
Impact of forest management on threatened epiphytic macrolichens: evidence from a Mediterranean mixed oak forest (Italy)
iForest - Biogeosciences and Forestry, Volume 12, Issue 4, Pages 383-388 (2019)
doi: https://doi.org/10.3832/ifor2951-012
Published: Jul 12, 2019 - Copyright © 2019 SISEF
Research Articles
Abstract
Forest management practices may heavily affect epiphytic cryptogams. This study was conceived in March 2016, as soon as we were informed about an authorized logging for timber within a Mediterranean mixed oak forest in Tuscany (central Italy), which threatened a large population of the forest macrolichen Lobaria pulmonaria (L.) Hoffm., composed of hundreds of fertile thalli. Lobaria pulmonaria is often used as an ecological indicator of high quality habitats hosting rare lichens, and in general, cryptogams worthy of conservation. The species has suffered a general decline throughout Europe as a consequence of air pollution and intensive forest management, and currently it is red-listed in several countries, where it is considered also as a “flag species”. We estimated that 40% of the lichen biomass (8.5-12.3 kg ha-1) has been lost due to logging operations (in the core area, up to 1.8 kg every 100 m2), in particular large and fertile thalli. One year after the conclusion of logging operations, the analysis of chlorophyll fluorescence emission (indicator of the photosynthetic performance of the lichen photobionts), revealed a significant reduction of the vitality of the thalli on retained-isolated trees. The article provides issues for conservation in Mediterranean oak forests and outlines the outmost importance of ensuring the safeguard of forest ecosystems hosting fertile populations of this model species, especially in the case of unprotected forests.
Keywords
Biomass Loss, Chlorophyll Fluorescence, Flag Species, Legal Protection, Lobaria pulmonaria, Red Lists
Introduction
Forest management strongly influences biodiversity ([16], [43]) and affects forest structure and dynamics, as well as environmental parameters relevant to dispersal, establishment and maintenance of epiphytic cryptogams ([1], [23]). This fully applies to the epiphytic forest lichen Lobaria pulmonaria Hoffm., which has suffered a general decline throughout Europe as a consequence of global change and forestry ([29], [26]), whose effects are expected to be further exacerbated by climate change ([26]). Its occurrence and abundance are used as indicators of forest ecological continuity ([34], [21]) and for mapping forest sites worthy of conservation ([5], [3], [22], [4]). Currently, L. pulmonaria is a red-listed species in several European countries ([39], [17]).
Forest management is associated with forest fragmentation and trees exploitation that cause a complete break in the availability of the primary habitat for epiphytic lichens ([29], [47]), strongly impacting the most sensitive species such as L. pulmonaria, whose distribution is limited by its dispersal capacity and the presence of suitable (high-quality) habitats ([28], [15]).
The negative effects of silviculture on L. pulmonaria were early reported at the beginning of the last century ([19]) and outstanding works summarize the interactions between different forest management practices and L. pulmonaria populations ([40], [1], [23], [4]). Intensive forest management may threaten L. pulmonaria, and in general sensitive forest lichens, causing habitat fragmentation, degradation and loss, that impact local populations structure, size and dynamics ([40], [2]). Logging may deplete species with poor dispersal ability (such as L. pulmonaria), which would remain restricted to the non-disturbed parts of the forest landscapes ([40]). Moreover, after logging epiphytic lichens are exposed to a sudden increase of sun irradiance and dry conditions, which, if in excess of the ecological range, may injure photosynthetic performances, and hence thallus vitality ([11], [12], [13], [18]). These aspects can be exacerbated in potentially arid environments, such as the Mediterranean region, where beech and oak-dominated forests are the main habitats for Lobaria populations ([36]). The impact of forestry on the abundance and fertility of L. pulmonaria populations has been assessed in boreal forests ([35], [9]). Nevertheless, the impact of logging on L. pulmonaria populations in the Mediterranean region has not been clearly quantified (i.e., measured as biomass loss).
