Establishment of a planted field with Mediterranean shrubs in Sardinia and its evaluation for climate mitigation and to combat desertification in semi-arid regions
iForest - Biogeosciences and Forestry, Volume 2, Issue 3, Pages 77-84 (2009)
doi: https://doi.org/10.3832/ifor0498-002
Published: Jun 10, 2009 - Copyright © 2009 SISEF
Research Articles
Abstract
Forested areas are important in arid and semi-arid regions primarily to combat desertification, but also to increase carbon sinks. To reverse the land degradation processes, restoration in the Mediterranean Basin had been frequently obtained by planting indigenous and exotic conifers, but it has been demonstrated that shrubs are nurse species for tree seedlings. Furthermore, planting indigenous shrubs is more efficient than allochthonous in restoring degraded soils. The aims of this work were: 1) to illustrate an experimental area in Sardinia used as a test-site to build up afforestation and reforestation activities in arid and semi-arid areas with autochthonous shrub species; 2) to show the results on plant survival and biomass one year after plantation trying to explain the role of different densities and specific compositions; 3) to hypothesize some trends of C accumulation of this “artificial” Mediterranean semi-arid shrubland by comparison with data found in the literature. The area is located in North West Sardinia, and is characterized by a Mediterranean climate. The revegetation was set up in February 2006, planting local species (Juniperus phoenicea, Pistacia lentiscus and Rosmarinus officinalis). Three densities and three specific compositions (monospecific plots with P. lentiscus, monospecific plots with J. phoenicea and mixed plots with the three cited species) were combined. One month after plantation, almost all plants were alive, but mortality increased after summer, independently of the treatments, likely due to summer drought. During the first year, no differences among the densities and the specific compositions were observed. Total above-ground biomass was in the range of 0.8-3.0 g m-2. Below-ground biomass was in the range of 0.9-1.7 g m-2. A significant lower biomass was measured in October, especially in the Pistacia plots. Higher densities and plant mixing seemed to better perform, allowing establishment of species with a strong pioneer behaviour and aridity resistant characteristics. Data collection on initial conditions and recurrent monitorings will be basic for evaluating in the long term the potential positive effects on C stock and on biodiversity of afforestation activities in arid and semi-arid areas.
Keywords
Introduction
Forested areas are important in arid and semi-arid regions primarily to combat desertification, but also to increase carbon sinks ([36]). The Mediterranean basin is particularly vulnerable to the desertification, as consequence of a mixed of environmental factors and human pressures. In the first category we can cite the occurrence of prolonged drought spells, the presence of highly erodible soils, the high flammability of the species; in the latter, the most relevant phenomena are land abandonment and urbanization of the coastal areas ([70], [39], [3], [16]). According to several studies ([17], see also ⇒ http://dismed.eionet.europa.eu/index_html), Sardinia is an island with high risk of desertification. To reverse the land degradation processes, restoration in the Mediterranean Basin had been frequently obtained by planting indigenous and exotic conifers ([56]), because of their rapid growth and aridity resistance. In Sardinia, in particular, this type of reforestation covered more than 40.000 ha in the 70’s ([5]). On one hand, these interventions provided job for local people, but on the other hand, they substantially altered the original landscape and made erosion processes start ([5]). Maestre & Cortina ([42]) wrote a comprehensive review of the ecological effects of Pinus halepensis plantations in the Mediterranean Basin. According to these authors, supported by several other studies ([44], [4], [7], [43]) classic reforestation by Pinus did not effectively facilitate the establishment of late-successional shrubs under tree canopy. Furthermore, a forest cannot be representative of later stages of succession under semi-arid conditions, because of the scarcity of water resources ([7]).
On the contrary, it has been demonstrated that shrubs are nurse species for tree seedlings, especially in the Mediterranean region ([11], [46], [13], [50]). Therefore, shrubs can protect and improve water and light regime, allowing the germination of seeds and the establishment of late-successional species. Planting shrubs improves physical and chemical soil properties, creating a favourable environment for soil microbes ([24], [53], [26]). Furthermore, the recovery of soil characteristics is the first step leading to reduction of desertification processes ([34], [36]) and increasing soil cover by plantation is a successful way to preserve soil ([53], [66]).
In the present study, we planted three species (J. phoenicea, R. officinalis, and P. lentiscus) selected among the pioneer species colonizing soil after disturbances in this area ([6]). Several studies reported that planting indigenous shrubs is more efficient than allochthonous in restoring degraded soils ([49], [8], [63], [30]). In Mediterranean areas, planting indigenous species adapted to limited nutrient and water resources is essential to obtain high survival rates. But at the same time, plenty of literature ([20], [21], [54], [25], [65], [8], [9], [10], [15], [66], [29], [64]) indicates seedling quality, weeding, irrigation, amendants, mychorrizal inoculation, and the right timing of planting as other important factors.
The aims of this work were: 1) to illustrate an experimental area in Sardinia used as a test-site to build up afforestation and reforestation activities in arid and semi-arid areas with autochthonous shrub species, using the minimum amount of external energy input (minimum tillage, no irrigation, etc.); 2) to show the results one year after plantation trying to explain the role of different densities and specific compositions; 3) to hypothesize some trends of C accumulation of this “artificial” Mediterranean semi-arid shrubland by comparison with data found in the literature.
Materials and methods
Experimental site and plant material
The experimental area is located in the peninsula of Capo Caccia (north-western Sardinia), within the Natural Reserve Porto Conte - Capo Caccia (40° 37’ N, 8° 10’ E). The geologic substrate is Mesozoic limestone. The main natural soil type is Terra Rossa (Lithic Xerorthent and Typic Rhodoxeralfs - [68], [69], [60]), rocky and shallow (20-30 cm), with a sandy loam texture, an ABC profile, and neutral pH. The experimental area (about 5 ha) was established in abandoned arable soil, which has been used in the past for wheat production.
The climate is semi-arid: the average rainfall amount is 640 mm yr-1; precipitation falls in autumn and spring with a long dry period from May to August, with about 60 mm. The mean annual temperature is 16.8 °C. The mean of minimum temperatures in the coldest month is 7 °C, the mean of the maximum temperatures in the hottest month is 28 °C (meteorological station of Fertilia Airport 40°38’ N 8°17’ E; altitude 40 m asl; sea distance 4273 m; period of observation 1961-1990).
In the Natural Reserve, four different vegetation types can be recognized: olm-oak forest, saved from the overexploitation of the XVI century ([6]); an afforested area, set up in the ’70s, with Pinus halepensis Mill., P. pinea L., Cupressus sempervirens L., Eucalyptus spp.; shrublands, which are widespread all over the area, particularly along the firebreak strips, the slopes and under Pinus plantations ([6]); Cistus shrublands, recolonizing abandoned land and burned areas. Cultivated surfaces are now abandoned.
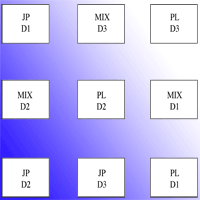
Planting was performed in February 2006. Autochthonous shrubs species were planted, all grown in local nurseries managed by the Regional Forest Service of the Autonomous Region of Sardinia. The species were Juniperus phoenicea L. (JP), Pistacia lentiscus L. (PL), andRosmarinus officinalis L. (RO). Plants derived from seed and were 2-3 years old. Juniperus seedlings were transplanted from 30 cm long phytocells; Rosmarinus and Pistacia seedlings were transplanted from squared pots (10 x 10 x 10 cm). Nine plots (36 x 18 m, 10 m spaced), were located according to a randomized block design with 4 replicates (36 plots). In each block (Fig. 1) three different species compositions were planted at three different densities: 2000 plant ha-1 (D1 - spaced 2 x 2.5 m), 2500 plant ha-1 (D2 - spaced 2 x 2 m), 3333 plant ha-1 (D3 - 2 x 1.5 m). The plot specific compositions were: monospecific with JP; monospecific with PL; plurispecific plots with JP, PL and RO (MIX). Before planting, soil was superficially ploughed at 35 cm depth. All the area was fenced to avoid grazing.
Fig. 1 - An example of a block. The drawing is not scaled.
Plant survival and plant biomass estimation
Plant status (alive or dead) was measured one, five and eight months after plantation, i.e., March, July and October 2006. Plants were recorded as dead when no live aboveground parts were found or when no plant were visible.
In February-March 2006, 16 plants in the monospecific plots and 12-13 per each species in the plurispecific ones were measured for biomass estimation.
Above- and below-ground biomass estimation was based on site species-specific allometric equations, based on the following model (eqn. 1):
These equations were calculated on 32 plants of Juniperus phoenicea and 10 plants of Rosmarinus officinalis, randomly selected among plantation seedlings.
For each seedling, basal diameter (Db), height (H) and the two orthogonal maximum crown diameters (Dc1 e Dc2) were measured. The plants were separated in stem, branches, leaves and roots, then dried at 70 °C until constant weight. The root/shoot ratio (R/S) was then calculated.
J. phoenicea aboveground and belowground biomasses were related to the basal diameter (Tab. 1). R. officinalis aboveground biomass was related to height (Tab. 1), while no relationship was found for the belowground biomass. For this reason, its belowground biomass in the field was estimated applying a fixed R/S (0.68), as found for the plants used in laboratory for the calculation of allometric regression. For Pistacia lentiscus, the allometric relation for the aboveground biomass was taken from a previous study in the same area ([18]). Belowground biomass of P. lentiscus was calculated applying the R/S (0.5) reported by Maestre et al. ([44]) for potted plants.
