Environmental factors affecting formation of lammas shoots in young stands of Norway spruce (Picea abies Karst.) in Latvia
iForest - Biogeosciences and Forestry, Volume 11, Issue 6, Pages 809-815 (2018)
doi: https://doi.org/10.3832/ifor2539-011
Published: Dec 14, 2018 - Copyright © 2018 SISEF
Research Articles
Abstract
Norway spruce is a relatively fast-growing tree species that is primarily regenerated by planting, which requires significant investments. The formation of lammas shoots (i.e., second flushing in late summer) have been proven to affect the quality and productivity of Norway spruce stands. The aim of our study was to assess the proportion of trees with lammas shoots in young stands of Norway spruce and to investigate the factors affecting their formation. Data have been collected at the end of 2011 in 102 three- to seven-year-old Norway spruce plantations randomly selected across different forest types in the central part of Latvia, and a subset of 21 stands were inventoried again at the end of 2012. In each stand, 20 sample plots were systematically established where trees with and without lammas shoots were counted and micro-environmental factors (moisture, competition, and browsing) were assessed on a three-grade scale. On average, the proportion of trees with lammas shoots was 6.5%. There was no significant effect of the stand age (from 3 to 7 years) on the proportion of trees with lammas shoots nor was there any age-related trend. The effect of forest type on the presence of lammas shoots was not significant. Micro-environmental factors had an important influence on the proportion of trees with lammas shoots. The proportion of lammas shoots in stands with no competition was significantly higher (14.5%) compared to stands with medium and high competition (6.0% and 2.2%, respectively). Similarly, a significantly higher proportion of trees with lammas shoots (11.7%) was observed in sites with normal moisture regime than in sites with slight or notable excess moisture, reaching 4.8% and 1.7%, respectively. Although the influence of browsing damage was not statistically significant, its trend was similar to that observed for competition and moisture regime. Overall, the proportion of trees with lammas shoots was highest in stands showing the most suitable micro-environmental conditions for Norway spruce.
Keywords
Second Flushing, Vegetation Competition, Tending, Moisture Excess, Browsing Damage
Introduction
Norway spruce is the most economically important tree species in the Baltic Sea region, due to its relatively large share in the total forest coverage ([3]), high value of coniferous tree saw logs, and notably shorter rotation period and higher productivity compared to Scots pine ([12]). In Latvia, Norway spruce is the dominant tree species in 18% of the total forest area ([15]). Most stands (83%) are regenerated by planting, which requires notable investments, starting from tree breeding programs and seed orchard maintenance up to plant production, soil preparation, and the planting itself ([20]). Norway spruce plantations in Latvia are mainly established in forest types with fertile mineral soils, with normal moisture regime (Hylocomiosa and Oxalidosa) and drained soil (Myrtillosa mel.) accounting for 34%, 39%, and 9%, respectively, of the total annual planted area by this species ([4], [15]).
Changes in climatic conditions might severely affect the survival and development of forest stands ([24]). Long-term phenological observations across Europe have indicated a trend towards an earlier and longer vegetation period ([41]), which is predicted to continue in the future. By the middle of this century, the length of the vegetation period in Latvia could increase by two to three weeks in comparison to the mean in 1961-1990 ([25]). The response of trees to longer periods of favourable growing conditions might include an increased frequency of formation of lammas shoots (i.e., the second flushing in late summer) after the end of the formation of the predetermined height increment. Formation of lammas shoots has long been studied; however, the underlying physiological mechanisms are not well understood yet ([7]). It has been previously reported that the formation of lammas shoots is significantly affected by both meteorological conditions ([10], [1], [43]) and genetic factors ([17], [34], [42], [9], [8], [39]).
Genetic determination of lammas shoot formation has been established by analysis of provenance trials. For example, Danusevičius & Persson ([9]) reported that the proportion of trees with lammas shoots ranged from 4% ± 2% for provenances from northern Sweden to 17% ± 3% for provenances from Austria at the age of five years in an experiment in Sweden. Significant provenance effects and similar trends (but higher mean values) were reported based on a trial at the same age in Norway. The proportion of trees with lammas shoots was lowest for Scandinavian provenances (18%), followed by Latvian provenances (28%) and Slovak and Czech provenances (54% - [39]). Eriksson ([13]) reported that, in Austria, the high-altitude provenances do not form lammas shoots, while such shoots are formed by valley-bottom provenances. Provenance and family were significant factors affecting the occurrence of lammas shoots, according to Steffenrem et al. ([40]), based on an analysis of open-pollinated progeny trials. However, the family (genetic) contribution to the total variance was relatively low (4%). Lammas growth is often among the traits included in the selection index due to its direct effect on or correlation with the stem and wood quality traits ([19]). For example, Skrøppa et al. ([37]), based on the evaluation of 15 provenances from Finland, Denmark, Germany, and Poland, reported a significant association between the occurrence of lammas shoots at juvenile age and the wood density at the age of 29 years.
Meteorological conditions of the previous year influence the predetermined growth, but conditions of the current year may affect the second flushing ([10], [23], [43]). The increased frequency of lammas shoots for Norway spruce due to the rise in temperature has been proven in laboratory conditions ([22], [26]). This increase has been observed in the field in recent years and has been attributed to changes in climatic conditions ([27], [39]).
The second flushing of Norway spruce and other conifer species has been linked to the increased frequency of trees with double leaders and/or spike knots ([14], [32], [39]). In some cases, the occurrence of these stem defects might be caused by several lammas shoots competing for dominance, as well as by insufficient frost hardiness of the apical shoot (and consequently damage) due to lammas growth, at least partly linked to maladaptation of the studied material. Some studies have indicated that most of the spike knots formed in this way disappear in a few years without causing permanent stem damage ([21]). Formation of lammas shoots is linked to a faster tree growth, thus contributing to increase the productivity of plantations. Indeed, correlations between the mean tree height and the proportion of trees with lammas shoots at five to 13 years of age have been reported in several countries: Germany (r = 0.93, p < 0.01), Sweden (r ≈ 0.3, p < 0.01), and Latvia ([17], [34], [9]).
A systematic inventory of the frequency of second flushing in Norway spruce has not previously been conducted in Latvia, though lammas shoots are demonstrated to have an increasing frequency and importance in the determination of the productivity and quality of spruce stands. Therefore, the aim of the study is to assess the proportion of trees with lammas growth in young stands of Norway spruce and to identify the environmental factors which affect their formation. The following hypotheses are tested: (i) optimal growing conditions have a positive effect on the formation of lammas shoots in young Norway spruce trees; (ii) tree age has a negative effect on the formation of lammas shoots. This knowledge could be further used in the preparation of recommendations for forest management.
Materials and methods
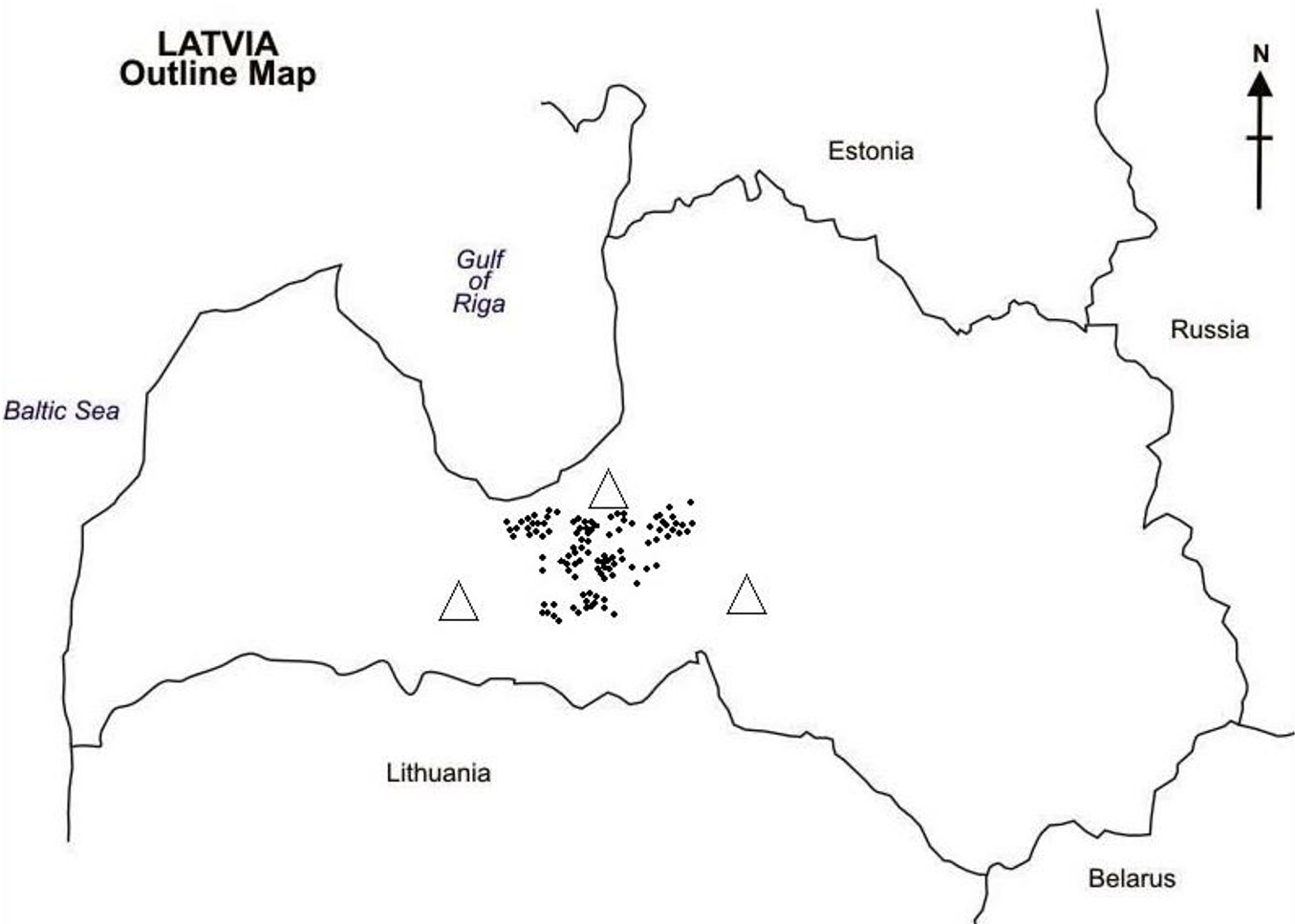
Data have been collected at the end of 2011 from 102 Norway spruce plantations located in the central part of Latvia (56° 37′-56° 57′ N, 23° 37′-24° 49′ E - Fig. 1). Three- to 7-year-old stands (in 2011) were randomly selected across different forest types (Tab. 1). A subset of 21 stands was further surveyed at the end of 2012. All the selected stands have been established using reproductive material from local seed orchards. The area of each stand was at least 0.5 ha and their elevation ranged from 10 to 40 m a.s.l.
Fig. 1 - Location of sampled Norway spruce stands (dots) and meteorological stations (triangles) in Latvia.
Tab. 1 - Number of plots sampled in the Norway spruce stands considered in this study, according to stand age and forest type.
| Forest type | Year of observation |
No. stands |
Age (years) | Total | |||||
|---|---|---|---|---|---|---|---|---|---|
| 3 | 4 | 5 | 6 | 7 | 8 | ||||
| Hylocomiosa | 2011 | 26 | 118 | 153 | 208 | 18 | - | - | 497 |
| 2012 | 5 | - | 20 | 20 | 59 | - | - | 99 | |
| Oxalidosa | 2011 | 9 | - | 20 | 74 | 54 | 17 | - | 165 |
| 2012 | 4 | - | - | 20 | 38 | 19 | - | 77 | |
| Myrtilloso-sphagnosa | 2011 | 13 | 83 | 76 | 43 | - | 16 | - | 218 |
| Myrtilloso-polytrichosa | 2011 | 5 | - | 56 | 39 | - | - | - | 95 |
| 2012 | 1 | - | - | 19 | - | - | - | 19 | |
| Myrtillosa mel. | 2011 | 24 | 69 | 39 | 19 | 191 | 116 | - | 434 |
| 2012 | 4 | - | 18 | - | - | 37 | 20 | 75 | |
| Mercurialiosa mel. | 2011 | 6 | - | - | 57 | 55 | - | - | 112 |
| 2012 | 1 | - | - | - | - | 20 | - | 20 | |
| Myrtillosa turf. mel. | 2011 | 15 | 39 | 72 | 106 | 10 | 37 | - | 264 |
| 2012 | 5 | - | 20 | 20 | 57 | - | - | 97 | |
| Oxalidosa turf. mel. | 2011 | 4 | 37 | - | - | - | 39 | - | 76 |
| 2012 | 1 | - | 20 | - | - | - | - | 20 | |
| Stands of all types | 2011 | 102 | 346 | 416 | 546 | 328 | 225 | - | 1861 |
| 2012 | 21 | - | 78 | 79 | 154 | 76 | 20 | 407 | |
The climate in the study area is predominantly characterised by western winds bringing warm and moist air masses from the Baltic Sea and the Atlantic. According to the Latvian Environment, Geology, and Meteorology Centre ([28]), the mean annual temperature in the region is +5.9°C. The average temperature of the warmest month (July) is +17.0°C (mean maximum temperature +21.5°C), and the annual precipitation is 667 mm. Data from the three nearest meteorological stations (Riga, Dobele, and Skriveri) were used to analyse the possible influence of meteorological conditions on lamma shoot formation (Fig. 1).
