
Contrasting resin-tapped and untapped Pinus pinaster Ait. trees of central Spain in a dendroclimatic research
iForest - Biogeosciences and Forestry, Volume 17, Issue 3, Pages 148-155 (2024)
doi: https://doi.org/10.3832/ifor4458-017
Published: May 30, 2024 - Copyright © 2024 SISEF
Research Articles
Abstract
We investigated whether there are significant differences in the climatic response of resin-tapped and untapped maritime pine (Pinus pinaster Ait.) trees in four forest stands in the central mountains of Spain, where the species is of natural origin. The stands are located in different provenance regions, with the westernmost site located on the northern slopes of Sierra de Gredos and the easternmost in the transition to the middle Duero basin, with altitudes ranging from 900 to 1350 m a.s.l. Dominant trees were sampled and standard dendrochronological methods were used to compare tree growth of both types of trees at each site. Climate-tree growth relationships were assessed using bootstrap correlations and response functions between tree-ring growth indices and climate variables, for both partials and full local chronologies of each stand, selecting climatic data from the most complete and closest meteorological stations. Our results showed that the trees selected for resin extraction were older, on average, and when site characteristics were favorable enough for maritime pine growth and resin extraction, all trees were resin-tapped. Contrastingly, when site characteristics were unfavorable for resin extraction, only the largest and the best-developed trees were tapped. No major differences were found between the climatic responses of resin-tapped and untapped trees, and it was therefore possible to use all the available tree-ring width series per site, both resin-tapped and untapped trees, to identify the main climatic drivers of maritime pine growth in central Spain. In addition, we have expanded the chronologies network of Pinus pinaster in Spain in number and length. This work may provide valuable information to improve forest management strategies for sustainable resin production.
Keywords
Resin Extraction, Tree-ring Width, Long-term Chronologies, Climate Response
Introduction
Pinus pinaster Ait. (Maritime pine) is the pine species that covers the larger extension in Spain due to its wide ecological range and long history of anthropic management. Palaeobotanical data describe the maritime pine as native to the Iberian Peninsula and proved the importance of its forests in the Iberian ecosystems for thousands of years ([12], [2], [36], [28]). Specifically, in the Central System mountain range of Spain, an early Holocene replacement of boreal pines (P. sylvestris) by Mediterranean pines (P. pinaster) has been documented in the mid-altitude areas of the NW region, a replacement that was most likely widespread in inland Spain ([26]). Other data confirm the establishment of maritime pine and monospecific P. pinaster forests on the southern slopes of the Sierra de Gredos, from around 400 cal yrs BP ([33]). Finally, in the Guadarrama mountain range, the main eastern section of the Central System, it is worth mentioning the presence of maritime pine pollen records for the last 1689 years in a periglacial hygroturbous deposit located in the Peñalara massif ([21]). P. pinaster has also been artificially extended in many areas of the Iberian Peninsula and other areas with Mediterranean and temperate climates worldwide ([3], [1]).
The natural and artificial populations of maritime pine have traditionally been exploited for the extraction of resin, one of the most important non-timber resources of the Spanish forests, especially from the 1840s to the 1980s. Spain and other European countries virtually ceased to be resin producers and exporters in the 1980s when cheaper Chinese production exceeded European production ([30], [43]). However, in recent years, the increased demand for natural resins and a decline in Chinese exportation have encouraged the resumption of resin extraction from the most productive Spanish forests ([43]).
It has long been known that resin tapping damages the trees and that the severity of the effects depends on the frequency of tapping and the type of wounds produced on the trunk. However, few studies have looked at the long-term effects on tree growth. Schweingruber ([42]) noted that some studies carried out between 1938 and 1992 showed that tree growth was reduced by up to 25% due to extreme and recurrent injuries. In the current century, very few studies have analyzed the effects of resin extraction on tree growth and vigor over time in Europe and China ([29], [25], [20], [13], [44], [46]).
Most maritime pine forests in Spain are quite young and trees older than 100 years are rare, due to traditional uses and intensive management for resin extraction. In the network of P. pinaster chronologies analyzed by Sánchez-Salguero et al. ([38]), 42% of the chronologies start before 1900, but only 20% before 1850. At least in central and southern Spain, it is difficult to find long-lived P. pinaster trees that have not been tapped ([20], [9]).
Accurate knowledge of the response of trees to climate change in the context of current global change is an increasingly pressing issue when planning adaptation and mitigation measures in forests. In this sense, the maritime pine is one of the most studied forest tree species in relation to the effects of increasing droughts in Spain in recent years. Numerous dendroecological studies have been carried out to analyze the diverse responses in regions with contrasting climates ([7], [15], [35], [8], [4], [38]). In addition, the decline of pine trees due to drought and other stressors has also been analyzed ([40], [17]).
We have posed our work with this in mind, especially because we think that the dendroecological studies on resin and non-resin trees may provide valuable information about tree performance to better adapt forest management strategies to sustainable resin production. For this reason, we focus on the long-term radial growth of maritime pine and its response to climate considering resin-tapped (RT) and resin-untapped trees (RU) in four native forest stands across the central mountain range of Spain, where the resin was extracted until the 1980s. Specifically, we focused on assessing whether there are significant differences in the climatic response between RT and RU trees. If the results do not provide significant evidence of different responses of both types of trees to climate change, then RT and RU trees could be used together in dendroclimatic studies. Moreover, we aim to provide relevant information to increase the knowledge about the growth-climate relationship of the maritime pine for the central region of the Iberian Peninsula and to compare it with other Iberian regions.
In summary, we try to answer the following questions: (i) which trees were selected for resin extraction in central Spain? (ii) Does resin extraction trigger lower growth in trees? (iii) Are there relevant disparities in the response to climate between RT/RU trees? (iv) What are the main climatic drivers for the P. pinaster growth in central Spain?
Furthermore, these results will be a valuable contribution to increase the current network of P. pinaster chronologies in Spain ([38]), and in particular to expand the scarce long-term chronologies of this species that have been built to date.
Material and methods
Study area
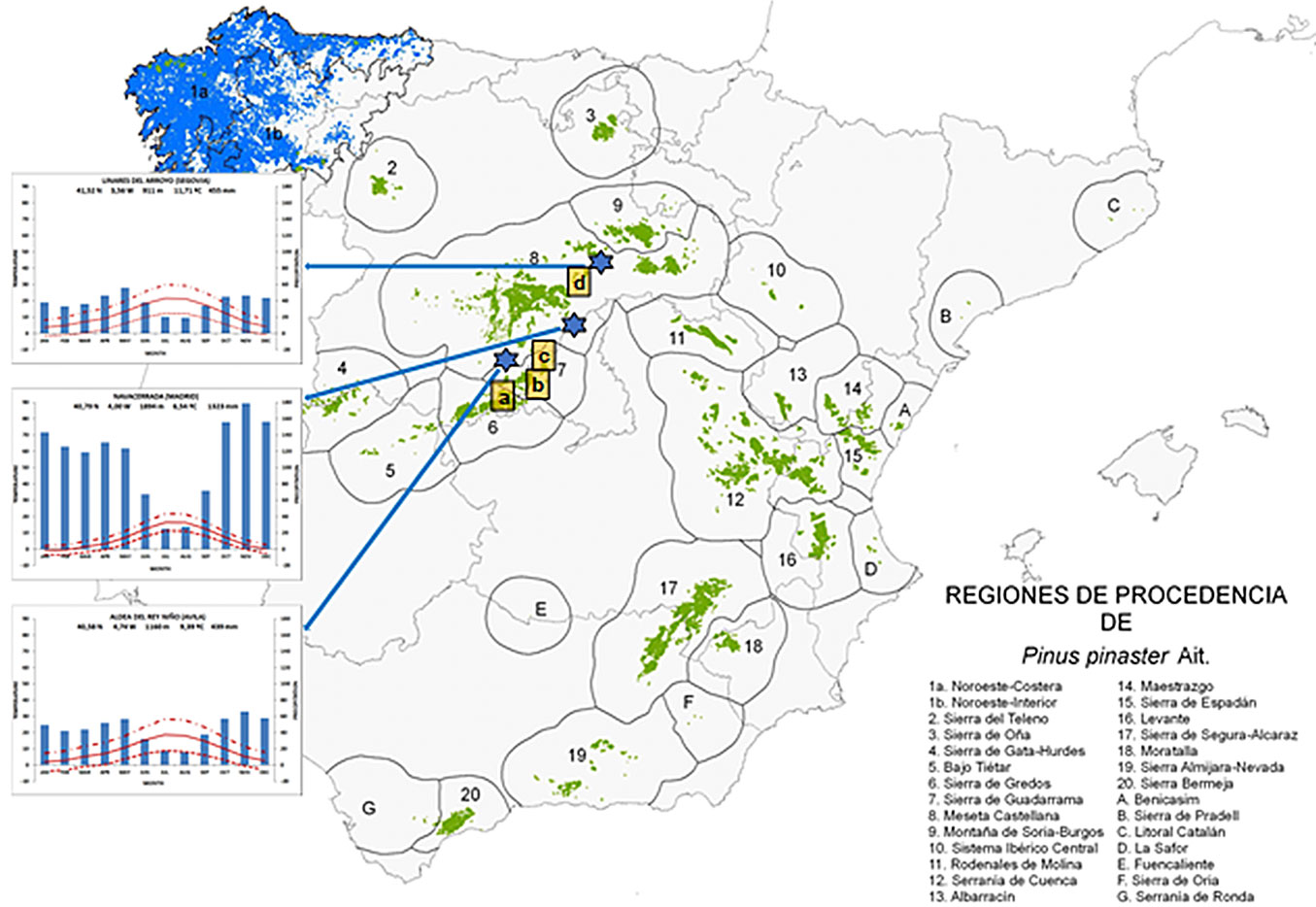
We selected four forest stands in the central mountains of Spain, where Pinus pinaster was of natural origin and the dominant tree species (Tab. 1, Fig. 1). From west to east, the sites sampled were Venero (VEN, Navaluenga, Ávila), Almenara (ALM, Robledo de Chavela, Madrid), Jarosa (JAR, Guadarrama, Madrid) and Hornuez (HOR, Moral de Hornuez, Segovia) that are located in distinct provenance regions according to Alía et al. ([3]), ranging in altitude from 900 (VEN) to about 1350 m a.s.l. (JAR).
Tab. 1 - Location and characteristics of sampling sites and the number of sampled trees. (Scc): Sandstones, conglomerates, and clays; (RT): resin-tapped trees; (RU): resin-untapped trees.
| Site | Latitude (N) |
Longitude (W) |
Mean altitude (m a.s.l.) |
Substrate | Aspect | No. RT trees |
No. RU trees |
|---|---|---|---|---|---|---|---|
| VEN | 40.39 | -4.64 | 1030 | Granite | NW | 15 | 22 |
| ALM | 40.45 | -4.26 | 1050 | Granite | W | 39 | 12 |
| JAR | 40.68 | -4.13 | 1230 | Gneiss | SW | 23 | 12 |
| HOR | 41.48 | -3.65 | 1200 | Scc | NE | 24 | 12 |
Fig. 1 - Sampled forest stands in the center of the Iberian Peninsula (indicated by a lowercase letter), meteorological stations location (indicated by stars) and their corresponding climatic diagrams are located in the map of provenance regions (“Regiones de Procedencia”) of Pinus pinaster ([3]), where the areas marked in blue correspond to repopulations and those marked in green correspond to native forest. Sites sampled: (a) Venero, (b) Almenara, (c) Jarosa, (d) Hornuez.
The Venero site is located in the river basin of the Arroyo Cabrera torrent, formed by the confluence of several secondary torrents descending from the Escusa summit (1960 m a.s.l.), on the northern slopes of Sierra del Valle (Gredos Mountain Range, Spanish Central System). Common granitic rocks covered by superficial quaternary formations of conglomerates, gravels, sands, and silts characterize the landscape. The forest stand is formed predominantly by Pinus pinaster, with some individuals of P. sylvestris and Quercus pyrenaica (Fig. 2d). In addition, riparian broadleaved species (Alnus glutinosa, Fraxinus angustifolia, Populus nigra) colonize both riverbanks ([5]).
Fig. 2 - (a) Resin extraction scars and callus formed to isolate damaged tissues in a cross-section from Hornuez; (b) scar caused by resin extraction several decades after in a live tree; (c) overview of Almenara, the steepest and stoniest site; (d) the pine forest of Venero; (e) the pine forest of Almenara with dead trees.

The Almenara site is located between the Gredos and Guadarrama ranges, forming part of a series of minor alignments of isolated hills that link both mountain ranges. The soil parent material is mostly granite and gneiss, dominating the former, and reaches a maximum altitude of 1259 m a.s.l. On the east side, Quercus ilex dominates the forest with Juniperus oxycedrus, Pistacia terebinthus, and Retama sphaerocarpa. On the western slope which is steeper and stony, Q. ilex locally shares dominance with Pinus pinea, in low and more southern areas, and with P. pinaster in the middle and higher areas, where it becomes more abundant (Fig. 2c). In Almenara we identified many dead trees in recent years (Fig. 2e), as it occurs in other areas of central Iberia ([17]).
The site called Jarosa is located in a valley in the southwest of the Sierra de Guadarrama, specifically on its southeast slope. The highest peak is Cabeza Líjar (1824 m) and, unlike the previous sites, the gneiss dominates over granite. The pine forest is the characteristic vegetation of the valley, P. pinaster forest in the lower areas is accompanied by Cistus ladanifer and Cytisus scoparius, which is replaced by P. sylvestris, as the altitude increases. Small pine forests and relict stands of P. nigra also survive on the highest and central sectors of the valley, between 1200 and 1600 m a.s.l. ([31]), some of them being more than 500-year-old ([18]).
The most northeastern site, Hornuez, is located in the mid-Duero basin at the transition with the Central System Range. The main substrates include Triassic sandstones, conglomerates, and clays, with very well-drained soils. The main tree species in the studied forest was P. pinaster, including also several isolated Quercus faginea and Q. ilex. In the summer of 2008, a big fire killed all the trees in the Hornuez forest stand (Fig. 2a). We sampled the dead trees and collected cross sections that were analyzed together with the cores previously sampled ([20]). In this work, we used the revised dedrochronological data to compare them with the other three sampled sites.
Resin extraction, especially from Maritime pines, was a common human activity throughout the 20th century that completely stopped in Spain in the 80s, although it has recently been restarted with new techniques ([30], [43]). With the exception of the Hornuez site, there are no precise dates of when the trees were resin-tapped. We used the disks collected in Hornuez to date the scars of the resin extractions, which occurred mainly during the period 1920-60 ([20] - Fig. 2a).
In each stand, we extracted cores from dominant and large trees, both resin-tapped (RT) and resin-untapped (RU). Resin-tapped trees that were usually sampled showed traces of scars in the trunk (Fig. 2b). At least two cores per tree were extracted at a height of 1.30 m from the base of the trunk and as far away from the resin scars as possible to avoid the influence of the overgrowth of the callus formation, which isolates the wounds and limits the spread of pathogens ([41]). For the Hornuez site, we added cross-sections of samples (13 RT trees and 6 RU trees) cut at a height of approximately 15 cm above the tree base ([20]).
Dendrochronological methods
Tree samples were prepared and cross-dated following standard dendrochronological methods. Tree-ring measurements were carried out using a Lintab device (accuracy 1/100 mm) and analyzed using TSAP software ([32]). For the cross sections from Hornuez, at least four radii per sample were dated and tree rings were measured ([20]). Cross-dating was checked using the software COFECHA ([22]).
We evaluated and compared the dated tree-ring growth series of both types of trees (RT and RU) at each site, selecting the same number of trees (12 trees) from the pool of sampled trees to give equal weight to each type of tree per site. As the RU trees are generally younger, those with similar tree-ring series lengths covering the whole trunk radius from the bark to the pith were selected. To compare RT and RU tree-growth, we analyzed the raw mean TRW (Tree-Ring Width) series and the growth change over time in the common period of the RT and RU chronologies (1929-2008).
Climatic data and climate-tree growth relationships
The general climate of the area is continental Mediterranean, typical of the mountain ranges of inland Spain. Summers are hot and dry, spring and autumn are temperate and humid, winters are cold and humid, and the rainfall is irregular, among and within years.
We selected climatic data from Aldea del Rey Niño (Ávila), Navacerrada (Madrid), and Linares del Arroyo (Segovia) meteorological stations, due to their proximity to the sampling areas and the wide and complete range of data they provide (Fig. 1). In addition, they capture a wide range of climates in central Iberia, from low (911 m a.s.l.) to high altitudes (1894 m a.s.l.), with the lower stations representing a warmer and drier climate. Most of the records began in the mid-1940s, with the exception of the Aldea temperature, which began in the early 1960s (Tab. 2).
Tab. 2 - Characteristics of the meteorological records used. (T): temperature; (P): precipitation; (MAT): mean annual temperature; (MAP): mean annual precipitation.
| Meteorological station | Province | Latitude (N) |
Longitude (W) |
Altitude (m a.s.l.) |
1st-year recorded T/P |
MAT (°C) |
MAP (mm) |
|---|---|---|---|---|---|---|---|
| Aldea del Rey Niño | Ávila | 40.58 | -4.74 | 1160 | 1945/1945 | 9.39 | 439 |
| Navacerrada | Madrid | 40.79 | -4.00 | 1894 | 1946/1946 | 6.54 | 1323 |
| Linares del Arroyo | Segovia | 41.52 | -3.56 | 911 | 1962/1943 | 11.71 | 455 |
In the records of Aldea del Rey Niño and Navacerrada there was a period when the temperatures were below the average (around 1962-1988) and a clear trend of temperature increase from 1970 onwards, whereas this trend is not so clear in the records of Linares del Arroyo (Fig. S1 in Supplementary material). Precipitation records from all the meteorological stations show high interannual variability and no trend, although the period 1990-1995 can be highlighted as dry, coinciding with years of generalized drought throughout Spain.
