin tree Pinus oocarpa
vol. 17, pp. 245-251 (online: 16 August 2024)
Research Articles
Preliminary study on genetic variation of growth traits and wood properties and superior clones selection of Populus ussuriensis Kom.
vol. 12, pp. 459-466 (online: 29 September 2019)
Review Papers
Genetic diversity and forest reproductive material - from seed source selection to planting
vol. 9, pp. 801-812 (online: 13 June 2016)
Research Articles
Patterns of genetic variation in bud flushing of Abies alba populations
vol. 11, pp. 284-290 (online: 13 April 2018)
Research Articles
Interaction between planting spacing and wood properties of Eucalyptus clones grown in short rotation
vol. 14, pp. 12-17 (online: 02 January 2021)
Research Articles
Genetic diversity of core vs. peripheral Norway spruce native populations at a local scale in Slovenia
vol. 11, pp. 104-110 (online: 31 January 2018)
Research Articles
Genetic variation of Fraxinus excelsior half-sib families in response to ash dieback disease following simulated spring frost and summer drought treatments
vol. 9, pp. 12-22 (online: 08 September 2015)
Research Articles
Comparison of genetic parameters between optimal and marginal populations of oriental sweet gum on adaptive traits
vol. 11, pp. 510-516 (online: 18 July 2018)
iForest Database Search
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords







