Multiple ways for enhancing flammability in Mediterranean woody species: a trait-based approach
iForest - Biogeosciences and Forestry, Volume 18, Issue 5, Pages 301-308 (2025)
doi: https://doi.org/10.3832/ifor4601-018
Published: Oct 20, 2025 - Copyright © 2025 SISEF
Research Articles
Abstract
Enhanced flammability is a key driver of community assembly and a favored characteristic in fire-dependent ecosystems. In this study, we examined the variability of flammability traits among Mediterranean woody species. We measured eight flammability traits, including leaf traits, branching architecture, and standing biomass traits, across 26 woody species in the understory of pine forests in southwestern Anatolia, Turkey. Given that these species are part of fire-dependent plant communities, where fire provides advantages for persistence, we hypothesized that species would use various ways to increase their flammability. All flammability traits exhibited substantial variation among species. The flammability of the studied species varied depending on the specific trait considered, with no single species consistently exhibiting higher or lower flammability across all traits. Moreover, many species employed distinct strategies to enhance their flammability, such as differences in leaf structure, branching patterns, and biomass characteristics. The interspecific variability in flammability was also evident through a calculated flammability score based on eight traits, providing a valuable framework for ranking species by their flammability. Our findings suggest that the traits driving increased flammability in Mediterranean woody plants are idiosyncratic at the species level. Therefore, multiple traits must be considered to comprehensively study flammability within a plant community. These results have practical implications for the selection of “fire-resistant” plants in land management strategies within the Mediterranean Basin.
Keywords
Among-species Variability, Fire, Flammability, Functional Traits, Mediterranean Basin
Introduction
Many biomes on Earth exhibit flammable vegetation due to climatic seasonality, the accumulation of aboveground biomass, and the presence of ignition sources, either of natural or anthropogenic origin ([16], [44], [22], [32], [23], [30]). In these fire-prone ecosystems - landscapes frequently experience fire, which is a critical factor in shaping vegetation dynamics - several plant traits have been shaped by fire regimes over thousands or even millions of years. Traits such as serotiny ([21]), resprouting ability ([4]), thick bark ([34]), and fire-stimulated germination ([20]) enable plants to survive or regenerate after fires. Another group of plant traits, known as flammability traits, determines how easily a species ignites, sustains combustion, and contributes to fire intensity, making species more (or less) flammable ([17]). Flammability traits significantly influence local fire regimes and therefore have significant ecological and socioeconomic consequences ([38]). Enhanced flammability can provide species with an advantage in fire-prone ecosystems, but only if they also possess traits that support post-fire survival or regeneration ([3]). Moreover, since flammability traits are under the selection pressure of local or regional fire regimes, among-population variability in flammability traits can also be observed ([33], [36]). This is because flammable species often exhibit additional adaptations, such as post-fire germination or resprouting ability, that enhance their recovery after fire events ([33]). As a result, fire regimes play a critical role in shaping both flammability traits and associated regeneration strategies in plant species.
Mutch ([28]) was the first to propose that the degree of flammability in plants is influenced by the extent to which the plant community is fire-dependent, suggesting that characteristics enhancing flammability are shaped by natural selection. However, Snyder ([49]) criticized this hypothesis, arguing that increased flammability may result from the selection of traits serving other functions, such as drought tolerance. Troumbis & Trabaud ([53]) questioned the validity of Mutch’s hypothesis and outlined specific criteria needed to support the selection of flammability traits, including a genetic basis and underlying mechanisms. Bond & Midgley ([3]) later proposed that flammable vegetation arises when flammable species gain a fitness advantage over less flammable neighbors in post-fire environments. A simulation model by Schwilk & Kerr ([47]) suggested that increased flammability can evolve as an adaptive strategy, even at the cost of sacrificing parent plants. More recently, empirical evidence suggests that flammability is a variable biological trait subject to natural selection ([43], [9], [33], [25]). These studies are major steps toward understanding the evolution and ecology of flammability in fire-prone ecosystems ([36]).
Traits that increase plant flammability contribute to the overall flammability of plant communities and vegetation in fire-prone ecosystems, shaping fire dynamics at both community and ecosystem levels. Understanding these traits is crucial for identifying their distribution within plant communities and gaining insights into the ecology and evolution of fire-prone vegetation. Additionally, as flammability traits vary among species, this knowledge can guide forest managers in selecting less flammable species for forest restoration efforts ([29], [2], [42]). While many studies have examined plant flammability at the species level, our understanding of the variability of flammability traits across species remains limited, especially at broader ecological scales, from plant communities to biomes ([33], [6], [45], [50]).
There are many approaches to studying flammability in plants. Conceptually, plant flammability can be measured by estimating parameters such as first ignition time, smoke temperature, heat release rate, burning time, maximum temperature, and burnt biomass of plant leaves or shoots when exposed to a heat or ignition source ([26], [1], [5], [27], [23], [13]). Alternatively, flammability can be assessed by burning a large section of plant material from several species within the same community to evaluate the vegetation-level flammability ([19], [1]). Another approach involves estimating fuel loading in plant individuals ([43]) or examining specific plant traits ([1], [27]).
