
Stem xylem traits in poplar genotypes (Populus L.): additional parameters for selection and breeding
iForest - Biogeosciences and Forestry, Volume 19, Issue 1, Pages 52-60 (2026)
doi: https://doi.org/10.3832/ifor4868-018
Published: Feb 15, 2026 - Copyright © 2026 SISEF
Research Articles
Abstract
Selection programs on Populus species are primarily aimed at developing fast-growing genotypes. This study aimed to examine the anatomical features of stem xylem and vessel traits in Populus clones, providing valuable parameters to guide future selection and breeding efforts. To achieve this objective, cross-sections of one-year-old shoots of two-year-old trees were analyzed using light microscopy. Measurements of xylem parameters were performed using automated image analysis, and theoretical hydraulic conductance (Kh), vulnerability index (Vi), and vessel grouping index were calculated. Based on our findings, we propose that stem anatomical characteristics related to xylem and vessel traits can serve as valuable tools for selecting poplar clones with enhanced productivity, water-transport efficiency, drought resistance, or cavitation resistance. A significant correlation was found between shoot anatomical properties and Kh and Vi. From an anatomical perspective, breeding efforts aimed at increased growth should prioritize developing clones with a higher xylem proportion (xylem/phloem ratio), larger-lumen vessels, a greater proportion of solitary vessels, and a lower percentage of grouped vessels. Conversely, breeding for drought-tolerant clones and those with enhanced resistance to cavitation should prioritize clones with more small-lumen vessels, a higher percentage of grouped vessels, more vessel groups, and vessel groups with more vessels. The anatomical approach described here is an efficient and accessible method for assessing the growth potential, cavitation tolerance, and drought resistance of poplar trees.
Keywords
Introduction
Populus L. species belong to the group of woody species of temperate regions characterized by fast growth and diffuse-porous wood ([29]). Throughout human history, Populus trees have been used for wind protection and erosion control. The broad genetic variability within and among Populus species, combined with the ability for spontaneous and controlled hybridization, has resulted in significant progress in poplar tree improvement ([26]). Initially, Populus selection programs were primarily oriented towards increasing growth rates, adaptability, and pathogen resistance ([3]), but recent breeding initiatives also started to focus on the provisioning of additional ecosystem services, such as carbon sequestration and phytoremediation, due to their fast biomass production and high evapotranspiration levels ([37]). Regardless of the final selection goal, creation of fast-growing genotypes remains the main objective.
Knowledge of the anatomical properties of wood is essential for understanding how wood affects tree growth. Xylem characteristics exert a considerable impact on plant growth. A positive correlation has been established between vessel lumen size and plant growth potential ([4], [38], [19], [40]). This relationship exists because larger-lumen vessels transport water more efficiently, enabling higher transpiration rates, thereby increasing photosynthetic rates. Vessel diameter and number determine hydraulic conductance, which is proportional to the sum of the vessel diameters raised to the fourth power ([31]). On the other hand, it has also been suggested that larger-lumen vessels increase cavitation vulnerability and embolism risk while reducing the mechanical strength of the plant ([20], [11]). Cavitation, as well as subsequent embolism, reduces xylem hydraulic conductivity ([32], [9]). The biological safety of vessels can be estimated using the vulnerability index (Vi), defined as the ratio of vessel diameter to frequency ([31]). Plants with numerous narrow-lumen vessels have lower indices, which is safer for water transport and reduces the risk of air embolism. Baas et al. ([2]) reported that important drought-resistance mechanisms in wood species include numerous small-lumen vessels. However, this can limit hydraulic conductance and consequently compromise growth ([10], [27]). Available evidence indicates that the growth of many species is correlated with vessel lumen size and vessel number ([28], [18]). Vessel grouping provides alternative water pathways when water transport through vessels is blocked by embolism, but it is sometimes disadvantageous because it enables cavitation to spread across vessels within the same group ([39]).
Vessels in Populus wood appear as solitary or in vessel groups ([11], [13]). Compared with other temperate tree species, poplars are generally vulnerable to xylem cavitation ([1], [32], [11]). However, P. tremula exhibits higher cavitation resistance than most other taxa of the same genus. Larger-lumen vessels in poplar trees tend to embolize, whereby hydraulic transport can be almost completely blocked when 40% of vessels are embolized ([12]). Vessel network structure also affects the impact of emboli on xylem hydraulic conductivity. The growth rate of Populus species is positively correlated with xylem conductivity, but not with cavitation resistance. Trade-offs among xylem resistance to cavitation, hydraulic efficiency, and mechanical strength were established by Guet et al. ([9]) at the within-population level in Populus nigra. However, these authors did not confirm the trade-off between xylem resistance to cavitation and growth. Hajek et al. ([11]) recorded moderate-to-low genetic variation in shoot xylem anatomical traits among the five Populus genotypes, but high heterogeneity in root-related anatomical and derived hydraulic traits. Despite demonstrating a close link between vessel size and hydraulic conductivity, the authors suggested that hydraulic conductivity must be at least partly dependent on other conduit properties, such as pit membrane structure. In their investigation on Populus trichocarpa, Jacobsen et al. ([13]) found that vessel diameter was positively correlated with organ diameter, sieve tube diameter, pit density, and pit membrane area. Conversely, Schreiber et al. ([32]) found that greater tree height in aspen and hybrid poplars was associated with narrower vessel diameters, which they explained as an adaptation to cold environmental conditions in the boreal areas of Canada, aimed at preventing winter-freezing-induced embolism. Water-stressed poplar plants have smaller-diameter, denser, and thicker-walled stem vessels that exhibit lower vulnerability to cavitation ([1]).
The aim of this study was to provide anatomical insights into the stem xylem and vessel characteristics of Populus clones in order to generate additional parameters essential for future selection and breeding programs.
Material and methods
Plant material was provided by the Institute of Lowland Forestry and Environment (ILFE), University of Novi Sad, Novi Sad, Serbia, which included nine poplar clones: 665 (P. deltoides Bartr.), 129-81 (Populus × euramericana (Dode) Guinier.), 181-81 (Populus × euramericana), Antonije (Populus × euramericana), Bora (P. deltoides), B-81 (P. deltoides), Pannonia (Populus × euramericana), PE19-66 (P. deltoides) and PE4-68 (P. deltoides). The selected clones are superior candidates identified during ILFE’s long-term complex testing for multiple traits and various uses ([22], [14], [23]). The study was conducted on a two-year-old plantation of poplar clones, grown at the experimental estate “Kaćka Šuma” managed by ILFE (45° 17′ 33.75″ N, 19° 53′ 22.87″ E). The experimental estate is located on the Danube alluvial plain, with predominantly poplar experimental plots grown on alluvial soils (fluvisol type) characterized by sandy conditions and a relatively high groundwater table. An experimental plot was established to simulate traditional timber production in poplar plantations with 1-year-old rooted seedlings, and samples were collected from 1-year-old sylleptic branches of 2-year-old trees. The experimental site is characterized by a moderate continental climate with cold winters and hot semi-arid summers. The average annual temperature is 11.7 °C, and precipitation is unevenly distributed, averaging approximately 630 mm ([15]).
Segments of one-year-old shoots (cut at 15 cm distance from the top) of ten plants of each clone were obtained and fixed in ethanol-water-glycerin fixative (at a 5:4:1 ratio). Using a hand microtome and a scalpel blade, ten cross-sections were obtained per clone. The sections were stained for lignin using the floroglucinol test ([30]). Measurements of xylem and vessels were performed in three fields of view, 120° apart, on each cross-section, and the average values were used in subsequent analyses. All vessels in the field of vision were measured.
Anatomical observations and measurements (cross-sectional areas of specific tissues, the number of vessel groups, the number of vessels per group, percentage grouped vessels in xylem) were performed using light microscopy and Image Analyzing System Motic Images Plus (Motic, Germany) using objectives 4×/0.1, 10×/0.25, 20×/0.45, and 40×/0.65. On the basis of the performed measurements, proportions of stem tissues (peridermis, primary cortex, secondary phloem, secondary xylem, and pith) were calculated.
An image-processing pipeline implemented in Python and Nextflow ([6]) was developed to quantify vessel structures in all 264 sample images across 9 clones. Vessel segmentation was primarily based on manual annotations drawn in the Napari image viewer, with polygon coordinates saved as comma-separated values (CSV) files. These annotations were used to generate integer label images of vessel structures for each sample image. Additionally, a StarDist ([36]) model was trained to recognize vessel structures using approximately 30% of the sample images, with pre- and post-processing steps applied to improve prediction accuracy. The pipeline outputs vessel metrics, including vessel lumen area (VL), vessel diameter (VD), and vessel frequency (VF, the number of vessels mm-2 of xylem) within a sample image. Vessel percentages (%V) were calculated as the sum of the vessel cross-sectional area relative to the total xylem cross-sectional area. The original image and the StarDist model prediction for vessels are shown in Fig. 1.
Fig. 1 - Light micrograph of the stem cross-section of a one-year-old shoot of the poplar clone Bora in the zone of secondary xylem (top panel) and the corresponding label image of vessels (bottom panel). Scale bar: 50 µm.

Theoretical hydraulic conductance (Kh) was calculated using the following formula based on Hagen-Poisseuille’s law ([31] - eqn. 1):
where d represents vessel diameter in meters, which was calculated from the vessel lumen area (VL = π d2/4), ρ denotes fluid density (assumed to be 103 kg m-3 for water at 20 °C), and η indicates viscosity (assumed to be 1.002 · 10-9 MPa s for water at 20 °C).
The vulnerability index (Vi) was calculated as the ratio of vessel diameter (d) to vessel frequency (VF) ([31]). Vessel grouping index (Vg) is a numerical measure of the percentage of grouped vessels in the xylem ([31]). It is defined as the number of all vessels per mm2 divided by the sum of solitary vessels and the number of vessel groups per mm2.
Statistical data analysis was conducted using STATISTICA® for Windows v. 14.0 ([34]). Calculated parameters included mean values, standard errors, standard deviation, and coefficients of variation. Differences in mean values were assessed using the Duncan test (p ≤ 0.05), and Principal Component Analysis (PCA) was performed to examine character variability. Specimen grouping was assessed using Discriminant Function Analysis based on parameters that significantly contributed to the observed differences among the analyzed clones. Correlation analysis was performed to assess relationships among variables. Auto-correlated variables were excluded from further statistical analyses.
Results
The analysis of wood anatomical characteristics revealed differences in proportions of individual tissues among clones (Tab. 1). The results indicate that the clone Pannonia stood out from other examined clones due to the highest percentage of peripheral tissues (periderm, primary cortex, and secondary phloem), as well as the lowest percentage of xylem in the stem (Tab. 1). Peripheral tissues were less developed in clones B-81 and Bora, which had the greatest proportion of xylem, and therefore the highest xylem/phloem ratio. Clone PE4-68 had low proportions of both phloem and xylem, but a high xylem/phloem ratio, and the largest part of the stem was occupied by the pith. A high percentage of xylem was also found in clones PE19-66, 129-81, and 665.
Tab. 1 - Percentages of stem tissues in poplar clones (mean value, coefficient of variation %). Different letters indicate significant differences among the clones for the analyzed parameter at p < 0.05, according to the Duncan test.
| Clone | % Periderm | % Primary cortex |
% Secondary phloem |
% Secondary xylem |
Xylem/phloem ratio |
% Pith |
|---|---|---|---|---|---|---|
| 665 | 3.6 (26.2) bc | 10.1 (19.7) b | 24.0 (7.5) cd | 53.7 (8.1) ab | 2.3 (12.4) bc | 8.5 (42.0) bc |
| 129-81 | 4.0 (8.9) bc | 10.6 (12.7) b | 25.9 (8.0) bc | 54.8 (6.6) ab | 2.1 (13.7) bc | 4.7 (25.0) d |
| 181-81 | 3.8 (23.1) bc | 9.6 (18.6) bc | 29.5 (33.6) ab | 52.0 (22.2) b | 1.9 (32.0) c | 5.1 (29.7) d |
| Antonije | 4.4 (22.2) b | 10.3 (19.2) b | 25.9 (14.4) bc | 51.6 (14.3) b | 2.1 (26.4) bc | 7.8 (29.2) bcd |
| Bora | 3.4 (25.7) c | 9.4 (24.3) bc | 23.7 (13.7) cd | 58.8 (6.9) a | 2.5 (19.5) b | 4.9 (28.8) d |
| B-81 | 3.1 (15.4) c | 7.8 (24.4) c | 19.9 (19.3) d | 59.7 (7.6) a | 3.1 (22.5) a | 9.7 (25.6) b |
| Pannonia | 7.7 (29.4) a | 16.0 (14.2) a | 31.4 (10.1) a | 39.3 (14.5) c | 1.3 (21.8) d | 5.7 (24.4) cd |
| PE19-66 | 3.8 (13.7) bc | 11.3 (18.3) b | 23.4 (11.3) cd | 56.0 (10.2) ab | 2.4 (19.8) b | 5.7 (71.9) cd |
| PE4-68 | 4.4 (16.0) b | 10.6 (17.0) b | 20.5 (11.6) d | 49.7 (13.4) b | 2.5 (16. 7) b | 14.9 (48.2) a |
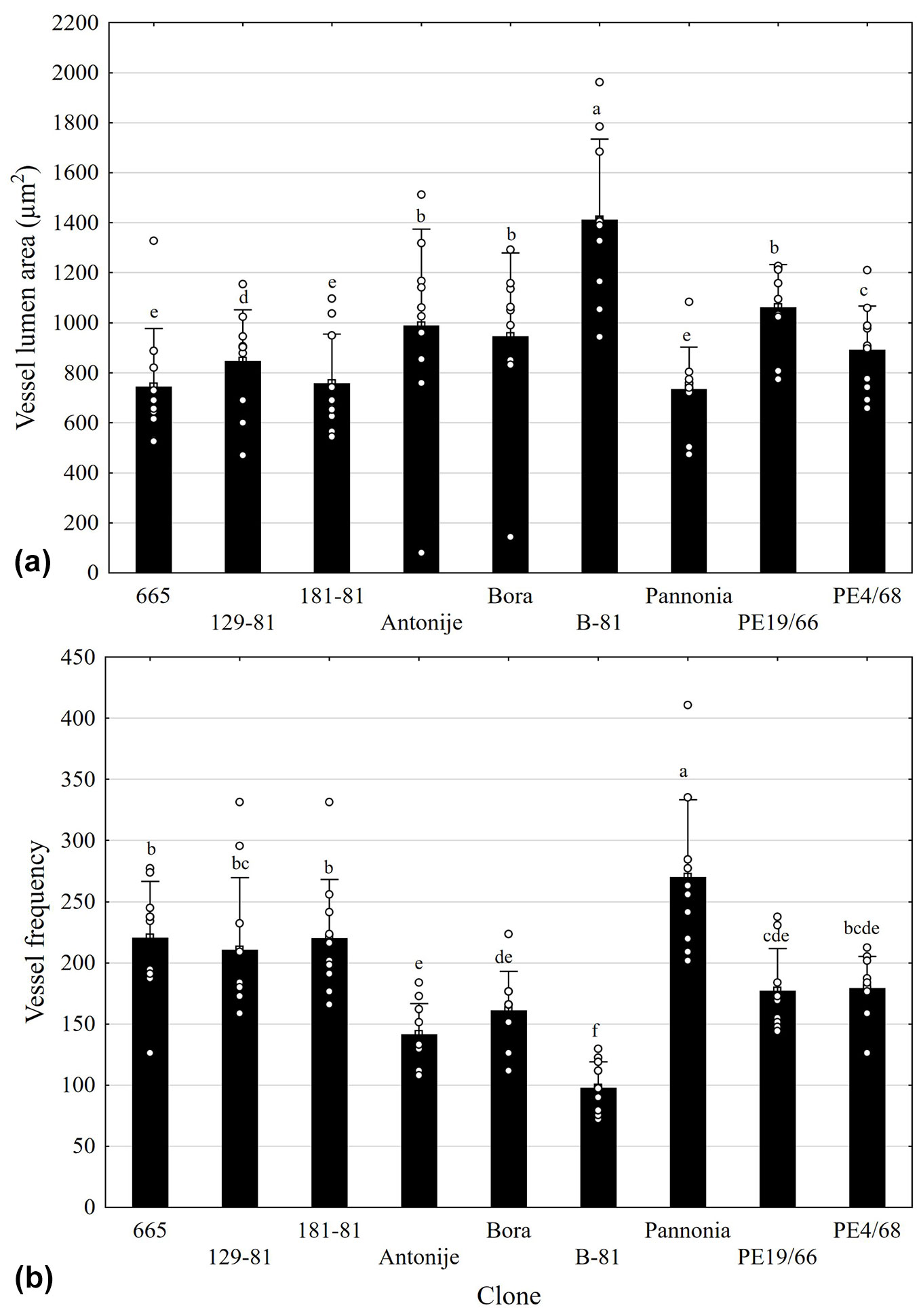
The average VL in the examined clones ranged from 735.6 µm2 in clone Pannonia to 1412 µm2 in clone B-81 (Fig. 2, Fig. 3). Similarly, the VD values, which are naturally highly correlated with VL, ranged from 29.7 µm in clones Pannonia and 665, to 41.4 µm in clone B-81. These two clones also exhibited the highest VF values, with B-81 showing 97.7 and Pannonia 269.9 vessels per mm² of xylem. A significant negative correlation was observed between VL and VF (r = -0.42), as well as between VD and VF (r = -0.53). Clones B-81, Bora, Antonije, and PE19-66 were characterized by vessels with the largest lumen areas, but had low VF. In contrast, clone Pannonia, followed by clones 665 and 181-81, had the highest number of vessels with small lumens.
Fig. 2 - Light micrographs of stem cross-section of one-year-old shoots of poplar clones in the zone of secondary xylem: (a) 665; (b) 129-81; (c) 181-81; (d) Antonije; (e) Bora; (f) B-81; (g) Pannonia; (h) PE19-66; (i) PE4-68 (magnification 200×).
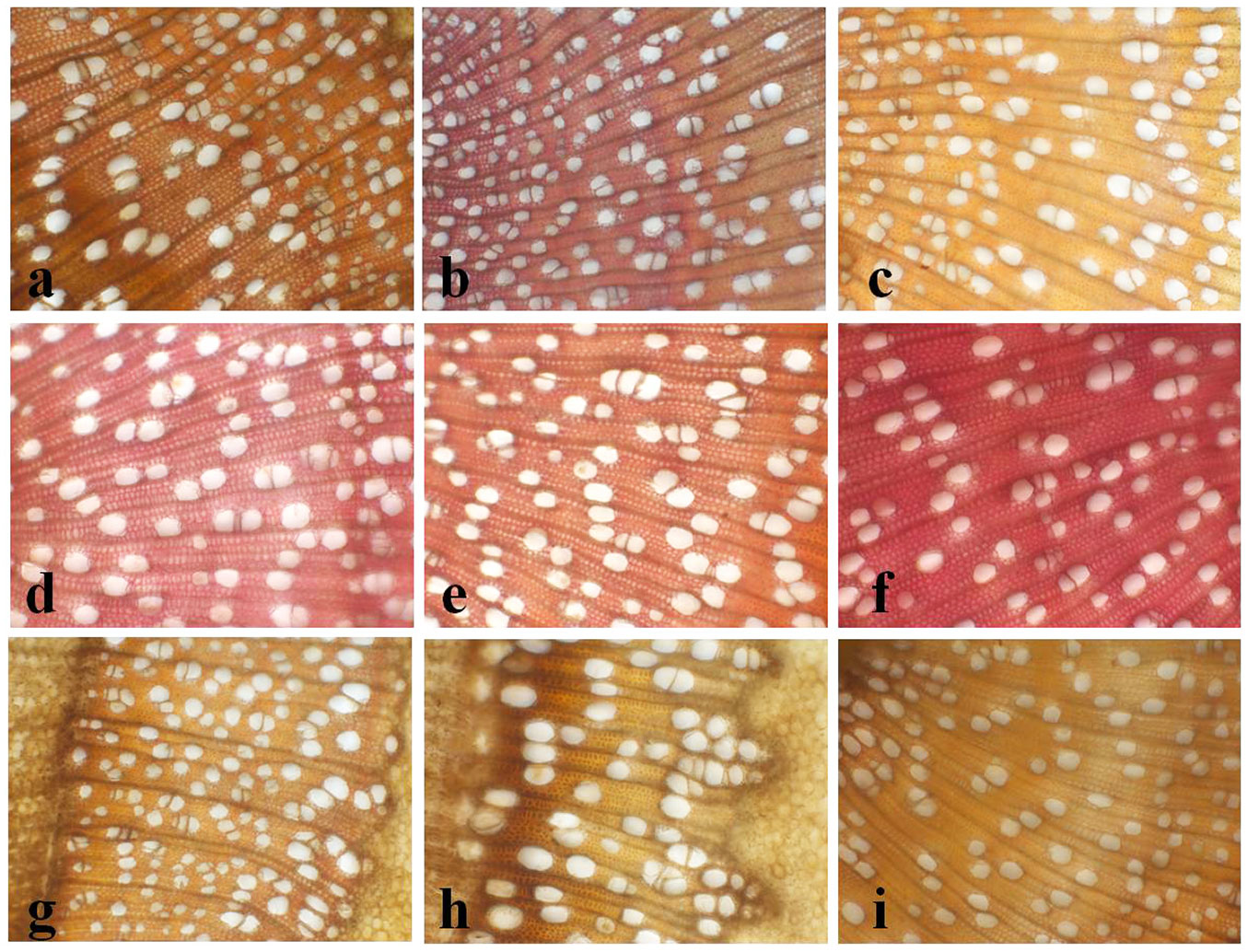
Fig. 3 - (a) Vessel lumen area (VL) and (b) vessel frequency (VF) in xylem of poplar clones (mean and standard deviation). Different letters indicate significant differences between the clones for the analyzed parameter (p<0.05), according to the Duncan test.