In March 2016, a large population of L. pulmonaria including hundreds of fertile thalli has been depleted and threatened by a legal logging for timber within a Mediterranean oak wood forest of central Italy. The damaged stand is a local hotspot for this model species. In this site, L. pulmonaria has a patchy distribution, colonizing several tree species (more than 1000 phorophytes): mostly Quercus ilex, Quercus cerris, Quercus pubescens, Arbutus unedo, Sorbus torminalis, Crataegus monogyna, Fraxinus ornus and even the bark of Hedera helix growing on these trees. Scattered thalli may also occur on fallen and standing deadwood and on soil-dwelling mosses. In fact, large stands with rich populations are core areas for several rare lichens and have conservation priority due to a higher genetic variability and evolutionary potential compared to small-sized stands ([33], [22]). In parallel with the cutting, we saved as much fresh material as possible to be used for conservation and experimental purposes; we estimated the overall distribution of the population and quantified the loss of biomass and fertile thalli, whose development may require up to 35 years in forest habitats under stable ecological conditions ([38]). We addressed the following questions: (i) which is the impact of logging on the model species within a Mediterranean mixed oak forest? (ii) do logging operations affect the vitality of the thalli growing on retained-isolated trees? Lastly, (iii) moving from local to broad-scale the study provides suggestions for the conservation of threatened macrolichens in Mediterranean oak forests and an overview on the status of L. pulmonaria (inclusion in red lists and legal protection) in Europe.
Materials and methods
Study site and experimental constraints
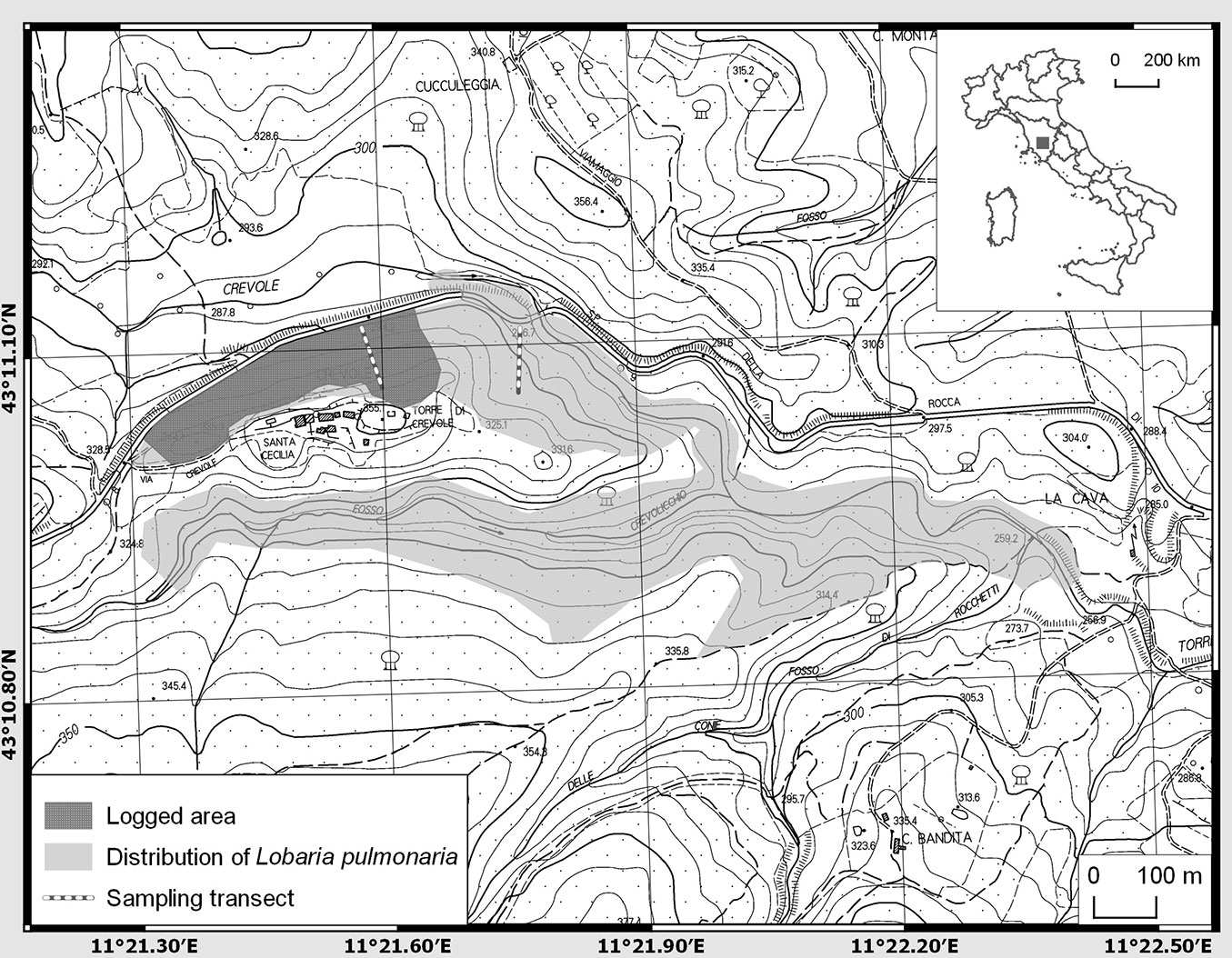
The study was conceived in March 2016 as soon as we became aware of a legal logging for timber in a Mediterranean mixed oak forest stand of 4.4 ha in Tuscany, central Italy (Fig. 1), that was previously recognized as a local hotspot of L. pulmonaria ([30]). The stand extends for about 500 m according with E-W direction on a hillside with a N slope. In the area, oak wood forests are generally managed by a coppice system with standards and rotation cycles of about 18-20 years. However, some stands with low management intensity have a longer logging cycle, which dates back to more than 40 years ago, as in our case. Upon our request, local authorities temporarily suspended the cutting for a few days (about 1 ha of the stand was already cut at that moment). This interruption offered the opportunity to investigate in the field the damage occurred to forest macrolichens, with a focus on the model species. Thanks to the cooperation with local authorities and woodcutters, the subsequent logging activity was addressed to an uneven-aged coppice system, allowing the preservation of several trees with L. pulmonaria (hereafter Lobaria-trees) and also some patches (islands) with groups of uneven-aged trees ([10]). In order to assess the impact of logging, the logged stand was compared with an adjacent unlogged area by means of sampling transects (Fig. 1). Both sites are located along a narrow valley, with similar orientation (N), soil type, tree age and composition, moisture and distance from the closest stream.
Fig. 1 - Map of the study area (Central Tuscany, Italy).
Assessment of the loss of L. pulmonaria
Occurrence
We estimated the occurrence of L. pulmonaria on the basis of a strip adaptive cluster sampling accounting for the spatial clustering of the lichen population ([44], [32], [45]). We surveyed 20 adjacent plots (5×5 m) along two randomly selected transects (Fig. 1), one being established in the logged stand and one in the neighbouring unlogged stand. When L. pulmonaria was found within a 5×5 m quadrat, two secondary adjacent quadrats, at the eastern and western sides, were adaptively included into the transect.
Within each plot, we recorded the number of colonized and not colonized trees, the circumference of each individual tree, the number of L. pulmonaria thalli as well as their size and aspect on the trunk, and the presence of structures for vegetative and sexual reproduction. For practical reasons, measurements were gathered up to 2 m for all stems with L. pulmonaria thalli. We also recorded the occurrence of additional epiphytic species of conservation concern ([24]). Nomenclature of lichens follows Nimis ([27]).
Biomass
In order to link thallus size to biomass of L. pulmonaria, we assessed the relationship between lichen surface area (cm2) and weight (mg) based on 100 randomly selected air dried thalli. By means of this procedure we translated the estimated surfaces of the thalli into lichen biomass.
For an estimation of the potential loss of L. pulmonaria biomass in the field, three 100 m2 (10×10 m) plots were randomly selected in the core area (the first logged sector, extensively colonized): in each plot a group of four volunteers carefully collected for 30 minutes as much material as possible from the logged trees as well as from the ground. To check the collection efficiency, a secondary 4 m2 (2×2 m) plot was randomly selected within each 100 m2 plot and volunteers retrieved the remaining fragments of the thalli until all the material remained was removed from the plot. This step (limited in space and time, since it was carried out during the short interruption of logging operations) allowed an estimate of the overall biomass of L. pulmonaria within any randomly selected 100 m2 plot in the core area. Since these plots were mainly located within the first 25 m from the forest edge, for a better estimation of the biomass of L. pulmonaria over the whole area, in a second step we harvested fallen thalli and measured those still standing after the cutting also from 16 square plots (4 m2) located at intervals of 25 m over a regular grid with the origin randomly selected.