Several plant traits are associated with flammability ([40]). Fuel moisture content is a substantial component of plant flammability ([52], [39], [10], [26]) and is the most frequently studied trait ([40]). Leaf traits, such as leaf dry matter content and leaf thickness, are also strongly linked to flammability ([38], [11]). Leaf curling is another important flammability parameter, as curled and larger leaves reduce fuel bed bulk density by generating air spaces between fuel particles, which increases flammability ([9], [31]). Branching architecture, particularly branch density, is a key trait associated with flammability ([38]), as dense branching structures increase the exposed surface area, resulting in more flammable canopies ([3]). Similarly, plant architecture, including the relative proportions of coarse to fine and dead to live standing fuel biomass, is a critical component of flammability, influencing fire intensity ([46], [43], [51]). Associations among flammability traits are also notable. For instance, the relationship between fuel moisture content and flammability can depend on leaf dry matter content ([48], [38]). While multiple traits drive the flammability levels of plant individuals and species, most studies focus on only a few traits ([46], [44], [43], [33], [26], [40]), and studies considering several flammability traits simultaneously remain relatively rare ([19], [1], [5], [23]).
Mediterranean Basin ecosystems are characterized by fire-prone vegetation ([18]) that exhibits diverse fire adaptations, including traits that enhance survival, regeneration, or reproduction following fire events. Examples of these adaptations include the serotinous cones of low-altitude pines, the high resprouting ability of shrubs, post-fire stimulation of flowering in geophytes, thick bark in high-altitude pines, and fire-stimulated germination triggered by heat or smoke in many herbaceous and woody species ([18], [51]). Flammability is another key fire adaptation associated with Mediterranean plants ([23]). In the Mediterranean Basin, various aspects of plant flammability have been extensively studied. Comparative studies have frequently examined the flammability levels of species within plant communities ([29], [8]). Functional hypotheses exploring the relationship between flammability traits and other plant traits have also been tested ([43], [2], [35], [23]). Furthermore, the role of fire regimes in shaping flammability has been acknowledged in Mediterranean ecosystems ([33]).
Although recent studies have revealed different aspects of plant flammability, gaps remain in understanding the patterns of variability in flammability traits across many ecosystems. In this study, we aimed to characterize the interspecific variability in flammability traits in Mediterranean woody species, anticipating significant trait variation among species. We employed a trait-based approach to assess the flammability of species, focusing on fuel loading and the characteristics of leaves and branches. Given that the study species are part of fire-dependent plant communities where fire provides advantages for persistence ([51]), we hypothesized that species adopt different strategies to enhance flammability. Specifically, we expected the traits responsible for increased flammability in Mediterranean plants to be species-specific. To test this hypothesis and clarify variability in flammability traits among Mediterranean species, we examined several leaf traits, branching architecture, and fuel loading properties in 26 woody plant species dominating the understory of Mediterranean pine forests in southwestern Turkey.
Methods
Study area and sampling design
The study was conducted in southwestern Anatolia, a region characterized by Mediterranean vegetation. The area experiences a Mediterranean climate, with dry summers and mild, wet winters, as classified by the Köppen system ([37]). Frequent crown fires, originating from both natural and anthropogenic sources, make this region one of the most fire-prone areas in Turkey ([30]). The fire regime in the region is primarily shaped by its climatic conditions, particularly summer drought, and the dominance of highly flammable vegetation types, including pine forests, maquis shrublands, and scrublands. These fires play a critical role in driving vegetation dynamics and influencing the composition and structure of local plant communities.
We selected 24 study sites, each 1 ha in size, within the study area to sample woody species for their flammability traits (Tab. S1 in Supplementary material). Although several burned sites exist across the region, all selected sites were mature Turkish red pine (Pinus brutia Ten.) forests that had remained unburned for at least 50 years. The study sites spanned elevations from sea level to over 1000 m and were situated on limestone bedrock.
We selected the five most dominant understory species at each study site to sample for their flammability traits. The dominant pine species (Pinus brutia) was excluded due to methodological limitations, as its height and large biomass made it impractical to sample for whole-plant and combustibility traits. For each of the five species at each site, three individuals were randomly selected for sampling. In sites with low woody species richness, we sampled fewer species, as it was not possible to identify five dominant woody species. In total, we measured and collected samples from 363 individuals representing 26 plant species across the study sites (Tab. 1, Tab. S2 in Supplementary material). These species belong to different taxonomic families and growth forms, including trees, shrubs, and lianas. Most of the sampled species are capable of resprouting after crown fires, while a few lack this ability (Tab. 1). Since not all species were present at every site, the number of individuals sampled per species varied from 3 to 69 (Tab. S2).
Tab. 1 - Plant species studied for flammability traits. GF is growth form, Resp indicates resprouting ability (yes: resprouter; no: non-resprouter), and Code shows the species acronyms used in the tables and graphs throughout the paper.
| Species | Family | Code | GF | Resp |
|---|---|---|---|---|
| Arbutus andrachne L. | Ericaceae | Aan | large shrub | Yes |
| Arbutus unedo L. | Ericaceae | Aun | large shrub | Yes |
| Asparagus aphyllus L. | Asparagaceae | Aap | liana | Yes |
| Astragalus sp. | Fabaceae | Ast | subshrub | Unknown |
| Cistus creticus L. | Cistaceae | Ccr | shrub | No |
| Cistus salviifolius L. | Cistaceae | Csa | shrub | No |
| Crataegus monogyna Jacq. | Rosaceae | Cmo | large shrub | Yes |
| Daphne gnidioides Jaub. & Spach | Tymelaeaceae | Dgn | shrub | Unknown |
| Euphorbia characias L. | Euphorbiaceae | Ech | subshrub | Yes |
| Genista acanthoclada DC. | Fabaceae | Gac | shrub | Yes |
| Hypericum empetrifolium Willd. | Hypericaceae | Hem | subshrub | Yes |
| Juniperus oxycedrus L. | Cupressaceae | Jox | large shrub | Yes |
| Laurus nobilis L. | Lauraceae | Lno | large shrub | Yes |
| Myrtus communis L. | Myrtaceae | Mco | large shrub | Yes |
| Olea europaea L. | Oleaceae | Oeu | tree | Yes |
| Origanum onites L. | Lamiaceae | Oon | subshrub | Yes |
| Phlomis grandiflora H.S.Thomps. | Lamiaceae | Pgr | shrub | Unknown |
| Phlomis lycia D. Don | Lamiaceae | Ply | shrub | Yes |
| Phillyrea latifolia L. | Oleaceae | Pla | large shrub | Yes |
| Pistacia lentiscus L. | Anacardiaceae | Ple | large shrub | Yes |
| Pistacia terebinthus L. | Anacardiaceae | Pte | large shrub | Yes |
| Quercus cerris L. | Fagaceae | Qce | tree | Yes |
| Quercus coccifera L. | Fagaceae | Qco | large shrub | Yes |
| Spartium junceum L. | Fabaceae | Sju | large shrub | Yes |
| Styrax officinalis L. | Styracaceae | Sof | large shrub | Yes |
| Teucrium divaricatum Sieber ex Heldr. | Lamiaceae | Tdi | subshrub | Yes |
Trait measurements
We performed trait measurements on individuals of the selected species both in the field and in the laboratory. To capture multiple components of plant flammability, we measured four leaf traits (leaf dry matter content, leaf moisture content, leaf thickness, and leaf curliness), one whole-plant trait (branching architecture), and three combustibility traits (dead to live fuel ratio, coarse to fine fuel ratio, and dead fine fuel proportion - Tab. 2). We followed published protocols to sample, measure, and estimate traits, drawing on a standardized methodology paper ([38]), plant trait database studies ([51], [15]), and specific research articles ([8], [9], [26], [31], [12]) - Tab. 2).
Tab. 2 - Flammability traits used in the study, along with their units, definitions, measurement or estimation methods, and related references. References for field or laboratory measurements specifically used in our study are underlined.
| Trait | Trait full name | Unit | Definition of trait | Measuring/Estimation | References |
|---|---|---|---|---|---|
| LDMC | Leaf dry matter content | mg g-1 | The ratio of the dry mass of the leaf to the corresponding water-saturated fresh mass | LDMC = Leaf dry weight / water-saturated leaf weight | Pérez-Harguindeguy et al. ([38]), Kattge et al. ([15]) |
| LMC | Leaf moisture content | % | Percentage of water in a leaf determined as a ratio of dry weight | LMC= [(fresh weight-dry weight)/dry weight)]·100 | De Lillis et al. ([7]) |
| LT | Leaf thickness | mm | The thickness of an individual leaf | Digital slide caliper | Engber & Varner ([9]), Pérez-Harguindeguy et al. ([38]), Grootemaat et al. ([12]), Kattge et al. ([15]) |
| LC | Leaf curliness | mm | The distance of a leaf on flat ground up to the highest curling point | Millimeter ruler | Engber & Varner ([9]), Parsons et al. ([31]), Grootemaat et al. ([12]) |
| BR | Branching architecture | cm | The ratio of the length of the lateral branches of the species to the corresponding number of branches | BR = Length of the lateral branches/number of branches | Pérez-Harguindeguy et al. ([38]) |
| DLF | Dead to live fuel ratio | Ratio | Dead to live fuel biomass ratio | DL = (Dead fine fuel biomass [f. b.] + Dead coarse f.b.) / (Live fine f.b. + Live coarse f.b.) |
Saura-Mas et al. ([43]) |
| CFF | Coarse to fine fuel ratio | Ratio | Coarse (≥ 6 mm diameter) to fine fuel (< 6 mm diam.) biomass ratio, including live and dead material |
CFF = (Live coarse f.b. + Dead coarse f.b.) / (Live fine f.b. + Dead fine f.b.) | Saura-Mas et al. ([43]), Tavsanoglu & Pausas ([51]) |
| DFF | Dead fine fuel amount | % | Standing fine (< 6 mm diam.) dead biomass attached to the plant during the dry season expressed in proportion of the total aboveground biomass | DFF = (Dead fine f.b. / Total biomass)·100 | Saura-Mas et al. ([43]), Tavsanoglu & Pausas ([51]) |
We sampled ten mature, undamaged leaves from each plant for leaf trait measurements. For species with smaller leaves, twigs containing more than ten leaves were collected to retain leaf moisture content until measurements were conducted. We placed these leaves in tea filter bags along with a piece of moistened filter paper. We later placed three filter bags from individuals of each species at a study site into air-inflated, sealed plastic bags. These were stored in a cooler during fieldwork and then in a refrigerator until measurements were conducted.
Leaf curliness and leaf thickness measurements were performed before measuring leaf fresh mass. Leaf curliness (LC) was determined by measuring the distance from the leaf surface to its highest curling point on a level surface using a millimeter ruler ([9], [31], [12] - Tab. 2). Average curliness values of ten leaves from each individual were calculated. Leaf thickness (Lt) was measured with a digital micrometer positioned at the center of the leaf, avoiding leaf veins ([38] - Tab. 2). We measured fresh weights of leaves using a digital scale within 24 h of sampling. The same leaves were oven-dried at 70 °C for 72 h to determine their dry weights ([38]). Leaf moisture content (LMC) was calculated as the ratio of the difference between fresh and dry weights to the dry weight ([26]), while leaf dry matter content (LDMC) was calculated as the ratio of dry mass to water-saturated fresh mass ([38]).
We examined two main branches per plant in the field to determine branching architecture (BR). For each main branch, we counted the number of lateral branches and measured the main branch length. Then, we calculated branching architecture as the number of lateral branches per unit length of the main branch (Tab. 2).
To reveal the combustibility component of plant flammability, we estimated standing fuel loading by measuring fine and coarse, as well as live and dead, standing plant material for each sampled individual. We collected biomass samples from the same individuals used for leaf sampling in the field. Immediately after collection, we sorted the living and dead plant material into separate paper bags and stored the samples in a dark, low-humidity box until transport to the laboratory. In the laboratory, we further divided the material into two fuel classes based on diameter: fine (<6 mm) and coarse (>6 mm). All leaf material was classified as fine fuel. This process resulted in data for four standing fuel classes for each individual: fine dead, fine live, coarse dead, and coarse live. The classified plant material was then dried at 80 °C for 48 h in a temperature-controlled oven to determine the dry mass of each fuel class ([38]). Using these data, we calculated three combustibility traits ([43], [51] ): dead to live fuel ratio (DLF), coarse to fine fuel ratio (CFF), and dead fine fuel proportion (DFF - Tab. 2).
The studied traits are among the most commonly used to assess plant flammability ([38] - see also references in Tab. 2) and exhibit variable relationships with it. Specifically, flammability increases with LDMC, LC, DLF, and DFF, while it decreases with LMC, BR, LT, and CFF ([46], [44], [43], [33], [26], [38]).
Data analysis
We used descriptive statistics and boxplots to examine the variability in flammability traits among species. Trait data were tested for normality and homoscedasticity using the Shapiro-Wilk test and histograms. Data were log-transformed prior to all analyses to better approximate normality and ensure homogeneity of variances. We performed all analyses in the R environment (⇒ https://www.r-project.org/).
For each trait, we performed a one-way analysis of variance (ANOVA) to identify differences among species, followed by Tukey’s HSD test for multiple comparisons. Based on significant differences identified through post-hoc tests (P < 0.05), we assigned each species to one of the following flammability level groups for each trait: high, moderate-high, moderate, moderate-low, and low. Threshold values for each flammability level group for each trait were determined using this assignment procedure. However, in a few cases where sample sizes were too small to detect significant differences, exceptions to this rule were made during the grouping process. To visualize flammability levels, we used different colors to represent the groups in trait variability graphs. This approach allowed us to identify specific groups of relative flammability levels by considering the relationship of each trait with flammability (i.e., some traits had a positive relationship, while others had a negative one). Additionally, we used one-way ANOVA to analyze among-species differences in the proportion of fuel fractions of standing biomass for each trait. We used a percent stacked bar chart to display the proportion of fuel fractions for each species.
To calculate the “flammability score”, representing the overall flammability of each species, we first assigned species to flammability level groups (low: 1, moderate-low: 2, moderate: 3, moderate-high: 4, and high: 5) for each trait based on thresholds derived from ANOVA and Tukey HSD tests (see Tab. S4 in Supplementary material). These thresholds were determined by dividing the trait ranges into meaningful intervals that reflected observed ecological variation. For highly correlated traits (e.g., DLF and DFF; r > 0.70 - Fig. S1), their contributions were averaged to avoid overrepresentation in the flammability score. The flammability level scores across traits were summed to calculate the overall flammability score, with equal weight assigned to each trait. This integrative approach enabled us to compare the relative flammability of species while minimizing biases arising from redundant trait information.
Results
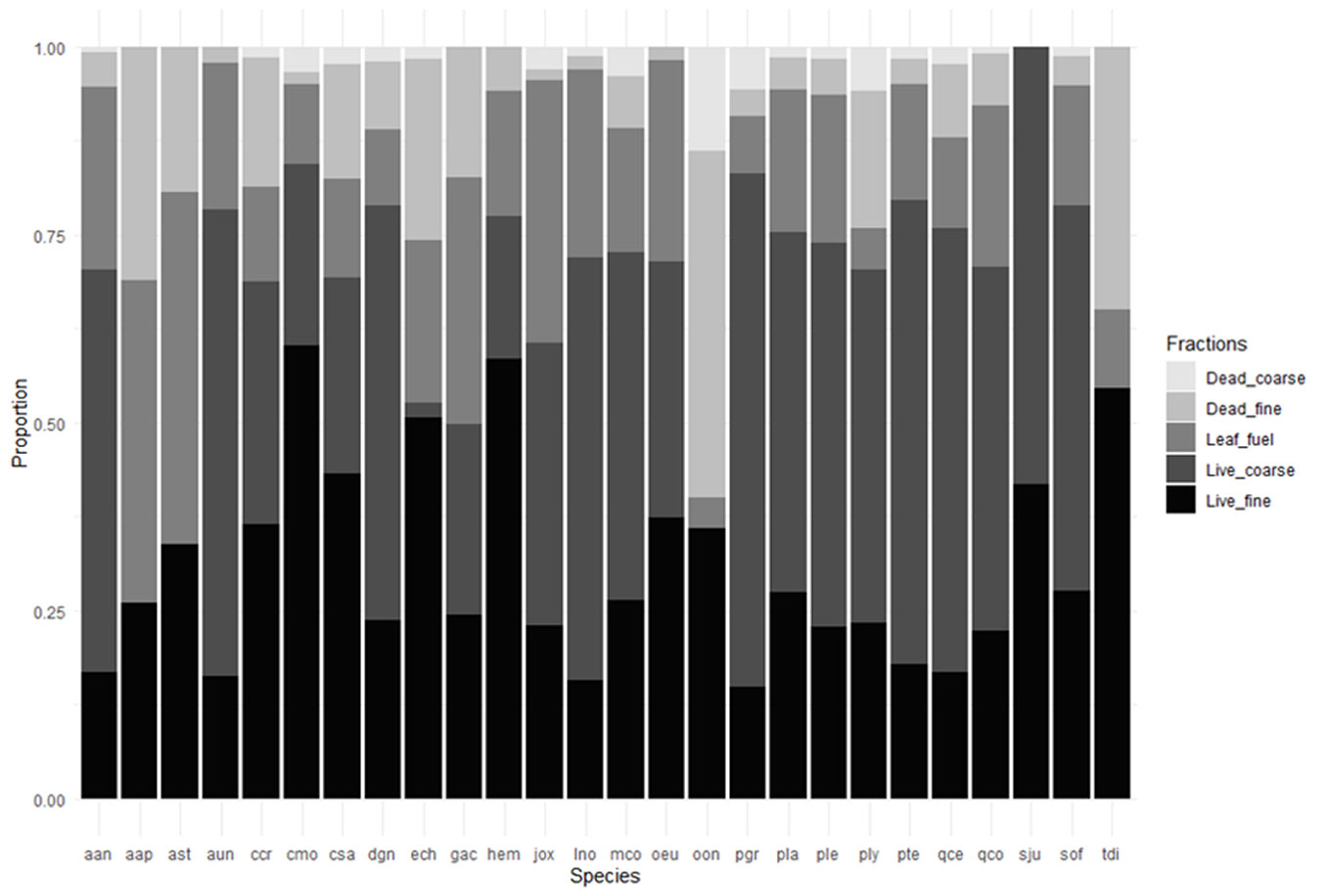
All flammability traits showed substantial variation among the studied species (P < 0.0001 - Tab. 3; Figs. S2-S9, Tab. S3 in Supplementary material). The range of leaf trait values varied considerably across species, with a sixfold difference observed for leaf dry matter content and a 31-fold difference for leaf moisture content. Branching architecture was among the most variable traits, exhibiting approximately a 36-fold difference between minimum and maximum values. Fuel loading traits also displayed pronounced variability among species, ranging from zero (i.e., lacking dead, coarse, or dead fine fuels) to 4.2 and 16.0 for DLF and CFF, respectively, and a 60.0% value for DFF (Tab. 3, Fig. 1).
Tab. 3 - Descriptive statistics of among-species variability in flammability traits. Results of one-way ANOVAs testing differences among species for each trait are also provided. n refers to the number of species considered for each trait. Species (codes as listed in Tab. 1) with minimum and maximum values are indicated in parentheses.
| Trait | n | Min | Median | Mean | Max | F | P |
|---|---|---|---|---|---|---|---|
| Leaf dry matter content (mg g-1) | 22 | 150.8 (ply) |
576.7 | 557.4 | 934.0 (qco) |
52.4 | <0.0001 |
| Leaf moisture content (%) | 22 | 9.6 (pte) |
100.2 | 115.3 | 298.4 (ech) |
22.6 | <0.0001 |
| Branching (cm) | 26 | 1.080 (ast) |
7.345 | 9.206 | 38.5 (ech) |
39.5 | <0.0001 |
| Leaf curliness (mm) | 22 | 0.16 (pla) |
0.52 | 0.60 | 2.72 (aan) |
60.6 | <0.0001 |
| Leaf thickness (mm) | 22 | 0.14 (jox) |
0.27 | 0.31 | 1.02 (ply) |
52.4 | <0.0001 |
| Dead to live fuel ratio | 26 | 0 (sju) |
0.09 | 0.23 | 4.23 (aap) |
4.6 | <0.0001 |
| Coarse to fine fuel ratio | 26 | 0 (aap,ast,tdi) |
1.67 | 2.02 | 16.0 (dgn) |
34.2 | <0.0001 |
| Dead fine fuel amount (%) | 26 | 0 (sju) |
5.61 | 9.14 | 60.0 (oon) |
10.1 | <0.0001 |
Fig. 1 - The proportion of plant fractions (dead and live, fine and coarse materials, and leaves) based on total dry weight for each species. Species codes are provided in Tab. 1.
Species exhibited varying levels of flammability across different traits, with no single species consistently showing higher or lower flammability across all traits. Consequently, many species ranked at the low end of the flammability continuum for one trait while appearing at the highly flammable end for another (Fig. 2, Figs. S2-S9 in Supplementary material). For example, Phillyrea latifolia exhibited highly flammable characteristics for leaf dry matter content and leaf moisture content (Fig. 2, Figs. S2-S3) but had lower flammability based on its leaf curliness (Fig. 2, Fig. S5). Moreover, in some species, multiple traits contributed significantly to their overall flammability. For instance, Quercus coccifera displayed high or moderate-high flammable characteristics in three leaf traits and branching architecture, while Arbutus andrachne ranked at the more flammable end of the trait continuum for four leaf traits (Fig. 2, Fig. S2-S6). Similarly, Asparagus aphyllus and Astragalus sp. demonstrated very high flammability in all four traits measured.
