Strong relationships between soil and vegetation in reference ecosystems of a riparian Atlantic rainforest in the upper Doce River watershed, southeastern Brazil
iForest - Biogeosciences and Forestry, Volume 16, Issue 4, Pages 226-233 (2023)
doi: https://doi.org/10.3832/ifor4313-016
Published: Aug 17, 2023 - Copyright © 2023 SISEF
Research Articles
Abstract
Habitat loss and fragmentation have been impacting ecosystem services essential for human survival. The Brazilian Atlantic rainforest, a biodiversity hotspot, has suffered from historical deforestation and, more recently, from an environmental disaster caused by the Fundão dam collapse that released ore tailings drastically affecting a large territory in the Doce River watershed. This work aims to assess the relationships between soil properties and vegetation in a reference ecosystem to provide guidelines for restoration projects in areas affected by the dam collapse. We conducted phytosociological (vegetation characteristic) and soil quality studies in three distinct natural sites and studied different vegetation strata to better understand plant species composition in reference sites along the impacted Doce River and their potential role in community structuring and functioning. We recorded 140 species, 78 in the tree stratum, and 90 in the sapling stratum. Furthermore, our results highlight the influence of soil on floristic composition in the Atlantic rainforest. Small-scale edaphic variation influenced species composition in both sapling and tree strata. We also identified species of the same genus with strong association with the extremes of the edaphic gradient. Therefore, we highlight that studies in various regions along the Doce River watershed are of utmost importance to evaluate the association between species and soils. The particularities of the species are crucial to the effectiveness of restoration processes since this plant-soil correlation should not be extrapolated even within the same genus. This knowledge is of strategic relevance to provide scientific-based guidance for restoring these environments, aiming at the recovery of biodiversity and ecosystem services.
Keywords
Dam Breach, Fundão Dam Collapse, Reference Ecosystem, Restoration Ecology, Soil-vegetation Relationships
Introduction
The increasing loss and fragmentation of tropical forests have led to the simplification of these environments, substantially affecting key ecosystem services essential for human survival. The simplification of environments comes along with biodiversity loss, which affects healthy soil maintenance and the provision of clean water and air, as well as compromises livelihoods, food security, and general quality of life worldwide ([47], [17]). Native ecosystems have a high resilience when faced with mild disturbances and reduce the effects of climate change ([17]). However, the degradation of native ecosystems has affected their regenerative capacity and ecological functions. To mitigate these impacts and promote the restoration of degraded areas, the United Nations (UN) has declared the present decade (from 2021 to 2030) as “the decade of restoration” ([49]).
Through ecological restoration, it is possible to recover the environment functionally by re-creating habitats, and increasing diversity, biological integrity, and ecosystem services ([6], [43]). One of the main steps in restoration is the identification of a reference ecosystem, which serves as a guide for planning and measuring the success of the project, so the recovered environment must resemble what it was before degradation ([6], [20], [48]). In forest restoration, the goal is to create plant assemblages with similar species composition found in the reference area previously established ([42], [43]). To be effective, these restored plant assemblages must modulate environmental conditions and ecosystem services, which in turn favor the establishment of other native species ([43]).
The success of restoration projects is strongly tied up to the recovery and re-establishment of soil properties, as they shape ecological succession and accelerate the recovery of tropical forests ([18], [44], [22], [50]), with an important role in creating microhabitats essential for forest diversity and taxonomic composition ([15]). Land use history and disturbance intensity determine the availability of seeds and propagules, as well as the presence of dispersing agents, which guide the regenerative capacity of the system ([29]). Thus, the analysis of soil factors concomitant with phytosociological (vegetation characteristic) surveys is an indispensable tool for understanding the differences among communities and their functional attributes ([7]). Besides creating guidelines for appropriate conservation and restoration policies ([30], [14]), studies of soil-vegetation relationships help to define properly the reference ecosystems.
Although biodiversity studies have been growing in the last decades, environmental degradation keeps rising at concerning levels ([25]). Tropical forests have been drastically devastated. For instance, the Brazilian Atlantic rainforest, one of the 25 most biodiverse and threatened ecoregions of the world ([34], [27]), now comprises only fragments corresponding to abount 12% of its original area ([41]), although new estimates point to a larger native cover area (28% - [40]). Even the riparian forests that are protected by specific environmental legislation have suffered intense degradation and disturbance ([40]) and this scenario is worsening because of the loosening of the laws ([21]). Riparian forests are associated with water courses and have high relevance in the preservation of species and natural resources, ensuring the maintenance of ecosystem integrity ([21]). However, stretches of riparian formations associated with the Doce River and its main tributaries have been degraded due to their history of inadequate management ([37]). The environmental disaster caused by the rupture of the Fundão dam in the municipality of Mariana (MG) is another major vector of stress imposed on the vanishing Atlantic rainforest. The dam breach released tailings that drastically affected the entire riparian vegetation of the Doce River ([14], [35]). The tsunami of iron ore mud affected the “ecological memory” of this site ([14]) and the gradual and natural capacity for resiliency ([3]), therefore requiring bold ecological restoration actions.
The assessment of the relationships between edaphic properties and vegetation composition in reference areas of threatened ecosystems is a first step towards understanding the particular species and edaphic conditions that promote diversity in these ecosystems, helping to expand the regenerative potential of impacted areas ([14]). Our objective was to evaluate the relationship between floristic composition of the tree and shrub strata and small-scale edaphic variation in Atlantic forests, assessing whether soil variation can explain vegetation structure and composition in these two strata. Our hypothesis is that small-scale edaphic heterogeneity plays an important role in tree and sapling communities in riparian Atlantic Forest remnants in the upper Doce River watershed, southeastern Brazil.
