
A comparative study of growth and leaf trait variation in twenty Cornus wilsoniana W. families in southeastern China
iForest - Biogeosciences and Forestry, Volume 10, Issue 5, Pages 759-765 (2017)
doi: https://doi.org/10.3832/ifor2288-010
Published: Sep 02, 2017 - Copyright © 2017 SISEF
Research Articles
Abstract
To investigate the genotypic differences associated with the growth potential and leaf traits of Cornus wilsoniana W., we planted twenty C. wilsoniana families in southeastern China and analyzed nineteen leaf morphological and physiological traits that have potential relationships with growth. Seedling growth and leaf traits exhibited high variability among the C. wilsoniana families. The phenotypic coefficients of variation (CVs) of these traits varied from 5.33% (leaf length/leaf width, LL/LW) to 23.17% (stomatal conductance, gs), and their heritabilities (H2) ranged from 0.17 (chlorophyll a/chlorophyll b, Chla/Chlb) to 0.75 (stem height, H and Chla). There was greater genetic variation in the physiological traits than in the morphological traits. H was significantly positively correlated with instantaneous water use efficiency (WUE), Chla, Chlb and total Chl, and diameter (D) was significantly positively correlated with net photosynthetic rate (Pn), gs, WUE, Chla, Chlb and total Chl and was negatively correlated with leaf phosphorus (LP). Based on cluster analysis, three families were selected as superior families for the study area due to their seedling growth and leaf traits. These results indicate that Pn, Chla, Chlb and total Chl are good indicators to use for selecting superior families of C. wilsoniana with better growth performance; additionally, high WUE and low LP are also critical leaf traits for cultivar selection because plant adaptation to environmental conditions is important for growth performance.
Keywords
Plant Growth, Leaf Traits, Intraspecific Variation, Genetic Heritability
Introduction
Cornus wilsoniana Wangerin is a deciduous tree that is native to South and West China. C. wilsoniana is not only a high-quality wood tree but also a biofuel tree with high yield and high oil content (the fruits contain 33-36% oil - [36], [15], [9]). Recently, this multi-functional tree species has rapidly developed in South China, and its area will increase to 2 million hectares by 2020 ([16]).
Cultivar selection is an effective method for increasing wood and fruit yield. Genetic diversity is the foundation of plant breeding. Previous studies have shown that intraspecific variation is lower than interspecific variability ([11], [12]). In fact, traditional community ecology has mainly focused on interspecific variation, and little work has involved intraspecific variation ([45], [42]). However, many studies have shown that there is high variability at the intraspecific level ([29], [10]), provenance level ([26], [32]) and at the family or clone level ([19], [48]).
Variation in leaf traits is an important component of plant phenotypic plasticity, and it is determined by genetics and the environment ([5], [3], [43]). Leaves are the main organs for photosynthesis and are tightly linked to plant biomass production; they are the main exchange platform for water, energy and carbon between the plant and the atmosphere. Leaf traits can reflect plant survival strategies to address environmental changes, and they also reflect plant adaptation strategies to maximize the harvest of carbon. Therefore, understanding variation in leaf traits not only has importance in ecological and biological evolution but also has great significance for plant growth and production. Some leaf traits are good indicators to use in selecting tree species with good growth performance and high biomass productivity ([1]). Among various leaf traits, leaf area, gas exchange, and intrinsic water use efficiency have been shown to be critical variables in determining plant biomass productivity ([18], [4], [8]). Other leaf traits, such as leaf nitrogen concentration and specific leaf area, are sometimes correlated with plant biomass and productivity, which are influenced by tree species, tree age, and growth conditions ([20]). However, the majority of these results were obtained from fast-growing trees, such as poplar and willow, and some studies were conducted in greenhouses with sufficient water and nutrient supplies. The stability of the relationships between intraspecific variation and leaf traits must be tested using much wider ranges of tree species and environmental conditions.
Although C. wilsoniana has recently received more attention and is well documented in silviculture and oil analysis, cultivar selection and the relationship between growth and leaf trait variation in cultivars are less well studied. The objectives of this study were to (1) compare the variation in growth and leaf traits among twenty C. wilsoniana families, (2) determine whether leaf traits can predict seedling growth performance, and (3) select superior families for the study area.
Materials and methods
Site description
The study was conducted at the Yangkou Forest Farm (117° 55′ E, 26° 47′ N, mean altitude 282 m), located in the north of Fujian Province, China. The average annual temperature in the study area is 18.5 °C, the average annual precipitation is 1880 mm, and the frost-free period is 280 days. The study area has a subtropical marine monsoon climate. The average slope is 15% in the experimental area. The soil texture in the study area is clay loam, and the pH is 4.5-4.8. The average contents of soil organic matter, soil available nitrogen and soil available phosphorus are 22.53 g kg-1, 172.71 mg kg-1, and 1.66 mg kg-1, respectively.
Experimental design
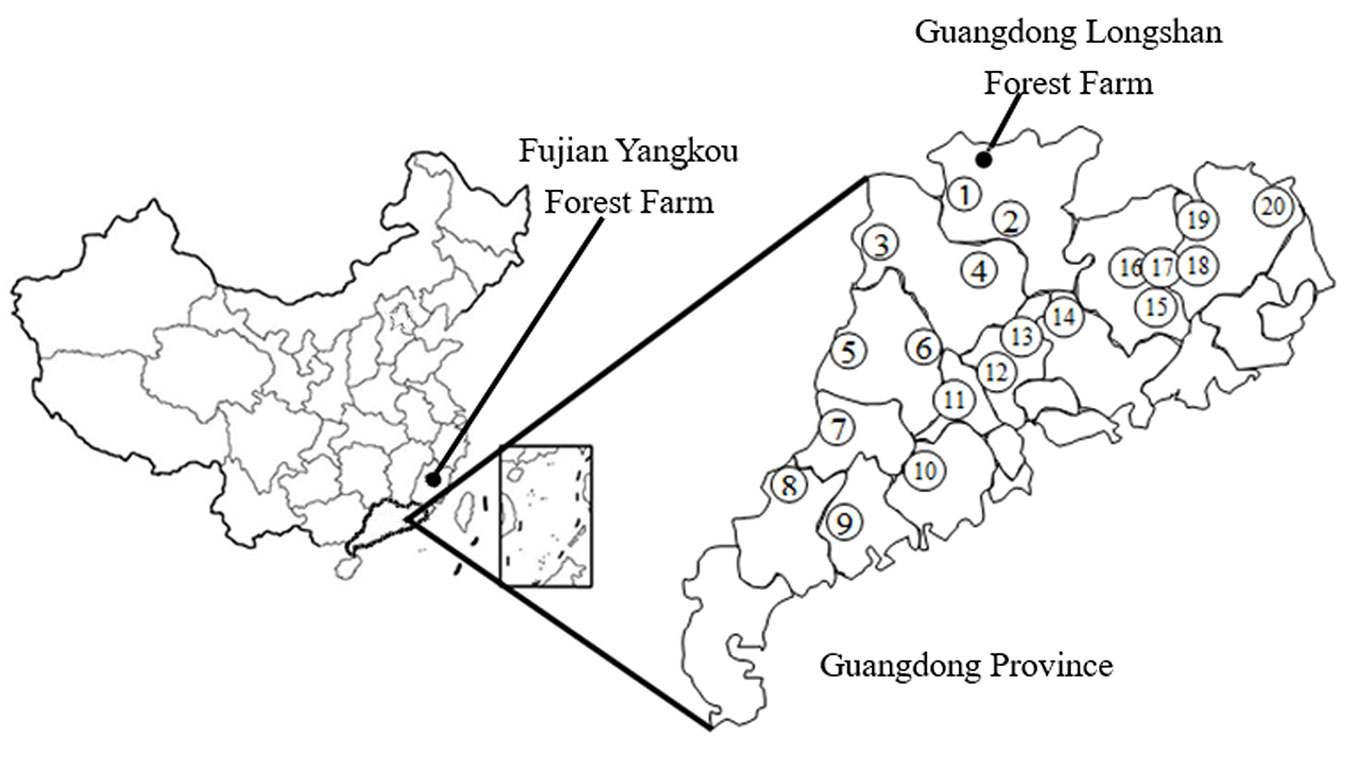
The experiment was performed in March 2013 using a randomized block design and included three blocks. Twenty C. wilsoniana families were randomly planted in the subplots of each block using 2 × 3 m spacing, and there were approximately 100 2-year-old seedlings in each family. The seedlings of the twenty families came from twenty plus trees in Guangdong Province. The seeds of these plus trees were collected and sown in the nursery of Guangdong Longshan Forest Farm. The sites of the selected plus trees, Longshan Forest Farm, and the experimental area are marked on the map in Fig. 1.
Fig. 1 - Map of China showing the location of the experimental area (Fujian Yangkou Forest Farm), Guangdong Longshan Forest Farm and the plus trees selected in Guangdong Province; numbers inside the circles represent the twenty families.
Seedling growth and leaf trait measurements
The stem height and ground diameter (5 cm above the ground) of each seedling were measured from July through August 2014. Nine seedlings in the center of each subplot (each family) were randomly selected for sampling. Over 50 mature leaves were collected from these seedlings, which were used to measure leaf length, leaf width and leaf area using the WinFOLIA® Analysis System (Regent Instruments Inc., Canada). The sampled leaves were dried at 75 °C until they reached a constant mass, and then, the dried leaves were ground and passed through a 2 mm sieve. The specific leaf area was calculated using the individual leaf area and dry mass. The total nitrogen and phosphorus concentrations in the leaves were determined using the Kjeldahl method and spectrophotometry, respectively. The leaf nitrogen and phosphorus concentrations are expressed on a dry mass basis.
The photosynthetic gas exchange of the leaves was measured with an LI-6400® portable photosynthesis system (Li-Cor, Inc., Lincoln, NE, USA). Six seedlings in each family were selected for the measurement of photosynthetic gas exchange. Four to five fully expanded and exposed leaves from each seedling were selected for the measurement of photosynthetic gas exchange. The photosynthetically active radiation was set to 1000 μmol m-2 s-1, and light was provided by the red/blue LED light source of the Li-6400. The measurements were carried out from 8:30 to 12:00 p.m. on sunny days. The gas parameters, such as net photosynthetic rate, stomatal conductance, intercellular CO2 concentration, transpiration rate and instantaneous water use efficiency (WUE), are expressed as average values.
Fresh leaf samples were washed thoroughly with distilled water in the laboratory, dried at room temperature (32 °C) and analyzed for the determination of chlorophyll a, chlorophyll b and total carotenoid concentrations. The chlorophyll a, chlorophyll b and total carotenoids in the leaves were extracted using acetone ethanol and then analyzed using a spectrophotometer (723N Jingke, China). See Tab. 1 for the abbreviations and units associated with these traits.
Tab. 1 - Traits measured in the study.