In each stand, 20 sample plots (radius 2.82 m, area 25 m2) were placed at regular distance along its longest diagonal. The number of trees with and without current season lammas shoots was recorded in each sample plot. A lammas shoot was defined as the second flushing of the apical or lateral buds of the top shoot, reaching at least 1 cm in length.
The growing conditions of trees in the plots were quite different and not always favourable, due to recent changes in habitat characteristics (e.g., the presence of a beaver dam across the melioration ditch) and browsing pressure (according to the State Forest Service, the density of roe deer, Capreolus capreolus L., was 30-40 inds. per 1.000 ha on average). Thus, for each plot the suitability of micro-environmental factors was assessed in a subset of 70 stands (56 in 2011, 14 in 2012) using a three-grade scale which considered the following factors:
- competition by the surrounding vegetation (within a radius of 0.5 m from the spruce tree), including herbaceous plants, shrubs, and sprouts of other tree species; a score (grade) was assigned to each plot based on the ratio between the mean height of trees and that of the surrounding vegetation (Grade 1: < 0.75; Grade 2: 0.75-1.25; Grade 3: > 1.25);
- moisture regime: Grade 1 was assigned to plots showing normal conditions, Grade 2 to plots with periodic excessive moisture, and Grade 3 to those showing permanent excessive moisture (e.g., due to clogging of a drainage system);
- browsing damage was recorded for trees with damaged tops and/or more than 50% of lateral shoots; a score was assigned to each plot based on the proportion of trees affected by browsing (Grade 1: < 1%; Grade 2: 1%-10%; Grade 3: > 10%).
The proportion of trees with lammas shoots (i.e., the number of trees with lammas shoots out of the total number of trees) at the stand level was calculated as the mean value (± 95% confidence interval) of sample plot proportions. The proportion of trees with browsing damage was calculated in the same way. Spearman’s correlation analysis was used to assess the relation between the proportion of trees with lammas shoots and stand density. To assess the effect of the above factors on the proportion of trees with lammas shoots, a binary logistic generalised linear mixed-effects model (GLMM) was implemented in software R ver. 3.4.3 ([33]) using the library “lme4” ([2]). Three models were developed using the relation between trees with and without lammas shoots at the plot level as the response variable. In all models, the stand ID was set as random effect to account for possible correlation (repeated measures) among sample plots within the same stand. The first model included stand age and forest type as predictors using the whole set of 102 stands inspected in 2011. The second model included the three micro-environmental factors (competition, moisture and browsing damage) and the year of observation as