Fig. 2 - Mean values of nine flammability traits for each species and their overall flammability scores (FS). The flammability level of each species and trait is represented by different colors: red (high), orange (moderate-high), yellow (moderate), light green (moderate-low), and dark green (low). Species are sorted by their overall flammability scores. Detailed graphs for each trait are provided in Figs. S2-S9. Trait names are listed in Tab. 2, species codes are provided in Tab. 1, and trait units are given in Tab. 3.
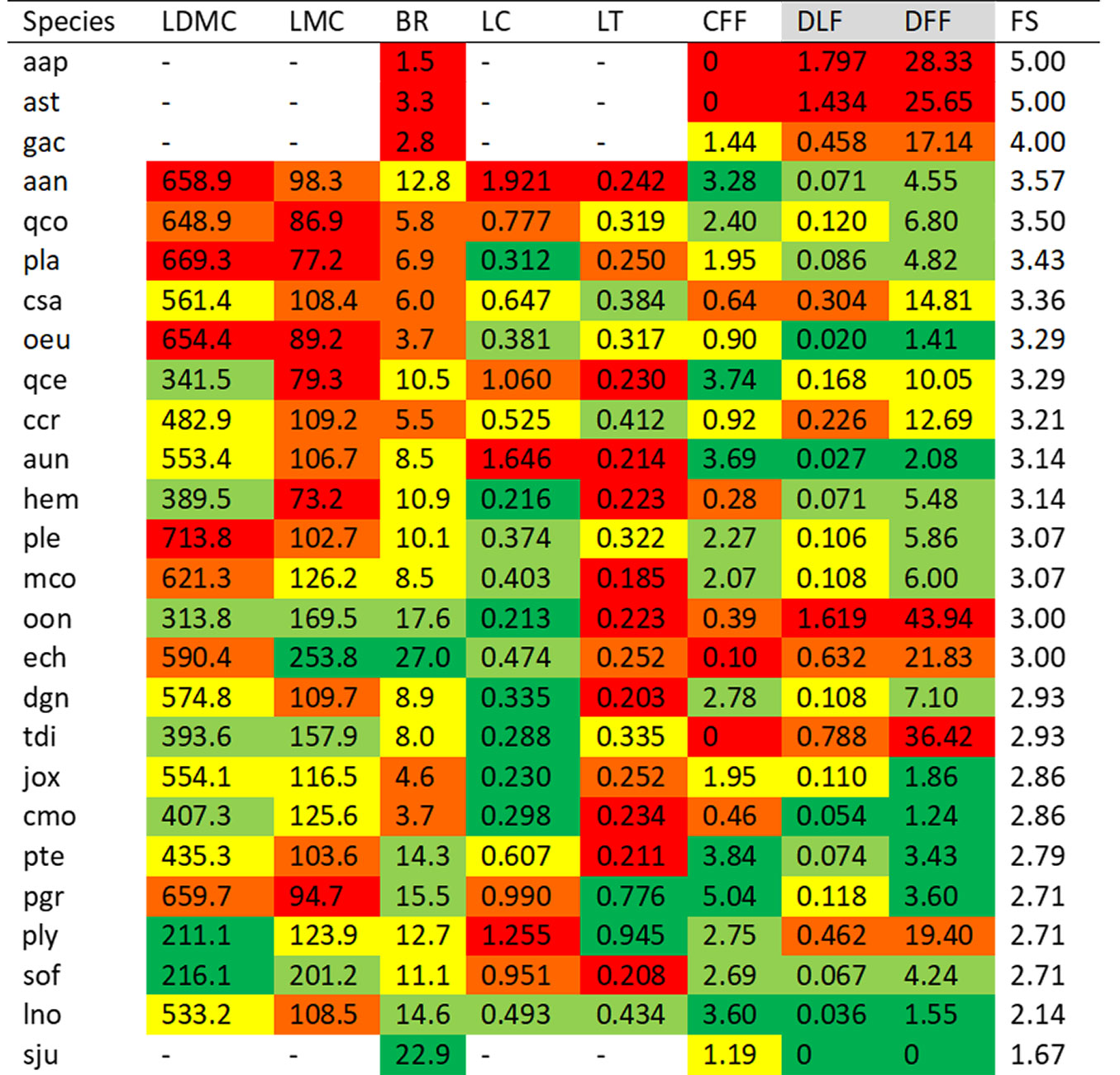
In contrast, some species (Laurus nobilis, Styrax officinalis, Spartium junceum, and Phlomis spp.) were at the less flammable end of trait continua for several traits (Figs. S2-S9 in Supplementary material). Even though these species had relatively lower overall flammability scores (Fig. 2), they exhibited flammable characteristics in a few specific traits. For instance, the flammability of L. nobilis was higher due to its leaf moisture content, while the leaf thickness and curliness of S. officinalis contributed to its flammability. Similarly, both Phlomis lycia and P. grandiflora exhibited higher flammability when leaf curliness was considered alone. Additionally, a few other traits uniquely contributed to the flammability of each of these species.
These results indicated that all but a few of the studied species exhibited a flammable character for at least one trait included in the study. Since traits contributing to higher flammability were species-specific, the ranking of species by flammability value varied significantly across traits (Figs. S2-S9 in Supplementary material). Consequently, interspecific variation in flammability persisted when considering the overall flammability score (FS) derived from the eight measured flammability traits (Fig. 2). In this sense, species could be ranked by their overall flammability, with Asparagus aphyllus, Astragalus sp., and Genista acanthoclada achieving the highest flammability scores (FS = 5.00, 5.00, and 4.00, respectively), while Spartium junceum presented the lowest score (FS = 1.67). Other notable species include Quercus coccifera (FS = 3.50) and Arbutus andrachne (FS = 3.57), which had relatively high scores, and Laurus nobilis (FS = 2.14), which had a lower score. The flammability score of other species ranged from 3.43 to 2.71 (Fig. 2).
Discussion
Our study demonstrated that the traits contributing to increased flammability in Mediterranean woody plants are idiosyncratic at the species level. Traits such as complex branching structure, retention of standing dead material, and curved, dense, or thin leaves, which enhance plant flammability, were found in various combinations across the studied species. These findings confirm our hypothesis that the Mediterranean woody species analyzed in the study area possess diverse traits contributing to higher flammability.
Our results revealed that several species exhibited very high flammability across multiple traits. Most of these species (Quercus coccifera, Arbutus spp., Cistus salviifolius, Phillyrea latifolia, and Genista acanthoclada, among others) are characteristic components of Mediterranean shrublands and also occur in the understory of low-altitude pine forests ([54]). Conversely, many species with relatively lower flammability scores (e.g., Phlomis grandiflora, Phlomis lycia, Styrax officinalis, and Laurus nobilis) could be considered more flammable when specific traits are evaluated in isolation. For instance, considering the negative relationship between leaf thickness and flammability ([24], [12], [6]), S. officinalis, C. monogyna, and P. terebinthus exhibited highly flammable characteristics due to their thinner leaves. Similarly, leaf curliness contributed to the overall flammability of P. lycia, P. grandiflora, and S. officinalis, even though these species had relatively lower flammability scores. L. nobilis, which had the lowest flammability score among species for which all eight traits were measured, could be mistakenly classified as flammable if only leaf moisture content were considered. We observed these inconsistencies across all the species examined, as each species could be considered highly flammable for certain traits but not for others. This finding suggests that Mediterranean plants in fire-dependent communities exhibit species-specific strategies for flammability, supporting the idea of alternative flammability syndromes that provide equivalent fitness under frequent fire regimes ([23]). Given the intra- and interspecific variability in flammability traits observed in our study and across different ecosystems ([26], [41], [23]), defining such flammability syndromes could improve our understanding of fire-related dynamics at regional or global scales. Jaureguiberry & Díaz ([14]) recently proposed that plant flammability may represent the third dimension of plant fire syndromes, alongside classical resprouting and seeding. This hypothesis is based on evidence that plant species in Mediterranean-type ecosystems are more flammable than those in many other ecosystems. Our results partially support this argument, as all species in our study, except Spartium junceum and Laurus nobilis, exhibited at least one highly flammable trait. Consequently, considering flammability as a component of plant fire syndromes may enhance our understanding of the drivers of trait variation in the plants from fire-prone ecosystems.
The species-specific nature of flammability traits makes species selection a critical stage in studies addressing flammability-related questions, especially when the number of species included is limited. In such cases, biased estimation of overall flammability can lead to erroneous interpretations of community-level patterns in flammability traits and vegetation flammability. Comparing and sorting plant species by flammability can be effectively achieved by incorporating multiple components of plant flammability ([50], [13]). The interspecific variability in flammability was evident in our study through the calculated flammability scores based on eight traits. Our findings on the flammability of woody species in southwestern Anatolia, derived from plant functional traits, do align with a recent study in a nearby region that grouped species based on leaf flammability using laboratory burnings ([13]). Specifically, for species common to both Güney et al. ([13]) and our study, Arbutus andrachne, Phillyrea latifolia, and Quercus coccifera were more flammable than Cistus creticus, Myrtus communis, and Pistacia terebinthus. In this context, our species sorting based on flammability scores, from most to least flammable, is robust, as it integrates multiple leaf traits, a branching trait, and three combustibility traits. Our results suggest that incorporating several components of flammability into studies involving a limited number of species can mitigate potential bias in estimating community flammability ([27]). This approach is also relevant for fire-protection or risk-reducing strategies in wildland-urban interfaces or firebreaks, where planting woody species with low flammability is a key objective. However, our study highlights the challenges of identifying fire-resistant (i.e., hard-to-burn) species in Mediterranean Basin fire-prone ecosystems. Consequently, the effort to plant “less flammable” species may be ineffective, primarily when species selection relies on studies comparing a few species and traits. Searching for less flammable species to construct fire-resistant vegetation belts near human settlements or to create less flammable forest patches is increasingly critical as climate change drives larger and more intense fires. For effective forest management, multiple traits should be considered when selecting less flammable species rather than relying on a single characteristic. For example, based on flammability scores in our study, Spartium junceum, Laurus nobilis, Styrax officinalis, and Phlomis spp. are suitable candidates for constructing a less-flammable green belt in forested areas. An integrative approach, such as calculating flammability scores based on multiple flammability traits as in our study, offers a robust framework for making these selections.
Our results are derived from a single region of the Mediterranean Basin and should, therefore, be interpreted cautiously by forest managers who are applying risk-reducing treatments with relatively less flammable species. While our selection of eight traits that capture different aspects of plant flammability likely reduced potential bias in the calculated flammability scores, we acknowledge that a different set of traits could have resulted in different species rankings based on flammability. Notably, there is no scientific consensus on which traits contribute most significantly to plant flammability, highlighting the need for further research to determine the relative importance of specific traits across various ecosystems. Additionally, although plant flammability can be predicted using functional traits, other dimensions of flammability exist at both the plant level (e.g., plant chemicals, litter flammability) and the vegetation level (e.g., meteorological conditions). More comprehensive studies that include a greater diversity of species and traits are urgently needed to identify low-flammability species for management purposes in wildland-urban interfaces and forested landscapes. Investigating interspecific variability in flammability traits within plant communities may also enhance our understanding of community assembly dynamics in fire-prone ecosystems. In this context, our study contributes to the understanding of the variability in flammability traits among Mediterranean Basin plants, particularly those from the studied region. It offers insights into why Mediterranean vegetation as a whole is highly flammable.
Acknowledgements
We thank D. Deniz Kazanci, Cansu Ülgen, Cihan Ü. Degirmenci, Cansu Özyapiciel, Anil Bahar, and Ü. Nihan Tavsanoglu for their assistance during fieldwork; Eray Aktepe, Pinar Kütküt, Nartjan Özden, and Z. Ladin Çosgun for their contributions to laboratory measurements; and Hasim Altinözlü for his support in identifying plant samples. We are also grateful to the anonymous reviewers for their constructive comments, which significantly improved the manuscript. This study formed part of the requirements for the Ph.D. dissertation of the first author, submitted to Hacettepe University, and was financially supported by the Hacettepe University Scientific Research Unit (project no: FHD-2017-14106).
Data availability statement
The raw data used and analyzed in this study are available in table and graph formats in the Supplementary material.
Conflicts of interest statement
The authors declare no conflicts of interest.
Author contribution
Nursema Aktepe: Conceptualization (supporting); data curation (lead); formal analysis (equal); investigation (lead); methodology (equal); resources (equal); software (equal); validation (equal); visualization (equal); writing - original draft (equal); writing - review and editing (equal). Çagatay Tavsanoglu: Conceptualization (lead); data curation (supporting); formal analysis (equal); funding acquisition (lead); investigation (supporting); methodology (equal); project administration (lead); resources (equal); software (equal); supervision (lead); validation (equal); visualization (equal); writing - original draft (equal); writing - review and editing (equal).
References
CrossRef | Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Institute of Science, Hacettepe University, Beytepe 06800, Ankara (Türkiye)
Division of Ecology, Department of Biology, Hacettepe University, Beytepe 06800, Ankara (Türkiye)
Corresponding author
Paper Info
Citation
Aktepe N, Tavsanoglu Ç (2025). Multiple ways for enhancing flammability in Mediterranean woody species: a trait-based approach. iForest 18: 301-308. - doi: 10.3832/ifor4601-018
Academic Editor
Lorenzo Mw Rossi
Paper history
Received: Mar 13, 2024
Accepted: Jun 05, 2025
First online: Oct 20, 2025
Publication Date: Oct 31, 2025
Publication Time: 4.57 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2025
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 4861
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 2108
Abstract Page Views: 1437
PDF Downloads: 1195
Citation/Reference Downloads: 1
XML Downloads: 120
Web Metrics
Days since publication: 279
Overall contacts: 4861
Avg. contacts per week: 121.96
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
(No citations were found up to date. Please come back later)
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Calibration assessment of forest flammability potential in Italy
vol. 7, pp. 300-305 (online: 07 May 2014)
Research Articles
Assessing the performance of fire danger indexes in a Mediterranean area
vol. 11, pp. 563-571 (online: 01 September 2018)
Technical Reports
Comparison of fire danger indices in the Mediterranean for present day conditions
vol. 5, pp. 197-203 (online: 02 August 2012)
Research Articles
Post-fire recovery of the plant community in Pinus brutia forests: active vs. indirect restoration techniques after salvage logging
vol. 11, pp. 635-642 (online: 04 October 2018)
Research Articles
Responses of European forest ecosystems to 21st century climate: assessing changes in interannual variability and fire intensity
vol. 4, pp. 82-99 (online: 05 April 2011)
Research Articles
Relevance of terpenoids on flammability of Mediterranean species: an experimental approach at a low radiant heat flux
vol. 10, pp. 766-775 (online: 02 September 2017)
Research Articles
Contribution of environmental variability and ecosystem functional changes to interannual variability of carbon and water fluxes in a subtropical coniferous plantation
vol. 9, pp. 452-460 (online: 25 January 2016)
Research Articles
Fire occurrence zoning from local to global scale in the European Mediterranean basin: implications for multi-scale fire management and policy
vol. 9, pp. 195-204 (online: 12 November 2015)
Review Papers
Monitoring the effects of air pollution on forest condition in Europe: is crown defoliation an adequate indicator?
vol. 3, pp. 86-88 (online: 15 July 2010)
Technical Reports
Air pollution regulations in Turkey and harmonization with the EU legislation
vol. 4, pp. 181-185 (online: 11 August 2011)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