| Variable | Unit | Description |
|---|---|---|
| H | cm | Stem height |
| D | cm | Diameter at 5 cm above the ground |
| Pn | μmol m-2 s-1 | Net photosynthetic rate |
| gs | mol m-2 s-1 | Stomatal conductance |
| Ci | mmol mol-1 | Intercellular CO2 concentration |
| Tr | mmol m-2 s-1 | Transpiration rate |
| WUE | μmol mmol-1 | Instantaneous water use efficiency |
| Chla | mg g-1 | Chlorophyll a concentration |
| Chlb | mg g-1 | Chlorophyll b concentration |
| Chl | mg g-1 | Total chlorophyll concentration |
| Chla/Chlb | - | Chlorophyll a to chlorophyll b ratio |
| Car | mg g-1 | Carotenoid concentration |
| Chl/Car | - | Chlorophyll to carotenoid ratio |
| LL | cm | Leaf length |
| LW | cm | Leaf width |
| LL/LW | - | Leaf length to width ratio |
| LA | cm2 | Average individual leaf area |
| SLA | cm2 g-1 | Average specific leaf area |
| LN | mg g-1 | Leaf nitrogen concentration on a dry mass basis |
| LP | mg g-1 | Leaf phosphorus concentration on a dry mass basis |
| LN/LP | - | Leaf nitrogen to phosphorus ratio |
Data analysis
One-way analysis of variance (ANOVA) was used to test for differences in seedling growth and in leaf physiological and morphological traits among the different C. wilsoniana families. The broad-sense heritability (H2) of each parameter was estimated based on a one-way ANOVA model as H2= σ2G /(σ2G +σ2R), where σ2G and σ2R are the genetic variance and residual variance, respectively ([37]). The correlations between the leaf traits were analyzed by Pearson’s correlations with a two-tailed test. Principal component analysis (PCA) was used to summarize the variability in the leaf traits. The traits were standardized, and orthogonal factors (i.e., the PC axes) were established as linear combinations of these traits to distinguish certain key traits. The relationships between seedling growth (such as stem height and diameter) and leaf traits were analyzed using ordinary least squares (OLS) linear regression. To select special functional types among the families, cluster analysis was performed on the seedling growth and leaf traits matrix using Euclidean distances and Ward’s linkage method. The number of clusters was determined by the visual assessment of cluster separation in the dendrogram and in the PCA ordination space. All data analysis was performed using SPSS® 17.0 (IBM SPSS Inc., NY, USA), and the significance level was set at P<0.05.
Results
Plant growth
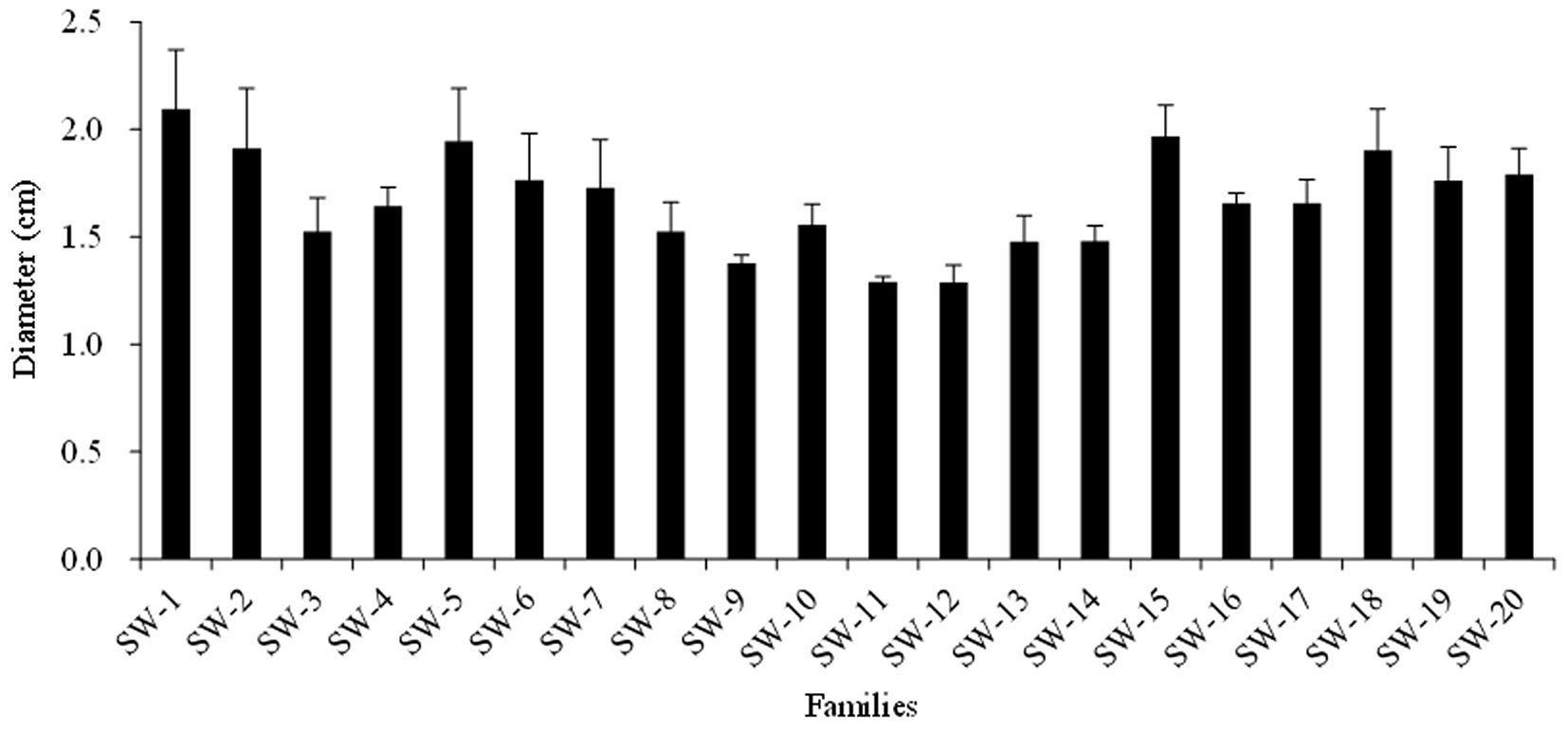
Stem height (H) and diameter (D) were significantly different among the twenty families (P<0.05). The average H was 107.3 cm; the maximum was 141.8 cm, and the minimum was 77.9 cm. The phenotypic coefficient of variance (CV) was 15.1%. The average D was 1.67 cm; the maximum was 2.10 cm, and the minimum was 1.29 cm. The CV was 13.7%. The H2 values were 0.75 and 0.73 for H and D, respectively (Fig. 2 and Fig. 3).
Fig. 2 - Mean ± standard deviation (error bars) of the seedling heights of the twenty families.