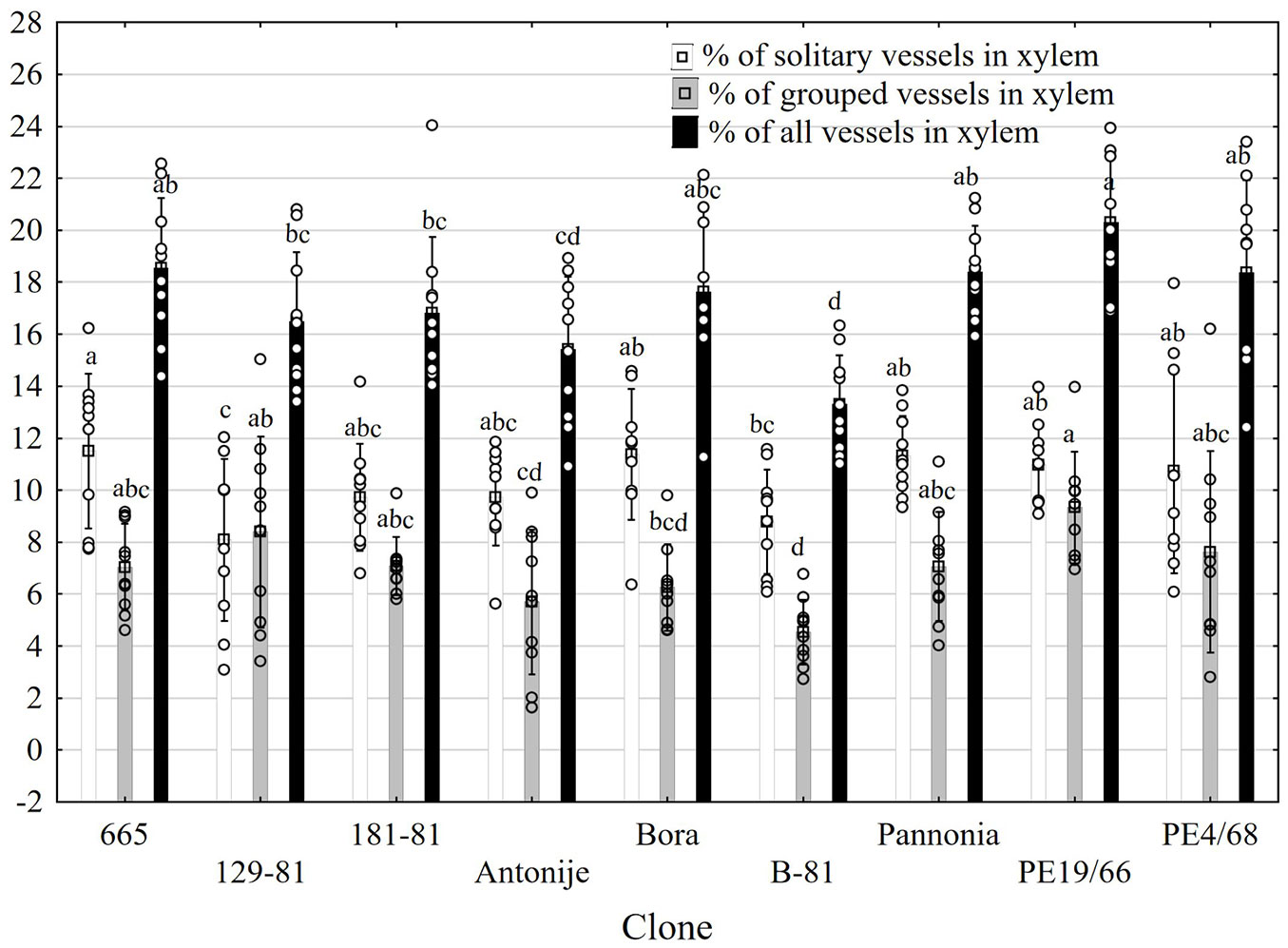
Vessels in the examined clones were present either as solitary or in groups. The highest total proportion of vessels in xylem (20.3%) was observed in clone PE19-66, followed by clones PE4-68, 665, Pannonia, and Bora (Fig. 4). In contrast, the lowest proportion of vessels was recorded in clone B-81 (13.3%). With the exception of clone 129-81, solitary vessels were significantly more numerous relative to grouped ones in all clones.
Fig. 4 - Percentages of solitary and grouped vessels in the xylem of poplar clones (mean ± standard deviation). Different letters indicate significant differences between the clones for the analyzed parameter (p<0.05), according to the Duncan test.
Grouped vessels were arranged in radial multiples. The number of vessel groups per mm2 of xylem showed a strong positive correlation with the number of vessels per group (r = 0.96). Clone 181-81 exhibited the highest number of vessel groups per mm² of xylem (57.6 - Tab. 2). A high number of vessel groups was also observed in clones 129-81 and Pannonia. Across these three clones, the vessel groups included more than seven vessels, on average. Significantly, the smallest proportion of grouped vessels was recorded in clone B-81, which, along with clone Antonije, had the lowest number of vessel groups per mm² of xylem (16.9 and 25.0, respectively). In these two clones, the vessel groups comprised a small number of vessels (two to three on average). The vessel grouping index contextualizes the total number of vessels relative to the number of vessel groups. It showed a significant negative correlation with VL (r = -0.24), and a positive correlation with VF (r = 0.51). The highest Vg values were recorded in clone 129-81, while the lowest were observed in clone Antonije (Tab. 2). Most clones exhibited similar Vg values.