Assessment of the vitality of the thalli
During April 2016 and in May-June 2017 (after the conclusion of logging operations), the vitality of the macrolichen L. pulmonaria was assessed through the analysis of chlorophyll (Chl) a fluorescence emission, measuring the potential quantum yield of primary photochemistry (FV/FM), where FV = (Fm - F0) is the variable fluorescence and F0 and Fm are minimum and maximum Chl a fluorescence. In the field, thalli were kept hydrated (sprayed with mineral water) and dark adapted overnight (covered with a black velvet cloth) before the measurements. Thalli were then lightened for one second with a saturating (up to 3000 μmol of photosynthetically active photons m-2.s-1) light pulse and fluorescence emission was recorded for one second. Measurements were carried out with a Plant Efficiency Analyzer fluorimeter (Handy PEA®, Hansatech Ltd, Norfolk, UK) on thalli of L. pulmonaria preserved in the logged area, on those occurring in the adjacent unlogged area and on those retrieved from the ground. Up to 40 thalli were analyzed for each condition and 3-5 replicates were recorded on each thallus, according to its size, in order to minimize error estimates owing to intrathalline variability. Furthermore, we evaluated the condition of the thalli on the basis of signs of discoloration and bleaching and the presence of necrotic parts. Thalli were attributed to the categories of damaged (presence of diffused discoloration, necroses or holes between the ridges), variable (moderately damaged, with occasional discolorations or necroses), or healthy (no visual injury).
For the statistical analysis, the Kruskal-Wallis non-parametric H test was used to detect differences in maximal PSII efficiency (P<0.05), using the Dunn’s test for post-hoc comparisons.
Results
Impact of logging on occurrence and biomass of L. pulmonaria
Forest logging reduced stem density from ~ 1100 to 165 stems ha-1, with a consequent increase of sun irradiance on retained-isolated trees (from 130-1100 to 900-1550 μmol s-1 m-2 PAR - Tab. 1). Owing to the depletion of host trees, occurrence and biomass of L. pulmonaria were significantly reduced. Due to logging operations, along the logged transect, only 26% of the plots (9 out of 34) resulted colonized by L. pulmonaria (284 individuals over 14 trees - Q. cerris and Q. Ilex), 13 plots remained without host trees and 12 with Lobaria-free trees. On the other hand, along the unlogged transect, 45% of the plots (18 out of 40) were colonized (288 individuals over 50 trees) and all plots included a potential host tree (Q. pubescens, Q. cerris, Fraxinus ornus and Q. ilex, including stumps). Most of the thalli (75%) in the logged stand were small and sterile, while the largest ones (> 26×16 cm) disappeared due to logging operations (Tab. 1). After cutting, the share of larger thalli with diaspores (vegetative and/or sexual) was 50% of the unlogged stand. In both cases, 75% of the colonized plots were located within the first 50 m from the forest margin, which hosted 84% of the overall lichen biomass. A previous study ([30]) reported the presence of rare lichens sharing the same habitat with L. pulmonaria (namely, Fuscopannaria mediterranea, Gyalecta derivata, Megalaria laureri, Pannaria conoplea, Parmeliella triptophylla, Pectenia plumbea, Ricasolia amplissima). Later, also the rare macrolichen Lobarina scrobiculata, was found. After logging, only P. plumbea, L. scrobiculata and R. amplissima were recorded within the logged transect.
Tab. 1 - Characteristics of the investigated oak forest, the sampling transects and the model species L. pulmonaria, comparing logged and unlogged stands. Dimension of the thalli: average length of larger and shorter diameters, in parenthesis the number of specimens. (*): tree clusters (islands) excluded; (**): according to Nascimbene et al. ([24]).
| Level | Parameters | Logged stand | Unlogged stand |
|---|---|---|---|
| Mixed oak forest | Density of potential host trees* | ~ 165 stems ha-1 | ~ 1050 stems ha-1 |
| Solar radiation (at noon, sunny days) | 900-1550 μmol s-1 m-2 PAR | 130-1100 μmol s-1 m-2 PAR | |
| L. pulmonaria biomass (0-2 m) | ~ 7.3 kg ha-1 | ~ 11.5 kg ha-1 | |
| Sampling transects | Plots colonized by L. pulmonaria | 26% | 45% |
| Host trees colonized along the transects | 14 stems | 50 stems | |
| Host tree circumference | 14 - 148 cm | 8 - 88 cm | |
| Other lichens of conservation concern** | 3 species | 8 species | |
| Model species level | L. pulmonaria specimens per transect | 284 | 288 |
| Sterile juvenile thalli | 6 cm × 4 cm (213) | 5 cm × 3 cm (141) | |
| Thalli with vegetative diaspores | 14 cm × 9 cm (40) | 13 cm × 9 cm (112) | |
| Fertile thalli with apothecia | 26 cm × 16 cm (31) | 36 cm × 23 cm (35) |
By measuring dimensions and weight of fragments and whole thalli of regular shape, the calculated surfaces of L. pulmonaria were converted into a biomass estimate of 15.2-19.8 mg cm-2 (95% confidence interval). The combination of data on occurrence and biomass of the specimens allowed estimating the biomass of L. pulmonaria still standing in the logged area up to 2 m from the ground (ca. 7.3 kg ha-1), which mainly consisted of sterile thalli (68%). The same procedure applied to the unlogged stand allowed estimating an average biomass of 11.5 kg ha-1, which largely consisted of thalli with vegetative diaspores (39%) or also sexual structures (12%). A comparison between thallus sizes (unlogged vs. logged stand) suggested that mainly larger thalli fell to the ground and hence were damaged. In fact, 60% of the samples saved from ground and bark of cut logs were fertile and (excluding thallus fragments) with dimensions (length or width) up to 40 cm.
Before logging, the biomass of L. pulmonaria (estimated as described above) in the study area was 15.8-19.6 kg ha-1 and plots in the highly colonized sector reached levels up to 1.8 ± 0.1 kg every 100 m2. Once we subtracted the biomass still standing in the logged area (7.3 kg ha-1), we evinced that 8.5-12.3 kg ha-1 (22-57% of the initial biomass, with an average of 40%) could have been lost due to forest cutting (37-54 kg over the whole area). Altogether, the collaboration with local authorities and a short field training of woodcutters allowed preserving a significant share of Lobaria-trees (65% of the standing trees) and small fragments of forest with Lobaria-trees which could act as reservoir.
Assessment of the vitality of L. pulmonaria
One year later (spring 2017), the thalli still growing on isolated-retained trees in the logged area showed visible symptoms of damage in 46% of Lobaria-trees, consisting in bronzing (melanisation), or in the worst case (14%) discoloration, bleaching and curling of the thalli. The remaining fraction (54% during spring 2017) still consisted of healthy thalli, with a dominant greenish colour and absence of discolorations and necrotic parts.
Kruskal-Wallis H test showed a statistically significant difference in maximal PSII efficiency between sites, χ2 (2, N = 300) = 142.85, P < 0.001. One year after forest cutting a significant decrease of the maximal PSII efficiency of the lichen photobiont (FV/FM) was observed in the thalli of the logged stand, when compared to those of the unlogged stand and to the thalli harvested (saved) from ground and cut logs in 2016. Overall, FV/FM values were within the range 0.726-0.760 (95% confidence interval) in the unlogged area and 0.476-0.601 in the logged area (Tab. 2).
Tab. 2 - Potential quantum yield of primary photochemistry (FV/FM), as indicator of the vitality of the samples. Mean ± standard deviation (SD) and 95% confidence interval (95% CI). (n): Fluorescence measurements. Values followed by a different letter are significantly different (P<0.05) after Dunn’s test.
| Lobaria pulmonaria | n | FV/FM | Conditions | |
|---|---|---|---|---|
| Mean ± SD | 95% CI | |||
| Thalli saved from the logged area (2016) | 100 | 0.727 ± 0.042 a | 0.705 - 0.750 | healthy |
| Logged area (2017) | 100 | 0.539 ± 0.117 b | 0.476 - 0.601 | variable |
| Unlogged area (2017) | 100 | 0.743 ± 0.031 a | 0.726 - 0.760 | healthy |
Discussion
Impact of forest logging on L. pulmonaria
Forest logging significantly reduced the occurrence and biomass of L. pulmonaria within the managed stand, mainly due to the depletion of host trees. Rare species that shared the same habitat with L. pulmonaria ([30]) were likely affected or lost after logging. Similarly, in Central Spain it was demonstrated that total species richness and in particular cyanolichens richness, were negatively affected by intensive forest management ([1]). Noteworthy, in our study a comparable number of thalli of L. pulmonaria has been recorded both in the logged and the unlogged transects, suggesting that the logged stand was densely colonized, much more than the unlogged one. Our measurements revealed an average estimated loss of 40% of the lichen biomass and showed that L. pulmonaria thalli remained on retained-trees were those of smaller size and chiefly sterile due the loss of wider and fertile ones. Edman et al. ([9]) estimated that after logging in a hardwood forest of Canada, the average abundance of L. pulmonaria per tree was five times higher in uncut stands than in cut stands; moreover, the reduction of suitable trees and, hence, the alteration of site quality, led to a significant decline of fertile thalli after 5-9 years.
Forest logging affected L. pulmonaria also indirectly, in particular increasing light influx to the thalli remained within the logged stand, leading to an overall decrease of their photosynthetic efficiency. This lichen, especially in the dry state, proved to be very sensitive to a high irradiance under laboratory conditions ([11]). High-light-intensity damage to L. pulmonaria thalli has been reported also in field studies as a consequence of abrupt changes in light conditions ([12]), such as after logging. In fact, long-term exposures under these conditions may produce visible symptoms of injury, such as bronzing and bleaching of the thalli ([12]), similar to those observed in our study. In the Mediterranean region, higher light irradiance in oak forests during summer, together with the decoupling of moisture and light availability across an annual cycle, have been hypothesized to shorten the generation time of L. pulmonaria in oak woods compared to beech forests ([36]).
Lobaria pulmonaria has been recognized as sensitive to the effects of logging and the consequent increase in habitat dryness even in boreal forests. A simulated logging carried out collecting L. pulmonaria thalli in old forests and transplanting them into clear-cuts showed that logging in winter was less harmful and could be associated to higher growth rates than logging in summer, which was associated with higher chlorophyll degradation and lower growth ([18]). Furthermore, partial cuts better supported L. pulmonaria growth rather than clear-cuts ([6], [7]). Under a climate change scenario, L. pulmonaria could face a high extinction risk associated with a possible reduction of their distribution range ([26]). Therefore, any habitat modification that implies an increase of dry conditions would likely affect this sensitive species (and other associated cryptogams) much more than in the past.
Practical suggestions for the conservation of threatened macrolichens in Mediterranean oak forests
The conservation of epiphytic lichens depends on the understanding of the main factors influencing their distribution, abundance, dispersal, establishment, growth and reproduction ([2]). Lobaria pulmonaria is an indicator of valuable forest habitats ([22], [25]) and important sites for the conservation of cryptogamic species belonging to understudied groups (e.g., bryophytes - [40], [4]). In our case, to mitigate further potential damages due to logging, several Lobaria-trees and patches (islands) with uneven-aged trees were preserved in the investigated stand ([10]), without a negative impact on the activity of woodcutters. Despite the fact that thalli on retained-isolated trees showed a lower vitality compared to the unlogged stand, they could still act as a source of propagules. Furthermore, specimens on retained-forest patches (islands) may represent a reservoir for future recolonizations.
An ecologically sustainable management should take into account that sensitive species with long generation cycles may not be able to recolonize available forest habitats if the sources of generative propagules have been depleted ([40], [14]). In this perspective, retaining patches with colonized trees, sufficient moist microclimate ([42], [31], [46]) and a mixed selection of younger as well as older host trees could be effective practices for the conservation of rare epiphytic lichens ([37]). These considerations on suitable forest management practices should be extended also for the safeguard of other understudied taxa of forest cryptogams. The survival of L. pulmonaria is compatible with non-intensive forest management practices, provided that in managed stands a significant portion of Lobaria-trees is preserved (see [41] and references therein), that the extent of the disturbance is not wider than the dispersal ability of the species and that all successional stages persist and continuously occur after logging ([20], [15]). Furthermore, forest clearings should be kept small to maintain a sufficiently moist microclimate ([46]) and strong interconnections between the stands ([23], [25]). These features are of outmost importance in potentially dry environments, such as Mediterranean oak forests. Moreover, populations with fertile individuals (as in our case) must be at a higher rank in terms of conservation priority, than strictly asexual populations.