Tab. 1 - Coefficients (± SE) of the allometric regressions applied for biomass estimation. AG: above-ground biomass; BG: below-ground biomass; Db: basal diameter; H: maximum height; Va: apparent volume (= orthogonal maximum crown diameters x maximum height).
| Parameter | J. Phoenicea | R. officinalis | P. lentiscus | |
|---|---|---|---|---|
| B = a · eb·Db | B = a · Hb | B = a · Va | ||
| AG | BG | AG | AG | |
| a | 3.43 (1.50) |
0.90 (0.36) |
0.006 (0.01) |
577.6 (17.4) |
| b | 0.23 (0.05) |
0.31 (0.05) |
1.981 (0.68) |
- |
| n | 32 | 32 | 10 | 4 |
| r2 | 0.47 | 0.64 | 0.53 | 0.99 |
Soil carbon and nitrogen
To estimate the initial values of soil total organic carbon (TOC) and nitrogen (TN), 144 soil cores (4 samples per plot) at two depths (0-20 cm and 20-40 cm) were collected just before the plantation. The samples were dried at air temperature and sieved at 2 mm. Sub-samples of each core were then dried at 105 °C and analysed by an Elemental Analyser (Flash EA 112 - Thermo Electron, Italy), after the reaction with HCl to remove the inorganic carbon (Method ISO 10694).
Statistical analysis
Statistical differences (p<0.05) of plant survival rates among the different specific compositions and densities were detected by ANOVA, with Bonferroni post-hoc test when the factor was significant. ANOVA was also run to test plant survival rates among species and densities inside the MIX plots. Prior to analysis, survival data were transformed by taking the arcsin of the square root of percentage survival.
Statistical differences (p<0.05) of plant biomass among the different specific compositions and densities were analysed by ANOVA, with Bonferroni post-hoc test when the factor was significant. ANOVA was also run to test biomass differences among species and densities inside the MIX plots.
ANOVA was used to highlight statistical differences (p<0.05) in soil total organic carbon (TOC) and total nitrogen (TN) content at the different depths (0-20 cm and 20-40 cm) and treatments (specific composition and density).
All analyses were performed by SYSTAT 11 (SPSS Inc., Chicago, Illinois).
Results
Living plants
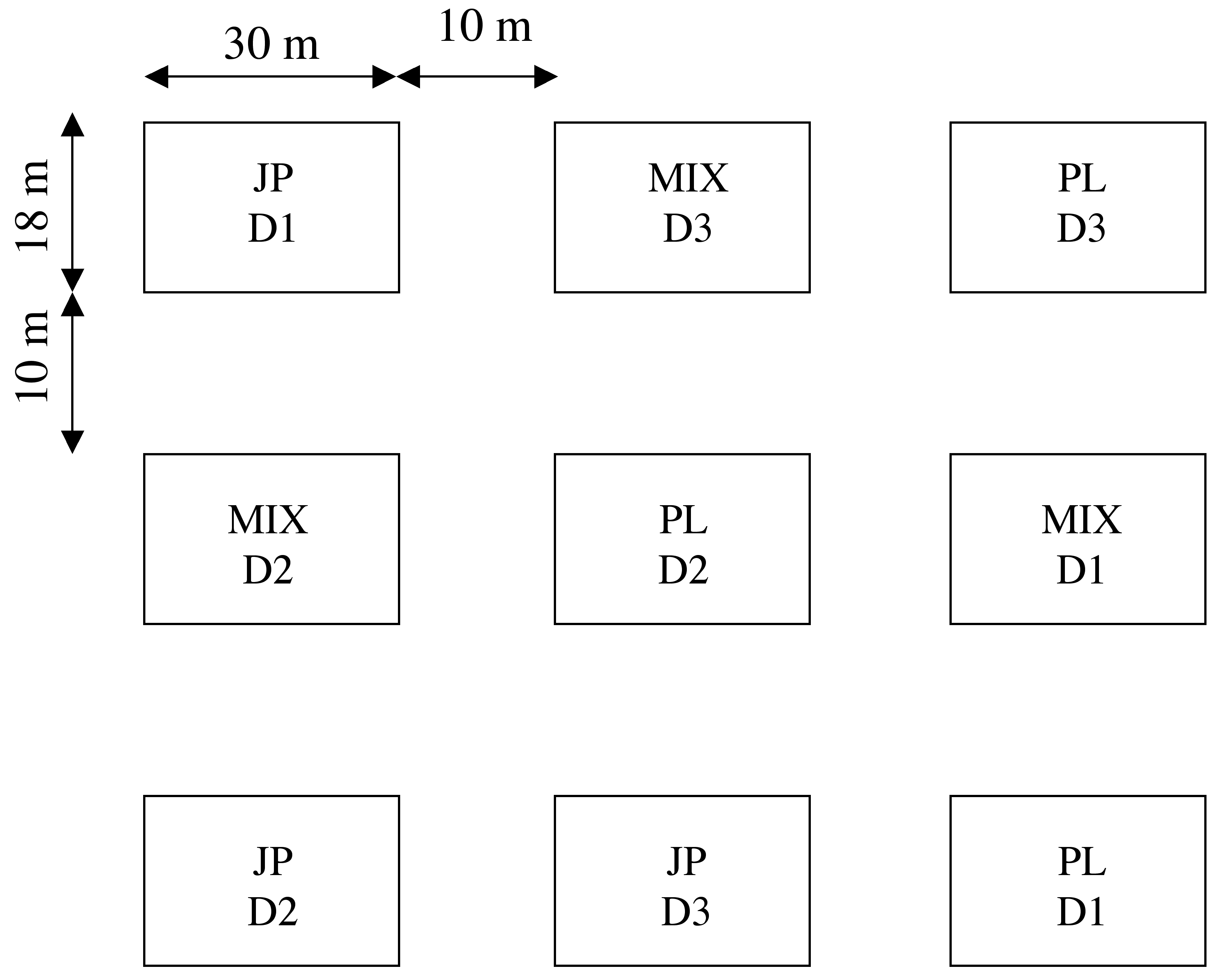
