
Linking nursery nutritional status and water availability post-planting under intense summer drought: the case of a South American Mediterranean tree species
iForest - Biogeosciences and Forestry, Volume 9, Issue 5, Pages 758-765 (2016)
doi: https://doi.org/10.3832/ifor1905-009
Published: Jun 03, 2016 - Copyright © 2016 SISEF
Research Articles
Abstract
Drought-avoidance traits of South American Mediterranean tree species are crucial attributes to be considered in nursery practices aimed at improving the performance of seedlings exposed to intense summer drought in dryland reforestation projects. In this study, we determined the relation between nursery fertilization doses and the development of drought-avoidance traits of the soapbark tree Quillaja saponaria (Mol.) under contrasting watering regimes following post-planting. Seedlings were grown for 6 months using four increasing doses of controlled-release fertilizer (0, 3, 6, and 12 g L-1 of Basacote® Plus 15:8:12). After outplanting, half of the seedlings were watered weekly and the other half were left unwatered for one growing season from September 2011 to May 2012. Seedlings were periodically measured for morphological and ecophysiological parameters, and carefully harvested for root measurements at the end of the study. Our results showed that high fertilization doses produced significantly larger seedlings in the nursery with high nitrogen and phosphorous foliar concentrations, which resulted in a significantly higher shoot dry mass after outplanting. Unfertilized seedlings grown with water application had a significantly higher stem diameter, root dry mass and lower shoot/root compared with seedlings with high fertilizer dose. These results highlight the ability of this species to maintain drought-avoidance traits, such as high xylem water potential and chlorophyll fluorescence, during the first 3 months of the 7-month drought period. High nutrient loading, although resulting in improved shoot productivity after outplanting, did not make a significant contribution to the early development of drought-avoidance traits in Q. saponaria.
Keywords
Dryland Forest Restoration, Plant Quality, Root Architecture, Seedling Fertilization, Water-stress Resistance
Introduction
The Mediterranean ecosystems are characterized by strong seasonality with high temperatures and low or no precipitation during the summer months; thus, soil water availability is a major limiting factor for the tree seedling survival and growth ([29], [28]). In South American Mediterranean ecosystems, the reforestation success is limited by both low water availability and poor nursery management and planting techniques, especially in highly degraded sites ([1]). In this area, tree seedling mortality can be close to 70% after first post-planting growing season ([1], [3]). Additionally, the sustainability of reforestation projects is threatened by climate change projections that indicate an increase in drought severity; therefore, plant production and reforestation techniques will need to be adapted to soil water limitations ([38]).
To adapt to water stress conditions and avoid early mortality, plants have evolved different structural and functional traits, such as deep roots, higher root length density, a reduced shoot:root balance, stomatal conductance, and/or osmotic adjustment ([19], [37], [27]). It has been reported that these adaptive traits could be considered in nursery or reforestation practices in areas with Mediterranean-type climate ([39], [26]). In this context, early nutrient loading is suggested to increase drought resistance of seedlings, which help to maintain a positive water balance during initial field establishment ([20], [5], [40]). Seedlings that received high levels of fertilizer increased their root growth, leading to an improved water potential and, consequently, to improved survival rates during the drought season following outplanting ([24]).
Seedlings with an excessive nutrient load and large size do not always respond well after outplanting in Mediterranean systems ([20]). The effect of the fertilization is dependent on the drought intensity during the establishment stage ([4]). Indeed, the positive relation between plant nutrition and growth could be negatively affected in regions with severe drought periods lasting five or more months ([31]). Under this scenario, seedling quality will be associated with the development of phenotypes with drought-avoidance traits, such as low leaf area, high root collar diameter, low shoot:root ratio, low shoot height, and large root volume ([33]). Previous studies carried out in xeric Mediterranean conditions concluded that limited nitrogen (N) application decreased the leaf area, thereby improving seedling resistance to drought after outplanting ([31], [34]). By contrast, high seedling N loading in the nursery produced a morphological imbalance by increasing shoot biomass over root biomass following planting ([34], [4]). Under dry conditions, this morphological imbalance increases transpiration rates and reduces stem water potential, root hydraulic conductivity ([12]), and stomatal conductance ([15]). Low shoot:root ratios could promote a more conservative water use and improve the likelihood of survival after outplanting ([17], [18]). Furthermore, seedlings ability to generate new roots immediately after planting generates a rapid soil colonization and field rooting ([40]). In terms of nutrients uptake and their effects on root development, Trubat et al. ([35]) found a positive effect of low N and phosphorous (P) on increases in specific root length.