Research results underline an international responsibility for the protection of forest habitats hosting sensitive species ([8]) and support the inclusion of Lobaria species within European conservation policies, such as the Habitat Directive. The current condition of L. pulmonaria (inclusion in red lists and legal protection) in most European countries has been summarized in Tab. S1 (Supplementary material). The lichen is generally red-listed, based on IUCN categories, in northern and central Europe (up to Russia) and in several countries of central Europe it is also legally protected. On the other hand, although widely declining, in Mediterranean countries the species has a lower level of implementation in conservation policies. Therefore, it has been highlighted the importance of encouraging stakeholders to identify and protect stands with high conservation priority that could act as sources of propagules to the surrounding environment ([25]). This is relevant especially in the case of unprotected forests. However, to prevent situations of damage (as the one described in this study), L. pulmonaria should be formally protected also in Mediterranean countries, at least ensuring the safeguard of those forest habitats hosting fertile populations.
Conclusions
Logging for timber in a Mediterranean mixed oak forest in central Italy depleted forest epiphytic cryptogams, including a large population of the macrolichen L. pulmonaria, red-listed in several European countries, but not in Italy. One year after logging operations, individuals on retained-isolated trees within the logged stand showed lower photosynthetic performances compared to those of the unlogged stand. Their vitality was significantly reduced. The study outlined specific issues for the conservation of macrolichens in Mediterranean forest ecosystems and the outmost importance of ensuring the safeguard of oak forest stands hosting fertile populations of L. pulmonaria in order to prevent unintended damages.
Acknowledgements
We are grateful to Elena Guerrini, Gabriele Sgroi, Tania Contardo (Italy) and Maya Thapa (Nepal) for helping with the lichen collection; to the students of the University of Florence for their help during field sampling; to Alessandro Ceppatelli and Mauro Pisaneschi (forest managers, “Unione dei Comuni della Val di Merse”, Tuscany, Italy) for their professional support and sensitivity; to Michelle Abajian (USA) for language revision. We would like to thank Eugenios Agathokleous (Cyprus), Gintare Sujetoviene (Lithuania), Silvana Munzi and Paula Matos (Portugal), Luciana Zedda (Germany), Maja Maslać (Croatia), Aino Hämäläinen (Finland), Polina Degtyarenko (Estonia), Martin Westberg (Sweden), Anna Fedosova (Russia), JiÅÂí Malíček (Czech Republic) and László Lökös (Hungary) for information concerning the protection of Lobaria pulmonaria in Europe. We acknowledge two anonymous reviewers for their valuable comments on a first draft of the manuscript.
Author contribution
LP, RB, EB, AV, BA, LDN, ML, SL, MM, SR (investigation); ZF, AG, SL, SR, AL (information concerning the inclusion in red lists and legislations); LP, JN, SLo, SR, RB, ZF, AG (planning of the study, data analysis and elaboration, writing and reviewing of the manuscript); LP (coordinator).