Material and methods
Study area
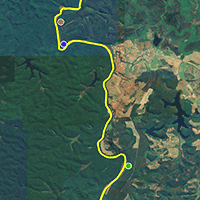
We carried out the study in three sites (Fig. 1) of riparian Brazilian Atlantic rainforest (Semideciduous Seasonal Forest) remnants located in the north of the municipality of Bom Jesus do Galho, district of Revés de Belém, state of Minas Gerais, southeastern Brazil (Fig. 1). The location of the three sampling sites were: Site 1: 19° 33′ 30.4″ S, 42° 31′ 25.6″ W; Site 2: 19° 34′ 32.2″ S, 42° 31′ 19.2″ W; Site 3: 20° 38′ 53.8″ S, 42° 28′ 50.0″ W. The study sites are located within the buffer zone of the Doce River State Park. Created in 1944, the park was the first state conservation unit in Minas Gerais. With an area of 36.000 ha, it is one of the largest Atlantic rainforest remnants in the country. According to the Köppen classification, the regional climate is categorised as Cwa, humid subtropical with dry winter (May to October) and hot rainy summer (November to April - [1]). The historical average annual temperature is 21.4 °C and the annual precipitation is 1325 mm ([1]). The soil use in Bom Jesus do Galho is predominantly anthropic, being occupied by crops and pasture, along with forest remnants ([26]). This municipality has been losing its native areas considerably. From 1985 to 2021 more than 38% of the native areas were lost, and the areas destined for monocultures and coffee plantations have had an increase of more than 4000 ha ([26]).
Fig. 1 - Map showing the location of three sampling sites in riparian Atlantic Rain Forest at the upper Doce River watershed, Bom Jesus do Galho-MG, southeastern Brazil.
Sampling vegetation and edaphic parameters
To characterize the vegetation structure, we used the fixed area plot sampling method proposed by Mueller-Dombois & Ellenberg ([33]). We selected three sites with a minimum distance of 2 km between them, with preserved riparian forest remnants. For the study of the tree stratum, we systematically demarcated 15 plots of 10 × 10 m (100 m2) in each sampling site, at least 10 m apart, totaling 45 plots. Within each plot, all woody individuals with DBH (diameter at breast height = 1.30 m above the ground) > 5 cm were inventoried. It is worth mentioning that all individuals of the tree stratum were woody species. We marked the sampled individuals with numbered aluminum plates attached by nails to their trunks. Samples of each plant individual were collected, identified with numbered adhesive tapes, and pressed to be later identified to the lower taxonomic level possible. Since many individuals were not flowering at the sampling time, several trips had to be done to the sites to collect material with reproductive structures for proper species identification.
For sampling the sapling stratum, we allocated one 5 × 5 m (25 m2) sub-plot in the lower left corner (watercourse direction) of each 10 × 10 m plot. Within each sub-plot, with the aid of a digital caliper, we inventoried all herbaceous and shrubby individuals with DAS (diameter at ground height) ≥ 1 cm and ≤ 5 cm. We marked the sampled individuals with numbered aluminum plates attached with nylon thread. Subsequently, we collected plant branches ideally with reproductive structures from all individuals for taxonomical identification. Sixteen species did not flower during sampling, which prevented taxonomic identification.
We treated the plant material collected for the tree and sapling strata according to conventional herborization techniques and deposited them in the Herbarium of Norte Mineiro (ICA-UFMG) and the Herbarium of Montes Claros Minas Gerais (Unimontes). The identification of the botanical material collected was done by consulting experts, using specialized literature, and comparing it with the existing exsiccata in these herbaria. We used the Angiosperm Phylogeny Group IV (APG IV 2016) system to classify the species into families. Synonymy verification, nomenclature, and species authors were obtained through the “WorldFlora” package ([23]) in the R environment ([39]), standardized according to World Flora Online (WFO - ⇒ http://www.worldfloraonline.org/). For species not found in WFO, we performed additional checks through the Flora e Funga do Brasil ([16]). We calculated the following parameters: absolute and relative values of density, dominance, frequency, and value of importance (IV). To calculate the IV, we used the average of the relative density, relative dominance, and relative frequency.
In each plot, at the four corners and the center, we collected 100 grams of soil obtained at 20 cm deep, then we mixed the five samples into one composite sample. Each composite sample represented a valid estimate of the average soil parameters. Each soil sample was shade dried, kept at room temperature, crumbled, completely homogenized, identified, and then sent for chemical and particle size analyses by the Soil Department of the Federal University of Viçosa. All soil granulometric analyses (coarse sand, fine sand, silt, and clay fractions) followed the description found in Donagemma et al. ([10]). The pH in water was measured using 1:2.5 (v/v) soil: solution ratios. The organic matter content was determined by the Walkley-Black method. The exchangeable cations Ca2+, Mg2+, and Al3+ were extracted using a 1 mol L-1 KCl solution. The Ca2+ and Mg2+ contents were determined in the extract by titration with EDTA 0.01 molc L-1, and the Al3+ contents by titration with NaOH 0.025 molc L-1, according to Silva et al. ([45]). The elements P, K, Zn, Fe, Mn, and Cu were extracted by the Mehlich 1 solution, the sulphur (S), by a solution of monocalcium phosphate in acetic acid, and the contents of these elements in the extracts were determined by spectrophotometry, according to Silva et al. ([45]). Potential acidity (H+Al) was extracted by a 0.5 mol L-1 calcium acetate solution at pH 7.0 and determined by alkalimetric titration of the extract ([45]). The base saturation (Sb) and aluminum saturation (SAl) were calculated, respectively, according to the following expressions (eqn. 1, eqn. 2):
Statistical analyses
To assess the adequacy of sampling effort, we constructed rarefaction curves for species diversity in R ([39]) using the function “rare_Rao” from package “adiv” ([36]), in which expected species diversity for each sampling site was computed as a function of the cumulative number of plots. We used the resampling approach with 10.000 iterations. We evaluated the pattern of exclusive and shared species among the three studied sites through Venn diagrams constructed for both tree and sapling strata using the package “VennDiagram” ([5]).