Despite the positive effect of nursery nutrient loading on the physiological responses of seedlings, there is little information about the influence of nursery fertilization on the development of morphological traits under different drought levels. We hypothesized that the positive effect of high nursery nutrient loading on the development of drought-avoidance traits is conditioned by the availability of soil water in the field. Thus, the objective of our study was to assess the role of nursery fertilization on the short-term development of drought-avoidance traits of a South American Mediterranean species growing under contrasting field water regimes. The study was conducted using the soapbark tree Quillaja saponaria Mol. (Quillajaceae) which is a representative evergreen tree species of South American Mediterranean forests (30° S, 38° W - [6]), and it is the most common tree species used for the reforestation of the highly degraded forests ([2]). In addition, this species has the ability to develop a strong taproot as one of the main survival strategies to extend drought periods ([26]).
Material and methods
Nursery treatments and measurements
Seeds of Q. saponaria were collected from a forest located in Cuesta Zapata (33° 23′ S, 71° 16′ W), central Chile. Seeds germination and initial growth took place in the Pumahuida Ltda. Nursery (Santiago, Chile). Seedlings were grown for 8 months in individual 200 cm3 pots filled with peat and were sprayed twice daily with water mist, following seedling production protocols in Chilean nurseries ([10]). The seedlings selected for the study (n = 240) were homogeneous in size (6-8 cm in height, 1-1.5 mm in stem diameter).
Seedlings were transplanted to 400 cm3 black polyethylene bags at a research site of the Pontificia Universidad Católica of Chile (33° 29′ S, 70° 36′ W). The potting substrate comprised a mixture of loam soil, sandy soil, and compost at a ratio of 2:1:1 prepared in the nursery. The chemical characteristics of the substrate were as follows: organic matter content (Walkley-Black method) 13.90%; pH (soil:water, 1:2.5) 7.00; electric conductivity (EC) 1.50 dS m-1; N concentration (Kjeldahl digestion procedure) 4.7 g kg-1; P (Olsen method - [25]) 4.1 g kg-1; exchangeable K (ammonium acetate method) 4.6 g kg-1; medium levels of Na+, Ca2+, Mg2+, and K+ were determined by wet-ashing techniques and analyzed using an ion chromatograph ([30]). Seedlings were grown under semi-controlled conditions covered with a 50% Rashell mesh to avoid excess radiation for 6 months starting from January 2011. All the seedlings were watered three times a week with 0.15 L plant-1. At the end of nursery period, seedlings were moved to an open area and watering was reduced to half to prepare them to final transportation to the field site.
In January 2011, the nursery treatments were established as a completely randomized design with four fertilization treatments. Each treatment was applied to 60 replicates (240 plants in total). The experimental unit and the sampling unit was an individual plant. To enable comparisons, the fertilizer treatments comprised incremental doses of N units of 0.00, 0.45, 0.90, and 1.80 g N L-1, equivalent to 0 (unfertilized or 0F), 3 (low dose: 3F), 6 (medium dose: 6F), and 12 g L-1 (high dose: 12F) of a controlled-release fertilizer (CRF), respectively. Basacote® Plus (COMPO, Germany) 15:8:12 (80 g kg-1 NH4+; 70 g kg-1 NO3-; 80 g kg-1 P2O5; 120 g kg-1 K2O; 20 g kg-1 MgO; 50 g kg-1 SO2; 0.4 g kg-1 Fe; 0.5 g kg-1 Cu; 0.6 g kg-1 Mn; 0.2 g kg-1 Zn; 0.2 g kg-1 B; 0.15 g kg-1 Mo) was used with a release period of 12 months (estimated for a constant temperature of 21 °C). This fertilizer has been commonly used in forest nurseries to extend the nutritional contribution to the field phase ([11]).
Upon completion of the nursery phase (June 2011) and before field plantation, a random sample (n = 15) of seedlings from each treatment was analyzed to estimate the effect of fertilization doses on shoot height (SH, cm), stem diameter (SD, mm), slenderness index (SI, cm mm-1), shoot dry mass (SDM, g), shoot:root ratio (g g-1), total root length (total RL, m), total root volume (total RV, cm3), and root dry mass (RDM, g g-1). Roots were separated from dry soil and the remaining soil particles separated by applying water at low pressure, taking care to collect all the fine roots that could be disconnected from the root plug. Shoots were cut at the cotyledon scar. Root morphological variables were quantified using a high-resolution scanner (1200 DPI resolution, Epson Perfection 4490 Scanner®, Nagano, Japan) and image analysis software (WinRHIZO®, Regent Instruments Inc., Quebec, Canada). SDM and RDM were obtained by forced-air oven drying at 65 °C until a constant weight was obtained. The shoot:root ratio was estimated as the quotient between SDM and RDM. The slenderness (SI, cm mm-1) was obtained from the quotient of SH and SD. Foliar concentrations (g kg-1) of N, P, K, Ca, and Mg were determined by the laboratory of the Pontificia Universidad Católica of Chile from a composite leaf sample of three seedlings per treatment that were dried in an oven at 65°C for 48 h. N concentration was determined using a LECO CNS-2000 Macro Elemental Analyzer® (Leco, Michigan, USA). P, K, Ca, and Mg concentrations were determined using wet ash extraction and perchloric acid extracts in an auto-analyzer by gas-segmented continuous flow coupled to a flame photometer ([30]).
Field treatments and measurements
Seedlings were planted in “Quebrada de la Plata” (33° 29′ S, 70° 52′ O; altitude 490 m), located in central Chile. The study site is located in a recently abandoned flat grazing area, near to piedmont. The summer drought lasted 7 months during this study period, with maximum mean temperatures of 30 °C and an absolute lack of rain for 5 months (Fig. 1). Soil is deep and has a colluvial origin, with clay loam textures and abundant gravel. Three composite soil samples (15 sub-samples) were taken from topsoil (0-20 cm) for determine the nutritional status and main chemical and physical properties of the soil in the study area. The results of the analysis in the laboratory indicated a total of 0.044 g kg-1 N, 0.031 g kg-1 P2O5, 0.334 g kg-1 K2O, 6.0 soil pH (H2O), 0.2 dS m-1 EC, and 3.1% total organic matter. Seedlings were outplanted during the winter in 2011. The study area was fenced using a protection mesh 1.50 m high to prevent large animal herbivory ([23]). Manually opened planting holes (0.40 m wide × 0.60 m deep) were distributed 1.5 × 2.0 m apart to prevent interspecific competition for soil water. To avoid transplant shock, seedlings received 2 L of water every 2 weeks for 6 weeks until the beginning of the growing season, with warmer temperatures and no rain until September 2011.
Fig. 1 - Monthly mean, minimum and maximum air temperature, and mean precipitation (PP - bars) in the field during the growing season 2011-2012 (Source: Estación Pudahuel, Dirección Meteorológica de Chile).
