
Retaining unlogged patches in Mediterranean oak forests may preserve threatened forest macrolichens
iForest - Biogeosciences and Forestry, Volume 12, Issue 2, Pages 187-192 (2019)
doi: https://doi.org/10.3832/ifor2917-012
Published: Apr 10, 2019 - Copyright © 2019 SISEF
Research Articles
Abstract
Forest management practices may heavily impact epiphytic (tree inhabiting) organisms. Retaining tree patches and buffer strips in logged stands may contribute to preserve ecosystem functioning and the vitality of epiphytic organisms in managed forests. To test these statements, the threatened forest macrolichen Lobaria pulmonaria (L.) Hoffm. was used as a model species, since it is a “flag” indicator species of forest ecosystems with long ecological continuity. To this purpose, photosynthetic performances, thallus anatomy and water holding capacity (WHC) of samples of L. pulmonaria were investigated in a logged mixed oak forest (Tuscany, Italy), confronting lichen thalli from retained-forest patches and retained-isolated trees, 18 months after logging. Compared with those of retained-forest patches, thalli on the trunks of retained-isolated trees were thinner and showed lower vitality (as indicated by the potential quantum yield of primary photochemistry - FV/FM and the index of overall photosynthetic performance - PIABS), as well as lower water holding capacity. In contrast, thalli from forest patches had performances comparable to those of healthy samples from unlogged forests.
Keywords
Biodiversity Conservation, Ecosystem Services, Forest Logging, Lobaria pulmonaria, Photosynthetic Performance, Water Holding Capacity
Introduction
A sustainable forest management scenario should face the conflicting goals of economic return from wood production and the conservation of biodiversity ([10], [17]). Retention forestry represents a valuable management option to conserve the structural, functional, and compositional diversity of forest ecosystems and to increase forest influence in harvested areas ([10]). On the contrary, intensive forest management practices may heavily affect terrestrial ecosystems and their components, altering animal, plant and fungal diversity, including forest lichens.
The epiphytic lichen Lobaria pulmonaria (L.) Hoffm. is such an example: an endangered forest macrolichen ([22]); considered as a “flag” indicator species ([24]) of forest ecosystems with long ecological continuity worthy of conservation ([18]); red-listed and legally protected in several European countries, but not in the Mediterranean area. The species is overall declining throughout Europe as a consequence of air pollution and intensive forest management (including habitat fragmentation), whose effects are expected to be further exacerbated by climate change ([19]). Oak-dominated forests are one of the main habitats for L. pulmonaria populations in the Mediterranean area ([23]). It is especially in these potentially dry environments that logging may damage species with poor dispersal ability such as L. pulmonaria, which would remain restricted to undisturbed parts of the forest landscapes ([24]). As a consequence of logging, forest lichens are exposed to a sudden increase of solar radiation and dry conditions, which, if in excess of their ecological range, may negatively affect their photosynthetic activity, and hence their overall vitality ([6], [14]). Within a certain range, forest lichens are able to withstand dry conditions, e.g., by increasing melanisation and consequently reducing photosynthetic performances ([16]). The photosynthetic performances are valuable proxies of the vitality of the lichen thallus ([6]).
So far, lichens are often neglected by nature conservation managers, despite the fact that their ecological role in the provision of ecosystem functions (e.g., water and nutrient cycling, metal chelation, air quality and microclimate regulation, primary colonization and soil formation) and services (e.g., food, habitat, shelter) is widely acknowledged ([29]). In fact, the presence of lichens (especially healthy macrolichens) increases the water storage capacity in forest canopies and positively influences their hydrology ([5]). Water may fill the intercellular capillarity and air spaces between fungal hyphae and algal cells (representing the water holding capacity of the thalli). Macrolichens (both epiphytic and epigeic) may contribute to passively regulate water flow at ecosystem level, increasing water availability for plants and animals, retaining nutrients and allowing the protection of soils from water erosion ([29]).
In the case of forest logging, retaining unlogged patches and buffer strips are supposed to improve biodiversity conservation and enhance ecosystem functioning of managed forests ([13]). Retained forests are beneficial to the functionality and health of adjacent harvested areas, acting as source population, especially for species with limited dispersal ability. Furthermore, maintaining refugia of mature-forest species may extend the area of forest influence in nearby harvested sites ([1]). The combination of a mixed retention system integrating retained forest patches and interconnected isolated trees should allow epiphytic organisms to survive and persist through the early (drier) stages of stand development in Mediterranean forests. In the long term, retaining forest-patches is expected to facilitate species re-establishment and on the whole, enhance forest biodiversity ([1], [2]).
Our working hypothesis is that in comparison with retained-isolated trees, retaining unlogged forest patches allows to preserve: (1) the vitality of the model species (L. pulmonaria); (2) the water holding capacity (WHC); and hence (3) ecosystem services provided by the presence of healthy forest macrolichens. To this purpose, we assessed photosynthetic performances, thallus anatomy and water holding capacity (WHC) of L. pulmonaria thalli within a logged mixed oak forest (Tuscany, Central Italy), comparing unlogged forest patches and retained-isolated trees.
Materials and methods
Experimental constraints and study area
In 2016, a large population of L. pulmonaria including hundreds of fertile thalli has been depleted and threatened by a legal logging for timber within a Mediterranean mixed oak forest of central Italy (WGS84: 43.1851° N, 11.3602° E). The study area extends for ca. 500 m according with E-W direction on a hillside with a N slope. The logged stand (~ 4.4 ha) was a local hotspot for this species. Lobaria pulmonaria has a patchy distribution there, colonizing more than 1000 phorophytes: Quercus ilex, Q. cerris, Q. pubescens, Arbutus unedo, Sorbus torminalis, Crataegus monogyna, Fraxinus ornus and even the bark of Hedera helix growing on these trees. Scattered thalli occur also on fallen and standing deadwood and on soil-dwelling mosses. Oak wood forests in the area are generally managed with a coppice system with standards and rotation cycles of 18-20 years. Some stands with low management intensity have a longer logging cycle, which dates back to more than 40 years ago, as in our case. Since L. pulmonaria is not protected by law, logging operations did not take into account the presence of such relevant population. Nevertheless, upon request from a group of lichenologists, local authorities temporary suspended the cutting for a few days: at that moment about 1 ha of the stand was already cut. The cooperation among stakeholders (scientists, local authorities and woodcutters) allowed redirecting the subsequent logging activity to an uneven-aged coppice system, which at least preserved several isolated trees with L. pulmonaria (Lobaria-trees) and moreover, selected patches composed of uneven-aged trees richly colonized by L. pulmonaria. Such patches (also indicated as “aggregates” or “tree-islands”) corresponded to retained (intact) forest fragments of about 0.05-0.15 ha area. A careful observation of the thalli after logging (between 12 and 18 months) suggested that individuals growing on the trunks within 5 m from the forest margin could be partially subjected to a forest margin effect due to increased light, higher temperature and lower humidity. Such individuals were sometimes characterized by bronzed or discolored thallus surfaces. In order to account for this aspect, in retained-forest patches only individuals growing at distances >5 m from the forest margin were sampled and investigated.