Tree-ring width series were standardized to establish the chronologies of indices, using the software ARSTAN ([14]). Climate-tree growth relationships were assessed using bootstrap correlations and response functions between tree-ring growth indices and climate variables, both for the partials and the entire local chronologies of each stand, using the program DendroClim2002 ([6]). Correlations were calculated from the previous August to the current October i.e., some months prior and during the year of tree-ring formation, based on previous analyses of growth responses of central Iberian maritime pine to climate ([20]).
Results
Tree-age and tree-ring width rate of RT and RU trees
The biometric characteristics and estimated age of the selected trees (12 trees of each type at each site) to explain and evaluate the differences in tree growth between resin-tapped (RT) and resin-untapped (RU) trees are shown in Tab. 3.
Tab. 3 - Characteristics of the tree-ring width series and the residual chronologies of the RT and RU trees. (RT): resin-tapped trees; (RU): resin-untapped trees; (MTRW): Mean Tree-Ring Width (mm); (IT): series intercorrelation; (MS): mean sensitivity; (EPS): Expressed Population Signal; (%varPC1): common variance explained by the first principal component.
| Site | Tree type |
Mean Age (range) |
MTRW (±SD) |
MTRW 1929-2008 (±SD) |
IT | MS | Time span | EPS>0.85 | %varPC1 |
|---|---|---|---|---|---|---|---|---|---|
| VEN | RT | 108 (72-129) | 3.18 ± 1.7 | 2.50 ± 0.69 | 0.60 | 0.23 | 1881-2009 | 1930-2007 | 54.14 |
| RU | 65 (56-81) | 3.71 ± 1.8 | 3.65 ± 1.17 | 0.60 | 0.23 | 1929-2009 | 1950-2007 | 52.93 | |
| ALM | RT | 107 (76-130) | 2.87 ± 2.0 | 2.50 ± 1.07 | 0.65 | 0.30 | 1885-2014 | 1920-2013 | 63.11 |
| RU | 108 (85-146) | 2.47 ± 1.6 | 2.23 ± 0.96 | 0.56 | 0.30 | 1869-2014 | 1920-2013 | 58.82 | |
| JAR | RT | 107 (70-133) | 1.99 ± 1.1 | 1.79 ± 0.46 | 0.53 | 0.25 | 1878-2010 | 1920-2010 | 48.06 |
| RU | 74 (56-96) | 3.89 ± 1.0 | 4.10 ± 1.36 | 0.60 | 0.24 | 1915-2010 | 1950-2010 | 50.08 | |
| HOR | RT | 94 (71-112) | 2.46 ± 1.6 | 2.09 ± 1.07 | 0.64 | 0.23 | 1897-2008 | 1910-2007 | 58.77 |
| RU | 91 (54-120) | 2.75 ± 1.4 | 2.77 ± 1.00 | 0.65 | 0.23 | 1889-2008 | 1930-2007 | 49.93 |
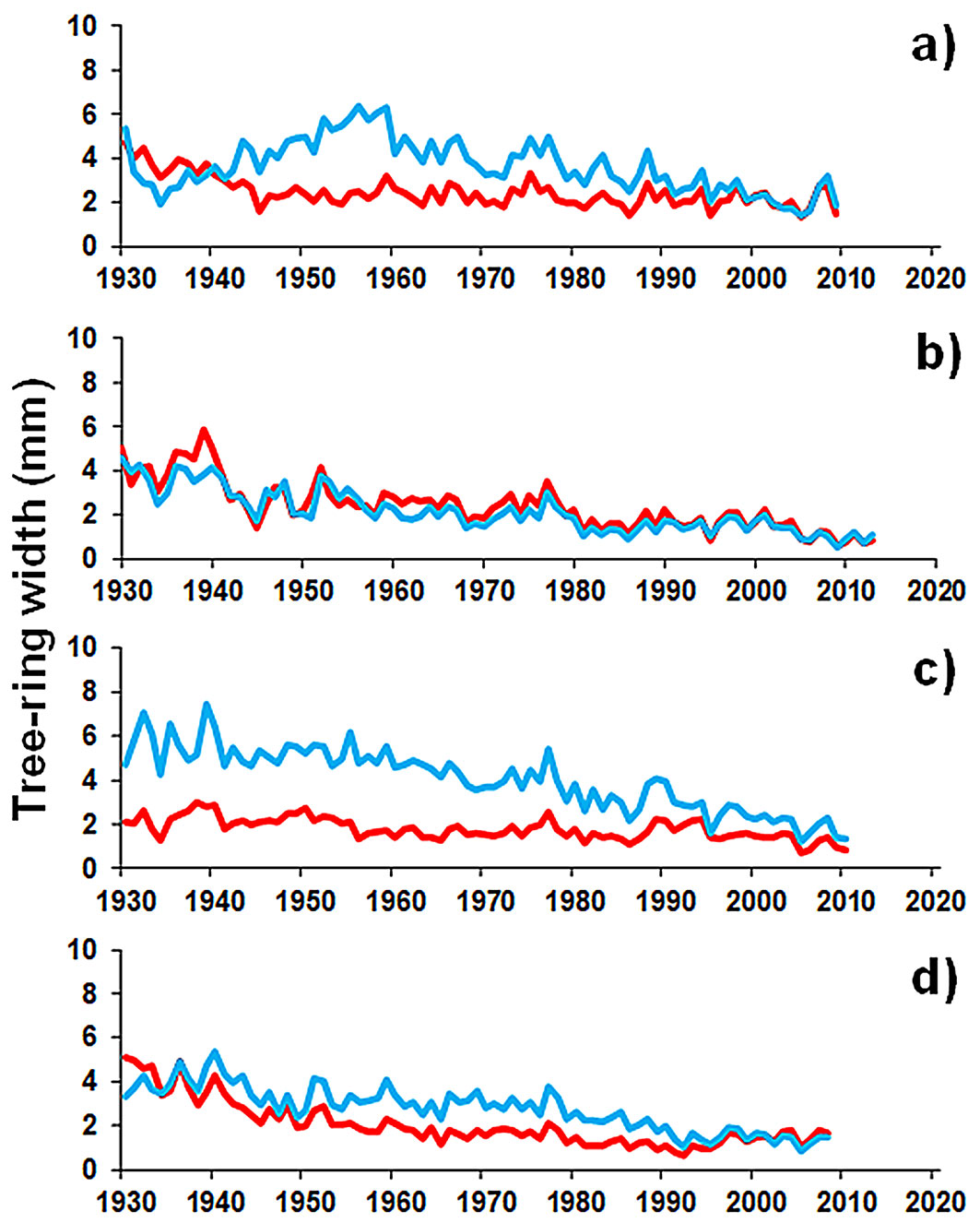
Mean tree-age ranged from 65 to 108 years and mean annual ring width ranged from 1.99 to 3.71 mm. RU trees were younger on average than RT trees, especially RU trees of VEN and JAR sites. Moreover, our results showed that the growth of RU trees was higher than that of RT trees in almost all sites, both in the full period and in the common period (Tab. 3, Fig. 3), with percentages ranging from 25%-56% in the common period. Analyzing the growth in this common period (1930-2008) divided into four 20-year segments, the growth of RU trees compared to RT is significantly higher (p<0.05) for all periods in the JAR site, in the VEN site except for 1930-1949, for the two intermediate segments (1950-1989) in the HOR site and only for the 1970-1989 segment in the ALM site.