To determine the relationship between soil factors and the plant species community, we made a co-inertia analysis (COIA). This robust and flexible analysis measures the agreement between two multivariate data sets, also called co-structure ([9], [11]). We performed the COIA for the tree and sapling strata separately. The edaphic matrix was defined as the values of 17 soil factors in the 45 plots (15 per site), while the floristic matrix was defined as the incidence (presence and absence) of 78 tree species or 90 sapling species in the 45 plots. We removed rare species (species that appear only once in each stratum) from the COIA in order to reduce bias in the detection of relationships between community composition and edaphic factors ([28]).
An RV coefficient, which measures the strength of the association between the two matrices, was calculated from COIA outputs. The RV coefficient is bounded to 0 (i.e., no association) and 1 (i.e., maximum association). The significance (p-value) of the RV coefficient was obtained through Monte Carlo permutation, performed with 10.000 randomizations. To analyse the COIA, a Principal Components Analysis (PCA - mean = 0; standard deviation = 1) was used for the soil matrix, and a centered PCA (mean = 0) was used for each floristic matrix, according to Dray et al. ([11]). To attain the assumptions of normality in the soil data, we performed square root transformation for potassium (K), iron (Fe), and organic carbon (C), and logarithmic transformation for sulphur (S), copper (Cu), silt, and clay. To assess the association between each soil factor and COIA axis 1, we performed a Pearson’s correlation between soil values and plot coordinates on COIA axis 1. The association between species and the COIA axis 1 was defined by the species coordinates on this axis. The COIA was performed using the “ade4” package ([12]). All analyses were done in the R environment ([39]).
Results
In this study, we found 140 plant species belonging to 28 families. Seventy-eight species belonged to the tree stratum and 90 to the sapling stratum (see Tab. S1 and Tab. S2 in Supplementary material). The rarefaction curves shown in Fig. S2 indicate that the sampling of species diversity for the three sites at both tree (Fig. S2a-c) and sapling strata (Fig. S2d-f) was solid, since the expected species diversity shows a trend for stabilization.
In the tree stratum, the richest families were Fabaceae (10 species), followed by Moraceae (6 species), Meliaceae (5 species), and Myrtaceae (5 species). Eight families were represented by only one single species (Tab. S1 in Supplementary material). Regarding the importance value (IV), Meliaceae accounted for 38.9% of IV, followed by Fabaceae (9.7%), Petiveriaceae (9.1%), and Sapindaceae (8.5% - Tab. S1). The most important tree species was Guarea macrophylla Vahl (IV: 22.6%), followed by Guarea guidonia (L.) Sleumer (Meliaceae, 11.2%), Gallesia integrifolia (Spreng.) Harms (Petiveriaceae, 8.0%), and Cupania emarginata Cambess. (Sapindaceae, 3.6%) (Tab. S1). Seventeen species were not identified, and these unidentified species accounted for 7.0% of IV. These species did not flower during sampling, which prevented taxonomic identification.
In the sapling stratum, the richest families were Fabaceae (13 species), followed by Myrtaceae (12 species), Meliaceae (8 species), Sapindaceae (7 species), Lauraceae (6 species), and Moraceae (5 species). Eight families were represented by only one species (Tab. S2 in Supplementary material). Regarding the IV, the family Sapindaceae accounted for 20.0% of the IV, followed by Myrtaceae (18.3%), Lauraceae (12.1%), and Meliaceae (8.3% - Tab. S2). The most important sapling species was Cupania emarginata (IV: 11.2%), Aniba firmula Mez (Lauraceae, 10.6%), Eugenia ligustrina (Sw.) Willd. (Myrtaceae, 5.6%), Trichilia pallens C.DC. (5.0%), and Eugenia umbrosa O.Berg (Myrtaceae, 4.5% - Tab. S2). Sixteen species were not identified, and these unidentified species accounted for 3.9% of the IV.
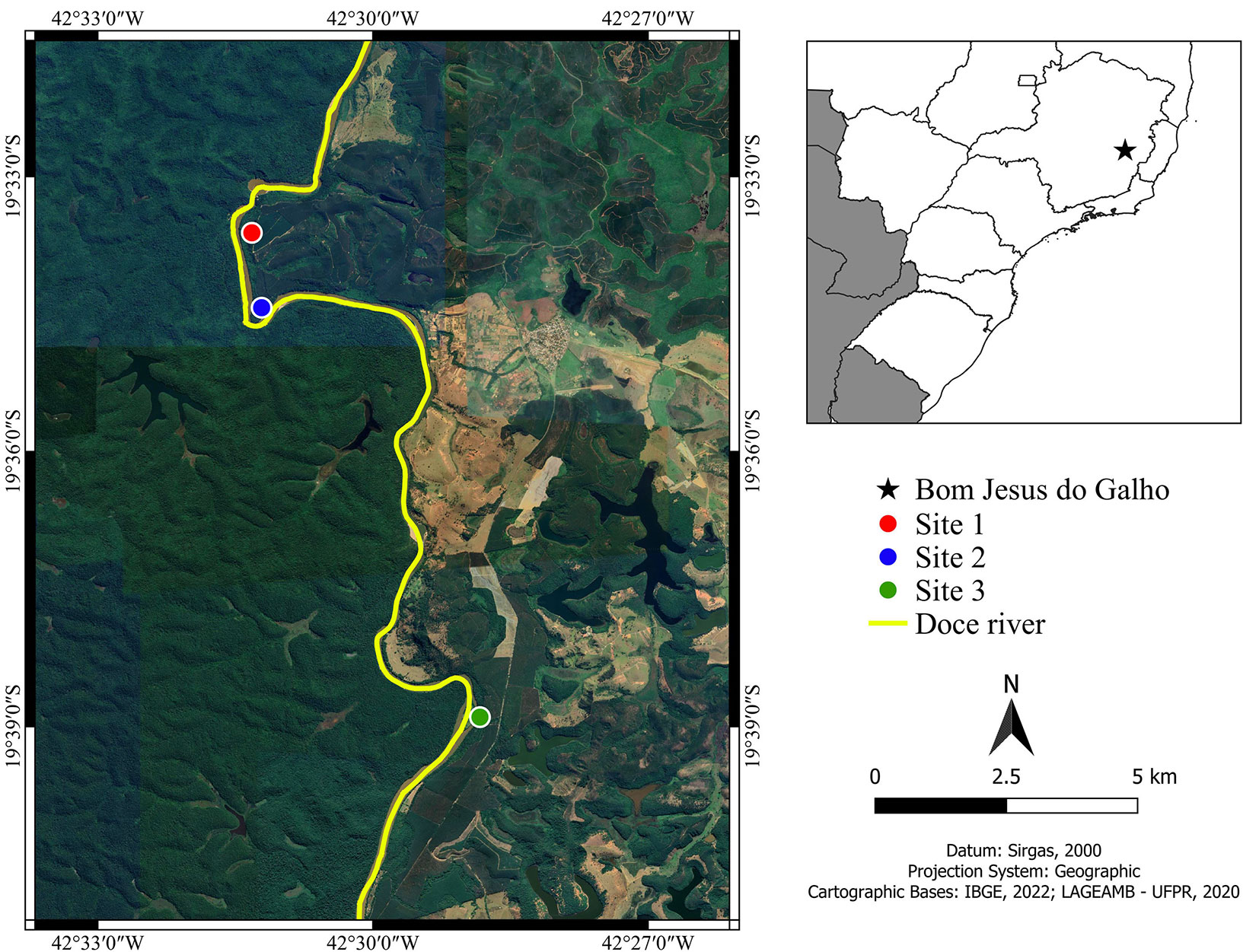
Two species of the tree stratum occurred at the three studied sites, Eugenia florida DC. (Myrtaceae), and Maclura tinctoria (L.) D.Don ex Steud. (Moraceae - Fig. 2a), and these two species stand out among the ten most important species from the tree stratum. However, most species were exclusive of each site. The number of exclusive tree species at sites 1, 2, and 3 was, respectively, 30 (75%), 16 (57%), and 17 (63%) (Fig. 2a). Regarding the sapling stratum, six species occurred at the three studied sites, A. firmula, Cu. emarginata, E. florida, Ga. integrifolia, Gu. macrophylla, and T. pallens (Fig. 2b), and except for Gu. macrophylla, these species stand among the ten most important species from the sapling stratum (Tab. S2). Similar to the tree stratum, the majority of sapling species were exclusive from each site. The number of exclusive sapling species at sites 1, 2, and 3 was, respectively, 37 (80%), 23 (70%), and 18 (62%) (Fig. 2b).
Fig. 2 - Venn diagram illustrating the exclusive and shared species among three studied sites from tree (a) and sapling strata (b) of riparian forests at the upper Doce River watershed, Bom Jesus do Galho-MG, southeastern Brazil.
The soils from the riparian Atlantic rainforests at the studied sites showed a considerable variation that indicated a high heterogeneity among the plots (Tab. S3 in Supplementary material). According to the chemical properties, the sampled soils were considered fertile, with the base saturation ranging from 49.3% to 84.6%, and the Al concentration was negligible. The pH was acidic (pH < 5.5) for 49% of plots and neutral for 51% of plots. The concentration of phosphorus ranged from 0.8 to 8.9 mg dm-3, the potassium ranged from 21 to 114 mg dm-3, and the organic carbon ranged from 0.1% to 3.0%. The iron content showed a wide variation, ranging from 10.3 to 438.1 mg dm-3, and the manganese ranged from 44.8 to 196.3 mg dm-3. Regarding the soil texture, there was also a wide variation in the sampling plots (Tab. S3). The proportion of fine sand ranged from 7.1% to 81.9%, while the proportion of clay ranged from 8.7% to 50.5%.
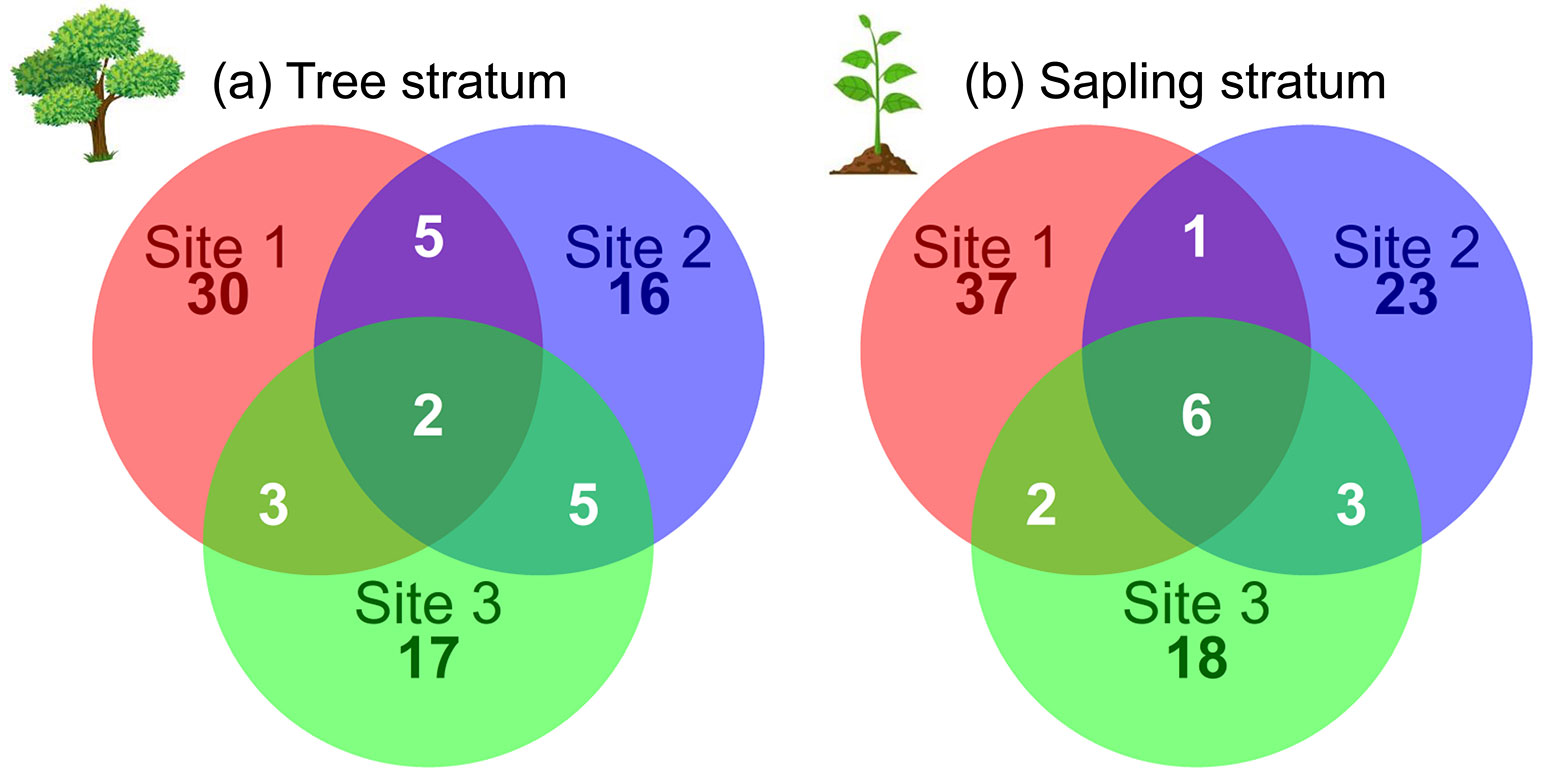
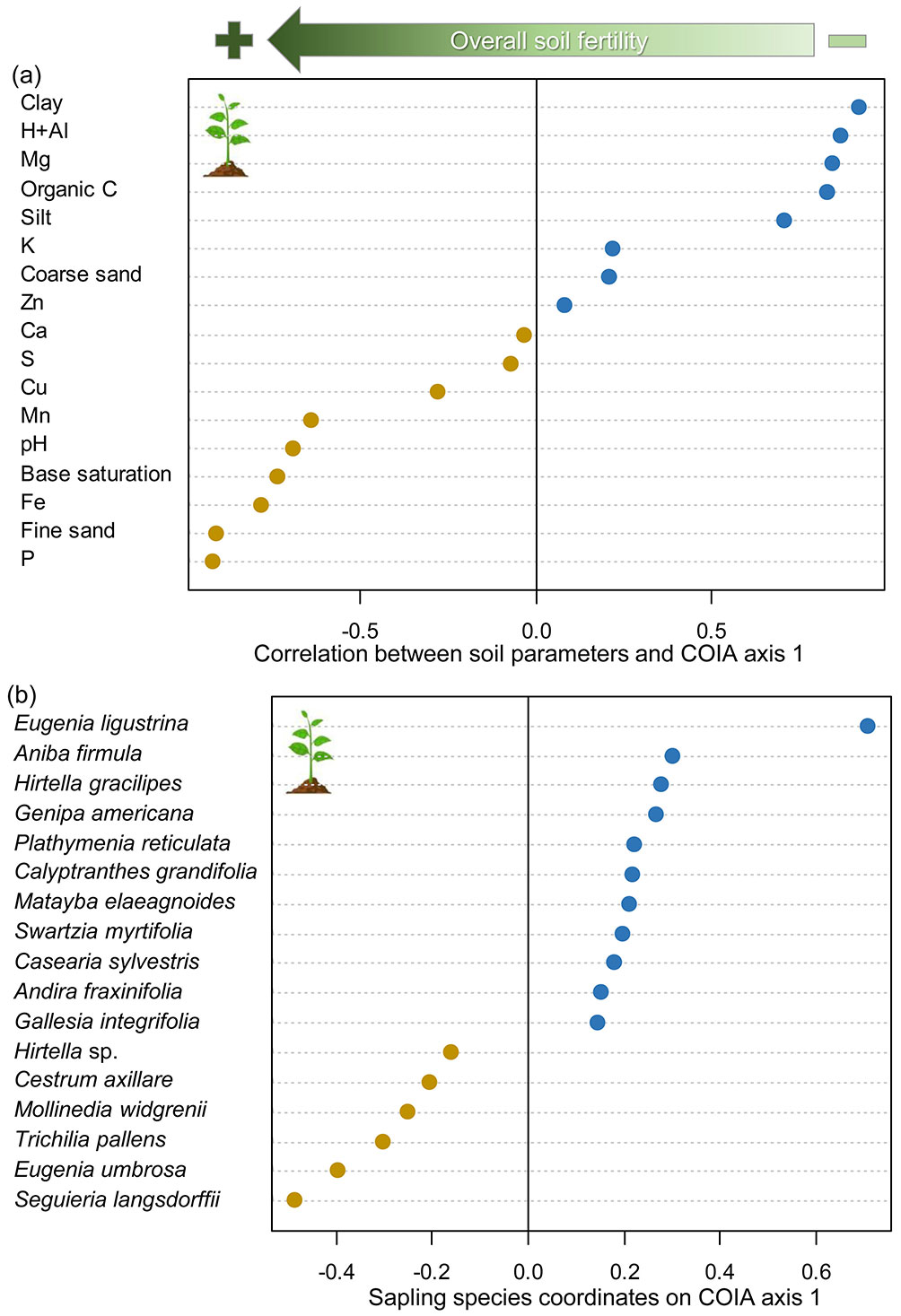
The co-inertia analysis (COIA) indicated a clear edaphic-floristic gradient at both tree and sapling strata. The overall association between tree species and edaphic factors was highly significant (RV = 0.445, p < 0.001), according to the COIA. We found a connection of 44.5% between the edaphic and tree floristic matrices. The percentage of covariance explained by the tree-COIA axis 1 was 70.8%, while axis 2 explained 17.5% of the covariance. Thus, we further explored only the tree-COIA axis 1. The positive side of the tree-COIA axis 1 showed plots with acidic soils with higher proportion of clay and silt, and higher concentration of magnesium and organic carbon (Fig. 3a). The tree species more strongly associated with the positive side of this axis were Gu. macrophylla, Cu. emarginata, T. pallens, Fabaceae sp.1, Macl. tinctoria, and Machaerium villosum Vogel (Fabaceae - Fig. 3b). On the other hand, the negative side of tree-COIA axis 1 showed plots with less acidic soils, nutritionally richer, with higher proportion of fine sand, and higher amounts of phosphorus, manganese, and iron (Fig. 3a). The tree species more strongly associated with the negative side of this axis were Gu. guidonia, Psychotria carthagenensis Jacq. (Rubiaceae), A. firmula, Cu. oblongifolia, Clarisia ilicifolia (Spreng.) Lanj. & Rossberg (Moraceae), and Seguieria langsdorffii Moq. (Petiveriaceae - Fig. 3b).
