Post-fire recovery of the plant community in Pinus brutia forests: active vs. indirect restoration techniques after salvage logging
iForest - Biogeosciences and Forestry, Volume 11, Issue 5, Pages 635-642 (2018)
doi: https://doi.org/10.3832/ifor2645-011
Published: Oct 04, 2018 - Copyright © 2018 SISEF
Research Articles
Abstract
Although reforestation is frequently utilized in many Mediterranean Basin countries to restore burned Mediterranean pine woodlands, post-fire recovery of the plant community is often neglected. To compare the post-fire recovery of the plant community following active and indirect post-fire restoration techniques, we studied three post-fire regeneration treatments in a salvage-logged Pinus brutia forest, including two active (plantation and seeding) restoration techniques and one indirect (natural regeneration). An unburned pine stand was also included in the study. We applied the point-intercept method to obtain data on the presence and cover of individual species and functional groups in six replicate one-hectare plots for each treatment. We found no significant differences in plant species richness among post-fire treatments; however, plant community composition and vegetation structure were significantly different between treatments. There was a shift in plant community structure when active restoration techniques were applied, from the woody- and resprouter-dominated plant community of the unburned site to an annual herbaceous- and non-resprouter-dominated one. Our results suggest that active restoration by planting tree saplings in Mediterranean pine forests after a fire may decrease the plant community’s resilience and provide empirical evidence that pine plantation treatments change the plant species composition of these forests. These results have important implications for post-fire management of Mediterranean Basin pine forests.
Keywords
Fire, Mediterranean Pine Forest, Plant Cover, Plant Functional Groups, Post-fire Restoration, Resilience, Species Diversity, Turkish Red Pine
Introduction
Pine forests cover large areas throughout the Mediterranean Basin and are of great ecological and economic importance ([51], [5]). Turkish red pine (Pinus brutia Ten.) and Aleppo pine (P. halepensis Mill.) form pine forests with the most prominent crown fire regimes in low altitudes in the Mediterranean Basin ([27]). In recent decades, especially in the western Mediterranean Basin, socio-economic changes (e.g., the rural-urban migration) and reforestation policies have resulted in homogenous pine stands that are sensitive to drought-driven large wildfires ([45], [46]).
Most of the low-altitude Mediterranean pine forests are resilient to fires and successful post-fire regeneration of these forests has been reported by several authors ([59], [50], [45]). However, the failure of post-fire establishment of pine seedlings in these forests has also been recorded, for instance on steep slopes or in poor soils ([59]), and especially if fires are very frequent ([15]). Field observations ([4], [58]) and modeling studies ([41]) suggest that if fires occur more frequently than 20 years, which is the time period needed for pine trees to develop a canopy seed bank (i.e., immaturity risk), then pine forests transform into open shrublands or grasslands.
In low-altitude Mediterranean pine forests, many species of angiosperms have acquired several adaptations to cope with fire ([40]), including resprouting from underground lignotubers and stimulation of germination by heat shock or smoke ([26]). Owing to these adaptations, post-fire plant diversity increases in the years following a fire until canopy closure is achieved ([27]) while post-fire successional process also contributes to the gamma diversity of the region as each seral stage has its own specific plant and animal species composition ([54], [22]). Many studies have documented a peak in plant diversity during the early stages of post-fire succession in fire-prone Mediterranean Basin habitats ([60], [23], [21]).
Post-fire regeneration of Mediterranean pine forests is a major economic concern in many countries within the Mediterranean Basin ([5], [42]). Therefore, national forest services frequently conduct active restoration by applying different techniques (e.g., pine plantation) for sustainable forestry in these pine forests ([63]). Owing to restoration priorities that favor pine plantations, many studies on post-fire regeneration in Mediterranean pine woodlands have also been performed, with a prominent focus on post-fire pine regeneration ([52], [62]). Few studies, however, have addressed the dynamics of plant community structure in burned Mediterranean Basin pine forests ([23], [21]). Contrary to the prevailing approach of post-fire management, natural regeneration and seeding methods have been suggested instead of pine plantation to restore burned areas in the Mediterranean Basin successfully and cost effectively ([5], [29], [34], [63]). It has also been suggested that less intensive post-fire management techniques would improve plant diversity in Mediterranean Basin forests ([30]), as has been concluded for fire-prone forests in the western United States ([37]). Moreover, owing to global change, post-fire restoration in Mediterranean forests requires a comprehensive approach ([12]). In short, “the restoration of a burned area is not just a matter of how to carry out reforestations” ([34]).
Turkey is one of the leading Mediterranean Basin countries in reforestation (mostly Pinus brutia - [45]); however, P. brutia stands are also utilized intensively for timber production ([33]). Various post-fire management techniques are applied to burned P. brutia forests in Turkey ([5]) without any consideration as to how different restoration techniques affect plant community structure and plant diversity, and with a prominent focus on pine regeneration for future wood production (Ç. Tavsanoglu, personal observation). In Turkey, salvage logging is the first application before any post-fire regeneration treatment in burned P. brutia forests due to economic concerns ([1]).