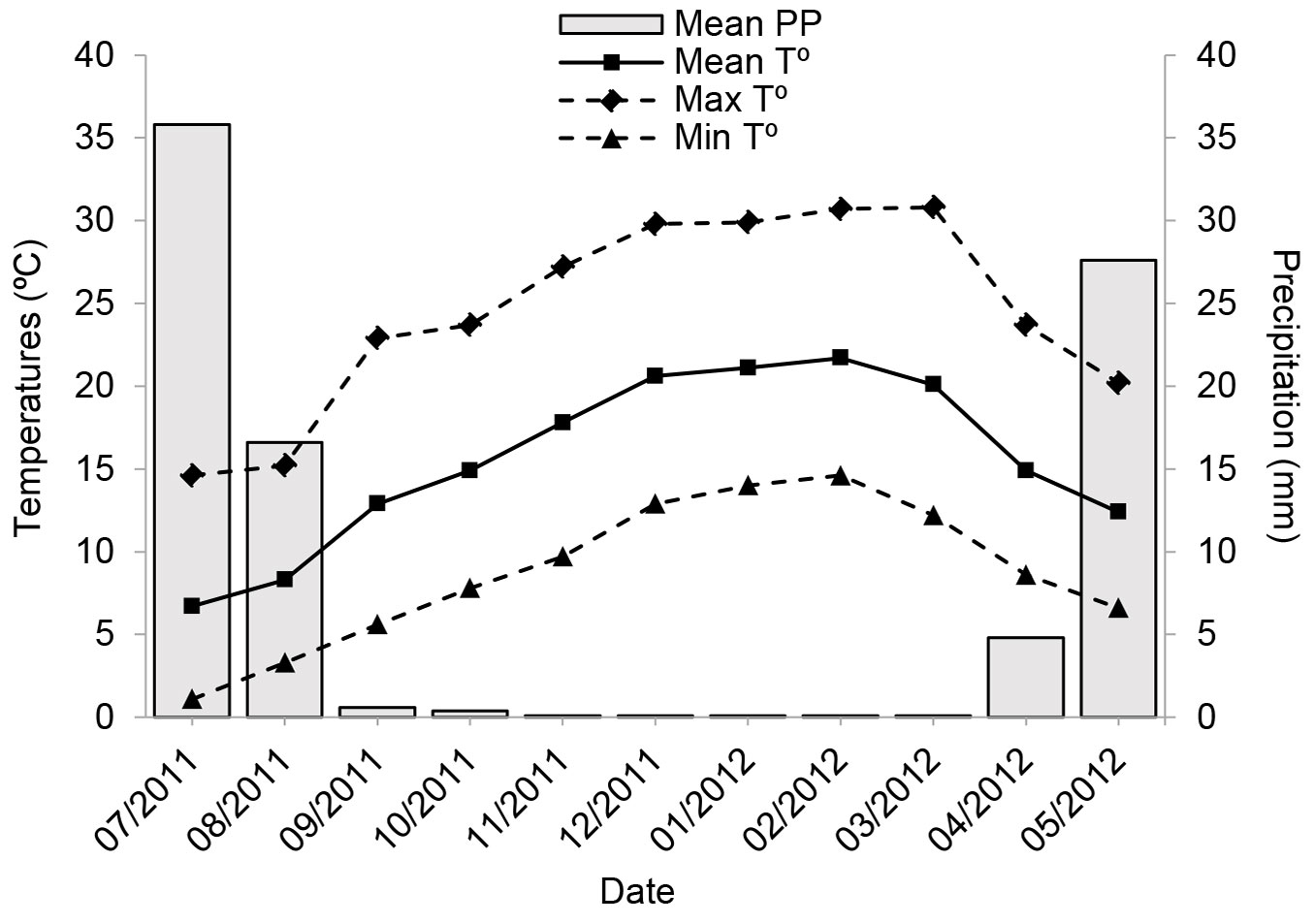
Watering treatments were established in mid-September 2011. The field experimental design was a 4×2 factorial (four levels of nursery fertilization and two levels of field water regimes). Half of the seedlings of each of the four fertilization treatments (0, 3, 6, and 12 g L-1) continued to receive weekly 2 L plant-1 (W+) and half were left unwatered (W-). These watering regimes corresponded to moderate and severe water stress, respectively, based on the criterion commonly used in reforestation operations of semiarid environments ([21], [36]). Precipitation was negligible because of the severe drought during the study period (Fig. 1). Each treatment had 15 replicates (120 plants in total) randomly assigned to each planting spot. The experimental unit and sampling unit were an individual plant. The study site conditions were homogeneous in luminosity, temperature, and airflow, which allowed random assignment without blocking for external factors.