Fig. 3 - Co-structure between edaphic factors and tree species community sampled at three sites from riparian forests at the upper Doce River watershed, Bom Jesus do Galho-MG, southeastern Brazil. (a): Pearson’s correlation between edaphic factors and plot coordinates on co-inertia analysis (COIA) axis 1; (b) coordinates of tree species with the highest associations with negative and positive sides of COIA axis 1. Green arrow on the top indicates the overall direction of the soil fertility gradient. Orange and blue circles represent, respectively, negative and positive values of correlation (a) or coordinates (b) on COIA axis 1.
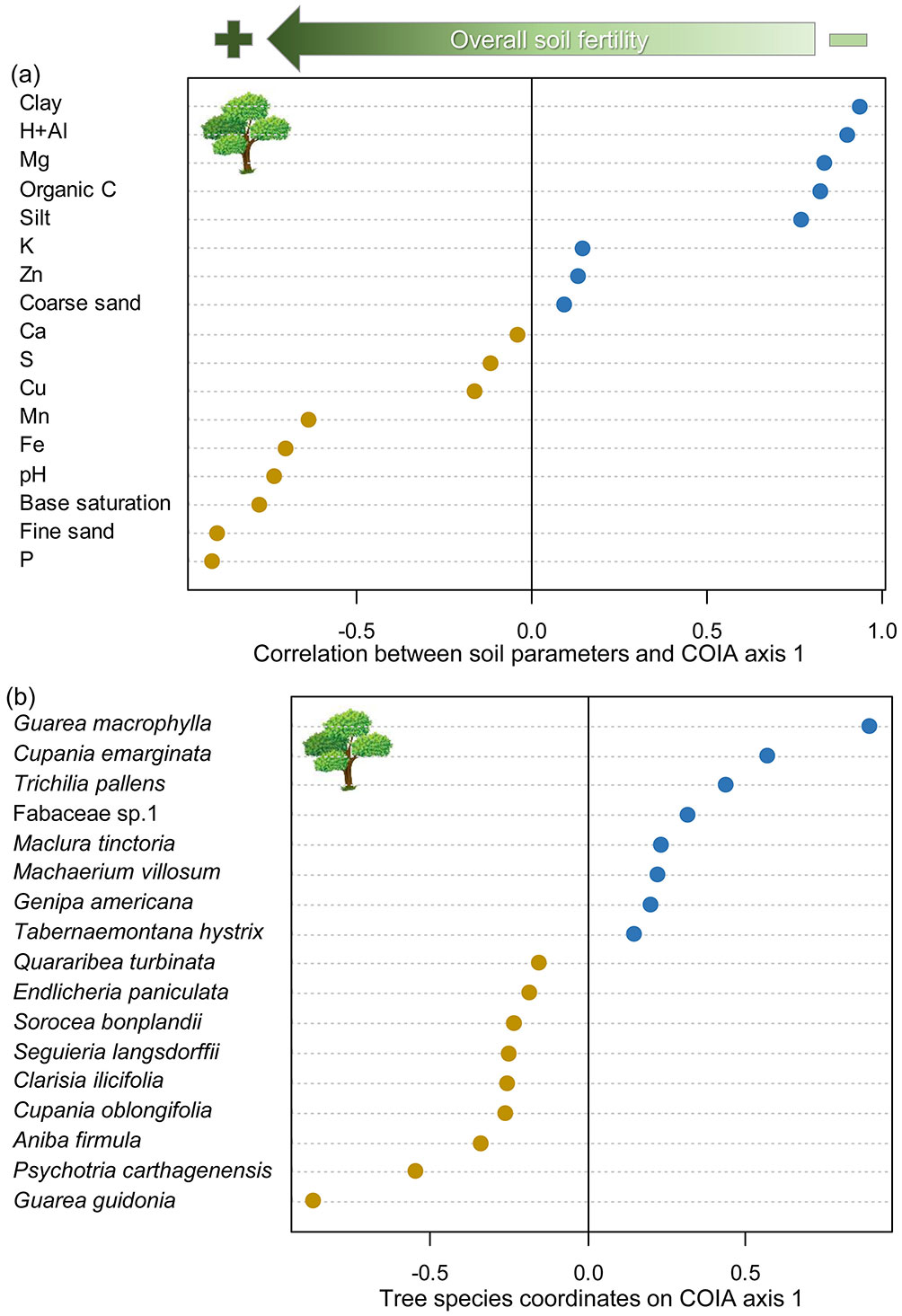
Regarding the sapling stratum, the overall association between sapling species and edaphic factors was also highly significant (RV = 0.357, p < 0.001) according to the COIA. We found a connection of 35.7% between the edaphic and sapling floristic matrices. The percentage of covariance explained by COIA axis 1 was 53.2%, while axis 2 explained 29.2% of the covariance. Thus, similar to the tree-COIA, we further explored only the sapling-COIA axis 1 for the sapling stratum. The positive side of the sapling-COIA axis 1 indicated a very similar pattern as the tree-COIA, showing plots with acidic soils with a higher proportion of clay and silt, and higher content of magnesium and organic carbon (Fig. 4a). The sapling species more strongly associated with the positive side of this axis were E. ligustrina, A. firmula, Hirtella gracilipes (Hook.f.) Prance (Chrysobalanaceae), Genipa americana L. (Rubiaceae), Plathymenia reticulata Benth. (Fabaceae), and Calyptranthes grandifolia O.Berg (Myrtaceae) (Fig. 4b). On the other hand, like the tree-COIA, the negative side of sapling-COIA axis 1 showed plots with less acidic soils, nutritionally richer, with higher proportion of fine sand, and higher amounts of phosphorus, iron, and manganese (Fig. 4a). The sapling species more strongly associated with the negative side of this axis were S. langsdorffii, E. umbrosa, T. pallens, Mollinedia widgrenii A.DC. (Monimiaceae), and Cestrum axillare Vell. (Solanaceae) (Fig. 4b).
Fig. 4 - Co-structure between edaphic factors and sapling species community sampled at three sites from riparian forests at the upper Doce River watershed, Bom Jesus do Galho-MG, southeastern Brazil. (a): Pearson’s correlation between edaphic factors and plot coordinates on co-inertia analysis (COIA) axis 1; (b) coordinates of sapling species with the highest associations with negative and positive sides of COIA axis 1. Green arrow on the top indicates the overall direction of the soil fertility gradient. Orange and blue circles represent, respectively, negative and positive values of correlation (a) or coordinates (b) on COIA axis 1.
Discussion
The Doce River basin, which has experienced a long historical degradation process ([42]) and more recently the large-scale drastic dam collapse, is still under successive waves of iron ore tailings due to the marked seasonality of the region. These affected riparian forests show no signs of natural regeneration ([14]). Thus, it is essential to select multiple reference ecosystems near the sites to be restored, encompassing abiotic factors necessary to accelerate the succession of the plant communities ([13]).
Our results highlight the strong influence of soil properties on the floristic composition of well-preserved Atlantic Rainforest sites studied here. Small-scale edaphic variation contributed significantly to explaining sapling and tree species composition. Soil conditions are among the most important factors shaping plant species composition ([7], [13], [22], [15]). We observed a soil gradient in which areas with a higher concentration of clay, organic carbon, and silt were distinct from areas with proportionally more phosphorus, fine sand, and less acidic soils. In this sense, the tree species with the highest value of importance were Guarea macrophylla followed by Guarea guidonia. These were also the species most strongly associated with the extremes of the soil quality gradient, as shown by our co-inertia analysis. Guarea macrophylla is a species commonly recommended in restoration processes of riparian Atlantic Rainforests for its dense canopy ([4]). We found that Guarea macrophylla had a strong association with poorer, more acidic soils with higher clay and silt ratios. Moreover, Guarea macrophylla is pollinated mainly by moths and coleoptera ([46]), and its seeds are dispersed by birds ([38]), thus promoting greater functional diversity in these areas. On the other hand, the species Guarea guidonia is a species of the secondary stage of succession ([8]), which may explain the strong association with less acidic and richer sandy soils of the edaphic gradient. This species has high crown density, biotic seed dispersal, and is tolerant to seasonal flooding ([4]). Similar to the genus Guarea, the congeneric species Cupania oblongifolia and Cupania emarginata from the tree stratum, and the sapling species Eugenia ligustrina and Eugenia umbrosa were also at opposite ends of the edaphic gradient. This niche differentiation observed in these congeneric species may reflect selective pressures acting on an evolutionary scale, such as competition, which led to the specialization of the species to distinct tolerable environments ([51]). Thus, we emphasize that what was observed in our results should not be extrapolated to other representatives of the genus without first understanding each particularity.
Recovery techniques should encompass various plant formations. The exclusive use of tree species may not adequately restore biodiversity in tropical forests ([19]). Life forms from other strata may provide resources in short time intervals for mutualistic and antagonistic animals ([32]) boosting the recovery of ecosystem functions. In this study, we did not find high plant species diversity in the evaluated areas (140 species, of which 78 occur at the tree stratum, and 90 at the sapling stratum), although it is worth noting that most of the sampled species occur uniquely in each site. Therefore, more studies need to be developed along the Doce River watershed, to better understand its plant diversity and community changes along the river. In the area affected by the collapse of the Fundão dam, 166 threatened species were described ([24]), which is not a surprising number, since this catastrophe affected one of the world’s biodiversity hotspots that is extremely endangered. The possibility of high turnover rates of plant species composition along the Doce River from its origin to its end at the Atlantic Ocean represents information of major relevance to the ecological restoration of biodiversity and ecosystem services in the region.
In both vegetation strata studied, the family Fabaceae had higher species richness, which was expected as this family is one of the most abundant and rich in tropical rainforests ([4], [18], [31]) as well as in forest remnants in the Doce River watershed ([15]). This group has representative species that are characteristic of the initial restoration processes ([18]). These are fast-growing native species adapted to poor soils, performing the function of quickly restoring the initial structure of the forest, allowing rapid ground cover, and avoiding the appearance of exotic species, which is one of the main problems for the success of forest restoration ([42], [3]). The family Myrtaceae was the second most diverse in the sapling stratum. Species of Myrtaceae are also often found in early succession processes ([4], [2]). In addition, species of this family promote the maintenance of the forest structure enabling greater functional diversity in the system, such as attracting pollinators and dispersers ([42], [49]). In this study, the species with the highest importance value sampled in both sapling and tree strata are dispersed by animals. Similarly, the species that occurred at the three sites in both strata also show zoochoric dispersal, except for Gallesia integrifolia, which is wind-dispersed. Therefore, including species with this type of dispersal should favor the reestablishment of ecological interactions important for the continuity and acceleration of successional processes ([4], [42]) and should be prioritized in the selection of species to be planted.
Conclusion
Our results confirmed that small-scale edaphic variation contributes significantly to explaining sapling and tree species composition. In addition, we identified species from both extremes of the edaphic gradient of the reference areas and highlighted the importance of studies in several regions of the Doce River watershed to evaluate the association among species and soils. The particularities of the species are crucial to the effectiveness of restoration processes since this plant-soil correlation should not be extrapolated even within the same genus. Neglecting that the floristic composition is a mosaic shaped by intrinsic local conditions can create erroneous knowledge to be used in restoration policies, which can be costly and inefficient, besides not restoring biodiversity and ecosystem services.
Acknowledgments
We thank Alysson R. Pereira, Vanessa M. Gomes, Suely A. Adriano, José G.S. Neves, Marly A. de Ávila, Cristina P.J. Veloso, Ricardo R. Fernandes, Michel Jacoby, Samuel Novais, and Júlio S. Lopes for their support in the fieldwork, and Heloisa P. Dinsmore for language review. We also thank two anonymous reviewers for their contributions in earlier versions of this manuscript, and logistical support by Instituto Chico Mendes de Conservação da Biodiversidade (ICMBio), Cmara Técnica de Conservação e Biodiversidade (CTBio), and Instituto Estadual de Florestas (IEF). This research was supported by Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG; APQ-03622-17, APQ-00031-19), and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).
The authors declare no conflict of interest with the content of this work.
References
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Online | Gscholar
CrossRef | Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Daniel Negreiros 0000-0002-4780-2284
Bárbara Stephanie Silva Ferreira
Yumi Oki 0000-0003-1268-9151
Wénita De Souza Justino 0009-0001-6527-493x
Ramiro Aguilar 0000-0003-4741-2611
G Wilson Fernandes 0000-0003-1559-6049
Ecologia Evolutiva & Biodiversidade, Departamento de Genética, Ecologia e Evolução/ICB, Universidade Federal de Minas Gerais, 31270-901, Belo Horizonte, Minas Gerais (Brazil)
Departamento de Biologia Geral, Centro de Ciências Biológicas e da Saúde, Programa de Pós-Graduação em Biotecnologia - PPGB, Universidade Estadual de Montes Claros, 39401-089, Montes Claros, Minas Gerais (Brazil)
Department of Biological Sciences, Institute of Environment, Florida International University, Miami, FL 33199 (USA)
Departamento de Ciências Florestais, Universidade Federal de Lavras, CP 3037, 37200-000, Lavras, Minas Gerais (Brazil)
Instituto Multidisciplinario de Biología Vegetal, Universidad Nacional de Córdoba - CONICET, CC 495, 5000 Córdoba (Argentina)
Programa de Pós-Graduação em Botnica Aplicada, Departamento de Biologia Geral, Universidade Estadual de Montes Claros, 39401-089, Montes Claros, Minas Gerais (Brazil)
Corresponding author
Paper Info
Citation
Ramos L, Negreiros D, Ferreira BSS, Figueiredo JCG, Paiva DC, Oki Y, Justino WDS, Santos RMD, Aguilar R, Nunes YRF, Fernandes GW (2023). Strong relationships between soil and vegetation in reference ecosystems of a riparian Atlantic rainforest in the upper Doce River watershed, southeastern Brazil. iForest 16: 226-233. - doi: 10.3832/ifor4313-016
Academic Editor
Michele Carbognani
Paper history
Received: Jan 22, 2023
Accepted: Jun 03, 2023
First online: Aug 17, 2023
Publication Date: Aug 31, 2023
Publication Time: 2.50 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2023
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 25029
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 18251
Abstract Page Views: 3932
PDF Downloads: 2395
Citation/Reference Downloads: 5
XML Downloads: 446
Web Metrics
Days since publication: 1062
Overall contacts: 25029
Avg. contacts per week: 164.97
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2023): 6
Average cites per year: 2.00
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Effect of restoration methods on natural regeneration in the Brazilian Atlantic Forest
vol. 18, pp. 23-29 (online: 15 February 2025)
Review Papers
The role of spatial criteria in enhancing forest restoration actions: a systematic review
vol. 19, pp. 157-167 (online: 11 May 2026)
Research Articles
A multisource approach helps to detect a forest as a reference site in an intensively used rural landscape (Uckermark, NE Germany)
vol. 14, pp. 426-436 (online: 21 September 2021)
Research Articles
Can the dynamics of forest restoration reduce landscape fragmentation in the Atlantic forest?
vol. 18, pp. 61-68 (online: 04 April 2025)
Research Articles
Soil fauna communities and microbial activities response to litter and soil properties under degraded and restored forests of Hyrcania
vol. 14, pp. 490-498 (online: 11 November 2021)
Review Papers
Shaping the multifunctional tree: the use of Salicaceae in environmental restoration
vol. 6, pp. 37-47 (online: 21 January 2013)
Research Articles
Nutrient cycling indicators in different models of successional groups and spacing for forest restoration
vol. 19, pp. 61-68 (online: 23 February 2026)
Research Articles
Changes in vegetation diversity and composition following livestock removal along an upland elevational gradient
vol. 8, pp. 582-589 (online: 22 April 2015)
Research Articles
Historical fire ecology and its effect on vegetation dynamics of the Lagunas de Montebello National Park, Chiapas, México
vol. 14, pp. 548-559 (online: 01 December 2021)
Research Articles
Outplanting performance of three provenances of Quillaja saponaria Mol. established in a Mediterranean drought-prone site and grown in different container size
vol. 13, pp. 33-40 (online: 21 January 2020)
iForest Database Search
Google Scholar Search
Citing Articles
Search By Author
- L Ramos
- D Negreiros
- BSS Ferreira
- JCG Figueiredo
- DC Paiva
- Y Oki
- WDS Justino
- RMD Santos
- R Aguilar
- YRF Nunes
- GW Fernandes
Search By Keywords