Taking root results are summarized in Fig. 2 and Tab. 2. In March, almost all plants were still alive (99% on average), with no differences among the densities and the specific compositions. In July, the average survival percentage was 93%. No difference was found among densities and specific compositions at this sampling date. In October, the living plants were on average 34%. Alive plants significantly (p<0.001) decreased from July to October, while no difference was observed between March and July. The decrease in living plants between July and October was affected by the specific composition (p<0.001): the highest mortality was observed in PL plots (from 95% of living plants to 11%), while in JP and MIX plots survival decreased from 92% to 45%. This reduction affected the average annual survival: PL was significantly poorer in plants (69%, p<0.001) than JP and MIX (79% and 78%, respectively). On average, D3 plots showed a slight tendency (p=0.1) to have more living plants (79% compared to 74% in the D1 and 73% in D2 plots), but differences among the three densities were not significant. The three specific compositions did not have different survival rates at the different densities. Only at one occasion (October) there were more alive plants in the JP planted at D3 (p<0.01) than in the PL plots at the same density.
Fig. 2 - Living plant percentage in the plots according to densities and specific composition. Error bars indicate SE of the mean (n=4). For details about the statistical differences among the treatments, see Tab. 2.
Tab. 2 - Results of the ANOVA run for alive plants in the JP, PL and MIX plots.
| Components | df | F | P |
|---|---|---|---|
| specific composition | 2 | 4.77 | <0.01 |
| density | 2 | 1.93 | 0.15 |
| time | 2 | 287.08 | <0.001 |
| specific composition * density | 4 | 2.92 | <0.05 |
| specific composition * time | 4 | 9.96 | <0.001 |
| density * time | 4 | 0.57 | 0.69 |
| specific composition * density * time | 8 | 0.42 | 0.91 |
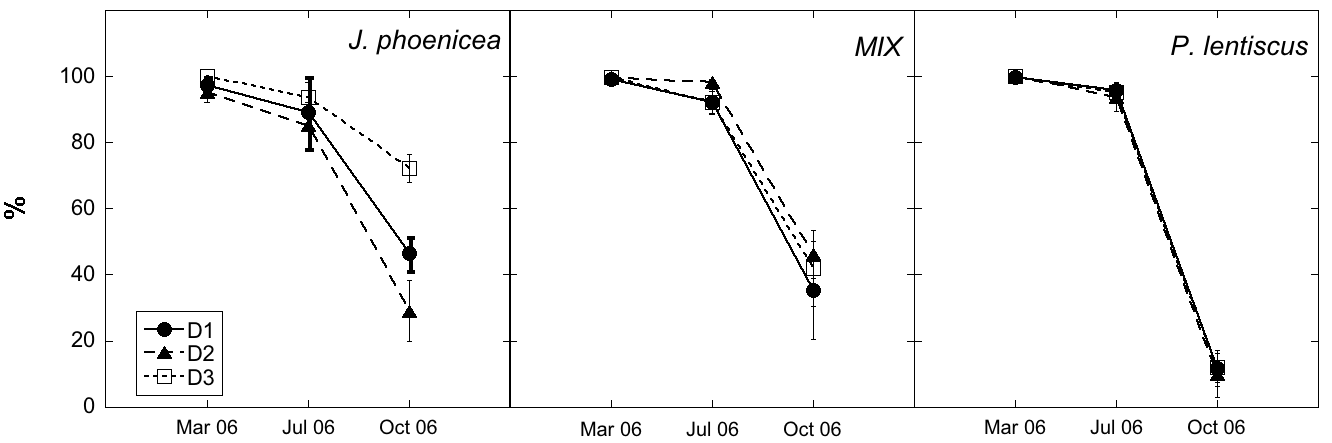
In the MIX plots (Fig. 3, Tab. 3), Pistacia had on average lower (p<0.001) survival rates than Juniperus and Rosmarinus. In March, all plants belonging to all species and densities were recorded as alive. In July, percentage of living plants was on average 94%, without any difference among species and densities. In October, the average survival rate was 41%, but significant lower rates (p<0.001) were recorded for Pistacia plants (21%, compared to 55% and 48% respectively for Juniperus and Rosmarinus), independently on the density. Survival rate changed over time (p<0.001), with lower average values in October, independently on species and density.