predictors, which were tested on the subset of 70 stands described above. Using the same data subset, a third model was developed which included the suitability of growing conditions as predictor, which was calculated as the sum of scores (grades) for all three micro-environmental factors, ranging from 3 (best) to 9 (worst). Generalised linear hypothesis tests, as implemented in the R library “multcomp” ([18]), were used as post-hoc tests after GLMM when the interaction of the factor or factors with more than two levels were significant compared to the mean proportions between levels.
Results
The proportion of trees with lammas shoots across the sampled stands ranged from 0% to 25% and was 6.5% on average. It was slightly higher in 2012 than in 2011 (in stands sampled in both years, the proportion was 12.9% and 9.1%, respectively), but differences between years were not statistically significant (χ2 = 0.5696, p = 0.450). Initial spacing in all stands was similar, but due to the differences in survival, the density at the age of assessment ranged from 800 to 3600 trees ha-1. The differences in stand density had no statistically significant relation to the proportion of trees with lammas shoots.
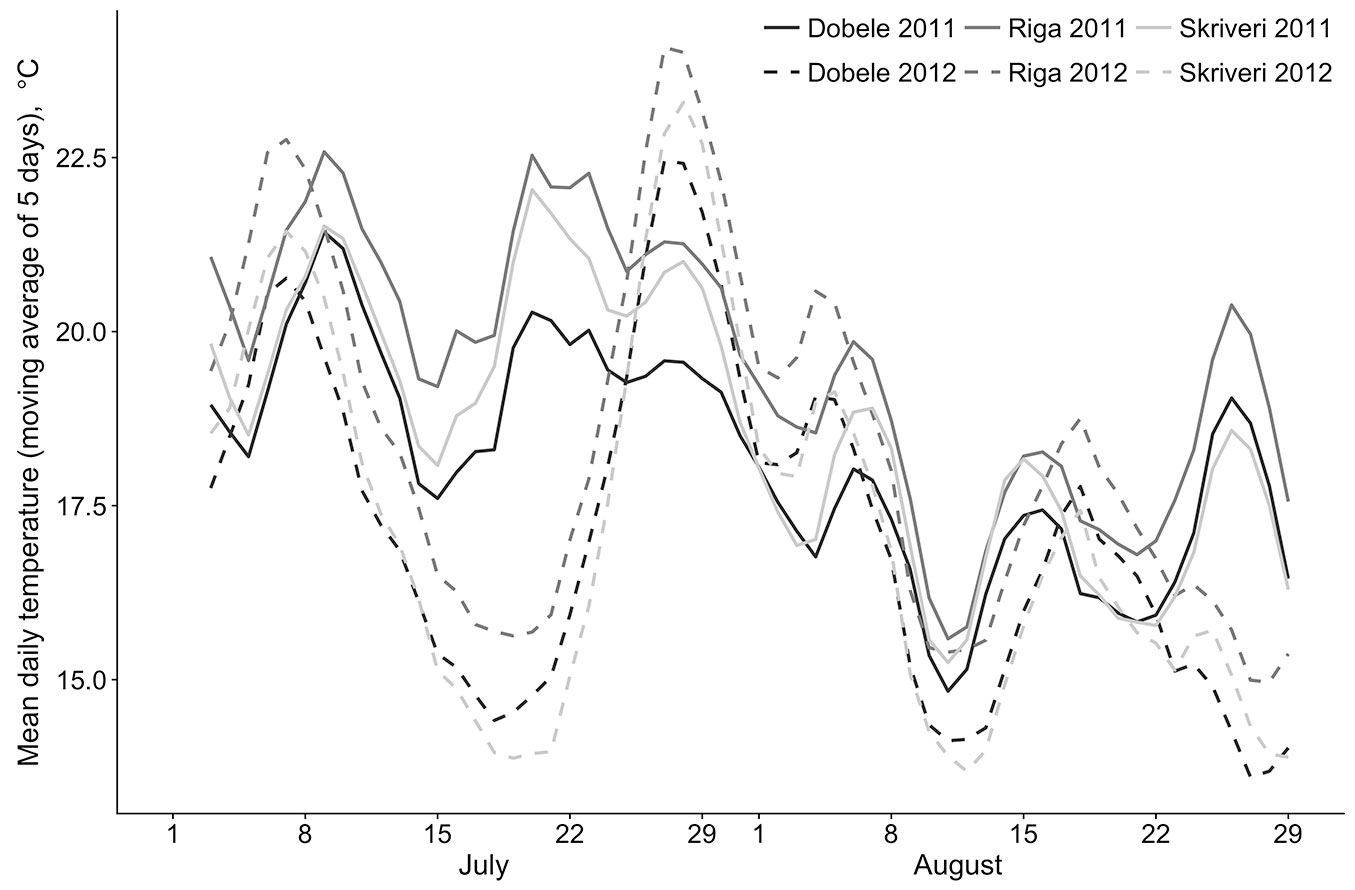
There were no notable differences in air temperature in the period of formation of lammas shoots (end of July to the beginning of August) between 2011 and 2012. The mean diurnal temperature in the last week of July and the first week of August at the Riga meteorological station was 20.4 and 20.7 °C, respectively. In Dobele, it was 18.6 and 19.3 °C, respectively, and in Skriveri it was 19.4 and 19.4 °C, respectively. However, a sharp temperature increase by the end of July 2012 in comparison to the slight decrease in the same period of 2011 was observed (Fig. 2). Temperature differences between the third and fourth weeks of July were 5 to 5.8 °C in 2012, but only 0.3 to 0.5 °C in 2011.
Fig. 2 - Mean daily temperature (moving average of five days) at three meteorological stations (Riga, Dobele, and Skriveri) in July-August 2011 and 2012 (data: [28]).
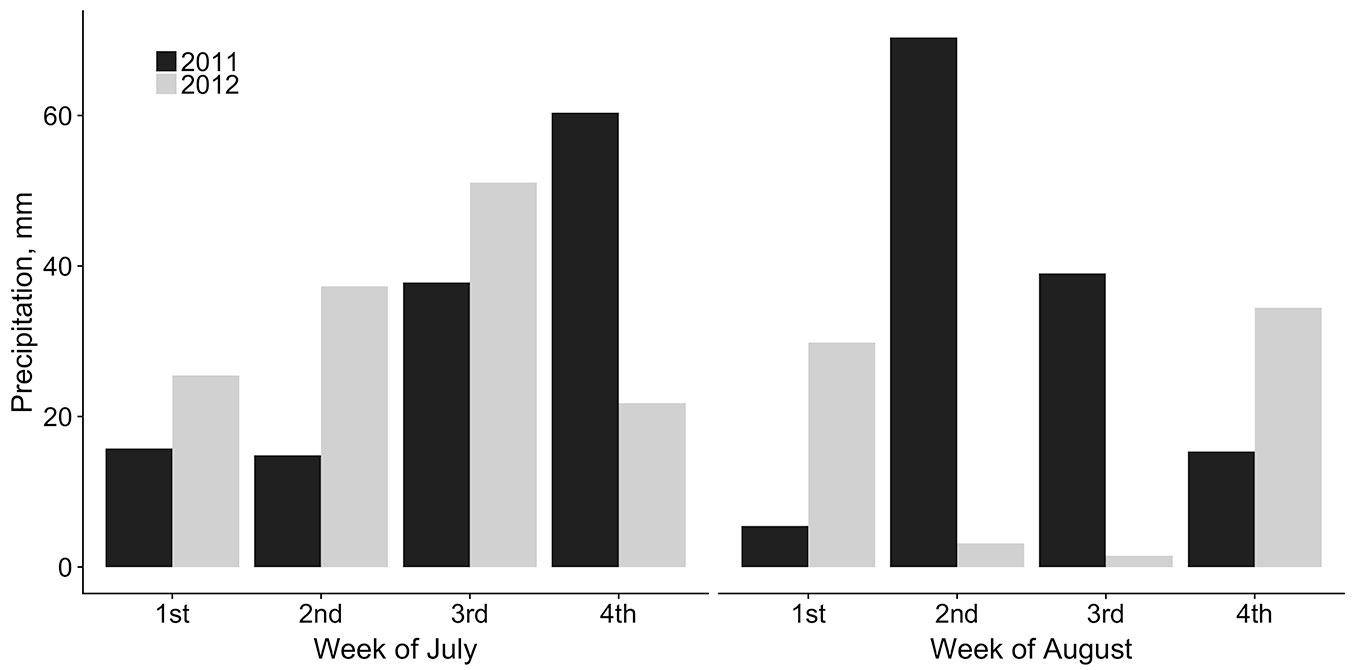
Minor changes in precipitation in the first week of August were observed in 2012 as compared to the last week of July (a slight increase in Dobele and decreases in Skriveri and Riga). In contrast, in 2011 (with a lower frequency of trees with lammas shoots), a sharp decrease in the amount of precipitation in the first week of August in comparison to the last week of July was observed (from 29 mm to 0 mm in Skriveri, from 46 mm to 3 mm in Riga, and from 106 mm to 14 mm in Dobele). In the second and third weeks of August, there was a lower amount of precipitation in 2012 than in 2011: 0.3-5 mm and 31-80 mm per week, respectively (Fig. 3).
Fig. 3 - Weekly amount of precipitation in July-August 2011 and 2012 (average of three meteorological stations - Riga, Dobele, and Skriveri; data: [28]).