Fig. 3 - Mean tree-ring width series of resin-tapped trees (red line) and resin-untapped trees (blue line) of the four sites for the common period. (a) VEN site; (b) ALM site; (c) JAR site; (d) HOR site.
Climate response of RU and RT trees
Cross-dating results (IT) show that the series intercorrelation is higher than 0.50, reaching a maximum of 0.65 in the HOR and ALM sites; the mean sensitivity ranged between 0.23 and 0.30 and the maximum was reached in the ALM-site (Tab. 3). We decided, therefore, to develop partial residual chronologies with the tree-ring series of RU and RT trees in each site. Most of these residual chronologies were reasonably representative from 1910 to 1930, although the common period in which EPS (Expressed Population Signal) > 0.85 extended from 1950-2007 (Tab. 3). Besides, the variance explained by the first principal component exceeded 50% in almost all chronologies (Tab. 3). Thus, we analyze the climate correlations of both sets of chronologies (RU and RT trees) and the average with respect to the meteorological record closer to each site, checking that they were almost indistinguishable (Fig. S2 in Supplementary material).
The local chronologies and the responses of Pinus pinaster to the climate
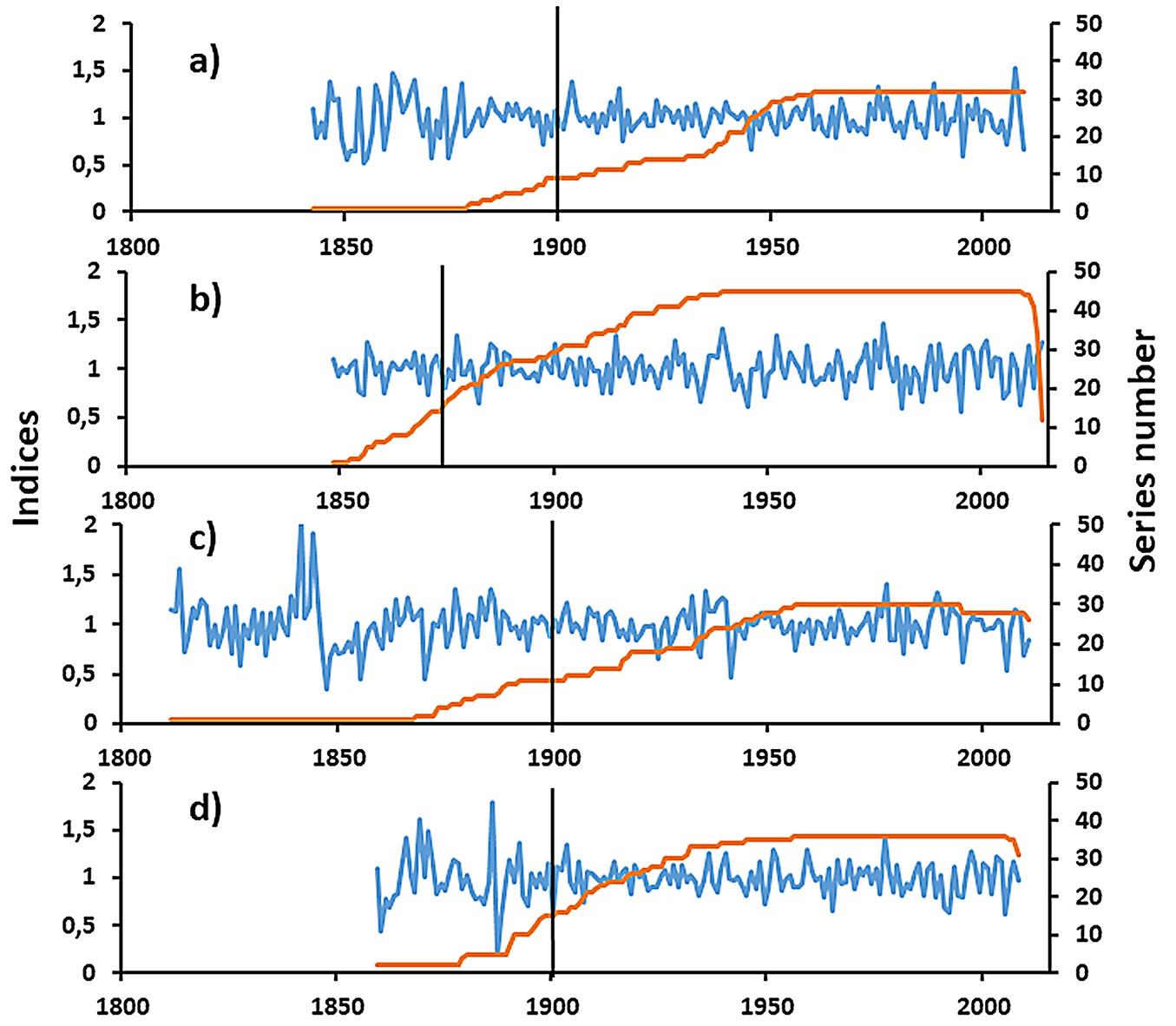
Because we found that resin extraction did not alter the overall short-wavelength variability of the tree-ring series and their climate response, we developed four local chronologies with all series dated and synchronized, one from each site. All of these sites show similar values of mean tree-ring width, intercorrelation, and mean sensitivity, although the VEN site had the maximum mean tree-ring-width, the JAR site had the lower intercorrelation, and the ALM site had the maximum mean sensitivity (Tab. 4). The chronologies extend up to a maximum of 200 years, with the HOR chronology showing the maximum intercorrelation and the variance explained by the first principal component of more than 62%. The agreement of the variance of the sample chronologies with that of the theoretical population chronology of more than 0.85 began in 1880, and it was common from 1900 onwards (Tab. 4, Fig. 4). In addition, the correlations of the 50-year dated segments were above the critical correlation below 0.32 in almost in all cases (Tab. 5). Lastly, five negative pointer years common since 1900 were identified: 1981, 1986, 1995, 2005, and 2009, all of them corresponding to the last four decades.
Tab. 4 - Characteristics of tree-ring width series and residual chronologies from each site. (MaxAge): maximum age; (MTRW): Mean Tree-Ring Width; (SD): standard deviation; (IT): intercorrelation; (MS): mean sensitivity; (EPS): Expressed Population Signal; (varPC1): variance explained by the first principal component.
| Site | N trees |
MaxAge | MTRW ± SD (mm) |
IT | MS | Time span | EPS>0.85 | %varPC1 |
|---|---|---|---|---|---|---|---|---|
| VEN | 32 | 170 | 3.50 ± 1.86 | 0.61 | 0.24 | 1842-2009 | 1900-2007 | 47.62 |
| ALM | 45 | 168 | 2.45 ± 1.66 | 0.60 | 0.28 | 1848-2014 | 1880-2012 | 44.82 |
| JAR | 30 | 201 | 2.40 ± 1.37 | 0.55 | 0.25 | 1811-2010 | 1900-2010 | 40.77 |
| HOR | 36 | 154 | 2.50 ± 1.50 | 0.66 | 0.25 | 1859-2008 | 1900-2008 | 62.18 |
Fig. 4 - Residual chronologies of the studied sites. a) VEN-site. b) ALM-site. c) JAR-site. d) HOR-site. The vertical bar indicates the year from which EPS>0.85.
Tab. 5 - Correlations of 50-year dated segments, lagged 25 years, between the residual local chronologies. (*): p < 0.01.
| Site | Time span | ||||||
|---|---|---|---|---|---|---|---|
| 1825-1874 | 1850-1899 | 1875-1924 | 1900-1949 | 1925-1974 | 1950-1999 | 1975-2024 | |
| VEN | 0.34 | 0.43 | 0.36 | 0.55* | 0.57* | 0.67* | 0.73* |
| ALM | 0.63* | 0.63* | 0.49* | 0.67* | 0.70* | 0.73* | 0.75* |
| JAR | 0.37 | 0.49* | 0.45 | 0.27 | 0.25 | 0.56* | 0.71* |
| HOR | - | 0.27 | 0.35 | 0.53* | 0.53* | 0.43 | 0.51* |
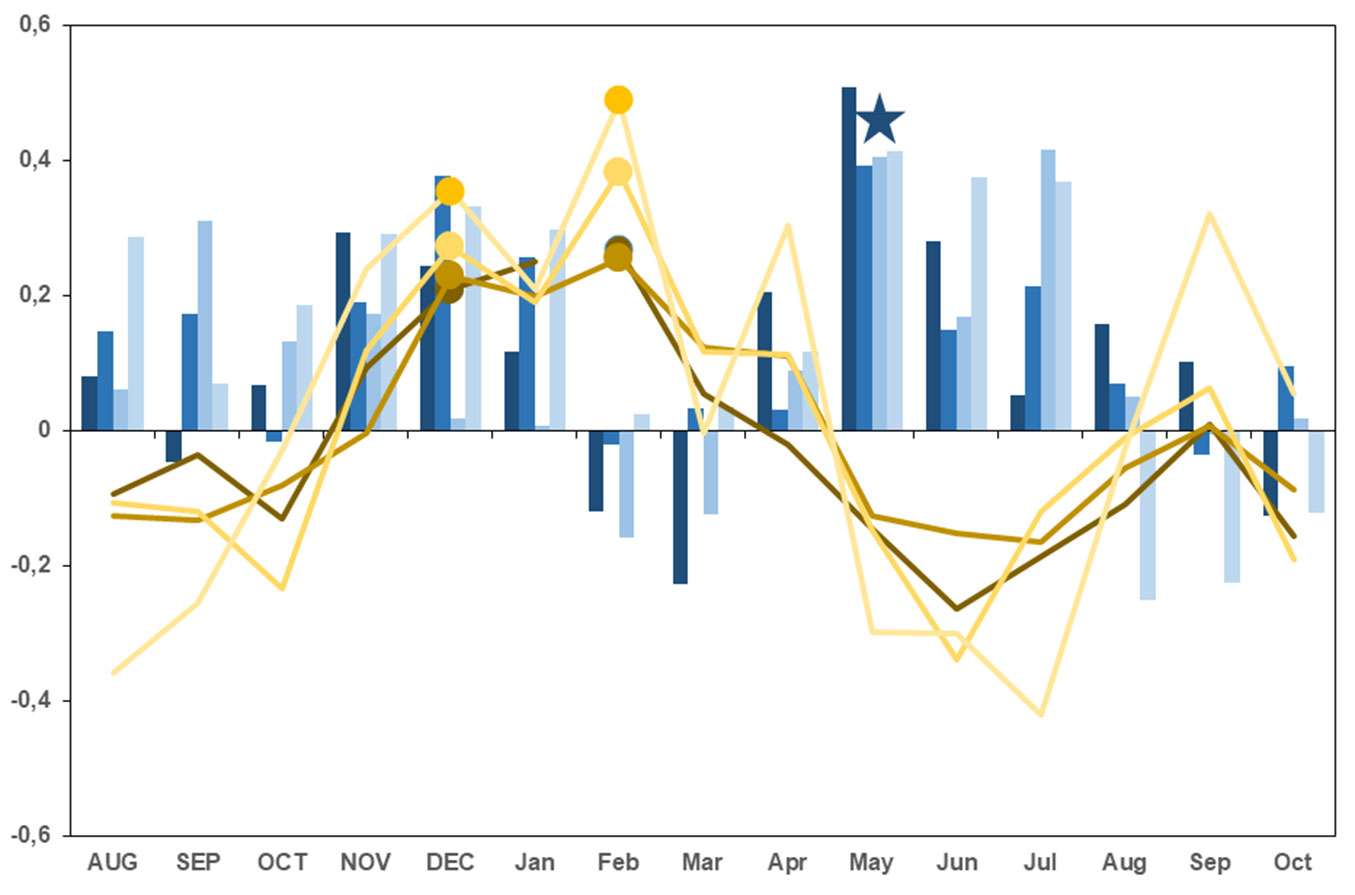
There were no significant differences between the responses of the four pine forests to the climate. The sign of the response function coefficients for the mean monthly temperatures varied depending on the seasons. It was mainly positive for the previous autumn and the current winter (mean February temperature was the most significant month) and negative for mid-spring and early summer (May-June-July). For the precipitation, the correlations were mostly positive. There was a highly significant relationship between the indices of the chronologies and precipitation for the previous autumn and the current winter (December was the most significant month) and, especially for the mid-spring and early summer (May-June-July) of the current year. Overall, the precipitation correlations were strongest and most significant in May (Fig. 5).