Chlorophyll a fluorescence emission as indicator of vitality
Eighteen months after the conclusion of logging operations (February 2018) the vitality of lichen thalli on retained-isolated trees and retained-forest patches was assessed by the analysis of chlorophyll (Chl) a fluorescence emission and compared with that of the samples from an adjacent unlogged area. One hundred individuals (50 per each condition) were randomly selected and harvested from the bark (0-2 m above the ground) of retained isolated-trees and trees within preserved forest patches (distance >5 m from the edge). In the laboratory, measurements of Chl a fluorescence were carried out by a Plant Efficiency Analyzer Handy PEA® (Hansatech Ltd., Norfolk, UK). After a dark adaptation of 10 min using a clip randomly placed over the lobes of the thalli, samples were illuminated for 1 s with a saturating excitation pulse [3000 μmol(photon) s-1 m-2] of red light (650 nm) from a LED into the fluorometer sensor. All fluorescence induction curves were recorded up to 1 s. Three fluorescence curves were recorded per each thallus and analysed by means of the so called JIP-test ([28], [27]), which is used to translate original fluorescence data to biophysical parameters that quantify energy fluxes and their ratios, physiological states, conformation, and overall photosynthetic performance of the samples. The “vitality” of the samples was expressed by the maximum of quantum yield of primary photochemistry as inferred from fluorescence data: φPo = (FM - F0)/FM = FV/FM. In addition, the performance index (PIABS) was calculated from another set of equations: γRC/(1 - γRC)·φPo/(1 - φPo)·ψEo/(1 - ψEo), where γRC = ChlRC/Chltotal is the probability that a PSII (photosystem 2) Chl molecule functions as RC (reaction centre); γRC/(1 - γRC) is approximated in the JIP-test by RC/ABS, with ABS/RC the inferred absorbed energy flux per active RC of PSII; φPo = TR0/ABS (= FV/FM) is the inferred maximum of quantum yield of primary photochemistry; ψEo = ET0/TR0 is the probability that an electron moves further than the electron acceptor QA ([28], [27]).
Water-holding capacity (WHC)
WHC was determined according with the procedure described in Gauslaa et al. ([8]) and Longinotti et al. ([15]). First, each investigated thallus was gently sprayed with mineral water until fully water-saturated, then the wet mass (WM) was measured in three steps: (1) each water-saturated thallus was weighed (WMtotal); (2) then gently shaken three times until no more drops were falling and weighed again (WMshaking); (3) then blotted (upper and lower cortex) with dry filter paper to quantify WMblotting. The specific thallus mass (STM) was calculated as dry mass over wet surface (DM/ Awet). The wet surface area (Awet) was estimated by direct measurements of thallus dimensions integrated using photographed images of wet (after shaking) and dry thalli, by the software ImageJ, version Fiji ([25]).
To calculate the water-holding capacity before (WHCtotal), after shaking (WHCshaking) and after blotting (WHCblotting), the respective WM (WMtotal, WMshaking or WMblotting) was used in the formula WHC = (WM-DM)/Awet. The water left after blotting was accounted as internal water (WHCinternal = WHCblotting), whereas the external water was computed as WHCexternal = WHCshaking - WHCblotting. Values are expressed as mg H2O cm-2. At the end of the experiment, whenever possible, the thalli were carefully re-exposed in their native environment. In order to prevent any further unintentional damage to the population, samples within the unlogged area were not harvested for WHC measurements.
Thallus anatomy
Anatomical features of L. pulmonaria thalli from isolated trees and forest patches were measured. At this purpose, 50 small discs of lichen material (one per each thallus) were randomly selected by means of a hole punch for paper sheets. A cross-section was created from each disc (5 mm diameter), hydrated, and then observed under a dissecting microscope at 400× magnification. At three positions, one near each end of the cross-section, and one in the middle, the thickness of the upper cortex, the photobiont layer, and the medulla (plus lower cortex), were separately measured. The thickness of a given layer in one disc (one observation) was the mean value from the three measuring positions, as described in Gauslaa & Coxson ([7]).
Statistical analyses
The significance of differences in the investigated parameters between samples in retained-forest patches and isolated-trees was tested using the t-test (p<0.05). Data not matching a normal distribution (Shapiro-Wilk test) were treated with a Box-Cox transformation, to meet normality criteria.
Results
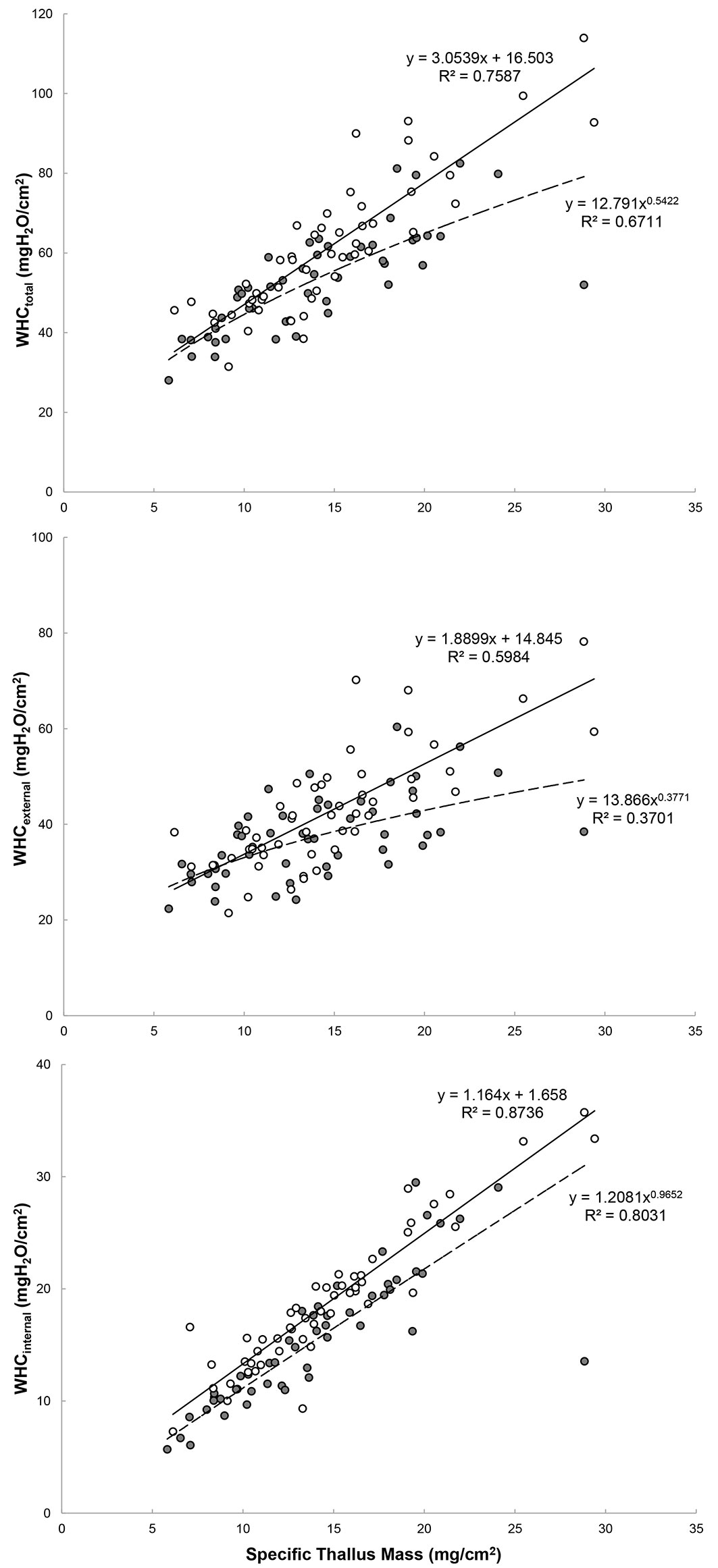
Forest logging reduced the density of stems from ~ 1100 to 165 per ha, with a consequent increase of sun irradiance on retained-isolated trees (from 130-1100 to 900-1550 μmol m-2 s-1 PAR at noon). Eighteen months after the conclusion of logging operations, L. pulmonaria individuals growing within retained-forest patches clearly showed higher photosynthetic performances than those growing on retained-isolated trees (Tab. 1), as suggested by higher values of the maximal quantum yield of primary photochemistry (φPo = FV/FM) and by the performance index (PIABS) of the photobionts. Fluorescence transients (grouped as shown in Fig. 1), reflected a higher photosynthetic activity of those individuals retrieved from retained forest patches. Such performance was similar to that of samples from unlogged forest stands in the same area (for comparison, fluorescence transients of thalli measured in situ in an adjacent unlogged area are given in Fig. 1). Individuals of L. pulmonaria on the trunks of retained-forest patches showed significantly higher values of internal (+21%), external (+14%) as well as total (+16%) WHC compared to the thalli growing on isolated Lobaria-trees (Tab. 1). WHC increased with specific thallus mass- STM (Fig. 2), especially in forest patches, where the strongest relationship was found between STM and WHCinternal (R2 = 0.87).
Tab. 1 - Photosynthetic parameters (FV/FM and PIABS, N=300), water holding capacity expressed as mg H2O cm-2 (WHC total, external and internal; N=100), specific thallus mass (STM - see text; N=100) and thickness of each thallus layer (µm; N=100) in Lobaria pulmonaria thalli from retained-isolated trees and retained-forest patches (average ± standard deviation). In parentheses, 95% confidence interval. (*): values differ significantly (t-test, p<0.05) comparing forest patches with isolated trees.
| Parameters | Retained-forest patches | Retained-isolated trees |
|---|---|---|
| FV/FM | 0.720 ± 0.051* (0.712 - 0.729) |
0.515 ± 0.146* (0.491 - 0.538) |
| PIABS | 0.600 ± 0.285* (0.553 - 0.646) |
0.172 ± 0.147* (0.148 - 0.196) |
| WHCtotal | 61.37 ± 17.47* (56.40 - 66.34) |
53.06 ± 12.73* (49.44 - 56.68) |
| WHCinternal | 18.76 ± 6.21* (17.00 - 20.52) |
15.55 ± 5.92* (13.87 - 17.24) |
| WHCexternal | 42.61 ± 12.17* (39.15 - 46.07) |
37.50 ± 8.50* (35.08 - 39.92) |
| STM (DM/Awet) | 14.69 ± 4.98 (13.28 - 16.11) |
13.95 ± 5.00 (12.53 - 15.37) |
| Upper cortex | 37.1 ± 8.9* (34.5 - 39.6) |
33.1 ± 6.2* (31.4 - 34.9) |
| Photobiont layer | 48.6 ± 11.3* (45.4 - 51.8) |
31.5 ± 9.3* (28.9 - 34.2) |