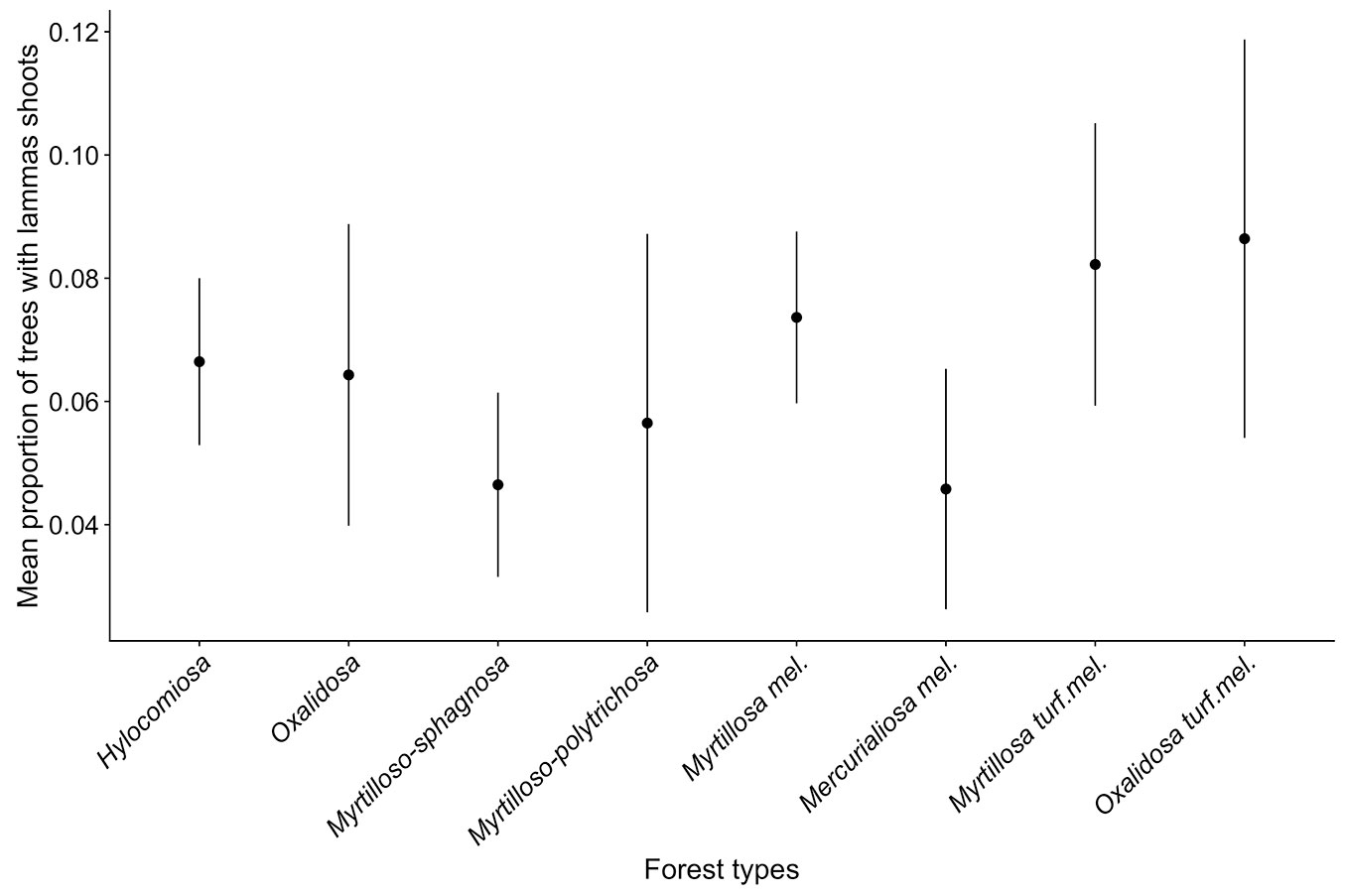
There was no significant effect of stand age on the proportion of lammas shoots (χ2 = 0.2175, p = 0.641) nor was there an age-related trend. The highest mean proportion of trees with lammas shoots was reached in 7-year-old stands, while the lowest value was observed in 6-year-old stands (Fig. 4). Notable variation in the proportion of trees with lammas shoots was detected for stands at different ages within a particular forest type; for example, in Myrtillosa mel. the proportions were 12.7%, 4.0%, and 10.8% at the ages of 4, 6, and 7 years, respectively. The effect of the forest type on the presence of lammas shoots was not significant (χ2 = 3.6892, p = 0.815). The highest and lowest proportion of trees with lammas shoots was detected in Mercurialiosa mel. and Oxalidosa turf. mel., respectively (Fig. 5). Also, the nteraction between forest type and stand age did not significantly affect the frequency of lammas shoots (χ2 = 5.0154, p = 0.6581).
Fig. 4 - Mean proportion of trees with lammas shoots (± 95% CI) in spruce stands of different ages.
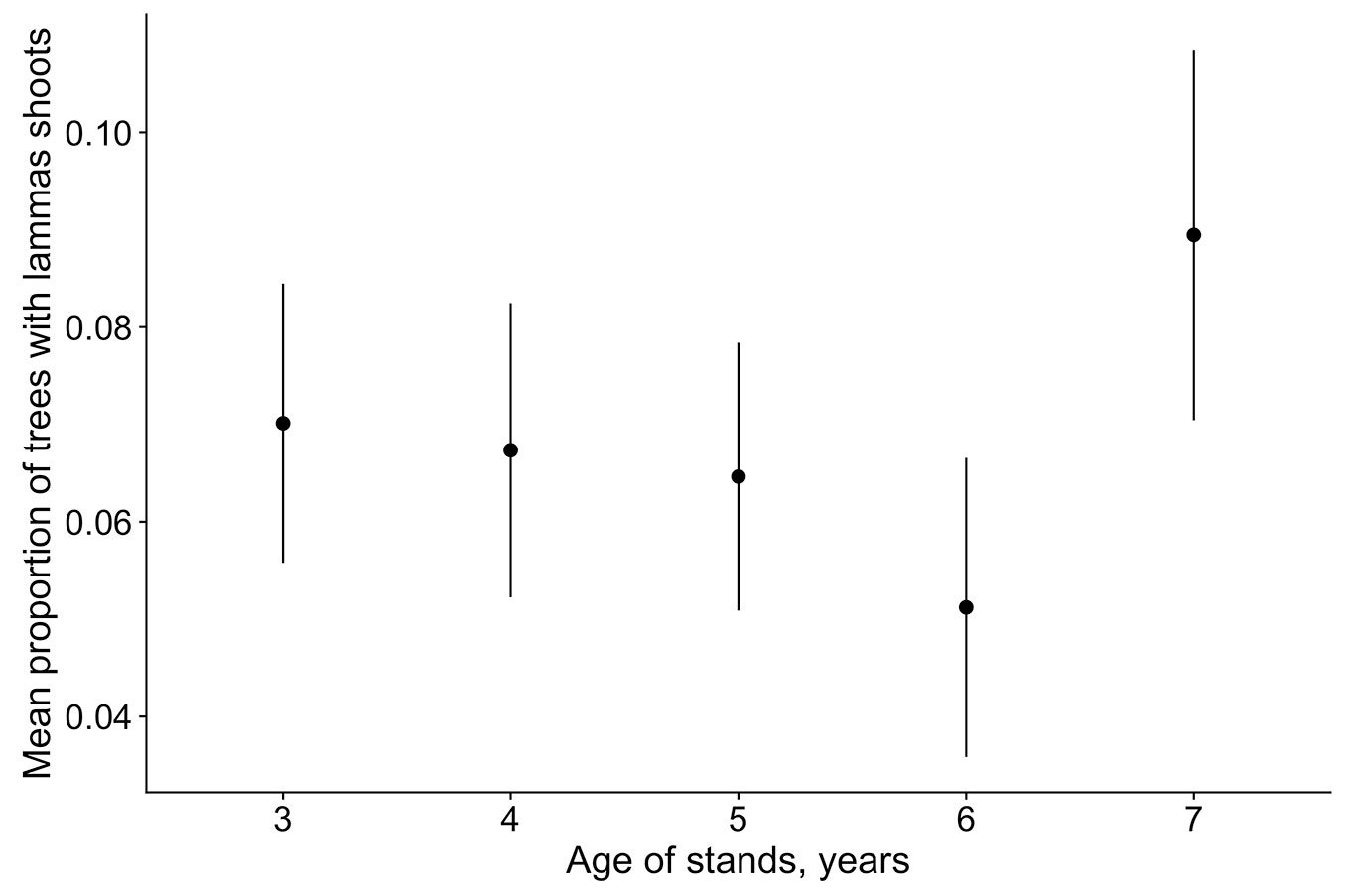
Fig. 5 - Mean proportion of trees with lammas shoots (± 95% CI) in spruce stands of different forest types.