SH, SD, and survival were evaluated monthly. Pre-dawn xylem water potential (ΨW), chlorophyll fluorescence (Fv/Fm), and stomatal conductance (gS, mmol m-2 s-1) were assessed every 2 months between July 2011 and May 2012. For pre-dawn ΨW, five seedlings per treatment were randomly selected. Each sample was obtained from the terminal parts of lignified twigs 5 cm long with three or four mature leaves (second growing season). Measurements were carried out during predawn hours (04:00-07:00) using a Scholander pressure pump (Model 1000®, PMS Instruments Inc., Corvallis, OR, USA). Midday measurements (11:00-13:00) were discarded because the pressure values reached at peak temperature exceeded the safety limit of the instrument (max ΨW = -7.00 MPa). Chlorophyll fluorescence, expressed as maximum photochemical efficiency (Fv/Fm), was measured on 12 randomly selected seedlings at peak radiation (12:00-14:00) using a portable modulated Chlorophyll Fluorometer (Model OS-30®, Opti-Sciences Inc., Hudson, NH, USA) following the protocols described by Maxwell & Johnson ([22]). Each recorded value was a composite of measurements from three different leaves (mature leaves exposed to direct radiation) from the same plant. To obtain gS values (n = 5), an infrared gas analyzer (IRGA) Model CI-340® (CID Inc., Camas, WA, USA) was used; the measurement was performed using the same sampling protocol described for Fv/Fm. We chose sampling days with similar temperatures and humidity for measurement of gS to diminish the variability of atmospheric conditions among dates.
After one growing season (May 2012), trees were dug up (n = 12) to evaluate the morphological variables of the shoot (SH, SD, SDM, and shoot:root ratio) and root systems (total RL, total RV, and RDM). Plants were extracted using a hand shovel. Carefully, we dig a hole 1 m in depth with a radio of 1 m from the base of the tree. Roots were washed applying abundant water at low pressure on a fine mesh sieve to remove soil while retaining thin roots. Roots were grouped according to their diameter as fine (<1 mm), medium (1-2 mm), and coarse (>2 mm) roots. Two new variables were evaluated: fine root volume (fine RV, cm3) and specific root length (SRL). SRL was the quotient of total RL and RDM. The remaining root morphological variables were measured following the same protocol as per the final nursery measurements.
Statistical analysis
For both nursery and field experiments, simple Pearson’s correlations were used to examine the pairwise relationship between measured variables. Before testing, compliance with normality, homogeneity of variance, and linearity assumptions were verified for all variables. In the field experiment, variables that did not match the normality assumptions (SH, SD, total RL, total RV, and RDM) were logarithmically transformed (pre-transformation values are presented). Differences among variables evaluated in the nursery phase of the experiment were determined by a one-way analysis of variance test (α = 0.05), and significant differences were identified by the Tukey’s multiple comparison test (α = 0.05). All data obtained in the field phase were subjected to a two-way ANOVA performed using a general linear model (GLM), to evaluate fertilization doses (F), watering regimes (W), and fertilization dose × watering regime interaction (F × W) effects (α = 0.05). When the F × W interaction was significant (P < 0.05, F test), we conducted the Tukey’s multiple comparison test (α = 0.05). To analyze seedling survival, a chi-square test was applied based on the Kaplan-Meier method with a log-rank test (Mantel-Cox). All statistical analyses were carried out using the SPSS® v 17.0 program (SPSS Inc., Chicago, IL, USA).