Fig. 5 - Bootstrapped tree-growth correlations to climate, estimated by ring-width indices and by monthly climatic variables (mean temperature - gold color lines - and precipitation - blue columns), for the studied sites. The window starts in August of the previous year (upper case) and ends in October of the current year (lower case). The color of the columns and lines becomes clear from W to E (VEN: Aldea del Rey Niño; ALM: Aldea del Rey Niño; JAR: Navacerrada; HOR: Linares del Arroyo). Significant values (p<0.05) coincident in all the response functions have been highlighted, the dots represent the temperature values and the star represents the precipitation values.
Discussion
Which trees were selected for resin extraction in central Spain?
To answer this question, we first tested whether the ages of the RT trees were significantly different from the ages of the RU-trees, verifying that the last were younger on average, though this does not fit in all the analyzed sites. We hypothesize that if site characteristics were favorable enough for both the development of maritime pine and the resin extraction (for example, ease of access), all the trees would be resin-tapped and, if not, only the largest and best-developed ones would be resin-tapped. For this reason, in the two most favorable sites (VEN and JAR), the RU-trees showed a lower mean age, probably because they were very young or small when the resin extraction started, or they established after the resin extraction started, whereas in the other two most unfavorable sites (ALM and HOR) the mean age of both tree groups was similar. The age and juvenile growth differences between RT and RU-trees will be of great interest to analyze the growth-rate of both tree types.
Does resin extraction trigger lower growth in trees?
Few studies have investigated the differences in growth caused by resin extraction. Previous studies on different species of Pinus (P. halepensis, P. pinaster, P. massoniana, P. tabuliformis) have found that pine growth is negatively affected by resin extraction ([29], [20], [46]), especially in old trees ([13]). In contrast, other studies in P. sylvestris determined a growth increase response to resin tapping, though these studies mainly analyzed the wood formation on the living part of the bole but not on the tapping face ([25], [44]). Therefore, these results are not exactly comparable (different species, ages, and types of samples). Furthermore, the cited studies were conducted in countries where different resin extraction methods were used, as summarized by Da Silva et al. ([16]).
In central Spain, the mean TRW-series showed that the growth of RU-trees was higher than that of RT-trees, with the exception of the ALM site, with variable percentages of growth divergence estimated between 56% and 25% depending on the sites. It should be noted that the TRW of the RT-trees generally remained lower for at least 50 years, although in recent decades this growth difference has decreased and disappeared, probably related to the end of resin extraction, which occurred throughout Spain in the 1980s ([30]). However, this growth difference persisted for more years in JAR, probably because resin extraction lasted longer at this site.
The site characteristics, that influenced the management of resin extraction, and the variability of tree-age, are the factors that are likely to explain the main divergences in growth-rate of both tree types among the sites studied. The largest differences were found in what is probably the most suitable environment for the maritime pine growth (the JAR site) and, conversely, the smallest differences were found in the steepest and stony site with poor and thin soils (the ALM site). In the latter site, resin extraction could have been difficulted because of the rugged orography and the long distance to the main transport routes, and trees with the poorest growth and appearance could have been excluded for resin tapping. That is, the variability in tree-age explained by tree selection for resin extraction, also plays a significant role in the interpretation of growth-rate divergences between RU and RT-trees, as already pointed out by Magnuszewski & Tomusiak ([25]) and Génova et al. ([20]).
Are there relevant disparities in the response to climate between RT/RU trees?
The potential impact of resin tapping on the growth-climate relationships has rarely been investigated ([20], [44], [46]). The first two studies conclude that there are no differences in the climate sensitivity of RT and RU trees, nor in the occurrence of growth responses to extreme events ([44]), but Zeng et al. ([46]) suggest that tapped trees are more vulnerable to drought. Our work, with extensive data on P. pinaster in central Spain, provides reliable evidence that there are almost no differences in the response to climate between these two groups of trees.
What are the main climatic drivers for the P. pinaster growth in central Spain?
The long-lived chronologies established in this study, spanning almost 200 years, extend and complete the current network of chronologies of Pinus pinaster from Spain ([38]). The great similarities found between the responses of maritime pine to the climate of central Spain in several sites with different ecological conditions, help outline in detail its main climatic drivers in the current context of global change. The main monthly climatic drivers are February temperature and May precipitation, albeit more broadly the tree response is focused on two periods: the previous autumn/current winter (from November to February) and the mid-spring/early summer (May-June-July).