Fig. 3 - Living plants (%) in the MIX plots at different densities and sampling time. Error bars indicate SE of the mean (n=4). Black column = J. phoenicea; grey column = P. lentiscus; white column = R. officinalis. For details about the statistical differences among the treatments, see Tab. 3.
Tab. 3 - Results of the ANOVA run for alive plants in the MIX plots for the three different species.
| Components | df | F | P |
|---|---|---|---|
| species | 2 | 13.45 | <0.001 |
| density | 2 | 2.02 | 0.14 |
| time | 2 | 194.01 | <0.001 |
| species * density | 4 | 0.27 | 0.90 |
| species * time | 4 | 2.67 | <0.05 |
| density * time | 4 | 0.47 | 0.76 |
| species * density * time | 8 | 0.65 | 0.73 |
Plant biomass
Total biomass in March 2006 ranged between 0.7 and 5.0 g m-2 with an average value of 2.3 g m-2 (Tab. 4). Aboveground (AG) biomass was on average 1.5 g m-2, ranging between 0.4 and 3.4 g m-2. On average, PL plots had less AG biomass (0.8 g m-2; p<0.01, after post-hoc test) than JP plots (2.3 g m-2), independently on density, but not than the MIX plots (1.7 g m-2). MIX and JP AG biomasses were not different between each other. On average, D3 plots had a tendency (p=0.061, after post-hoc test) to have more AG biomass (2.3 g m-2) than the other two densities (1.4 and 1.1 g m-2, respectively).
Tab. 4 - Estimated above-ground (AB) and below-ground biomass (BG) ± SE in March 2006 (A) and October 2006 (B).
| Date | Species | Plots | AG biomass (g m-2) |
BG biomass (g m-2) |
|---|---|---|---|---|
| (A) March 2006 | JP | D1 | 2.43 (0.67) | 1.17 (0.29) |
| D2 | 1.10 (0.59) | 0.60 (0.22) | ||
| D3 | 3.36 (1.28) | 1.62 (0.56) | ||
| PL | D1 | 0.45 (0.02) | 0.22 (0.01) | |
| D2 | 0.71 (0.09) | 0.35 (0.05) | ||
| D3 | 1.13 (0.32) | 0.56 (0.16) | ||
| MIX | D1 | 1.34 (0.11) | 0.64 (0.05) | |
| D2 | 1.49 (0.21) | 0.79 (0.08) | ||
| D3 | 2.27 (0.51) | 1.33 (0.16) | ||
| (B) October 2006 | JP | D1 | 0.43 (0.24) | 0.17 (0.10) |
| D2 | 0.23 (0.09) | 0.10 (0.03) | ||
| D3 | 1.70 (0.74) | 0.79 (0.36) | ||
| PL | D1 | 0.00 (0.00) | 0.00 (0.00) | |
| D2 | 0.00 (0.00) | 0.00 (0.00) | ||
| D3 | 0.01 (0.01) | 0.00 (0.00) | ||
| MIX | D1 | 0.33 (0.18) | 0.15 (0.08) | |
| D2 | 0.70 (0.24) | 0.34 (0.11) | ||
| D3 | 0.89 (0.39) | 0.47 (0.21) |
Mean belowground (BG) biomass in March was 0.8 g m-2, varying between 0.2 g m-2 and 1.6 g m-2. Mean BG biomass in PL plots was lower (p<0.01; 0.4 g m-2) than in JP (1.1 g m-2) and MIX (0.9 g m-2) plots. On average, in D3 plots there was a higher (p<0.05) amount of BG biomass compared to both the other two densities.
Biomass in October was on average 80% of that of March (p<0.001), having values in the range of 0 to2.5 g m-2. PL plots had significant lower AG biomass (about zero; p<0.05) than in the other two specific compositions (0.8 and 0.6 respectively for JP and MIX), independent of density. D3 plots had on average higher AG biomass (0.9 g m-2; p<0.05) compared to D1 (0.3 g m-2) but not compared to D2 (0.3 g m-2).
BG biomass in October was on average 0.2 g m-2, with a lower amount in PL plots (p<0.05). BG biomass in D3 plots (0.4 g m-2) was significantly higher than what observed in D1 (0.1 g m-2; p<0.05), but not in D2. D1 and D2 BG biomass were not different.
Concerning Juniperus phoenicea, its R/S in March was in the order of 0.9-0.6 in the monospecific and mixed plots, respectively, with no differences among the densities. Similarly, no effect of specific composition and density was observed in October, with values in the range of 0.5-0.6. Time did not affected R/S at all specific composition and densities (Tab. 5).
Tab. 5 - ANOVA results of biomass data, calculated in February and March 2006.
| Biomass | Components | March | October | ||||
|---|---|---|---|---|---|---|---|
| df | F | P | df | F | P | ||
| Above-ground biomass | specific composition | 2 | 5.75 | <0.01 | 2 | 5.39 | <0.05 |
| density | 2 | 3.26 | 0.054 | 2 | 3.57 | <0.05 | |
| specific composition * density | 4 | 0.95 | 0.45 | 4 | 1.97 | 0.13 | |
| Below-ground biomass | specific composition | 2 | 8.07 | <0.01 | 2 | 0.46 | <0.05 |
| density | 2 | 5.36 | <0.05 | 2 | 0.34 | <0.05 | |
| specific composition * density | 4 | 1.21 | 0.33 | 4 | 0.17 | 0.15 | |
| Total biomass | specific composition | 2 | 6.38 | <0.01 | 2 | 5.28 | <0.05 |
| density | 2 | 4.03 | <0.05 | 2 | 3.64 | <0.05 | |
| specific composition * density | 4 | 0.97 | 0.44 | 4 | 1.92 | 0.14 | |
Soil carbon and nitrogen
The soil N content was 0.11-0.17%; the soil C content varied between 1.2% and 2% (Tab. 2). No significant differences were found between the TOC content at the two depths and among densities and specific compositions, as also found for the TN values (Tab. 6). The C/N ratio was on average 11.5, not different at the two depths and among densities and specific compositions (Tab. 6).
Tab. 6 - Soil nutrient (± SE; n=3) just before the plantation at two depths. Different lower-case letters indicate significant differences (p<0.05) between the two depths inside each parameter.
| Layer (cm) |
N% | C% | C/N |
|---|---|---|---|
| 0-20 | 0.15 (0.01) a |
1.71 (0.07) a |
11.68 (0.16) a |
| 20-40 | 0.14 (0.01) a |
1.56 (0.07) a |
11.33 (0.16) a |
Discussion
Seedling survival after eight months ranged between 40-50%. These values are similar to those reported in other studies concerning plantations in Mediterranean region with different species ([15], [63], [47]). Only P. lentiscus showed a very high mortality rates, similar to those found by Garcia-Fayos & Verdu ([23]) and Maestre et al. ([45]).
The high mortality rate is normally associated to summer aridity ([12], [13], [45]). In this study the late planting time (February), added to an early aridity in April 2006, when it rained only 7 mm (Fig. 4, Tab. 7), might have reduced root development before the occurrence of summer drought, affecting seedling survival ([37], [51]).
Fig. 4 - Monthly values of air temperature and precipitation during the years 2006: a) mean temperature (full circle with continuous line), mean maximum temperature (open square with short dashed line) and mean minimum temperature (closed triangles with long-dashed line); b) monthly precipitation. The climate station is located 1 km far from the plantation.
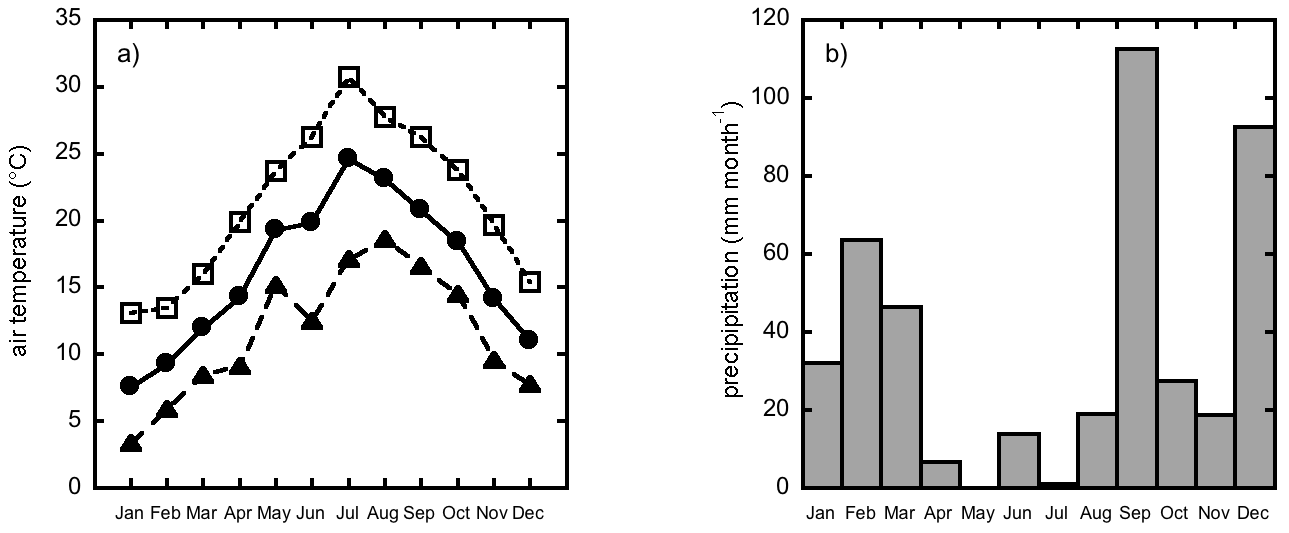
Tab. 7 - Average values of global radiation and air temperaturte and total precipitation during the year 2006. Concerning air temperature, the three values displayed are annual averages of the mean, minimum and maximum temperatures, respectively.
| Variable | Units | Value |
|---|---|---|
| Global Radiation | MJ m-2 d-1 | 15.0 |
| Air temperature | °C | 15.5 11.1 20.7 |
| Total precipitation | mm | 434 |
The different mortality observed among the three planted species could be related to the plant functional types (PFT) they belong. P. lentiscus is a hygrophytic sclerophyll shrub, while R. officinalis and J. phoenicea are both xerophytic with a semi-deciduous and a sclerophyll habit, respectively ([1]). P. lentiscus has an evergreen broad-leaf habit compared to the other two species: this characteristic makes it weaker in front of summer drought, when water table is not achievable, due to a lower control of water balance. Similarly, Gratani & Varone ([28]) asserted that during drought R. officinalis has a higher decrease of photosynthesis and stomatal conductance followed by a higher recovery capacity than P. lentiscus. In the future, physiological measurements will be necessary to make clear this issue.
Plants compete for water and nutrient resources, and this competition may result in a negative effect on growth and on seedlings survival ([38], [59]). Our results showed that higher densities had more biomass and had a tendency towards higher survival rates. This positive effect of density on biomass, concomitant with the unchanged R/S (at least for Juniperus) can be interpreted as a lack of interaction and competition among plants for water and nutrient resources, likely due to the small dimension of plants. The lack of difference in mortality rates of J. phoenicea and P. lentiscus in the mixed plots compared to those of the same species in the monospecific plots support the same conclusion.
Frequently in the past, afforestation practices were done in areas where crops were cultivated, where seed bank of forest species were not available, making difficult the succession and evolution of a stable ecosystem. Our intervention, on the contrary, was set up near a pine forest with an understory of P. lentiscus, Phillirea spp. and other maquis species. As reported by Bonet ([7]) and Pareliussen et al. ([52]), reforestation and restoration are likely to be more successful if efforts are focused on expanding the existing fragments of natural remnant vegetation supplying seed bank, rather than on establishing new fragments in openings.
Because of land abandonment, shrubs in the future will encroach ([27], [33]), whereas, because of climate change, Mediterranean forests will likely face considerable threats ([62]). In a recent work, Harris et al. ([30]) wondered how appropriate is to restore historical ecosystem types when facing rapidly changes of biophysical conditions. According to the same authors, recovery of threatened species and ecosystems will be more difficult to achieve in the future. Therefore, we used plants that can cope with the altered climate conditions ([32], [48], [2]). Experimental evidence showed that once established, those species have the ability to recover rapidly after sever drought effects ([31], [40], [57]), and to sustain elevated Net Ecosystem Exchange (NEE), comparable to forests, as a consequence of the relative low C loss via respiration.
According to the FAO Forest Resource Assessment ([22]), 30 million ha are covered by “other wooded lands”, including shrublands, in the Mediterranean basin. As far as Italy, the last National Inventory of Forest and Carbon INFC dated 2005 (⇒ http://www.sian.it/inventarioforestale/jsp/home.js) reported that shrublands cover about 991.000 ha, that include about 690.100 ha of Mediterranean maquis and shrubs, while the remaining surface is covered by shrubs of sub-alpine and temperate climate. Considering their vast diffusion, efforts should be done to evaluate the potential of these ecosystems in sequestering C in order to achieve the Kyoto Protocol commitments. In this study, we measured plant biomass and soil C, representing the first step of the inventory along the time series. Therefore, only previsions can be merely assumed, given the need of long term monitoring, at least 5-10 years ([61], [67]). Some authors reported a woody biomass increment by 0.25-0.60 t ha-1 yr-1 ([58], [19]) for similar vegetation in Mediterranean climate.
In a Mediterranean environment, La Mantia et al. ([35]) showed a linear increase from 13 to 85 t C ha-1 of soil organic C along a soil chronosequence of 0-30 years after land abandonment. In an area close to our experimental plantation, covered by a 30-year old maquis, soil C is about 87 t C ha-1, which is in agreement with the data given by La Mantia et al. ([35]). Presently in our plantation, assuming a reasonable soil bulk density of about 1.1-1.2 g cm-3 ([9], [14]), soil C is about 0.7-0.8 t ha-1. Based on our soil CO2 emissions measurements (data not shown, but a likely estimation would be 7 t C ha-1 yr-1) and on the low biomass accumulation, the plantation will be a net C source in the first stages. This is in agreement with Paul et al. ([55]), who showed that soil C generally decreased in the first years after plantation, as an effect of the initial low C input from litter, while contemporarily the old C of the preceding agricultural phase continued to decompose, until this effect disappeared in the long term (> 30 years), when soil C became higher than in the preceding agricultural soil.
Conclusions
Forest policy has changed in Italy and Europe during the last decades. The main goals that characterized forest plantations until last decades were to give job to people and to involuntary develop non-regenerating forests, especially in case of lack of management, as occurred for the Italian forests. The likely positive long-term impacts of afforestation activities in arid and semi-arid environments by shrubs species can make wonder if these types of plantations might be a valid alternative to classic revegetation activities when the goal of increasing biological diversity, reducing soil erosion and creating C sink are pursued ([56]). Higher densities and mixed specific composition seemed to better perform, allowing establishment of species with a strong pioneer behaviour and aridity resistant characteristics. Data collection on initial conditions and recurrent monitorings will be effective for evaluating the potential of C sequestration in the long-term by afforestation in semi-arid Mediterranean areas.
Acknowledgments
The research was funded by the Ministry of Environment (Ministero dell’Ambiente e della Tutela dell’Ambiente e del Mare) under the frame of a bilateral collaborative international programme (2006). We also would like to acknowledge the Regional Forest Service in the persons of Dr. Nudda, Dr. Caporali, Dr. D’Angelo, for the support on this research. Finally, we acknowledge Mr. Bianchi and all the workers of the Regional Forest Service for the support in the practical works and for logistics. We are particularly thankful to Dr. Alberto Masci and Dr. Abou Jaoudé Renée for their comments and suggestions.
References
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Online | Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
L Loperfido
P De Angelis
R Valentini
Department of Forest Environment and Resources, University of Tuscia, v. S. Camillo De Lellis snc, I-01100 Viterbo (Italy)
Corresponding author
Paper Info
Citation
De Dato GD, Loperfido L, De Angelis P, Valentini R (2009). Establishment of a planted field with Mediterranean shrubs in Sardinia and its evaluation for climate mitigation and to combat desertification in semi-arid regions. iForest 2: 77-84. - doi: 10.3832/ifor0498-002
Academic Editor
Marco Borghetti
Paper history
Received: Jan 08, 2009
Accepted: Apr 02, 2009
First online: Jun 10, 2009
Publication Date: Jun 10, 2009
Publication Time: 2.30 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2009
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 53491
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 42990
Abstract Page Views: 4400
PDF Downloads: 4957
Citation/Reference Downloads: 51
XML Downloads: 1093
Web Metrics
Days since publication: 6260
Overall contacts: 53491
Avg. contacts per week: 59.81
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Jul 2026)
Total number of cites (since 2009): 13
Average cites per year: 0.72
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Using field and nursery treatments to establish Quercus suber seedlings in Mediterranean degraded shrubland
vol. 13, pp. 114-123 (online: 26 March 2020)
Review Papers
The role of spatial criteria in enhancing forest restoration actions: a systematic review
vol. 19, pp. 157-167 (online: 11 May 2026)
Research Articles
Effect of restoration methods on natural regeneration in the Brazilian Atlantic Forest
vol. 18, pp. 23-29 (online: 15 February 2025)
Technical Reports
Canopy Chamber: a useful tool to monitor the CO2 exchange dynamics of shrubland
vol. 10, pp. 597-604 (online: 15 May 2017)
Research Articles
Methods for biomass stock estimation in Mediterranean maquis systems
vol. 10, pp. 108-114 (online: 21 August 2016)
Research Articles
Rewilding beech-dominated temperate forest ecosystems: effects on carbon stocks and biodiversity indicators
vol. 18, pp. 1-9 (online: 02 February 2025)
Research Articles
Outplanting performance of three provenances of Quillaja saponaria Mol. established in a Mediterranean drought-prone site and grown in different container size
vol. 13, pp. 33-40 (online: 21 January 2020)
Research Articles
The effects of post-pasture woody plant colonization on soil and aboveground litter carbon and nitrogen along a bioclimatic transect
vol. 6, pp. 238-246 (online: 13 June 2013)
Research Articles
Effects of warmer and drier climate conditions on plant composition and biomass production in a Mediterranean shrubland community
vol. 1, pp. 39-48 (online: 28 February 2008)
Research Articles
Can the dynamics of forest restoration reduce landscape fragmentation in the Atlantic forest?
vol. 18, pp. 61-68 (online: 04 April 2025)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