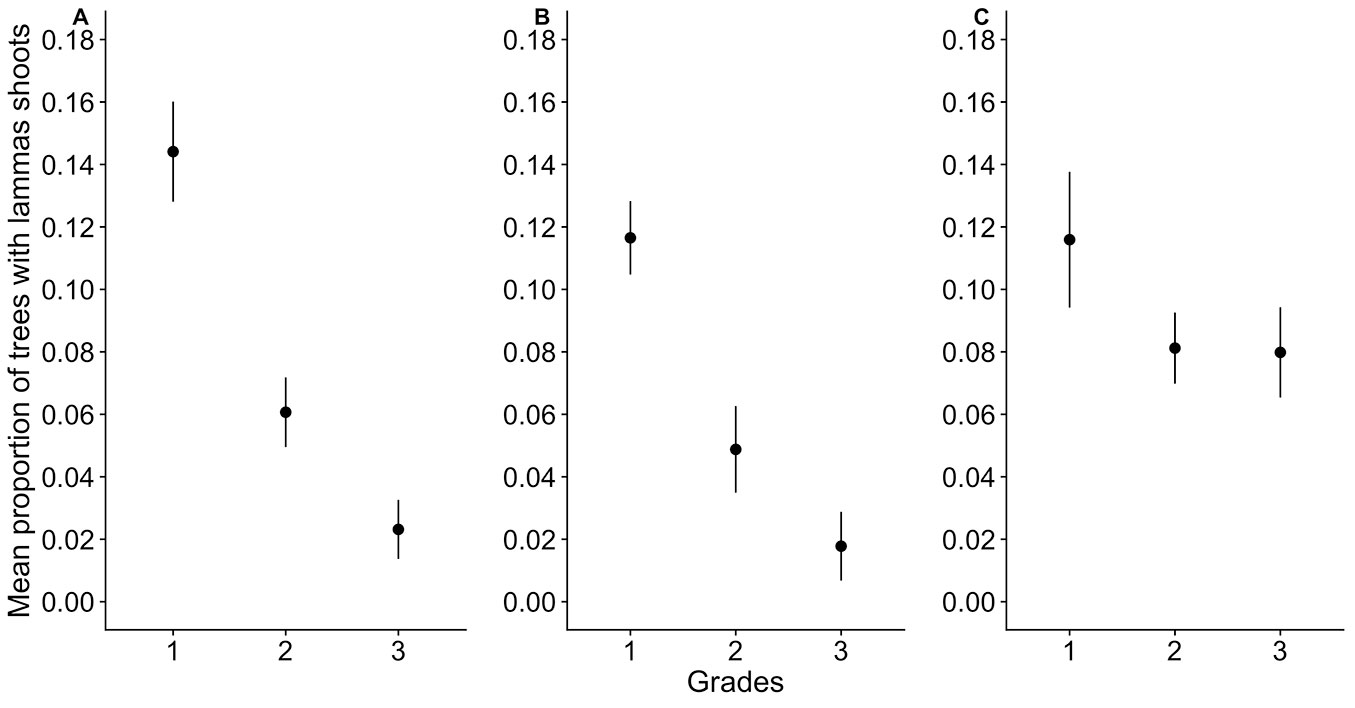
Micro-environmental factors had an important influence on the proportion of trees with lammas shoots (Fig. 6). The effect of competition from the surrounding vegetation and moisture regime was statistically significant (χ2 = 45.0218, p < 0.001). The proportion of trees showing lammas shoots in stands with no competition (Grade 1) was significantly higher (14.5%) in comparison to stands with medium (Grade 2) and high (Grade 3) competition (6.0% and 2.2%, respectively), being the difference between the two latter grades not significant (p = 0.066). Similarly, there was a significantly higher proportion of trees with lammas shoots (11.7%) in sites with a normal moisture regime (Grade 1) than in sites with periodic (Grade 2: 4.8%) or permanent (Grade 3: 1.7%) excessive moisture. Similar to that observed for competition, the difference between Grade 2 and Grade 3 of moisture was not significant (p = 0.184). Although the influence of the browsing damage was not statistically significant (p = 0.094), the observed trend was similar to that detected for competition and moisture regime. The proportion of trees with lammas shoots was 11.3%, 8.2%, and 7.7% in Grade 1, Grade 2, and Grade 3, respectively. The effect of the interactions (of the second or third level) was not significant, suggesting that each factor had independent influence on the formation of lammas shoots.
Fig. 6 - Mean proportion of trees with lammas shoots (± 95% CI) in spruce stands with different micro-environmental factors characterised by (A) competition of surrounding vegetation, (B) excessive moisture, and (C) browsing damage. Each factor was assessed in a three-grade scale, where (1) the negative effect of the factor on growth of the spruce was not observed, (2) the minimal negative effect of the factor was observed, and (3) the notable negative effect of the factor was observed.