Tab. 2 - Calculated values of Kh, Vi, and Vg and parameters of vessel groups (mean value, coefficient of variation % in parentheses). Different letters indicate significant differences among the clones for the analyzed parameter (p < 0.05), according to the Duncan test. (Nvg mm-2): Number of vessel groups per mm2 of xylem; (Ng): Number of vessels in one group.
| Clone | Kh × 10-6 (kgm MPa-1 s-1) |
Vi | Vg | Nvg mm-2 | Ng |
|---|---|---|---|---|---|
| 665 | 0.46 (42.1) bcd | 0.15 (32.7) de | 1.27 (23.2) ab | 45.2 (35.3) abc | 5.87 (33.7) ab |
| 129-81 | 0.37 (35.2) cd | 0.16 (27.4) cde | 1.38 (18.7) a | 52.0 (44.0) ab | 7.60 (30.7) a |
| 181-81 | 0.31 (46.3) d | 0.15 (23.8) de | 1.14 (20.5) b | 57.6 (17.8) a | 7.17 (21.7) ab |
| Antonije | 0.43 (41.7) bcd | 0.25 (32.2) b | 1.13 (12.6) b | 25.0 (38.7) d | 3.17 (33.8) d |
| Bora | 0.54 (31.6) ab | 0.21 (25.7) bc | 1.23 (21.3) ab | 30.8 (18.6) cd | 3.89 (17.7) cd |
| B-81 | 0.48 (31.3) abc | 0.45 (23.9) a | 1.18 (18.9) ab | 16.9 (37.0) d | 2.17 (43.0) d |
| Pannonia | 0.38 (24.8) cd | 0.12 (17.2) e | 1.28 (9.7) ab | 51.4 (50.6) ab | 6.97 (51.7) ab |
| PE19-66 | 0.63 (20.0) a | 0.22 (23.3) bc | 1.23 (21.8) ab | 41.4 (32.7) bc | 5.37 (29.9) bc |
| PE4-68 | 0.44 (35.3) bcd | 0.19 (21.0) bcd | 1.14 (15.7) b | 40.1 (40.1) bc | 5.43 (39.2) bc |
Vulnerability index ranged from 0.12 in clone Pannonia to 0.45 in clone B-81 (Tab. 2). Clone B-81 exhibited a significantly higher Vi value due to the presence of large-lumen vessels. In contrast, clones Pannonia, 129-81, 181-81, and 665, characterized by small-lumen vessels, had correspondingly low Vi values. The correlation analysis revealed that vessel vulnerability strongly depends on various xylem and vessel characteristics (Tab. 3). It was significantly positively correlated with the proportion of xylem and with the xylem-to-phloem ratio. Vessel diameter/lumen size was found to significantly increase vulnerability, whereas vessel frequency exhibited a strong negative correlation with it. Additionally, all vessel-grouping parameters were significantly negatively correlated with Vi.
Tab. 3 - Correlation coefficients between stem anatomical parameters and Kh and Vi. Marked correlations (*) are significant at p < 0.05.
| Parameter | Vi | p-value | Kh | p-value |
|---|---|---|---|---|
| % periderm | -0.40* | 0.000 | -0.30* | 0.005 |
| % primary cortex | -0.48* | 0.000 | -0.31* | 0.004 |
| % secondary phloem | -0.44* | 0.000 | -0.36* | 0.001 |
| % secondary xylem | 0.45* | 0.000 | 0.41* | 0.000 |
| % pith | 0.15 | 0.173 | -0.02 | 0.830 |
| xylem/phloem ratio | 0.59* | 0.000 | 0.42* | 0.000 |
| Vessel frequency (VF) | -0.65* | 0.000 | -0.36* | 0.000 |
| Vessel lumen area (VL) | 0.87* | 0.000 | 0.79* | 0.000 |
| Vessel diameter (VD) | 0.85* | 0.000 | 0.80* | 0.000 |
| % of solitary vessels in xylem | -0.09 | 0.387 | 0.45* | 0.000 |
| % of grouped vessels in xylem | -0.30* | 0.005 | 0.32* | 0.002 |
| % of all vessels in xylem | -0.32* | 0.002 | 0.64* | 0.000 |
| No. of vessel groups/mm2 of xylem | -0.69* | 0.000 | -0.33* | 0.002 |
| No. of vessels per group | -0.72* | 0.000 | -0.34* | 0.001 |
| Kh | 0.42* | 0.000 | - | - |
| Vi | - | - | 0.42* | 0.000 |
| Vg | -0.54* | 0.000 | -0.27* | 0.012 |
The Kh values ranged from 0.31 · 10-6 to 0.63 · 10-6 kg m MPa-1 s-1, with the highest values observed in clones PE19-66, Bora, and B-81 (Tab. 2). Clone PE19-66 had the highest percentage of relatively wide vessels, resulting in the largest Kh value. Similar lower Kh values, without statistically significant differences, were observed in the remaining clones. The correlation coefficients indicated that water conductivity is significantly positively influenced by the proportion of xylem, the xylem-to-phloem ratio, and the percentages of solitary and grouped vessels within the xylem (Tab. 3). VL and VD showed highly positive and significant correlations with Kh. Vessel frequency, however, played a critical role in conductivity, as it was negatively correlated with Kh. Moreover, several parameters associated with vessel grouping, such as the number of vessel groups and the number of vessels per group, were significantly negatively correlated with Kh. It is important to note that Kh was significantly positively correlated with Vi, indicating a trade-off between water conductivity and cavitation resistance.
PCA analysis indicated that, because the stem anatomical parameters exhibited similar patterns, the studied clones could not be separated based on their variation. The parameters exhibiting the greatest variability and showing the highest loading on the first principal axis were the percentages of primary cortex and secondary xylem, the xylem/phloem ratio, VF, Vi, and parameters related to grouped vessels (Tab. 4). Together, these factors explained 39.5% of the total variation. Variability along the second axis was significantly explained by the percentage of grouped vessels in the xylem, which accounted for an additional 18.4% of the variance.
Tab. 4 - Principal Component Analysis based on quantitative stem anatomical parameters - factor loadings on the three principal axes (*: loadings ≥ 0.7). Only characters that significantly contributed to variation are listed.