| Medulla + lower cortex | 109.5 ± 29.9 (102.4 - 116.6) |
101.9 ± 26.1 (94.5 - 109.3) |
Fig. 1 - Chlorophyll a fluorescence transients of Lobaria pulmonaria thalli from retained-isolated trees (grey circles) and retained-forest patches (white circles). For comparison, the average curve of (in situ) individuals in the unlogged area is given (white squares). Vertical bars represent standard deviations.
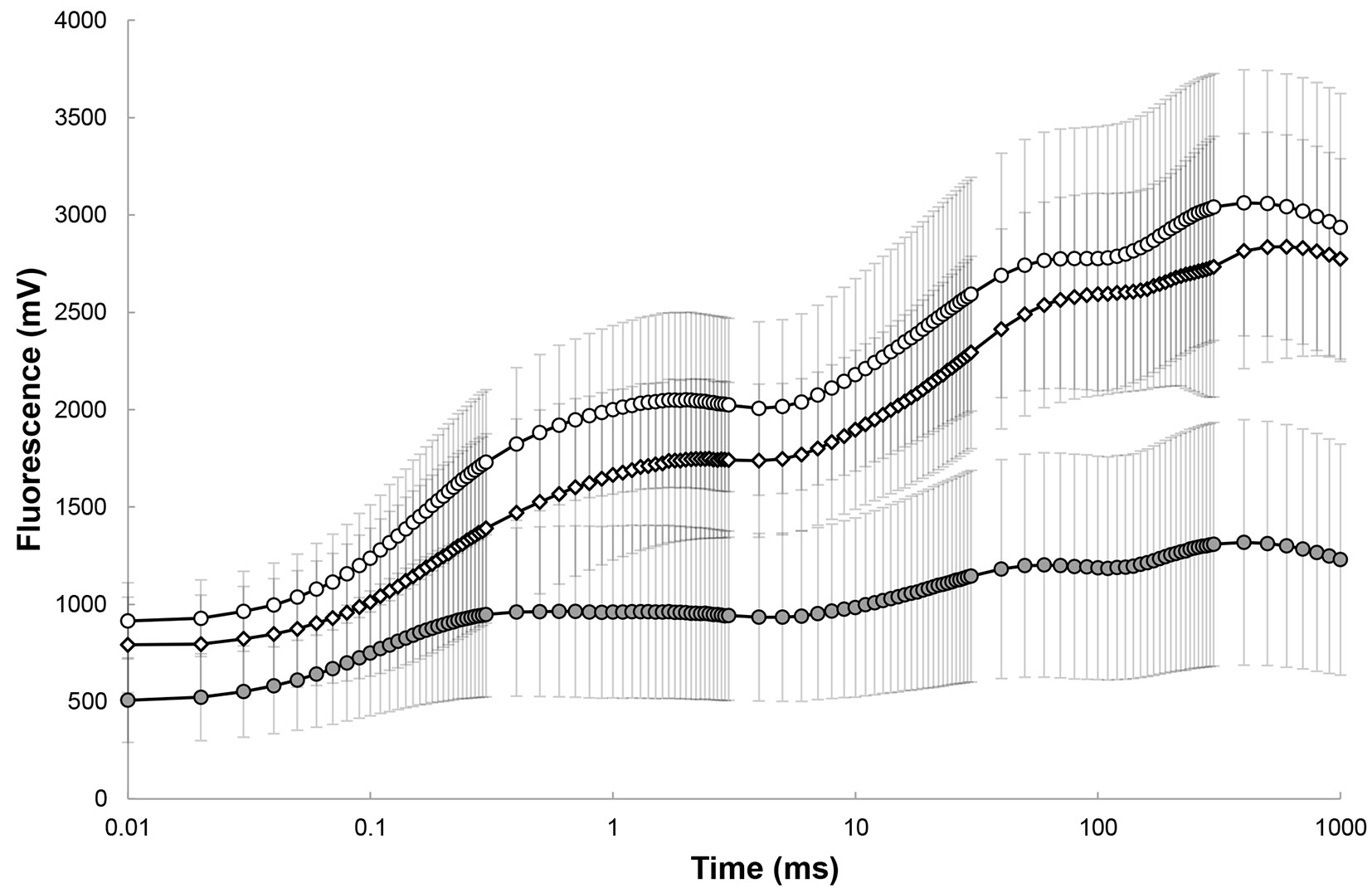
Fig. 2 - Water-holding capacity (WHC total - upper panel; WHC external - middle panel; WHC internal - lower panel; N=100) in relation to the specific thallus mass (STM) of Lobaria pulmonaria thalli from retained-isolated trees (grey circles) and retained-forest patches (white circles). Regression lines/curves, best fit equations and R2 (p<0.05) are given (solid = forest patches; dashed = isolated trees).
With respect to thallus anatomy, the thalli from isolated trees had a thinner upper cortex (33.1 μm) and moreover a thinner photobiont layer (31.5 μm) than the thalli from forest patches (37.1 and 48.6 μm of cortex and photobiont layer, respectively - see Tab. 1 for details), meaning a 35% reduction of the algal layer in thalli from isolated trees.