References
CrossRef | Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
Online | Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Department of Biology, University of Pisa, v. Ghini 13, 56126, Pisa (Italy)
Elisabetta Bianchi
Luca Di Nuzzo
Department of Biology, University of Florence, v. La Pira 4, I-50121, Florence (Italy)
Anna Lackovičová
Anna Guttová
Plant Science and Biodiversity Centre, Slovak Academy of Sciences, Dúbravská cesta 9, SK-84523 Bratislava (Slovakia)
Department of Biological, Geological and Environmental Sciences, University of Bologna, v. Irnerio 42, I-40126 Bologna (Italy)
Marco Marchetti 0000-0002-5275-5769
University of Molise, Campobasso (Italy)
Associazione Culturale di Murlo, p.zza Cattedrale, I-53016 Murlo, Siena (Italy)
Reparto Carabinieri per la Biodiversità di Siena, Loc. Il Braccio 4, I-53100 (Italy)
Department of Natural and Land Sciences, University of Sassari, Loc. Piandanna, I-07100 Sassari (Italy)
Stefano Loppi 0000-0002-3404-1017
Department of Life Science, University of Siena, v. Mattioli 4, I-53100 Siena (Italy)
Corresponding author
Paper Info
Citation
Paoli L, Benesperi R, Fačkovcová Z, Nascimbene J, Ravera S, Marchetti M, Anselmi B, Landi M, Landi S, Bianchi E, Di Nuzzo L, Lackovičová A, Vannini A, Loppi S, Guttová A (2019). Impact of forest management on threatened epiphytic macrolichens: evidence from a Mediterranean mixed oak forest (Italy). iForest 12: 383-388. - doi: 10.3832/ifor2951-012
Academic Editor
Rodolfo Picchio
Paper history
Received: Aug 20, 2018
Accepted: Apr 28, 2019
First online: Jul 12, 2019
Publication Date: Aug 31, 2019
Publication Time: 2.50 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2019
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 45919
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 37827
Abstract Page Views: 4081
PDF Downloads: 3043
Citation/Reference Downloads: 6
XML Downloads: 962
Web Metrics
Days since publication: 2559
Overall contacts: 45919
Avg. contacts per week: 125.61
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2019): 12
Average cites per year: 1.71
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Distribution factors of the epiphytic lichen Lobaria pulmonaria (L.) Hoffm. at local and regional spatial scales in the Caucasus: combining species distribution modelling and ecological niche theory
vol. 17, pp. 120-131 (online: 30 April 2024)
Research Articles
Retaining unlogged patches in Mediterranean oak forests may preserve threatened forest macrolichens
vol. 12, pp. 187-192 (online: 10 April 2019)
Short Communications
Effect of four levels of shade on survival, morphology and chlorophyll fluorescence of Nothofagus alessandrii container-grown seedlings
vol. 8, pp. 638-641 (online: 08 January 2015)
Research Articles
Calibration of a multi-species model for chlorophyll estimation in seedlings of Neotropical tree species using hand-held leaf absorbance meters and spectral reflectance
vol. 9, pp. 829-834 (online: 17 May 2016)
Research Articles
Relationships between leaf physiognomy and sensitivity of photosynthetic processes to freezing for subtropical evergreen woody plants
vol. 12, pp. 551-557 (online: 17 December 2019)
Research Articles
Effects of substrate and ectomycorrhizal inoculation on the development of two-years-old container-grown Norway spruce (Picea abies Karst.) seedlings
vol. 8, pp. 487-496 (online: 10 November 2014)
Research Articles
Magnolia grandiflora L. shows better responses to drought than Magnolia × soulangeana in urban environment
vol. 13, pp. 575-583 (online: 07 December 2020)
Research Articles
Needle traits of understory Silver fir, Norway spruce, and Scots pine in response to increased canopy openness in a birch-dominated stand
vol. 18, pp. 273-282 (online: 14 October 2025)
Research Articles
Analysis of biometric, physiological, and biochemical traits to evaluate the cadmium phytoremediation ability of eucalypt plants under hydroponics
vol. 10, pp. 416-421 (online: 24 March 2017)
Research Articles
Wintertime photosynthesis and spring recovery of Ilex aquifolium L.
vol. 12, pp. 389-396 (online: 31 July 2019)
iForest Database Search
Search By Author
- L Paoli
- R Benesperi
- Z Fačkovcová
- J Nascimbene
- S Ravera
- M Marchetti
- B Anselmi
- M Landi
- S Landi
- E Bianchi
- L Di Nuzzo
- A Lackovičová
- A Vannini
- S Loppi
- A Guttová
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
- L Paoli
- R Benesperi
- Z Fačkovcová
- J Nascimbene
- S Ravera
- M Marchetti
- B Anselmi
- M Landi
- S Landi
- E Bianchi
- L Di Nuzzo
- A Lackovičová
- A Vannini
- S Loppi
- A Guttová
Search By Keywords