The present study aims to assess the response of the plant community to various post-fire regeneration treatments in salvage-logged Pinus brutia forests. We hypothesize that plant diversity differs substantially between active (i.e., planting and seeding) and indirect (i.e., natural regeneration) restoration techniques. We therefore expect more diverse and fire-resilient plant communities in areas subjected to indirect restoration.
Materials and methods
Study area
The study was conducted in Çetibeli district, Marmaris region, south-western Turkey (36.98° N, 28.32° E). The study area is dominated by native Turkish red pine (Pinus brutia) forests, with maquis shrubs (such as Quercus infectoria, Phillyrea latifolia, and Myrtus communis) also found in the understory with high coverage ([56]). The Marmaris region is one of the most frequently burned areas in south-western Turkey, with many burned sites co-occurring with unburned forests in the landscape. Consequently, both monopyric (i.e., annuals and post-fire seeders) and polypyric (i.e., post-fire resprouters) species exist in the region as in many typical crown-fire ecosystems ([47]), while the area’s vegetation consists of plant communities adapted to crown fire regimes ([54]). Stunning examples of monopyric species in southwestern Turkey include Chaenorhinum rubrifolium, whose germination is stimulated by multiple fire-related cues ([57]), many woody Lamiaceae species that has smoke-stimulated germination ([8]), several Cistus species in which germination is stimulated by various heat-shocks, and Pinus brutia, a serotinous pine tree ([56]). Many large resprouter shrubs, such as Phillyrea latifolia and Arbutus andrachne, are good examples of the region’s polypyric species ([56]). The climate is typically Mediterranean, with a prominent summer drought period, and wet winters. Serpentine soils cover most of the study area because of ophiolite rocks formed by underwater volcanic activity in the early Mesozoic. The study area is located between 70 and 400 m above sea level.
Fire and regeneration treatments
In the summer of 2002, a high-intensity, stand-replacing crown fire burned 1775 ha of Pinus brutia forest in the study region. After this fire event, fire-killed trees were salvage-logged within the first few months before several regeneration treatments (hereafter “treatment”) were applied to the burned area within the first year after the fire by the local forest service (see below). Although the extent of the burned area was sizeable, treatments were applied at a fine scale (in 1-5 ha sites) throughout the burned area.
The treatments applied to the burned area can be classified as active or indirect restoration techniques ([63]), which are both frequently applied in the regeneration of salvage-logged Pinus brutia forests by forest services ([5]). The first active restoration technique was the pine plantation (hereafter “plantation”) treatment, in which the vegetation remaining after salvage logging was removed by plowing before planting P. brutia saplings. The second active restoration technique was the “seeding” treatment, in which, after salvage logging, the cone-bearing branches of the burned trees and the remaining parts of the shrubs were homogeneously placed on the soil surface before additional Pinus brutia seeds from seed orchards from a neighboring forest locality were spread by hand over the site. The indirect restoration technique was “natural regeneration”, which meant leaving the site as found without applying any treatment after salvage logging.
Study sites and sampling design
We selected the three treatments described above (planting, seeding, and natural regeneration) within the area burned in 2002 and an unburned site next to the burned area. The unburned site had the same characteristics as the burned site in terms of the type of the geological material and soil structure while the pre-fire plant community of the burned site was similar to the unburned site ([55], [56]). Each treatment and unburned site included six 1-ha replicate plots for a total of 24 plots, evenly distributed over three aspects (north, south, and flat; see Tab. S1 and Fig. S1 in Supplementary material). We applied point-intercept methodology to obtain data on the presence and cover of individual species in the selected plots. At a location close to the center of each plot, two line transects, 50 m in length, were established. The distance between two transects in each plot was 20 m. We sampled 100 consecutive points at 50 cm intervals along each transect line, and recorded the plant taxa present at each point to estimate the cover of each species in each transect. This methodology allowed us to estimate mean cover values at the individual taxon level in each plot by averaging the values of the two transects. We also obtained mean cover values for functional groups by summing the cover values of the taxa included in the same functional group. Field sampling was conducted between August 2007 and February 2008. Where possible, plant individuals found at the sampling points were identified to the species level in the field and, if not possible, a specimen was taken for identification at the herbarium. Nomenclature followed Davis ([9]), Güner et al. ([18]), and Stevens ([53]) for the current family names.
Functional groups
We applied regeneration mode and growth form as functional grouping systems to better understand the effects of post-fire treatments on the plant community and vegetation structure at the functional group level. These functional grouping systems are fundamental ones for the fire-prone Mediterranean Basin ecosystems ([56]). Regeneration mode grouping was based on the regeneration traits of plants in crown fire ecosystems ([43]) and included the following groups: (i) obligate resprouters; (ii) facultative resprouters; (iii) obligate seeders; and (iv) species with no specific post-fire regeneration traits (hereafter “none”). Obligate resprouters regenerate only from under- or above-ground buds by resprouting after fire (R+P- in [43]); obligate seeders regenerate only from seeds, have fire-stimulated germination, and lack any resprouting ability (R-P+); and facultative resprouters can regenerate both by resprouting and seeding after fire (R+P+). The none group consisted of non-resprouter species lacking any evidence of post-fire germination stimulation (R-P-). We also considered obligate seeders with a soil seed bank as a separate functional group to distinguish their effects from Pinus brutia, which has a canopy seed bank. The regeneration modes of individual taxa were based on Paula et al. ([40]), Tavsanoglu & Gürkan ([56]), and field observations.
We also applied a binary growth form grouping to distinguish woody and herbaceous species, and these groups were further divided into additional sub-groups (tree, large shrub, shrub, scrub, liana, perennial forb, perennial graminoid, geophyte, annual forb, and annual graminoid) to describe how different growth forms are affected by each treatment. We also analyzed the responses of species belonging to different families to determine if there were any associations between treatments and species taxonomic status.
Data analysis
To obtain the overall cover of the functional group in each plot, we summed the cover of all species in each functional group. Mean cover values of functional groups for each treatment site and the unburned site were obtained by averaging plot values. Therefore, the cover of any functional group could exceed 100%. We also summed the number of species for each functional group. We utilized chi-square analysis to test the statistical significance of the associations between treatments and number of taxa in different functional groups.
Differences in the cover of each taxon and functional group across treatment groups (including the unburned site) were analyzed by one-way analysis of variance (ANOVA). Tukey’s HSD post-hoc test was applied to analyze the differences between treatments. Two-way ANOVAs were also conducted to test the effects of treatments and aspect on the cover of functional groups. For each functional group, we performed two separate two-way ANOVAs: one included the unburned site and all three post-fire treatments (planting, seeding, and natural regeneration); the other excluded the unburned site. The latter analysis was conducted to reduce the possibility of obtaining overestimated p-values indicating the presence of significant differences among groups, especially considering that the majority of variance in the data came from the unburned site. This enabled us to show more clearly the differences in the cover of functional groups across treatments (without the noise from the unburned site). Cover data were log-transformed before each analysis for a better approximation to the normal distribution.
Permutational multivariate analysis of variance was used to test whether the species composition of the plant community differed across treatments, based on 999 permutations of a Bray-Curtis dissimilarity matrix of the presence/absence of species in study plots. Non-metric multidimensional scaling (NMDS) was also performed to visualize the differences in species composition across treatments. We performed both analyses twice to determine the effects of the unburned site on the overall results: one included the unburned site and all three post-fire treatments (planting, seeding, and natural regeneration); the other excluded the unburned site. For these two analyses, we used “adonis” and “metaMDS” functions in the “vegan” package, respectively ([38]).
All analyses were performed using R statistical software (version 3.4.2, R Foundation for Statistical Computing, Vienna, Austria - ⇒ http://www.R-project.org/).
Results
A total of 60 taxa were recorded in the study area (listed in Tab. S2, Supplementary material), belonging to 24 families and 51 genera; however, the study area was dominated by members of the Poaceae, Lamiaceae, and Fabaceae families. Many of the woody taxa were obligate resprouters whereas many of the herbaceous taxa were non-resprouters (Tab. 1). However, there was no relationship between the number of taxa within functional groups (regeneration mode and growth form) and treatments (Tab. 2).
Tab. 1 - Number of species according to their regeneration mode, growth form, and family in each treatment group. (Natural Reg): natural regeneration.
| Group | Subgroup | Unburned | Restoration technique | ||
|---|---|---|---|---|---|
| Plantation | Seeding | Natural Reg | |||
| All Species | Total | 28 | 35 | 40 | 32 |
| Obligate resprouters | 15 | 12 | 11 | 9 | |
| Facultative resprouters | 5 | 6 | 7 | 4 | |
| Obligate seeders | 4 | 8 | 10 | 9 | |
| None | 4 | 9 | 12 | 10 | |
| Resprouters | Total | 20 | 18 | 18 | 13 |
| Woody | 17 | 11 | 14 | 10 | |
| Herbaceous | 3 | 7 | 4 | 3 | |
| Non-resprouters | Total | 8 | 17 | 22 | 19 |
| Woody | 3 | 5 | 5 | 5 | |
| Herbaceous | 5 | 12 | 17 | 14 | |
| Woody species | Total | 20 | 16 | 19 | 15 |
| Trees | 2 | 1 | 2 | 1 | |
| Large shrubs | 9 | 3 | 6 | 4 | |
| Shrubs | 3 | 5 | 5 | 4 | |
| Scrubs | 5 | 6 | 5 | 5 | |
| Liana | 1 | 1 | 1 | 1 | |
| Herbaceous Species | Total | 8 | 19 | 14 | 17 |
| Perennials | 3 | 8 | 6 | 6 | |
| - Perennial forbs | 2 | 2 | 4 | 4 | |
| - Perennial graminoids | 0 | 5 | 2 | 2 | |
| - Geophytes | 1 | 1 | 0 | 0 | |
| Annuals | 5 | 11 | 15 | 11 | |
| - Annual forbs | 2 | 6 | 7 | 6 | |
| - Annual graminoids | 3 | 5 | 8 | 5 | |
| Family | Lamiaceae | 4 | 3 | 3 | 4 |
| Fabaceae | 1 | 5 | 5 | 3 | |
| Asteraceae | 1 | 3 | 3 | 4 | |
| Cistaceae | 1 | 2 | 2 | 2 | |
| Poaceae | 3 | 9 | 10 | 7 | |
Tab. 2 - Association between treatments and number of taxa in different regeneration modes and different growth forms. The raw data is presented in Tab. 1. (df): degrees of freedom; (RM): regeneration mode; (GF): growth form.
| Group | χ2 | df | Prob. |
|---|---|---|---|
| RM (all) | 2.0 | 6 | 0.916 |
| RM (resprouting/non-resprouting) | 6.6 | 3 | 0.086 |
| RM × GF (woody/herbaceous) | 3.1 | 6 | 0.790 |
| GF (all) | 17.7 | 24 | 0.817 |
| GF (woody/herbaceous) | 4.8 | 3 | 0.190 |
There were significant differences between the unburned site and treatment sites in the cover of one-third of the studied taxa (Tab. S3 in Supplementary material). Most differences were related to the regeneration mode of the taxa. That is, cover of seeder taxa, such as Cistus spp. and Cytisopsis pseudocytisus ssp. reeseana, were significantly higher in the seeding and natural regeneration treatments than in the unburned site. Cover of dominant resprouters either decreased (Quercus infectoria, Myrtus communis, and Styrax officinalis) or remained unchanged (Phillyrea latifolia) in burned sites compared to the unburned site. Significant increases in the cover of taxa in the none group were observed in the plantation treatment (e.g., Bupleurum orientale, Avena fatua, and Bromus spp. - see Tab. S3 in Supplementary material).
The results of the two-way ANOVAs showed that the unburned site was clearly different from the burned sites regarding the cover of functional groups (Tab. 3). By excluding the unburned site, less significant results were obtained in almost all functional groups, although the effects of the treatments were still significant for some functional groups. This analysis also showed that different post-fire treatments resulted in different cover values only for resprouters, woody species, and trees (Tab. 3). Aspect made a relatively limited contribution to the total variance and had a significant effect only on the cover of trees and liana (Tab. 3).
Tab. 3 - Results of two-way analysis of variance for the effects of treatment and aspect on the cover of regeneration modes, growth forms, and plant families. Two separate analysis were performed: including and excluding unburned site. (T × A): interaction of treatment and aspect; (ns): non-significant; (*): p < 0.05; (**): p < 0.01; (***): p < 0.001.
| Groups | With unburned site | Excluding unburned site | ||||
|---|---|---|---|---|---|---|
| Treatment | Aspect | T × A | Treatment | Aspect | T × A | |
| Total | **** | * | ns | ns | ns | ns |
| Obligate resprouters | *** | ns | ns | * | ns | ns |
| Facultative resprouters | ns | ns | ns | * | ns | ns |
| Obligate seeders | *** | ns | ns | ns | ns | ns |
| Obligate seeders (soil seed bank) | ** | ns | ns | ns | ns | ns |
| None | * | ns | ns | ns | ns | ns |
| Woody species | **** | ns | ns | *** | ns | ns |
| Trees | **** | ** | ** | ** | ** | * |
| Large shrubs | **** | ns | * | * | ns | ns |
| Shrubs | *** | ns | ns | * | ns | ns |
| Scrubs | * | * | ns | ns | ns | ns |
| Liana | * | ** | * | ns | ** | ns |
| Herbaceous species | * | ns | ns | * | ns | ns |
| Perennials | ** | ns | ns | ns | ns | ns |
| - Perennial forbs | ns | ns | ns | ns | ns | ns |
| - Perennial graminoids | *** | ns | ns | * | ns | ns |
| - Geophytes | ns | ns | ns | ns | ns | ns |
| Annuals | ns | ns | ns | ns | ns | ns |
| - Annual forbs | * | ns | ns | * | ns | ns |
| - Annual graminoids | ns | ns | ns | ns | ns | ns |
| Lamiaceae | ns | * | ns | ns | ns | ns |
| Fabaceae | **** | ns | ns | ns | ns | ns |
| Asteraceae | ns | ns | ns | ns | ns | ns |
| Cistaceae | **** | ns | ns | ** | ns | ns |
| Poaceae | ** | * | ns | * | ns | ns |
More detailed analysis of the cover of functional groups in different treatments indicated that there were significant increases or decreases in different treatment groups compared to the unburned site (Tab. 4). Considering the woody species, the total cover of trees (mainly Pinus brutia) was dramatically reduced in the burned sites compared to the unburned site (from 88% down to 1%) while that of shrubs and scrubs increased (from 13% up to 50% and from 2% up to 19%, respectively). Total cover of herbaceous species significantly increased only in the plantation treatment compared to the unburnt site (from 9% to 56%) whereas no significant changes were found in either the seeding or natural regeneration treatments (Tab. 4). Increase in the cover of annuals was responsible for the significant change in total herbaceous cover in the plantation treatment; however, the lack of any increase in the cover of annuals resulted in an insignificant increase in the seeding and natural regeneration treatments. Specifically, the cover of perennial graminoids was significantly higher in the natural regeneration treatment than in the unburnt site (18% and 0%, respectively). The cover of Cistaceae, Poaceae, and Fabaceae families was significantly higher in the burned sites than in the unburned site whereas no significant change was observed in the cover of Lamiaceae and Asteraceae families.