| Parameter | PCA 1 | PCA 2 | PCA 3 |
|---|---|---|---|
| % primary cortex | -0.712* | -0.334 | -0.150 |
| % secondary xylem | 0.720* | 0.527 | 0.076 |
| xylem/phloem ratio | 0.789* | 0.452 | 0.100 |
| Vessel frequency (VF) | -0.865* | -0.034 | -0.126 |
| % of solitary vessels in xylem | 0.124 | -0.165 | -0.884* |
| % of grouped vessels in xylem | -0.341 | 0.790* | -0.095 |
| % of all vessels in xylem | -0.181 | 0.520 | -0.810* |
| Number of vessel groups/ mm2 of xylem | -0.794* | 0.521 | 0.189 |
| Number of vessels per group | -0.810* | 0.502 | 0.171 |
| Vi | 0.830* | -0.127 | 0.130 |
| % Total Variance Explained | 0.395 | 0.184 | 0.140 |
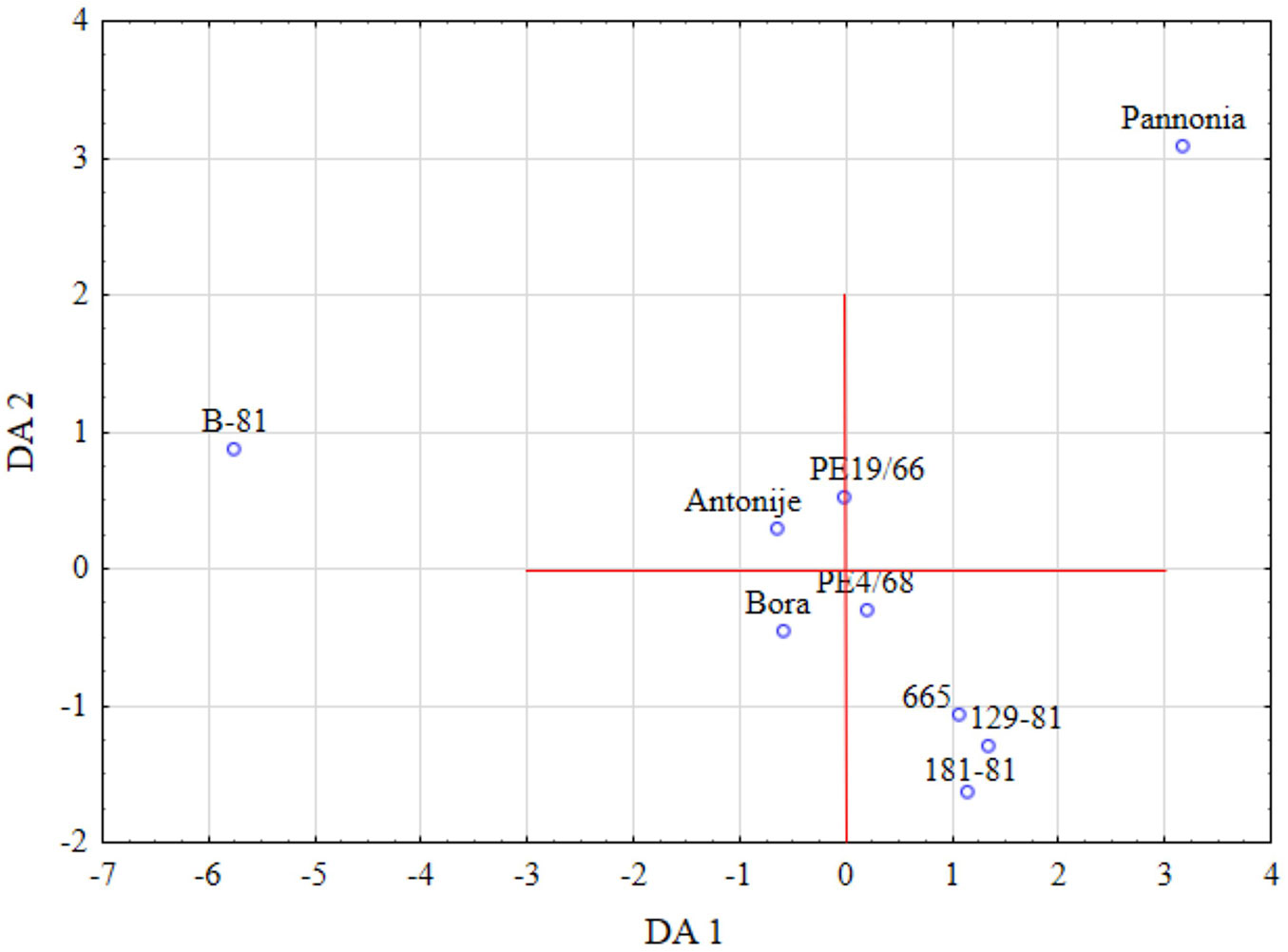
Discriminant analysis revealed that the xylem-to-phloem ratio and Vi were the primary factors discriminating the clones along the first axis, accounting for 53.6% of the discrimination. Clone B-81 was distinctly separated from all other clones along this axis (Fig. 5). Clone Pannonia, as the other clone with extreme values, was positioned on the opposite side of the graph from B-81. The second discriminant axis was defined by parameters related to grouped vessels (the number of vessels per group and Vg), which accounted for an additional 18.1% of the discrimination. These parameters grouped clones 665, 129-81, and 181-81 as those with a high number of vessel groups and a high number of vessels per group.
Fig. 5 - Results of the discriminant analysis, showing the clone grouping based on quantitative xylem characters along the first two discriminant axes.
Discussion
A comparative study of quantitative stem anatomical characters, particularly those of the xylem, in poplar clones revealed significant variability, which could serve as a useful starting point for improving poplars through selection and breeding programs. Anatomical data on xylem and vessel characteristics could serve as new parameters for programs that focus on developing more productive poplar genotypes with enhanced growth potential, improved water-transport efficiency and safety, increased drought tolerance or resistance to cavitation, and advanced breeding strategies.
Because the primary function of the xylem is water transport, its anatomical properties are crucial to plant growth. Tree growth rate is not always correlated with xylem proportion; therefore, more precise data on the properties of xylem elements, especially vessels, are crucial for determining and predicting tree growth ([40]). Xylem also contains mechanical and parenchyma cells, which do not primarily conduct water, and therefore do not affect tree productivity. Consequently, a higher proportion of xylem in poplars does not necessarily imply higher productivity. Among the examined clones, clone B-81 and Bora exhibited the highest xylem proportion and xylem/phloem ratio, accompanied by weakly developed peripheral tissues. In contrast, clone Pannonia exhibited well-developed peripheral tissues and phloem, but low xylem proportion, resulting in the lowest xylem/phloem ratio. This observation aligns with other anatomical data on tree growth rates, suggesting that xylem proportion may, to some extent, predict poplar growth.
Previous studies have shown a positive correlation between vessel lumen size and plant growth potential ([4], [38], [19], [40]). Hydraulic conductance is influenced by vessel diameter and number, and is directly proportional to the fourth power of the sum of the vessel diameters ([31]). Previous research suggests that the growth of many species is linked to both vessel lumen size and vessel number ([28], [18]). This connection arises because larger lumen vessels facilitate water transport, increase transpiration rates, and, consequently, increase photosynthetic rates. The key parameters for predicting cherry tree vigor were identified as vessel frequency, vessel lumen area, and the percentage of vessels in the wood cross-section ([40]). Tombesi et al. ([35]) reported that vigorous peach rootstocks had larger vessel diameters, but a smaller number of vessels per visual field, while Olmstead et al. ([21]) proved that smaller and fewer vessels in the scion and graft union contribute to reduced scion growth of cherry trees. However, Guet et al. ([9]) argued that the relationship among cavitation resistance, hydraulic conductivity, and plant growth is open to multiple interpretations, as these traits are often unrelated across tree species. In their study on Populus nigra, they found no evidence of a trade-off between xylem cavitation resistance and growth, but observed that xylem density was negatively correlated with shoot height increment and tended to scale negatively with total above-ground dry mass. They attributed this to the weak though significant relationship between xylem density and cavitation resistance, indicating that variation in xylem density explains only a small portion of the variation in cavitation resistance and growth. These findings suggest that additional xylem traits, beyond vessel lumen size and vessel density, likely contribute to determining tree growth. Among the examined poplar clones, B-81, Bora, and PE 19-66 were the most promising based on their xylem parameters. Klašnja et al. ([14]) investigated the potential of poplar clones for energy biomass production, showing high productivity of clones PE 19-66, B-81, and Bora (labeled B-229 in their study). The high growth potential of these clones is also documented in phytoremediation research ([23], [24], [25]). These clones shared some common xylem properties. While all three clones had low to medium frequencies of large-lumen vessels, B-81 had the lowest total proportion of vessels because vessel groups were sparse, averaging only 2.2 vessels per group. These characteristics were shared by clone Bora, whilst PE19-66 had the highest %V value, due to the significant contributions of both solitary and grouped vessels. For these reasons, the three clones had the highest Kh values, ranging from 0.48 to 0.63 × 106 kg m MPa-1 s-1. It can thus be concluded that the presence of large lumina and predominantly solitary vessels (albeit at low to medium frequency) facilitates more efficient water transport, which is expected to stimulate tree growth. Following the same logic, Pannonia is supposed to be the least productive among the examined clones. It was characterized by xylem composed of numerous small-lumen vessels, with a large number of grouped vessels per mm2. Its low Kh value was attributed to greater resistance to water transport by small-lumen vessels. The resulting reduction in water transport could lead to slower tree growth and lower productivity. Therefore, when selecting poplar clones for maximizing growth potential, priority should be given to those with high hydraulic conductivity (Kh > 0.5 × 10-6 kg m MPa-1 s-1), large-lumen vessels (>1000 µm2), and a high proportion of xylem tissue (>55%). However, such clones are also more vulnerable to cavitation and are not recommended for cultivation in arid regions, as the risk of cavitation increases under drought conditions, unless adequate irrigation is provided. For breeders seeking clones with high growth potential, the most suitable option is one that exhibits the mentioned anatomical traits. Among the clones studied, clone B-81 stands out for the most suitable combination of stem anatomical characters and promising growth potential, though its cultivation is advisable only in mesic environments due to its high vulnerability to cavitation.