Discussion
Critical aspects of retaining only isolated trees
Differences in photosynthetic parameters (FV/FM and PIABS) based on fluorescence measurements indicated that retaining unlogged forest patches in a harvested Mediterranean oak forest enhances the vitality of the model species L. pulmonaria, as compared to retained isolated-trees. The photosynthetic performance of samples from isolated trees, as reflected by the overall index of photosynthetic performance (PIABS), was three times lower than that in samples from forest patches. Moreover, samples from isolated trees also showed a lower WHC. Fluorescence induction curves (Fig. 1) well reflected a different photosynthetic behaviour, confirming a lower performance of those individuals from retained-isolated Lobaria-trees as compared with those from forest patches and the adjacent unlogged area. Furthermore, samples on isolated trees showed a reduction of the photobiont layer, as a likely sign of stress to the photosynthetic partner due to the sudden increase of light irradiance occurred after logging. In line with such data, in 46% of retained-isolated trees L. pulmonaria thalli showed visible symptoms of damage, consisting in bronzing (melanisation), or in the worst case (14%) discoloration, up to bleaching (unpublished data), which were not present within forest patches (at a distance > 5 m from their margin). The conservation of threatened forest lichens depends on the understanding of the factors that influence their distribution, abundance, dispersal, establishment, growth and reproduction ([4]). However, the level of influence of these factors is still unclear for most lichens ([24]). Dry conditions under high light intensity, such as after logging, can be deleterious to lichen thalli ([6]). Lichens are able to adapt to drought and high-light stress through a variety of mechanisms, such as energy dissipation, melanisation of the thalli, reduction of photosynthetic activity ([16]). Lobaria pulmonaria synthesizes melanic pigments when exposed to high solar radiation: such melanisation may effectively reduce high-light stress, although it also reduces photosynthetic activity of the photobionts ([16]). In our case, melanisation was much more evident in samples from retained-isolated Lobaria-trees, which were likely subject to rapid desiccation and hence to a shrinkage of the surface and a curling of the lobes, with respect to the individuals within unlogged forest patches. Despite curling of the lobes during desiccation reduces light exposure and photoinhibition ([3]), such processes may be not enough effective to prevent long lasting or irreversible physiological alterations to forest lichens adapted to the shade, such as L. pulmonaria ([6]), especially when the environment becomes suddenly drier and well-lit. Therefore, it is likely that thalli of the model species from retained-isolated trees will be further affected during the early stages of stand development. Our data provide an early warning on the processes ongoing after logging, and more extensive analyses divided by multiple years and growing seasons would be useful to delineate a long-term overview on the survival of the species.
The benefits of retaining forest-patches
Our results have shown that retaining forest-patches in a Mediterranean oak forest enhances water storage, as reflected by the WHC of L. pulmonaria. Positive relationships were found between photosynthetic parameters (FV/FM and PIABS) and WHC (total, external and internal), the strongest being between WHCinternal and the overall index of photosynthetic performance - PIABS (Pearson’s correlation coefficient: r=0.35, p<0.001), suggesting that active and vital thalli in forest patches have a higher ability to adsorb and retain water from the surrounding environment with respect to thalli from isolated trees. On the whole, WHC values in our samples were comparable to those reported for L. pulmonaria and other forest macrolichens in N Europe ([15]) and N America ([7]). It has been reported that WHC and STM may increase with light exposure, presumably as a result of acclimation to higher evaporation demands ([7]). If a successful acclimation occurs after logging, the lichen mycobiont may respond by thickening the upper cortex, which increases the protection against low humidity and high irradiance and prolongs hydration periods ([11], [7]). Noteworthy, positive correlations between WHC and thickness of the photobiont layer suggested that also the photobiont as such may play a significant role in the water economy of a lichen ([7]). In our study, this was evident in those individuals from forest patches where higher WHC corresponded to higher photosynthetic performances and to a thick photobiont layer. In the case of isolated-trees (in a context of a Mediterranean environment), lower WHC, thinner upper cortex and photobiont layer, and lower values of the photosynthetic parameters are clear signs of stress in L. pulmonaria thalli. On the whole, such data reflect a difficult acclimation to the new conditions after logging, and hence a potential loss of ecosystem functions. In fact, luxuriant populations of L. pulmonaria may contribute to several ecosystem functions also within Mediterranean oak forests, including the fixation and regulation of atmospheric nitrogen. As for plants, a significant water holding capacity contributes to the hydrological regulation of ecosystems and buffers water seasonal extremes ([26]), in the case of macrolichens it enhances productivity (photosynthesis, carbon fixation, nutrient cycling) and contributes to regulate water flow and ecosystem humidity.
From a conservation perspective, the creation or preservation of micro-reserves as well as longer rotation cycles within productive forests increase the resilience of forest ecosystems to anthropogenic disturbances ([1], [17], [9]), maximizing the availability of microhabitats in Mediterranean oak forests ([21]).
The presence of undisturbed forest aggregates allows the survival of mycorrhizal fungi ([12]), plants and invertebrates ([2]) compared to adjacent harvested areas. However, the effective benefits for biodiversity may vary among different taxa and the size and connections between forest patches should be considered to support forest-dependent species ([1], [2] and references therein, [20]).