The positive effect of warmer February may be due to the fact that it induces the earliest cambial activation and has also been detected in central Portugal ([10]) and central and southern Spain ([9], [17], [34]). On the other hand, lower precipitation in May has a noteworthy negative effect on P. pinaster growth, as was also determined in many regions of Spain ([7], [35], [11], [8], [9], [27], [38]) and Portugal ([45], [23]). High temperatures and precipitations during the previous autumn and the current winter enhanced tree-growth in the four stands studied, with the most crucial period being mid-spring/early summer of the current year (May-June-July). In mid-spring/early summer, the negative relationship with temperature converges with a positive relationship with precipitation, suggesting that the growth of P. pinaster in central Spain is mainly limited by water availability during the growing season, as postulated by Bogino & Bravo ([7]), Sánchez-Salguero et al. ([38]) and Gea-Izquierdo et al. ([17]).
Of all the identified negative pointer years, only 1995 and 2005 have been mentioned before, especially 2005, for example in Sánchez-Salguero et al. ([39]), Madrigal-González et al. ([24]), Rubio-Cuadrado et al. ([37]) or Gea-Izquierdo et al. ([17]). The year 1986 was also reported by Génova ([19]) as a pointer year for P. sylvestris and P. nigra on the Guadarrama mountains in central Spain. In the ALM and JAR sites, we add another pointer year, 2009, just as outstanding as the previous ones. We point out that these growth reductions in annual ring width seem to be recurring more and more in recent decades.
Conclusions
Four native Pinus pinaster forests in central Spain were used as indicators to assess the effects of resin tapping on tree-ring growth. Tree-ring growth in tapped trees was in general significantly lower than that in untapped trees, but this fact was related to several factors. Our results showed that when site characteristics were favorable enough for both maritime pine development and resin extraction, all trees were resin-tapped, otherwise only resin was extracted from the largest and the best-developed trees. No significant differences were found between the climatic response of resin-tapped and untapped trees, making it possible to use both tree-ring width series to identify the main climatic drivers for maritime pine growth in central Spain. By selecting native areas of Pinus pinaster in Spain, it was possible to establish longer chronologies than if populations from artificial plantations were studied. Dendroecological studies may provide valuable information to improve forest management strategies for sustainable resin production.
List of abbreviations
RT: resin-tapped, RU: resin-untapped, VEN: Venero site, ALM: Almenara site, JAR: Jarosa site, HOR: Hornuez site, IT: Intercorrelation, EPS: Expressed Population Signal, TRW: Tree-Ring Width.
Acknowledgments
We thank Miguel Salguero, Agentes Forestales of Comunidad de Madrid, and Agentes medioambientales of Comunidad de Castilla-León for their support during fieldwork; J. A. Ballesteros kindly gave data on untapped trees of the Venero site and Enrique Sadornil help us with some statistical analysis. This research was partially funded by the Senscom (CGL2008-06005), Dendro-Avenidas (CGL2007-62063), Dinecofor (CGL 2011-27229) and Dipave (CGL2011-27229) Research Projects.
Author contibutions
MG conceived the study, collaborated in the fieldwork and the methodological investigations, and drafted the original manuscript; LC carried out the fieldwork and performed the statistical analysis; EG collaborated in the methodological investigations. All the authors reviewed, and edited the manuscript.
References
Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Authors’ Info
Authors’ Affiliation
Leocadia Caminero 0009-0008-0897-1369
Departamento de Sistemas y Recursos Naturales, Universidad Politécnica de Madrid, 28040 Madrid (Spain)
Department of Evolutionary Biology, Ecology and Environmental Sciences, University of Barcelona, 08028 Barcelona (Spain)
Corresponding author
Paper Info
Citation
Génova M, Caminero L, Gutiérrez E (2024). Contrasting resin-tapped and untapped Pinus pinaster Ait. trees of central Spain in a dendroclimatic research. iForest 17: 148-155. - doi: 10.3832/ifor4458-017
Academic Editor
Jesus Julio Camarero
Paper history
Received: Aug 24, 2023
Accepted: Feb 09, 2024
First online: May 30, 2024
Publication Date: Jun 30, 2024
Publication Time: 3.70 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2024
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 15517
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 10374
Abstract Page Views: 2260
PDF Downloads: 2495
Citation/Reference Downloads: 3
XML Downloads: 385
Web Metrics
Days since publication: 757
Overall contacts: 15517
Avg. contacts per week: 143.49
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2024): 1
Average cites per year: 0.50
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
The effect of provenance of historical timber on tree-ring based temperature reconstructions in the Western Central Alps
vol. 13, pp. 351-359 (online: 25 August 2020)
Research Articles
Climatic fluctuations trigger false ring occurrence and radial-growth variation in teak (Tectona grandis L.f.)
vol. 9, pp. 286-293 (online: 28 September 2015)
Research Articles
The effects of fire on Pinus sylvestris L. as determined by dendroecological analysis (Sierra de Gredos, Spain)
vol. 15, pp. 171-178 (online: 09 May 2022)
Research Articles
Tree-ring-based reconstruction of larch budmoth outbreaks in the Central Italian Alps since 1774 CE
vol. 12, pp. 289-296 (online: 27 May 2019)
Research Articles
Role of photosynthesis and stomatal conductance on the long-term rising of intrinsic water use efficiency in dominant trees in three old-growth forests in Bosnia-Herzegovina and Montenegro
vol. 14, pp. 53-60 (online: 28 January 2021)
Research Articles
Individual-based approach as a useful tool to disentangle the relative importance of tree age, size and inter-tree competition in dendroclimatic studies
vol. 8, pp. 187-194 (online: 21 August 2014)
Research Articles
Response to climate and influence of ocean-atmosphere phenomena on annual radial increments of Pinus oocarpa Schiede ex Schltdl. & Cham in the Lagunas de Montebello National Park, Chiapas, Mexico
vol. 16, pp. 174-181 (online: 30 June 2023)
Research Articles
Growth-climate relations and the enhancement of drought signals in pedunculate oak (Quercus robur L.) tree-ring chronology in Eastern Hungary
vol. 11, pp. 267-274 (online: 29 March 2018)
Research Articles
Species-specific responses of wood growth to flooding and climate in floodplain forests in Central Germany
vol. 12, pp. 226-236 (online: 03 May 2019)
Research Articles
Earlywood vessel features in Quercus faginea: relationship between ring width and wood density at two sites in Portugal
vol. 8, pp. 866-873 (online: 27 April 2015)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