In the examined clones, the number of vessels was inversely proportional to vessel lumen size and, consequently, to the theoretical hydraulic conductance. This result is consistent with the findings reported by Fan et al. ([8]), which indicate a similar relationship in tropical tree species, and is supported by the observations of Tombesi et al. ([35]). In most tree species, vessel diameter is strongly negatively related to vessel frequency but positively related to hydraulic conductivity ([33], [8], [7]).
Studies suggest that larger-lumen vessels increase susceptibility to cavitation and embolism and compromise the plant’s mechanical strength ([20], [11]). Cavitation and the resulting embolism reduced xylem hydraulic conductivity ([32], [9]). According to Awad et al. ([1]), easily measurable anatomical parameters, such as vessel diameter and vessel wall thickness, are relevant traits for assessing vulnerability to cavitation. The vulnerability index is used to assess the biological safety of vessels ([31]). The correlation analysis revealed that vessel vulnerability was heavily influenced by various xylem and vessel characteristics. Increased VF appears to be the most important factor improving resistance to cavitation, as it has a statistically significant negative correlation with Vi (r = -0.76), whereas VL and VD were found to significantly increase vulnerability due to strong positive correlations. Plants with more narrow-lumen vessels exhibit lower Vi, thereby making water transport safer and reducing the likelihood of air embolism. Poplars are typically more susceptible to xylem cavitation than other temperate tree species ([1], [32], [11]), as their larger vessels are prone to embolism ([12]). Our results concur with these findings, as the highest vulnerability index was observed in clone B-81, which has the largest-lumen vessels. In clone Pannonia, small-lumen vessels provide substantially better mechanical support and resistance to cavitation and embolism, resulting in the lowest recorded vulnerability index. The most susceptible to cavitation, aside from clone B-81, are clones Bora, Antonije, and PE 19-66.
Plants face a compromise between increasing vessel diameter, which enhances water conductivity, and the vulnerability of these vessels to cavitation. Guet et al. ([9]) established trade-offs between xylem resistance to cavitation, hydraulic efficiency, and mechanical strength in Populus nigra. Genotypes of this species that were more resistant to drought-induced cavitation exhibited lower Kh values. This finding aligns with our study, which demonstrated a significant inverse relationship between Vi and Kh in poplar clones. Both parameters are determined by vessel traits, particularly lumen size. High conductivity is primarily associated with large-lumen vessels, which are more susceptible to cavitation. In contrast, genotypes with narrower vessels are less efficient in water transport but exhibit greater resistance to cavitation and can withstand higher mechanical pressure.
Cavitation resistance is closely linked to increased mechanical strength, which may also confer water stress resistance. Schreiber et al. ([32]) observed that taller aspen and hybrid poplar trees in Canada’s boreal region had narrower vessel diameters. They attributed this adaptation to cold climates, designed to prevent embolism caused by winter freezing. Under water stress, wood species tend to reduce vessel diameter and form numerous small-lumen vessels, which lowers hydraulic conductivity but increases resistance to embolism ([2]). This highlights an adaptive response to balance efficiency and safety. However, this combination of characters could compromise growth ([10], [27]). A revision of Carlquist’s work ([5]), conducted by Ewers et al. ([7]), highlighted Carlquist’s identification of several xylem and vessel traits associated with xeromorphy. These included narrow vessels, high vessel density, short vessel elements, vessel grouping, and increased vessel wall thickness. Carlquist emphasized that vessel diameter, density, and element length were the most responsive to environmental selection. He proposed that Vi values below 1, which was the case in our study for all of the examined clones, reflect a high degree of xeromorphy, whereas values approaching 3 indicate true mesomorphy. Awad et al. ([1]) found that decreased vulnerability to cavitation of poplars grown under drought conditions was correlated with decreased vessel diameter, larger density, and increased vessel wall thickness. Guet et al. ([9]) further observed that Populus nigra genotypes that were highly resistant to drought-induced cavitation had narrower vessels, lower theoretical xylem hydraulic conductivity, and lower vessel grouping index. According to the results obtained, the clone most adapted to water stress is Pannonia, as it possesses several drought-resistance anatomical adaptations: well-developed peripheral stem tissues (protective), numerous small-lumen vessels, and low Vi and Kh values. Clones 129-81 and 181-81 could also be considered more drought-tolerant and more resistant to cavitation than other clones. In general, for selecting poplar clones with improved drought tolerance, we recommend those characterized by numerous, small-lumen vessels (VL <800µm2; VF >200 vessels per mm2 of xylem), low hydraulic conductivity (Kh <0.4 × 10-6 kg m MPa-1 s-1), and Vi values below 1, an approach also supported by Carlquist, as cited in Ewers et al. ([7]). Clones exhibiting this combination of stem anatomical traits (e.g., clone Pannonia in this study) should be prioritized for cultivation on arid sites and in dry or cold environments where drought or physiological drought is prevalent.
Vessel grouping provides alternative water pathways when embolism blocks water transport; however, it can also be disadvantageous, as it facilitates cavitation within the same group of vessels ([39]). Zhao ([39]) found that in Betula plathyphylla, vessel grouping was inversely related to vessel width, consistent with our findings for poplar clones, indicating a significant negative correlation between Vg and VL. Zhao ([39]) also found that in B. plathyphylla, vessel grouping improved hydraulic conductivity while reducing xylem vulnerability. Carlquist ([5]) found that the number of vessels per group is higher in plants adapted to dry habitats (about 5.20), whereas the average value for mesophytes is 3.04. We agree that poplar clones having high Vg would be more resistant to vulnerability (a significant negative correlation was recorded between Vg and Vi). However, our results show that Kh and Vg were significantly inversely correlated, suggesting that vessel grouping reduced conductivity. It is important to note that plants with more grouped vessels, more vessel groups in the xylem, and more vessels per group tend to have greater vessel safety and a lower risk of embolism, owing to significant negative correlations among these parameters. More densely distributed vessels contribute to their stability. From this perspective, the most advantageous clone under drought conditions among the examined clones is Pannonia, with a relatively high percentage of grouped vessels (more than seven vessels per vessel group), a high Vg index, and a high frequency of low-lumen vessels.
Solitary vessels occupy the larger part of the xylem and thus play a more significant role than grouped vessels in water transport efficiency. This assertion is echoed by the results reported by Lens et al. ([16]), who found a negative correlation between vessel grouping index and hydraulic conductivity in Aster species. Our findings concur with these observations, as the correlation between these two parameters was significantly negative. It is also worth noting that, according to Loepfe et al. ([17]), hydraulic conductivity and vulnerability to embolism increase with vessel grouping due to connectivity of xylem vascular elements. Additionally, Guet et al. ([9]) found that more cavitation-resistant genotypes of Populus nigra exhibited a lower degree of vessel grouping, though these assertions were not confirmed in our study. The opposite trend observed in our study, reduced vulnerability to cavitation and lower water conductivity associated with vessel grouping, can be explained by the fact that plants with a higher number of grouped vessels also had smaller vessel lumens and higher vessel frequency. A significant negative correlation was found between Vg and VL, and a significant positive correlation between Vg and VF. When small-lumen vessels are grouped, their contribution to overall hydraulic conductivity decreases; however, they provide greater mechanical support and increased resistance to cavitation.
Discriminant analysis along the first two axes distinctly separated two clones, B-81 and Pannonia, which exhibited extreme values in quantitative xylem characteristics, compared to the other examined clones. However, all clones exhibited similar variability in xylem parameters, making further separation by PCA statistically insignificant.
Conclusion
Based on our results, we believe that stem anatomical data and the quantitative parameters related to xylem and vessels could be used as additional criteria in the selection of poplar clones for increased productivity, water use efficiency, drought tolerance or resistance to cavitation. The use of this methodology, combined with physiological, biochemical, and molecular methods, can shorten breeding and selection processes for poplars with desired properties.
From the anatomical perspective, further breeding for increased growth should be targeted to clones with a higher proportion of xylem (xylem/phloem ratio), composed of large-lumen vessels, with a higher proportion of solitary, and a lower percentage of grouped vessels. Breeding for drought-tolerant clones and clones with increased resistance to cavitation should aim to produce clones with more low-lumen vessels, a higher percentage of grouped vessels, and more vessel groups, each composed of a larger number of vessels. The anatomical method described here could be used as a new, efficient means of assessing poplar growth potential and as an additional, readily accessible tool for predicting drought tolerance and cavitation vulnerability.
Data availability
Publicly available source code repository for the image processing: ⇒ https://github.com/Turku-BioImaging/2024-lana-zoric-populus-vasculature
Acknowledgements
The authors gratefully acknowledge the financial support of the Ministry of Science, Technological Development and Innovation of the Republic of Serbia (Grants no. 451-03-137/2025-03/200125 & 451-03-136/20 25-03/200125). We extend our gratitude to the Institute of Lowland Forestry and Environment (ILFE) at the University of Novi Sad, Serbia, for supplying the poplar material used in this analysis. Turku BioImaging, part of the Finnish Advanced Microscopy Node of Euro-BioImaging Finland, is funded by the Research Council of Finland, FIRI 2023 (grant no. 358879 to P.K.). The authors acknowledge Dr. John G. Du Plissis for English language editing.
References
Gscholar
Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Authors’ Info
Authors’ Affiliation
Sonja Davidović
Dunja Karanović 0000-0002-8007-4610
Jadranka Luković 0000-0003-2384-7610
University of Novi Sad, Faculty of Sciences, Department of Biology and Ecology, Novi Sad (Serbia)
University of Minnesota, Natural Resources Research Institute (NRRI), Duluth, MN (USA)
Ramish Bibi
Irina Belaya 0000-0001-8129-3711
Dado Tokic 0009-0007-8471-3006
Pasi Kankaanpää 0000-0002-5890-0883
Turku BioImaging, Abo Akademi University and the University of Turku, Turku (Finland)
Corresponding author
Paper Info
Citation
Zorić L, Pilipović A, Solis J, Bibi R, Belaya I, Tokic D, Kankaanpää P, Davidović S, Karanović D, Luković J (2026). Stem xylem traits in poplar genotypes (Populus L.): additional parameters for selection and breeding. iForest 19: 52-60. - doi: 10.3832/ifor4868-018
Academic Editor
Marco Borghetti
Paper history
Received: Apr 08, 2025
Accepted: Jul 15, 2025
First online: Feb 15, 2026
Publication Date: Feb 28, 2026
Publication Time: 7.17 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2026
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 2458
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 933
Abstract Page Views: 826
PDF Downloads: 601
Citation/Reference Downloads: 0
XML Downloads: 98
Web Metrics
Days since publication: 140
Overall contacts: 2458
Avg. contacts per week: 122.90
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
(No citations were found up to date. Please come back later)
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Earlywood vessel features in Quercus faginea: relationship between ring width and wood density at two sites in Portugal
vol. 8, pp. 866-873 (online: 27 April 2015)
Short Communications
Changes in Populus sp. wood subjected to heat treatment: anatomy and silica content
vol. 18, pp. 223-226 (online: 09 August 2025)
Research Articles
The physicomechanical and thermal properties of Algerian Aleppo pine (Pinus halepensis) wood as a component of sandwich panels
vol. 15, pp. 106-111 (online: 21 March 2022)
Research Articles
Diurnal dynamics of water transport, storage and hydraulic conductivity in pine trees under seasonal drought
vol. 9, pp. 710-719 (online: 21 August 2016)
Research Articles
Interaction between planting spacing and wood properties of Eucalyptus clones grown in short rotation
vol. 14, pp. 12-17 (online: 02 January 2021)
Research Articles
Tree growth, wood and bark water content of 28 Amazonian tree species in response to variations in rainfall and wood density
vol. 9, pp. 445-451 (online: 16 January 2016)
Research Articles
Deploying an early-stage Cyber-Physical System for the implementation of Forestry 4.0 in a New Zealand timber harvesting context
vol. 17, pp. 353-359 (online: 13 November 2024)
Research Articles
Comparison of alternative harvesting systems for selective thinning in a Mediterranean pine afforestation (Pinus halepensis Mill.) for bioenergy use
vol. 14, pp. 465-472 (online: 16 October 2021)
Research Articles
Improving dimensional stability of Populus cathayana wood by suberin monomers with heat treatment
vol. 14, pp. 313-319 (online: 01 July 2021)
Research Articles
Identification of wood from the Amazon by characteristics of Haralick and Neural Network: image segmentation and polishing of the surface
vol. 15, pp. 234-239 (online: 14 July 2022)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