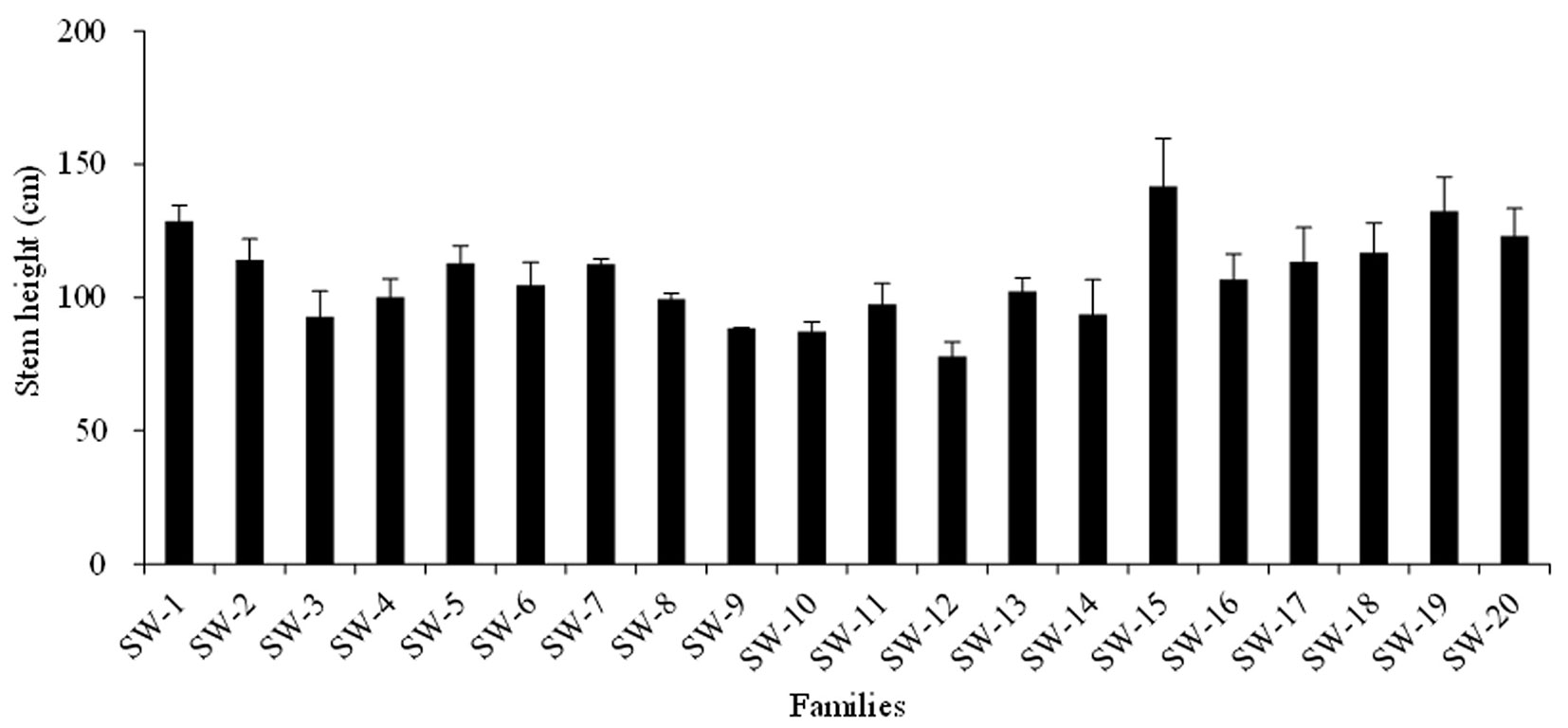
Fig. 3 - Mean ± standard deviation (error bars) of the seedling diameters of the twenty families.
Leaf traits
The majority of the leaf traits showed significant differences among the twenty families (P<0.05), but no significant differences were observed for the ratio chlorophyll a/chlorophyll b (Chla/Chlb, P>0.05 - Tab. 2). The CVs were greater than 20% for gs and Ci; the CVs varied from 10% to 20% for the net photosynthetic rate (Pn), transpiration rate (Tr), WUE, Chla, Chlb, Chl, carotenoid concentration (Car), Chl/Car, leaf area (LA), leaf nitrogen (LN) and leaf phosphorus (LP); and the CVs were less than 10% for the other variables. The H2 was higher for Pn, WUE, Chla, Chlb, Chl, and LN, and it was lower for the other leaf traits, especially for Chla/Chlb (0.17) and Chl/Car (0.31).
Tab. 2 - Descriptive statistics of the leaf traits of twenty families.
| Variable | Min | Average | Max | SD | H2 | CV (%) | P |
|---|---|---|---|---|---|---|---|
| Pn (μmol m-2 s-1) | 5.90 | 8.41 | 10.52 | 1.52 | 0.70 | 18.04 | <0.01 |
| gs (mol m-2 s-1) | 0.044 | 0.073 | 0.102 | 0.017 | 0.56 | 23.17 | <0.01 |
| Ci (mmol mol-1) | 93.36 | 141.97 | 207.92 | 30.74 | 0.54 | 21.65 | <0.01 |
| Tr (mmol m-2 s-1) | 0.91 | 1.29 | 1.62 | 0.18 | 0.47 | 13.62 | <0.01 |
| WUE (μmol mmol-1) | 5.21 | 6.58 | 8.40 | 0.80 | 0.70 | 12.17 | <0.01 |
| Chla (mg g-1) | 1.38 | 1.93 | 2.57 | 0.34 | 0.75 | 17.78 | <0.01 |
| Chlb (mg g-1) | 0.51 | 0.77 | 1.04 | 0.15 | 0.64 | 19.93 | <0.01 |
| Chl (mg g-1) | 1.89 | 2.71 | 3.59 | 0.49 | 0.74 | 17.97 | <0.01 |
| Chla/Chlb | 2.18 | 2.52 | 3.10 | 0.24 | 0.17 | 9.67 | 0.12 |
| Car (mg g-1) | 0.69 | 0.99 | 1.39 | 0.18 | 0.44 | 17.85 | <0.01 |
| Chl/Car | 1.78 | 2.79 | 3.37 | 0.49 | 0.31 | 17.70 | 0.02 |
| LL (cm) | 8.53 | 9.81 | 11.11 | 0.75 | 0.54 | 7.69 | <0.01 |
| LW (cm) | 3.53 | 4.27 | 4.97 | 0.41 | 0.48 | 9.65 | <0.01 |
| LL/LW | 2.11 | 2.34 | 2.58 | 0.12 | 0.49 | 5.33 | <0.01 |
| LA (cm2) | 18.32 | 25.76 | 33.07 | 4.23 | 0.50 | 16.43 | <0.01 |
| SLA (cm2 g-1) | 77.73 | 89.79 | 105.29 | 7.34 | 0.58 | 8.18 | <0.01 |
| LN (mg g-1) | 11.75 | 14.62 | 17.09 | 1.52 | 0.71 | 10.44 | <0.01 |
| LP (mg g-1) | 0.75 | 0.93 | 1.18 | 0.12 | 0.61 | 13.27 | <0.01 |
| LN/LP | 13.44 | 15.82 | 18.03 | 1.26 | 0.48 | 7.99 | 0.01 |
Correlations between leaf traits
Pearson’s correlation coefficients were determined for all pairwise combinations of leaf traits (Tab. S1 in Supplementary material). The gas exchange variables (Pn, gs and WUE) were positively correlated with Chla, Chlb and Chl, but only Pn was positively correlated with SLA and negatively correlated with LA. Chl was positively correlated with LN and LP, and Car was positively correlated with SLA and LP. SLA showed a positive correlation with LP and was negatively correlated with LN/LP. There was a positive correlation between LN and LP.
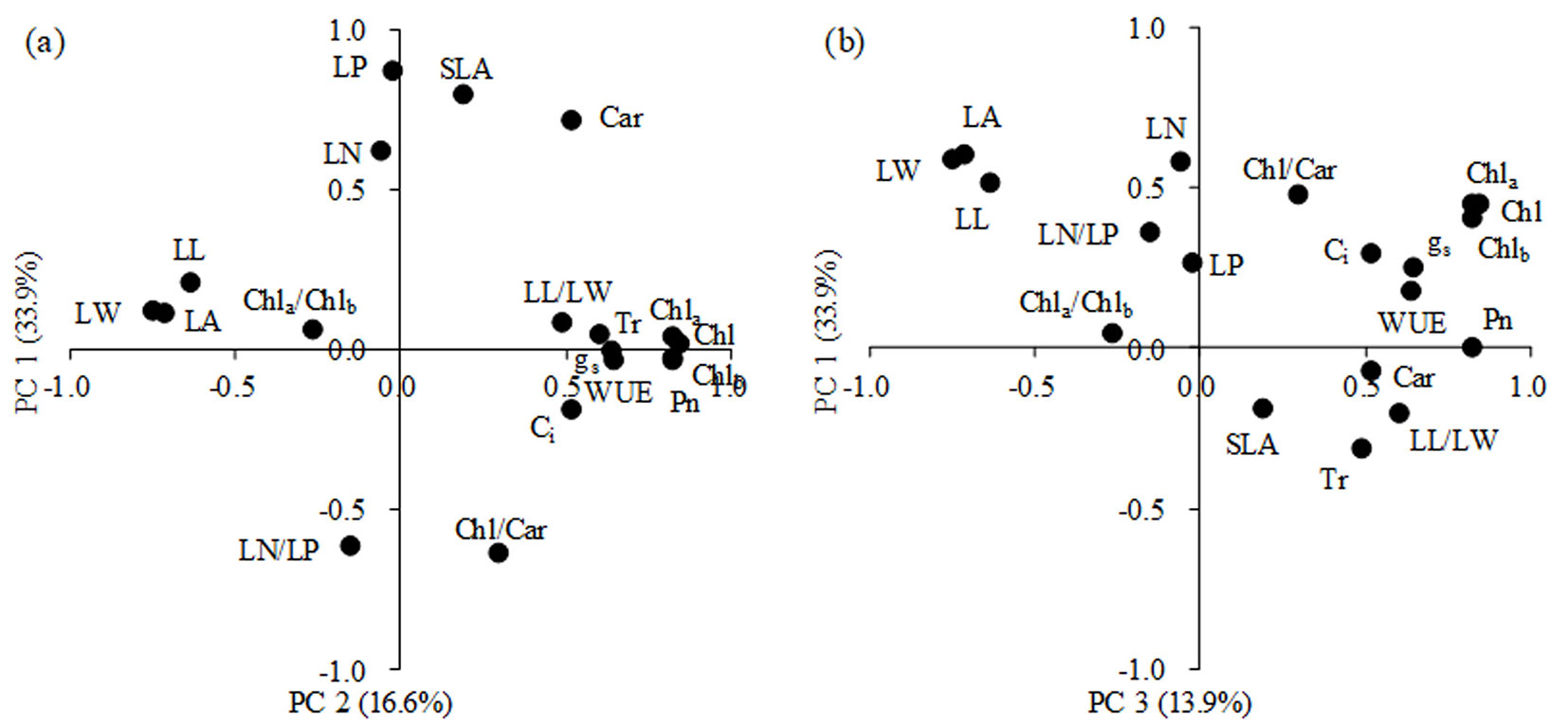
PCA showed that the top 3 principal components explained more than 64% of the total variance (Fig. 4). The first principal component (PC1) accounted for 33.9% of the total variance, which was mainly associated with changes in Chla, Chlb, Chl and Pn. The second principal component (PC2) accounted for 16.6% of the total variance and represented the variation in LP and SLA. The third principal component (PC3) accounted for an additional 13.9% of the total variance, and LA exhibited the highest loading.
Fig. 4 - Results of the principal component analysis (PCA) of the leaf traits, PC1 against PC2 (a) and PC1 against PC3 (b). The percentages in brackets represent the percent of total variance.
The linear least squares regression showed that WUE, Chla, Chlb and Chl were significantly positively correlated with H. The other leaf traits showed no significant linear relationships with H (Tab. 3). D was significantly positively correlated with Pn, gs, WUE, Chla, Chlb, Chl, LL and LL/LW and negatively correlated with LP but showed no significant correlation with the other leaf traits (Tab. 3).
Tab. 3 - Ordinary least squares regression results for the nineteen bivariate relationships.
| X variable | Y variable = H | X variable | Y variable = D | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Slope | Intercept | R 2 | P | Slope | Intercept | R 2 | P | ||
| Pn | 4.581 | 68.772 | 0.183 | 0.060 | Pn | 0.103 | 0.801 | 0.469 | 0.001 |
| gs | 348.170 | 207.903 | 0.135 | 0.111 | gs | 6.278 | 1.213 | 0.222 | 0.036 |
| Ci | 0.087 | 94.967 | 0.027 | 0.488 | Ci | 0.002 | 1.419 | 0.056 | 0.315 |
| Tr | 1.179 | 105.790 | 0.001 | 0.958 | Tr | 0.456 | 1.080 | 0.122 | 0.131 |
| WUE | 11.166 | 33.869 | 0.302 | 0.012 | WUE | 0.169 | 0.558 | 0.349 | 0.006 |
| Chla | 29.635 | 50.024 | 0.393 | 0.003 | Chla | 0.410 | 0.876 | 0.380 | 0.004 |
| Chlb | 51.516 | 67.436 | 0.239 | 0.029 | Chlb | 0.766 | 1.075 | 0.267 | 0.020 |
| Chl | 19.957 | 53.286 | 0.357 | 0.005 | Chl | 0.281 | 0.906 | 0.360 | 0.005 |
| Chla/Chlb | 12.088 | 76.860 | 0.033 | 0.442 | Chla/Chlb | 0.051 | 1.539 | 0.003 | 0.818 |
| Car | 17.106 | 90.425 | 0.034 | 0.433 | Car | 0.314 | 1.358 | 0.059 | 0.302 |
| Chl/Car | 11.203 | 76.068 | 0.115 | 0.143 | Chl/Car | 0.141 | 1.275 | 0.092 | 0.193 |
| LL | -0.613 | 113.322 | 0.001 | 0.905 | LL | -0.092 | 2.575 | 0.094 | 0.190 |
| LW | -8.978 | 145.692 | 0.052 | 0.335 | LW | -0.282 | 2.871 | 0.256 | 0.023 |
| LL/LW | 49.944 | -9.735 | 0.145 | 0.098 | LL/LW | 0.932 | -0.517 | 0.254 | 0.024 |
| LA | -0.495 | 120.066 | 0.017 | 0.588 | LA | -0.022 | 2.242 | 0.170 | 0.071 |
| SLA | -0.420 | 144.981 | 0.036 | 0.423 | SLA | -0.002 | 1.882 | 0.006 | 0.748 |
| LN | -0.357 | 112.512 | 0.001 | 0.887 | LN | -0.051 | 2.410 | 0.117 | 0.140 |
| LP | -33.648 | 138.585 | 0.065 | 0.277 | LP | -0.852 | 2.460 | 0.212 | 0.041 |
| LN/LP | 1.876 | 77.626 | 0.021 | 0.539 | LN/LP | 0.023 | 1.299 | 0.017 | 0.587 |
Cluster analysis
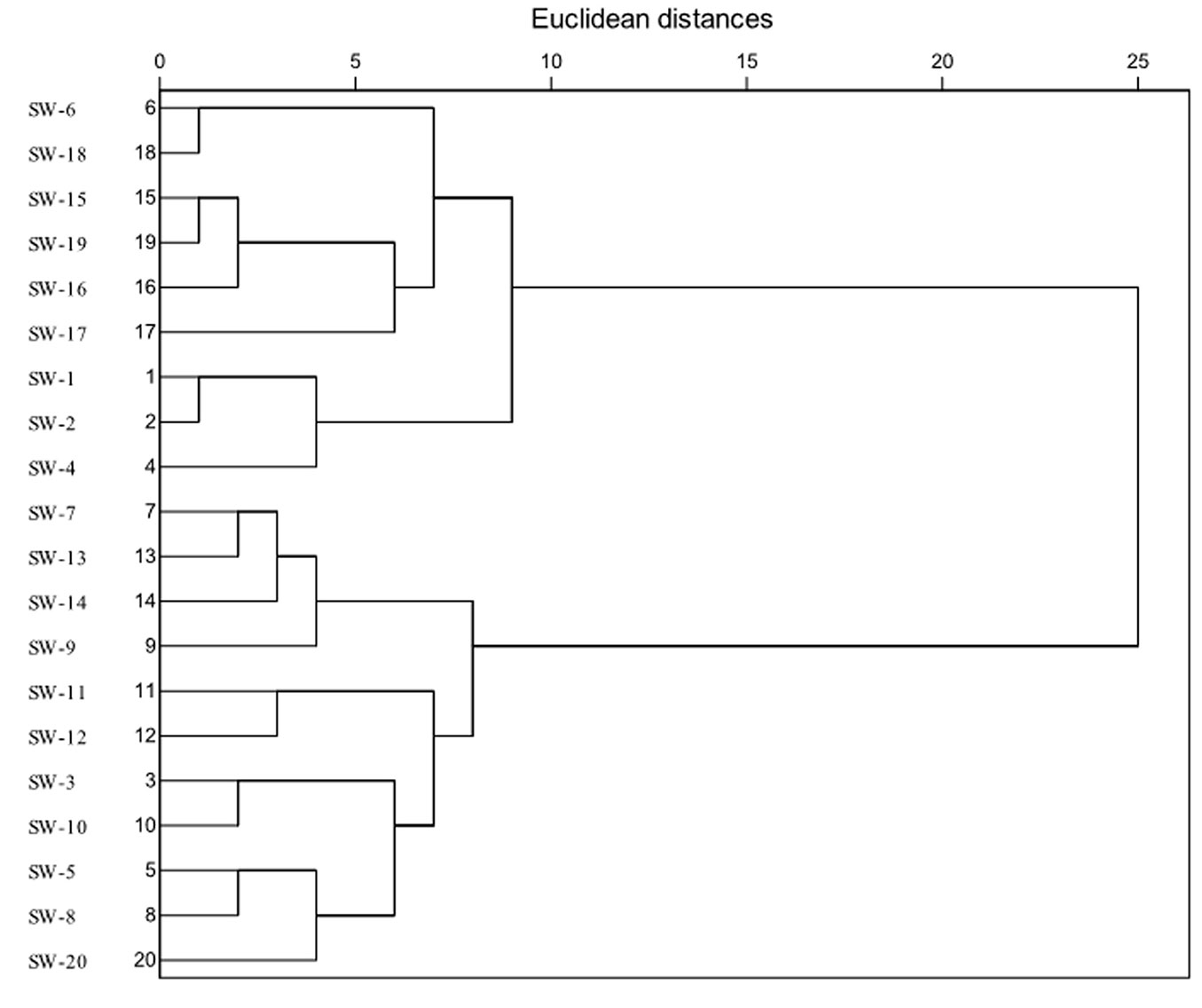
Based on a cluster analysis of leaf traits and seedling growth, the families were classified into four groups (Fig. 5). The first group consisted of SW-6, SW-15, SW-16, SW-17, SW-18 and SW-19; the second group consisted of SW-1, SW-2 and SW-4; the third group consisted of SW-7, SW-9, SW-13 and SW-14; and the fourth group consisted of SW-3, SW-5, SW-8, SW-10, SW-11, SW-12 and SW-20. Collectively, the families in the first and second groups exhibited higher performance in most of the studied traits than did those in the third and fourth groups. The three families in the second group performed better than those in group 1 in terms of Pn, WUE, SLA, and diameter growth rate, and the third group exhibited lower performance in Pn and SLA than the fourth group.
Fig. 5 - Cluster analysis of different families of C. wilsoniana.
Discussion
Variation in growth and leaf traits
It is very important to understand the genetic variation within species, families or clones for plant breeding ([34]). The CV and H2 are two key parameters for evaluating the extent of variability in a breeding population and in selecting cultivars ([17]).High phenotypic variability in a species benefits cultivar selection. In the current study, we found that growth and most leaf traits exhibited significant differences and great variation among the twenty families, with the CVs of different traits varying from 5.33% to 23.17%. This finding is in accordance with previous studies ([19], [18], [14], [13], [41], [43]). Zhao et al. ([48]) showed that the CVs of H and diameter at breast height (DBH) varied from 15.37% to 20.85% for 44 Betula platyphylla families. Pliura et al. ([28]) reported that the CVs of H and DBH varied from 10.05% to 31.60% among poplar clones at different sites. The CVs of H and D in the present study were lower than those in these previous studies, which may be due to differences in the plant species, the numbers of families or clones or the environment.
High heritability was observed for stem height (H2=0.75), diameter (H2=0.73), Pn (H2=0.70), WUE (H2=0.70), Chl (H2=0.64-0.75) and LN (H2=0.71). This result indicates that these traits showed high diversity among the families and low sensitivity to the microenvironmental heterogeneities in the study area. By contrast, Chla/Chlb (H2=0.17), Chl/Car (H2=0.31) and leaf size (H2 varied from 0.48 to 0.54) displayed low heritability, which may be due to low interfamily variability.
Plant physiological and morphological plasticity may play different roles in response to environmental changes. Recently, a review by Gratani ([7]) indicated that in colonized gaps and open areas, the plasticity of physiological traits plays a dominant role because it ensures adjustments in gas exchange in response to changes in environmental factors. Leaf morphology tends to be maintained under stress conditions to avoid the production of structures too expensive to be sustained ([2], [39]). Additionally, morphological plasticity is closely linked to the capacity of a plant to acquire resources in forest understories ([23], [25]). In this study, the variation in leaf physiological traits was greater than that in morphological traits (Tab. 2). The results from the PCA also confirmed that the proportion of the leaf physiological traits accounting for the total variance was greater than that of the morphological traits (Fig. 4). However, some studies have found that the genetic variation among tree genotypes in terms of morphological traits was greater than that for physiological traits ([22], [21]). The difference might be explained by two factors. First, the present study was conducted at a single site, which results in less variation in environmental conditions; the physiological traits with high CVs also exhibited greater H2, which implies that a difference in tree species might be an important factor that leads to different results. Second, the present study included a smaller number of morphological traits.
The twenty families in this study represent a very small subset of the actual population. Additionally, phenotypic plasticity results from phenotypic variation due to genotype and the environment, as well as their interactions; thus, the levels of phenotypic variation may be higher in natural C. wilsoniana stands.
Leaf traits related to growth
The parameters of tree stem growth, such as H and D (or DBH), are often used to evaluate tree growth performance because these two parameters are directly associated with aboveground biomass ([27], [19], [35]). In the present study, H was significantly positively correlated with WUE, Chla, Chlb and Chl and showed a relatively weak correlation with Pn (P = 0.060). D was significantly positively correlated with Pn, gs, WUE, Chla, Chlb and Chl and was negatively correlated with LP. Pn is closely associated with biomass and productivity in many plant species; however, it is not always positively correlated with tree growth and biomass, which might be due to photosynthetic complexity, different carbon allocation patterns, or sampling error while collecting photosynthesis data ([4], [6]). Reddy et al. ([33]) confirmed that Chla, Chlb, the Chla/Chlb ratio and the total Chl showed significant positive correlations with leaf area and yield in different genotypes of mulberry (Morus spp. L.). Therefore, our results indicate that families with higher Pn and chlorophyll content have the potential for better growth performance. A higher WUE would be beneficial for seedling survival and growth in arid habitats, where reducing water loss allows a tree to avoid dehydration and extend water use. In recent years, the seasonal drought frequency has increased in the study area. At the same time, the families with low LP may be better adapted to phosphorus-deficient soils.
Leaf area, total leaf area and leaf number per plant have been considered to be good indicators and have been used to predict the productivity of fast-growing poplar and Salix species ([47], [44]). Some studies have shown that H and D are positively correlated with leaf area and total leaf area ([31], [19]). In the current study, although D was positively correlated with LW and LL/LW, no significant correlation was observed between D and LA. Therefore, compared to the average leaf area, the total leaf area per plant may be more important for predicting plant growth because a larger total leaf area suggests that the plant can capture more light. However, the present study did not investigate the total leaf area per plant.
Many studies have shown that Pn is significantly correlated with LN or SLA and that LN and SLA play key roles in the photosynthetic process. We found that only Pn was positively correlated with SLA; no significant correlations were observed between the two variables and H and D. Some studies have shown a positive correlation between SLA and relative growth rate (RGR - [30], [40]). Other studies have reported that SLA was negatively correlated with biomass production ([24], [46], [38]). These different results for the relationship between SLA or LN and growth or productivity depended on species, growth conditions and tree age ([20]).
In this context, additional studies are needed to test: (i) whether the ranking of these families in terms of growth performance based on leaf traits remains stable over time; (ii) whether these results can be extended to clones; and (iii) the relationship between leaf traits and fruit yield at the level of families or clones.
Conclusion
This study confirmed that seedling growth and leaf traits display high variability among twenty C. wilsoniana families and suggested greater genetic variation in physiological traits than in morphological traits. Pn, WUE, Chla, Chlb, Chla/Chlb, total Chl and LP could be used together to select families with the best growth performance. Plant adaptation to specific environments is a key factor to consider to increase growth and productivity. Based on the variation in seedling growth and leaf traits, 3 superior families were selected for this study area.
Acknowledgments
This work was supported by the Science and Technology Planning Project of Guangdong Province (No. 2014A020208033), the Forest Research Project of Fujian Provincial Department of Forestry (No. 2013-5) and the National Key Technology R&D Program (2015BAD15B02).
Conflict of Interest
The authors declare that they have no conflicts of interest.
References
CrossRef | Gscholar
Gscholar
Gscholar
Supplementary Material
Authors’ Info
Authors’ Affiliation
Hongtao Xie
Mukui Yu
East China Coastal Forest Ecosystem Long-term Research Station, Research Institute of Subtropical Forestry, Chinese Academy of Forestry, Hangzhou, 311400 (P.R. China)
Changzhu Li
Hunan Engineering Research Center of Biodiesel, Hunan Academy of Forestry, Changsha, 410004 (P.R. China)
Guangdong Provincial Key Laboratory of Silviculture, Protection & Utilization, Guangdong Academy of Forestry, Guangzhou, 510520 (P.R. China)
State Key Laboratory of Pharmaceutical Biotechnology, School of Life Sciences, Nanjing University, Nanjing 210093 (P. R. China)
Corresponding author
Paper Info
Citation
Cheng X, Xie H, Zhang L, Wang M, Li C, Yu M, He Z (2017). A comparative study of growth and leaf trait variation in twenty Cornus wilsoniana W. families in southeastern China. iForest 10: 759-765. - doi: 10.3832/ifor2288-010
Academic Editor
Gianfranco Minotta
Paper history
Received: Nov 21, 2016
Accepted: Jun 21, 2017
First online: Sep 02, 2017
Publication Date: Oct 31, 2017
Publication Time: 2.43 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2017
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 49614
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 40855
Abstract Page Views: 3770
PDF Downloads: 3869
Citation/Reference Downloads: 18
XML Downloads: 1102
Web Metrics
Days since publication: 3220
Overall contacts: 49614
Avg. contacts per week: 107.86
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2017): 9
Average cites per year: 1.00
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Seedling emergence capacity and morphological traits are under strong genetic control in the resin tree Pinus oocarpa
vol. 17, pp. 245-251 (online: 16 August 2024)
Research Articles
Genetic variation and heritability estimates of Ulmus minor and Ulmus pumila hybrids for budburst, growth and tolerance to Ophiostoma novo-ulmi
vol. 8, pp. 422-430 (online: 15 December 2014)
Research Articles
Age trends in genetic parameters for growth and quality traits in Abies alba
vol. 9, pp. 954-959 (online: 07 July 2016)
Research Articles
Comparison of genetic parameters between optimal and marginal populations of oriental sweet gum on adaptive traits
vol. 11, pp. 510-516 (online: 18 July 2018)
Research Articles
Patterns of genetic variation in bud flushing of Abies alba populations
vol. 11, pp. 284-290 (online: 13 April 2018)
Research Articles
Genetic variation of Fraxinus excelsior half-sib families in response to ash dieback disease following simulated spring frost and summer drought treatments
vol. 9, pp. 12-22 (online: 08 September 2015)
Research Articles
Preliminary study on genetic variation of growth traits and wood properties and superior clones selection of Populus ussuriensis Kom.
vol. 12, pp. 459-466 (online: 29 September 2019)
Research Articles
Genetic control of intra-annual height growth in 6-year-old Norway spruce progenies in Latvia
vol. 12, pp. 214-219 (online: 25 April 2019)
Research Articles
Comparison of range-wide chloroplast microsatellite and needle trait variation patterns in Pinus mugo Turra (dwarf mountain pine)
vol. 10, pp. 250-258 (online: 11 February 2017)
Research Articles
Genetic diversity of core vs. peripheral Norway spruce native populations at a local scale in Slovenia
vol. 11, pp. 104-110 (online: 31 January 2018)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