Tab. 4 - Mean (± standard error) cover values for each functional group in different treatment sites. The results from the one-way analysis of variance for the effects of treatments on the cover of regeneration modes, growth forms, and plant families are shown. The same letter on the cover values indicates no significant difference (p > 0.05) across treatments. (Natural Reg): natural regeneration.
| Groups | Unburned | Restoration technique | F | p | ||
|---|---|---|---|---|---|---|
| Plantation | Seeding | Natural Reg | ||||
| Total | 185.5 ± 14.0 a | 98.7 ± 9.1 b | 100.0 ± 5.8 b | 107.7 ± 6.6 b | 18.0 | <0.0001 |
| Obligate resprouters | 74.9 ± 16.1 a | 12.8 ± 3.1 b | 18.6 ± 7.5 b | 40.1 ± 6.3 ac | 9.3 | 0.0005 |
| Facultative resprouters | 12.0 ± 7.8 | 5.4 ± 2.4 | 13.7 ± 1.2 | 9.6 ± 2.0 | 2.0 | 0.141 |
| Obligate seeders | 93.7 ± 2.4 a | 39.8 ± 3.9 b | 59.8 ± 7.5 c | 52.9 ± 5.6 bc | 14.3 | <0.0001 |
| Obligate seeders (soil seed bank) | 5.5 ± 2.9 a | 31.8 ± 4.5 b | 57.2 ± 7.0 c | 51.5 ± 5.5 bc | 20.2 | <0.0001 |
| None | 4.9 ± 2.4 a | 40.7 ± 11.2 b | 7.9 ± 2.6 ab | 5.1 ± 2.4 ab | 3.7 | 0.029 |
| Woody species | 114.7 ± 6.1 a | 37.2 ± 6.8 b | 75.2 ± 7.2 c | 64.5 ± 4.8 c | 22.4 | <0.0001 |
| Trees | 88.5 ± 2.3 a | 8.0 ± 2.6 b | 2.7 ± 0.9 b | 1.4 ± 0.4 b | 20.7 | <0.0001 |
| Large shrubs | 62.2 ± 10.3 a | 5.7 ± 2.0 b | 11.2 ± 5.5 b | 16.5 ± 4.0 b | 12.8 | <0.0001 |
| Shrubs | 12.8 ± 7.7 a | 23.0 ± 3.2 b | 50.4 ± 3.4 b | 44.8 ± 8.0 b | 11.6 | 0.0001 |
| Scrubs | 2.3 ± 1.0 | 5.7 ± 3.4 | 18.7 ± 7.7 | 15.6 ± 8.1 | 2.3 | 0.109 |
| Liana | 11.1 ± 5.0 | 0.5 ± 0.3 | 3.5 ± 2.8 | 2.7 ± 2.2 | 2.0 | 0.140 |
| Herbaceous species | 8.7 ± 3.3 a | 55.8 ± 12.4 b | 13.7 ± 6.4 a | 26.7 ± 3.8 ab | 8.1 | 0.001 |
| Perennials | 0.7 ± 0.5 a | 6.5 ± 2.6 bc | 4.8 ± 3.0 ac | 18.8 ± 2.7 b | 10.1 | 0.0003 |
| - Perennial forbs | 0.6 ± 0.5 | 0.2 ± 0.1 | 1.2 ± 0.4 | 1.2 ± 0.7 | 1.4 | 0.279 |
| - Perennial graminoids | 0 a | 5.9 ± 2.8 bc | 3.6 ± 2.8 ab | 17.7 ± 3.0 c | 14.0 | <0.0001 |
| - Geophytes | 0.1 ± 0.1 | 0.1 ± 0.1 | 0 | 0 | 0.7 | 0.582 |
| Annuals | 8.0 ± 3.5 | 49.3 ± 12.4 | 8.9 ± 3.7 | 7.8 ± 3.8 | 2.4 | 0.093 |
| - Annual forbs | 3.5 ± 2.3 ab | 12.8 ± 3.5 b | 1.8 ± 1.4 a | 2.4 ± 1.6 ab | 3.4 | 0.038 |
| - Annual graminoids | 4.5 ± 2.3 | 36.5 ± 10.5 | 7.0 ± 2.4 | 5.4 ± 2.5 | 3.0 | 0.057 |
| Lamiaceae | 4.5 ± 2.8 | 1.7 ± 0.8 | 1.1 ± 0.5 | 0.6 ± 0.3 | 0.7 | 0.584 |
| Fabaceae | 0.2 ± 0.2 a | 13.8 ± 4.6 b | 23.9 ± 5.5 b | 21.8 ± 6.2 b | 57.4 | <0.0001 |
| Asteraceae | 0.1 ± 0.1 | 1.5 ± 1.1 | 0.8 ± 0.4 | 1.0 ± 0.5 | 1.6 | 0.219 |
| Cistaceae | 2.3 ± 0.6 a | 18.0 ± 1.0 b | 39.1 ± 3.9 c | 35.9 ± 6.3 bc | 51.4 | <0.0001 |
| Poaceae | 4.5 ± 2.3 a | 41.3 ± 10.2 b | 10.6 ± 4.9 ab | 23.1 ± 2.6 b | 7.8 | 0.001 |
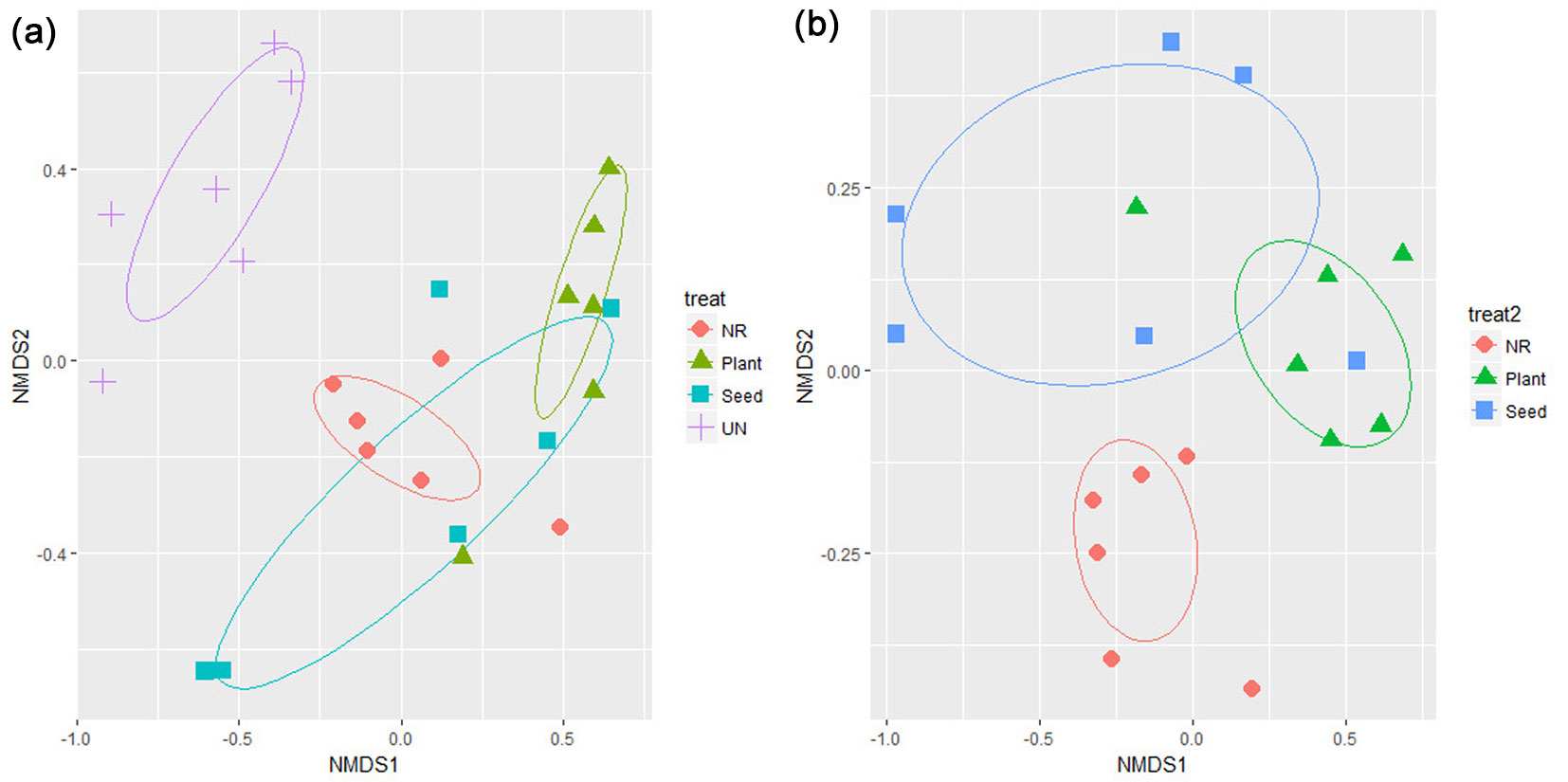
NMDS of species presence data differentiated the unburned site from burned sites (i.e., treatment groups - Fig. 1a), as was confirmed by the permutational multivariate analysis of variance (F = 5.6, R2 = 0.46, p = 0.001). Moreover, after excluding the unburned site, the cross-treatment differences were still significant (F = 3.4, R2 = 0.31, p = 0.003) and NMDS analysis revealed three separate groups (Fig. 1b). However, a clear difference in the two-dimensional space of the NMDS graph appeared between indirect (i.e., natural regeneration) and active (i.e., seeding and planting) restoration treatments after excluding the unburned site from the analysis (Fig. 1b). These results indicate a change in community composition in the burned sites compared to the unburned site. Moreover, they show that the natural regeneration treatment sites had different plant species compositions to the plantation and seeding treatment sites.
Fig. 1 - NMDS ordination graph of species presence across treatments. Two graphs are given: including (a) and excluding (b) the unburned site (UN) from the analysis (see Materials and methods section for details). Each point represents the species composition of the plant community in a single 1-ha plot. Confidence ellipses are also shown for each treatment. (NR): natural regeneration; (Seed): seeding; (Plant): plantation.
Discussion
Our study showed that plant community composition and vegetation structure significantly changed in burned sites compared to the unburned site in Pinus brutia forests. Moreover, this change became more drastic in areas where the plantation treatment was applied. Considering the coverage of plant functional groups, seeding and natural regeneration treatments had less impact on the vegetation structure than the plantation treatment did in P. brutia forests after fire.
Short-term increases in the number of plant species in burned pine forests of the Mediterranean Basin is very common ([23], [21]), which results from fire adaptations observed in many plants ([40]). Management decisions in favor of recruiting pine plantations, however, may lead to the disruption in the natural recovery process in such forests ([31], [16]). On the contrary, it is reported that plowing has minor and temporary effects on plant species richness and vegetation cover; however, it negatively affects pine regeneration in Mediterranean pine woodlands ([19]). As the benefits of planting seedlings offset by its negative effect on the small mammal community in a pine woodland dominated by the non-serotinous species Pinus nigra, it has been suggested that a multi-criteria approach should be implemented before selecting post-fire management practices in such forests ([14]). In our study, although there were no significant differences in plant species richness across the post-fire treatments, plant community composition and vegetation structure for natural regeneration and the other two (seeding and plantation) restoration techniques differed significantly. Consequently, many of the plants found in the seeding and plantation areas were not found in the natural regeneration areas and vice versa. This result indicates that there was a shift in plant community structure when active restoration techniques were applied, specifically from the woody- and resprouter-dominated plant community of the unburned site to an annual herbaceous- and non-resprouter-dominated one. Consequently, we rejected our first hypothesis that plant diversity differs across treatments. However, our results supported the second hypothesis - that indirect restoration sites have a more fire-resilient community than active ones. This result also shows that such a shift in the plant community composition can be masked if only species diversity measures are considered ([35], [2]). The case mentioned above was more dramatic in the plantation treatment in which annual species had approximately 40% more cover than others. The active restoration treatments we included in our study also differed from each other in plant species composition and the cover of the seeder functional group in that both were lower in the plantation than the seeding treatment. This difference suggests that fire-adapted seeder species were eliminated in the plantation treatment and replaced by non-resprouter species without any adaptation to fire. On the other hand, tree (i.e., pine) cover in the plantation treatment was six- and three-times higher than in the natural regeneration and seeding treatments, respectively. This result indicates that plantation is a successful treatment to regenerate the target species Pinus brutia in burned pine woodlands in areas where wood production is the main ecosystem service ([39]). However, the negative effects of pine plantations on several ecosystem services, such as providing suitable animal habitat, preventing soil erosion, and maintaining biodiversity, have also been acknowledged ([31], [39]). Even if the only management goal is regenerating the dominant pine species of the plant community in some areas, biodiversity is still an important part of these ecosystems, which provides services like promoting forest resilience ([61]). In some cases, moreover, even salvage logging is not the best option for post-fire management and it could have long-term negative impacts on ecological and socio-economical services ([7]). Unfortunately, because our sampling design did not include plots of natural regeneration without salvage logging, we cannot evaluate the possible effects of salvage logging on the post-fire recovery of plant community. Therefore, future studies about post-fire management in P. brutia forests should include burned plots in which salvage logging is not applied for a better understanding of the effects of salvage logging on the plant community in such forests.
Resilience is high in resprouter-dominated vegetation in fire-prone Mediterranean Basin areas ([25], [11], [4], [45], [17]) since resprouters can recover quickly after fire. Post-fire resilience in pine-dominated plant communities is also dependent on fire-persistent understory vegetation, including both resprouters ([11]) and seeders ([3]). Consequently, resilience to disturbance is a key functional element of Mediterranean plant communities ([25], [28]), and a deviation from the plant community composition that can be expected from an autosuccessional model (i.e., direct regeneration) is a potential indicator of low resilience in Mediterranean pine forests ([24]). The apparent change in plant community structure in our study, as indicated by significantly lower cover values of resprouters and seeders in plantation sites, therefore suggests that active restoration by planting Mediterranean pine forests after fire (i.e. ploughing and then replanting pine saplings) may decrease the resilience of the plant community compared to less artificial restoration tools (i.e., seeding and natural regeneration in our case). Furthermore, large shrubs had a significant place in the plant community in both the unburned and naturally-regenerated sites in our study, which suggests that these species are sensitive to active restoration techniques. As one of the providers of high forest resilience, this functional group could be used as a substitute for the tree component of the community for ecosystem services such as wood production (i.e., coppicing) and soil protection.
Successful pine regeneration can be achieved through indirect restoration techniques in burned Mediterranean pine forests ([52], [42], [62]). No effect of using log and branch barriers has been demonstrated in post-fire regeneration in Mediterranean pine forests ([49]) whereas the existence of branches on the forest floor is reported to have positive effects ([13], [44], [7]). In this respect, our results support the suggestion that indirect (i.e., natural regeneration) and relatively less artificial (i.e., seeding) restoration techniques should be applied instead of plantation in many cases while restoring burned fire-prone Mediterranean pine forests ([36], [48], [5], [31], [10], [63], [32], [30]). The functional group approach we used in our study provided practical knowledge to improve resilience in Mediterranean pine forest ecosystems and may help to predict the outcomes of future changes in fire regimes to which the plant community is adapted.
Conclusions
Our results provide empirical evidence that plantations are the least beneficial treatment for resilience in Mediterranean pine forests ([31], [20]). We therefore recommend that seeding and natural regeneration techniques should be preferred in salvage-logged Pinus brutia forests after fire. On the other hand, other restoration options that we did not test in our study may also help to improve the diversity and resilience of post-fire plant community, such as the plantation of resprouter species ([17]) and pine planting without removing regenerating vegetation. Our findings on the plant community shift also indicate that plant functional groups provide an effective tool for evaluating the effects of post-fire management approaches in fire-prone ecosystems ([6]). The results of our study have important implications for post-fire management of Mediterranean Basin pine forests.
Acknowledgements
We thank Yasin Ilemin and Kaan Ürker for their assistance in the field, and Baris Özüdogru, Evren Cabi, and Sinasi Yildirimli for their help during plant identification. We also thank three anonymous referees who made comments that significantly improved the manuscript. The plant specimens were identified in the Hacettepe University Herbarium. The data was obtained from field studies conducted as part of the Master’s of Science thesis work of Okan Ürker submitted to Hacettepe University. The present study was financially supported by the State Planning Organization of Turkey (DPT, project no: 2007K120920). The authors declare that they do not have any conflict of interest regarding this paper.
References
Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Authors’ Info
Authors’ Affiliation
Çagatay Tavsanoglu
Fire Ecology and Seed Research Laboratory, Division of Ecology, Department of Biology, Hacettepe University, 06800 Beytepe, Ankara (Turkey)
Institute of Natural Sciences, Yasar University, Izmir (Turkey)
Corresponding author
Paper Info
Citation
Ürker O, Tavsanoglu Ç, Gürkan B (2018). Post-fire recovery of the plant community in Pinus brutia forests: active vs. indirect restoration techniques after salvage logging. iForest 11: 635-642. - doi: 10.3832/ifor2645-011
Academic Editor
Davide Ascoli
Paper history
Received: Oct 08, 2017
Accepted: Jul 04, 2018
First online: Oct 04, 2018
Publication Date: Oct 31, 2018
Publication Time: 3.07 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2018
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 54442
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 42219
Abstract Page Views: 5880
PDF Downloads: 5213
Citation/Reference Downloads: 14
XML Downloads: 1116
Web Metrics
Days since publication: 2829
Overall contacts: 54442
Avg. contacts per week: 134.71
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2018): 9
Average cites per year: 1.13
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Post-fire effects and short-term regeneration dynamics following high-severity crown fires in a Mediterranean forest
vol. 5, pp. 93-100 (online: 30 May 2012)
Technical Reports
Post-fire forest management in southern Europe: a COST action for gathering and disseminating scientific knowledge
vol. 3, pp. 5-7 (online: 22 January 2010)
Research Articles
Post-fire recovery of Abies cephalonica forest communities: the case of Mt Parnitha National Park, Attica, Greece
vol. 11, pp. 757-764 (online: 15 November 2018)
Short Communications
Upscaling the estimation of surface-fire rate of spread in maritime pine (Pinus pinaster Ait.) forest
vol. 7, pp. 123-125 (online: 13 January 2014)
Review Papers
Post-fire soil hydrology, water erosion and restoration strategies in Andosols: a review of evidence from the Canary Islands (Spain)
vol. 9, pp. 583-592 (online: 17 March 2016)
Research Articles
Assessing the performance of fire danger indexes in a Mediterranean area
vol. 11, pp. 563-571 (online: 01 September 2018)
Research Articles
Three prescribed fire regimes on the restoration of flooded savannah grasslands under encroachment of Vochysia divergens Pohl, Pantanal, Brazil
vol. 17, pp. 165-171 (online: 17 June 2024)
Research Articles
Role of serotiny on Pinus pinaster Aiton germination and its relation to mother plant age and fire severity
vol. 12, pp. 491-497 (online: 02 November 2019)
Research Articles
The economic value of fire damages in Tuscan agroforestry areas
vol. 14, pp. 41-47 (online: 14 January 2021)
Research Articles
Historical fire ecology and its effect on vegetation dynamics of the Lagunas de Montebello National Park, Chiapas, México
vol. 14, pp. 548-559 (online: 01 December 2021)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