Relatively small patches in mixed oak forests seem suitable for L. pulmonaria, since this species generally avoids both excess exposure to light irradiance and full shading (e.g., within Q. ilex dominated stands), rather preferring a combination between diffuse light and air humidity ([4]). The effective extension and distance between forest patches suitable for epiphytic organisms in Mediterranean oak forests, compatible with the goals of wood production and the conservation of biodiversity, still requires investigations. As a concluding remark, it is suggested that retained-forest patches should prudentially include a marginal buffer zone, where samples growing at a distance generally >5 m from the forest margin may maintain a healthy status.
References
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Online | Gscholar
CrossRef | Gscholar
Authors’ Info
Authors’ Affiliation
Anna Guttová 0000-0001-9394-1077
Plant Science and Biodiversity Centre, Slovak Academy of Sciences, Dúbravská cesta 9, SK-84523 Bratislava (Slovakia)
Department of Biology, University of Florence, v. La Pira 4, I-50121 Florence (Italy)
Department of Life Sciences, University of Siena, v. Mattioli 4, I-53100 Siena (Italy)
Luigi Sanità di Toppi 0000-0002-8731-4904
Luca Paoli
Department of Biology, University of Pisa, v. Ghini 13, I-56126 Pisa (Italy)
Corresponding author
Paper Info
Citation
Fačkovcová Z, Guttová A, Benesperi R, Loppi S, Bellini E, Sanità di Toppi L, Paoli L (2019). Retaining unlogged patches in Mediterranean oak forests may preserve threatened forest macrolichens. iForest 12: 187-192. - doi: 10.3832/ifor2917-012
Academic Editor
Gianluca Piovesan
Paper history
Received: Jul 10, 2018
Accepted: Jan 18, 2019
First online: Apr 10, 2019
Publication Date: Apr 30, 2019
Publication Time: 2.73 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2019
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 45914
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 38158
Abstract Page Views: 3761
PDF Downloads: 3156
Citation/Reference Downloads: 3
XML Downloads: 836
Web Metrics
Days since publication: 2677
Overall contacts: 45914
Avg. contacts per week: 120.06
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Jul 2026)
Total number of cites (since 2019): 13
Average cites per year: 1.63
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Distribution factors of the epiphytic lichen Lobaria pulmonaria (L.) Hoffm. at local and regional spatial scales in the Caucasus: combining species distribution modelling and ecological niche theory
vol. 17, pp. 120-131 (online: 30 April 2024)
Research Articles
Impact of forest management on threatened epiphytic macrolichens: evidence from a Mediterranean mixed oak forest (Italy)
vol. 12, pp. 383-388 (online: 12 July 2019)
Review Papers
Towards better practices of salvage logging for reducing the ecosystem impacts in Mediterranean burned forests
vol. 13, pp. 360-368 (online: 25 August 2020)
Research Articles
Investigating the effect of selective logging on tree biodiversity and structure of the tropical forests of Papua New Guinea
vol. 9, pp. 475-482 (online: 25 January 2016)
Research Articles
Biodiversity conservation and wood production in a Natura 2000 Mediterranean forest. A trade-off evaluation focused on the occurrence of microhabitats
vol. 12, pp. 76-84 (online: 24 January 2019)
Review Papers
The forest biodiversity artery: towards forest management for saproxylic conservation
vol. 9, pp. 205-216 (online: 26 October 2015)
Short Communications
Biodiversity and ecosystem functioning: exploring large-scale patterns in mainland China
vol. 5, pp. 230-234 (online: 02 October 2012)
Research Articles
The effects of forest management on biodiversity in the Czech Republic: an overview of biologists’ opinions
vol. 15, pp. 187-196 (online: 19 May 2022)
Book Reviews
National forest inventories: contributions to forest biodiversity assessments (2010)
vol. 4, pp. 250-251 (online: 05 November 2011)
Short Communications
An approach to measuring biodiversity and its use in analysing the effect of nitrogen deposition on woodland butterfly populations in the Netherlands
vol. 2, pp. 46-48 (online: 21 January 2009)
iForest Database Search
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords