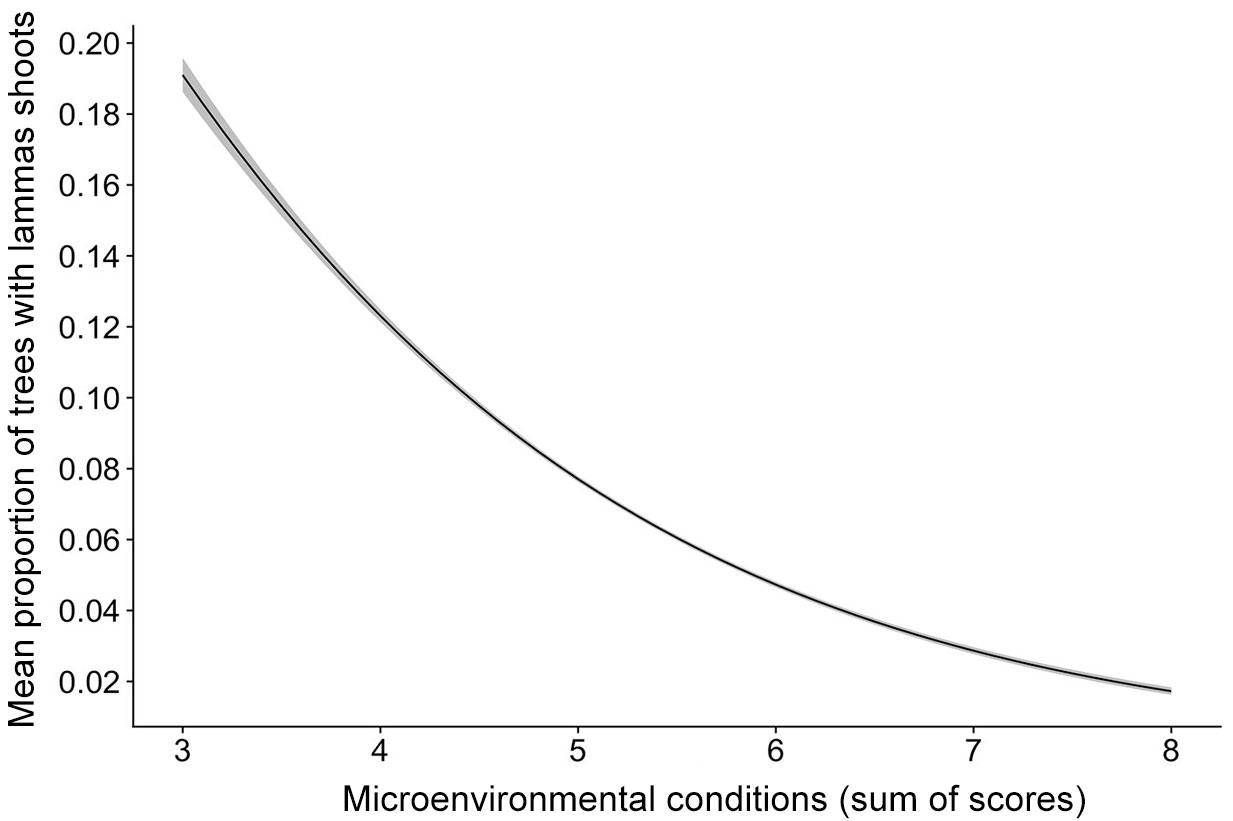
Finally, the combination of the scores for all micro-environmental factors (moisture, competition, and browsing pressure) revealed a clear significant trend (Fig. 7). The highest proportion of trees with lammas shoots occurred in the most favourable conditions (lower score). If at least one of the factors scored 3 in any stand (a notable negative influence), the proportion of trees with lammas shoots in that stand did not exceed 10%.
Fig. 7 - Mean proportion of trees with lammas shoots (shown by the black line) with the 95% CI (grey band on both sides of the line) in the Norway spruce stands analyzed in relation to the suitability of the growing conditions (x-axis), expressed as the sum of grades (scores) for three micro-environmental factors (competition, moisture, and browsing). For each factor, its negative effect on Norway spruce growth was quantified as: (1) not observed or negligible; (2) minimal; (3) notable.
Discussion
The average proportion of trees with lammas shoots in our study (7% ± 1%) was similar to that found for Norway spruce provenances from Baltic States at the age of five years in an experiment in Sweden ([9]). An even higher proportion of trees with lammas shoots was found in Norway, based on the analysis of the national forest inventory sample plots (located at an elevation up to 200 m a.s.l.). Moreover, Kvaalen et al. ([27]) reported that as many as 80% of trees had lammas shoots in two of the 58 sample plots analyzed.
The formation of predetermined and additional height increment is defined by meteorological conditions, such as temperature and precipitation, in combination with the photoperiod ([10], [1], [43], [14]). Skrøppa & Steffenrem ([38]) described the negative relationship between the presence of lammas shoots and the timing of shoot growth for full-sib families in a provenance trial of Norway spruce in the fifth growing season, pointing out that the occurrence of lammas shoots is generally positively associated with a favourable climate and soil fertility. Analysis of different coniferous tree species, such as red pine (Pinus resinosa Ait.), white pine (Pinus strobus L.), California red fir (Abies magnifica A. Mur.), and white fir (Abies concolor (Gord. and Glend.) Lindl.), suggests that abundant precipitation at the end of the vegetation period can stimulate the formation of lammas shoots ([5], [6], [29], [16]). This is in contrast to our results, where a lower proportion of trees with lammas shoots was observed in 2011 - a year with higher amounts of precipitation in the middle part of August in comparison to 2012, but only slightly higher than the long-term average and with an increase of precipitation only two weeks after the end of the predetermined height growth. After the completion of the predetermined height growth (second part of July), a sharp temperature increase was observed in the year 2012, along with the highest proportion of trees with lammas shoots. The results suggest that a relative change (not necessarily the mean values of any of the meteorological parameters temperature and precipitation) at the right period of the annual cycle of tree growth could trigger the formation of lammas shoots. In addition, there might be some indirect influence of spring temperatures since previous studies have suggested that earlier flushing trees are more likely to form lammas shoots ([30], [38]), presumably due to the longer period between the end of the predetermined growth and the drop in temperature in autumn. However, the observations in the current study were conducted in autumn, and the spring phenology data were not available.
A decrease in the frequency of formation of lammas shoots with tree age was observed in a number of studies ([5], [11], [1], [35], [43], [42], [39]). For example, in a Norway spruce trial in Norway, 38% of 5-year-old trees had lammas shoots, in contrast to 22% at 6 years and 17% at 7 years. This trend was similar in almost all represented provenances ([39]). In contrast, no age-related trend was observed in our study. Prolonging or expanding the study (adding one more inventory or older stands) could further improve our understanding of this phenomenon. After analysing the genetic and environmental variation of internodal and whorl branch formation (including whorls formed by lammas growth) for open-pollinated families of Norway spruce at the age of 27 years, Steffenrem et al. ([40]) pointed out that, the effects at the juvenile stage might be confounded with the year (meteorological) effect. Retrospective study has also demonstrated higher frequency of the occurrence of extra whorls formed by lammas growth in sites with higher soil fertility (assessed in terms of site indices). In other studies carried out on Norway spruce in south-eastern Norway, the forest type (i.e., availability of nutrients) was one of the major factors determining the proportion of trees with lammas shoots: by height index H40 = 11 and H40 = 23, the proportion of trees with lammas shoots was 5%-25% and 30%-90%, respectively ([39]). We did not find significant differences in the proportion of trees with lammas shoots between forest types representing growing conditions for more than 90% of the spruce-dominated stands in Latvia (data: [31]). However, the range of site indexes included in our study (roughly corresponding to H40 = 8-22) was smaller than that of the aforementioned study (H40 = 8-26 - [39]).
Our results showed that differences in the proportion of trees with lammas shoots (even within forest types or stand ages) were strongly linked to micro-environmental factors like moisture, competition, and browsing pressure. For example, in a 5-year-old stand belonging to the Hylocomiosa forest type with little evidence of tending and therefore a high competition of herbaceous plants and shrubs, as well as visible browsing damage on 22% of trees, only 1.8% of the individuals had lammas shoots, while the proportion was 25.5% in another 5-year-old stand of the same forest type with visible signs of regular tending and no browsing damage. Similar to our results, Roth & Newton ([36]) found a notable increase in the occurrence of lammas shoots for the Douglas fir after weed control.
Sporadic observations gathered during our study (conducted in 2011 and 2012) also suggest that an increase in frequency of lammas shoots might be linked not only to differences in weather conditions between the years but also to the effect of tending conducted in 2011. For example, in a 3-year-old Oxalidosa turf. mel. forest type stand, where vegetation control was done in the second part of the vegetation period of 2011, the proportion of trees with lammas shoots increased from 10% in 2011 to 26% in 2012. Similarly, in a 5-year-old Hylocomiosa stand, it increased from 18% in 2011 to 33% in 2012.
Browsing damage could notably reduce carbon accumulation capacity and growth of Norway spruce trees. However, it did not show as much pronounced effect on the formation of lammas shoots as the other two micro-environmental factors assessed in this study. Low average browsing pressure (average share of damaged trees 10.7%) and the intensity of damage - a notable effect on height increment was only observed in a few cases - could be the possible reasons for the limited influence of this factor.
For the preparation of recommendations for forest regeneration and tree breeding, further studies should address the issue of actual risks (mainly the formation of long-lasting stem defects) and the long-term positive relationship to growth which could be associated with the formation of lammas shoots.
Conclusions
The proportion of trees with lammas shoots in juvenile (from 3 to 7 years) Norway spruce stands ranged from 0% to 25% and was 6.5% on average. Improvement of micro-environmental conditions (intensive forest management) increased the proportion of trees with lammas shoots in juvenile Norway spruce stands, but there was no clear link between the occurrence of lammas shoots and the age of stands or the forest type.
Acknowledgement
The study was conducted in the project “Forest management risks: prognosis and minimization”, funded by JSC Latvijas valsts meži (Latvia).
References
Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Una Neimane
Baiba Dzerina
Mara Kitenberga
Janis Jansons
Aris Jansons
Latvian State Forest Research Institute “Silava”, Rigas Street 111, Salaspils, LV-2169 (Latvia)
Corresponding author
Paper Info
Citation
Katrevics J, Neimane U, Dzerina B, Kitenberga M, Jansons J, Jansons A (2018). Environmental factors affecting formation of lammas shoots in young stands of Norway spruce (Picea abies Karst.) in Latvia. iForest 11: 809-815. - doi: 10.3832/ifor2539-011
Academic Editor
Angelo Nolè
Paper history
Received: Jun 27, 2017
Accepted: Oct 08, 2018
First online: Dec 14, 2018
Publication Date: Dec 31, 2018
Publication Time: 2.23 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2018
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 44924
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 36736
Abstract Page Views: 3460
PDF Downloads: 3749
Citation/Reference Downloads: 4
XML Downloads: 975
Web Metrics
Days since publication: 2710
Overall contacts: 44924
Avg. contacts per week: 116.04
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2018): 7
Average cites per year: 0.88
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Large-scale spatial distribution of deer browsing damage to young tree plantations
vol. 14, pp. 34-40 (online: 14 January 2021)
Research Articles
Understory vegetation dynamics and tree regeneration as affected by deer herbivory in temperate hardwood forests
vol. 10, pp. 837-844 (online: 26 October 2017)
Research Articles
Patterns of genetic variation in bud flushing of Abies alba populations
vol. 11, pp. 284-290 (online: 13 April 2018)
Research Articles
Modelling the moisture status of habitats by using NDVI on the example of the Cerrado and Atlantic Forest biomes borderland (Brazil)
vol. 18, pp. 375-381 (online: 16 December 2025)
Research Articles
Case study of a new method for the classification and analysis of Dryocosmus kuriphilus Yasumatsu damage to young chestnut sprouts
vol. 5, pp. 50-59 (online: 10 April 2012)
Review Papers
Avoidance by early flushing: a new perspective on Dutch elm disease research
vol. 2, pp. 143-153 (online: 30 July 2009)
Research Articles
Changes in moisture exclusion efficiency and crystallinity of thermally modified wood with aging
vol. 12, pp. 92-97 (online: 24 January 2019)
Research Articles
Strong relationships between soil and vegetation in reference ecosystems of a riparian Atlantic rainforest in the upper Doce River watershed, southeastern Brazil
vol. 16, pp. 226-233 (online: 17 August 2023)
Research Articles
Oak often needs to be promoted in mixed beech-oak stands - the structural processes behind competition and silvicultural management in mixed stands of European beech and sessile oak
vol. 13, pp. 80-88 (online: 01 March 2020)
Research Articles
Damage assessment to subtropical forests following the 2008 Chinese ice storm
vol. 10, pp. 406-415 (online: 24 March 2017)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