Results
Nursery fertilization
Fertilization treatment with the highest dose (12F) significantly affected all seedling morphological variables at the end of the nursery-growing period, with the exception of SD and foliar nutrient concentrations measured before field transplanting (Tab. 1). Seedlings grown at 12F tended to have better shoot and root development compared with seedlings in the other treatment groups. The 0F seedlings had the lowest SDM (Tab. 1). There was a significant effect of 12F on the slenderness (SI) seedlings compared with the 0F. Foliar nutritional analysis showed higher foliar N for the 12F seedlings in comparison to the 0F and 3F. Foliar P concentrations were significantly higher in the 12F seedlings in comparison to all other treatments. Foliar K concentration was higher in the 0F seedlings in comparison to all the other treatments (Tab. 1).
Tab. 1 - Morphological characteristics and foliar nutrient concentrations (N-P-K, g kg-1) of Quillaja saponaria seedlings grown in nursery with different controlled-release fertilizer (CRF) doses (0, 3, 6, and 12 g L-1 Basacote® Plus). Data recorded at the end of the nursery phase in June 2011 before planting. Means ± SE (n = 15) with different letters indicate significant differences after Tukey’s HSD test (P <0.05).
| Variables | Fertilization dose (g L-1) | F value | P value | |||
|---|---|---|---|---|---|---|
| 0 | 3 | 6 | 12 | |||
| Shoot height (SH, cm) | 18.88 ± 0.39 b | 19.65 ± 0.54 b | 20.31 ± 0.22 b | 23.28 ± 0.34 a | 23.681 | <0.001 |
| Stem diameter (SD, mm) | 3.13 ± 0.09 | 2.95 ± 0.05 | 3.08 ± 0.10 | 3.45 ± 0.09 | 1.148 | 0.338 |
| Shoot dry mass (SDM, g) | 1.21 ± 0.03 c | 1.28 ± 0.03 b | 1.38 ± 0.04 b | 1.67 ± 0.01 a | 44.768 | <0.001 |
| Slenderness (SI, cm mm-1) | 6.10 ± 0.21 b | 6.68 ± 0.21 ab | 6.72 ± 0.31 ab | 6.82 ± 0.21 a | 3.858 | 0.014 |
| Root dry mass (RDM, g) | 0.53 ± 0.06 b | 0.56 ± 0.03 b | 0.59 ± 0.07 a | 0.63 ± 0.06 a | 14.908 | <0.001 |
| Shoot/root ratio | 2.28 ± 0.08 b | 2.29 ± 0.07 b | 2.34 ± 0.10 b | 2.65 ± 0.04 a | 8.969 | <0.001 |
| Foliar N (g kg-1) | 22.40 ± 1.30 b | 24.30 ± 0.40 b | 25.50 ± 0.70 ab | 27.70 ± 0.50 a | 7.917 | 0.004 |
| Foliar P (g kg-1) | 2.00 ± 0.10 b | 2.00 ± 0.00 b | 1.90 ± 0.10 b | 2.30 ± 0.10 a | 5.619 | 0.012 |
| Foliar K (g kg-1) | 26.10 ± 1.20 a | 21.00 ± 0.80 b | 19.50 ± 0.30 b | 20.80 ± 0.20 b | 15.028 | 0.001 |
Field survival
Seedling field survival ranged from 80% to 93% in all treatments. Previous nursery fertilization doses (0, 3, 6, and 12 g L-1) and field watering regimes (W+ and W-) had no effect on seedling field survival at the end of the study (χ2 = 1.312, P < 0.726).
Shoot and root morphology
There was a significant main effect of nursery fertilization and field watering regimes on SH, SDM, and total RL (Tab. 2). A significant interaction effect was also detected on SD, total RV, fine RV, SRL, RDM, and the shoot:root ratio (Tab. 2).
Tab. 2 - Two-way analysis of variance (ANOVA) results for treatment main effects for nursery fertilization effect (4 levels) and field watering regimes (2 levels) and their interaction for morphological and ecophysiological variables measured on Quillaja saponaria seedlings after outplanting. ΨW data were analyzed based on ΨW averages of the driest months (September, November, March, and April).
| Variables | Fertilization dose (F) | Watering regime (W) | Interaction F × W | |||
|---|---|---|---|---|---|---|
| F value | P value | F value | P value | F value | P value | |
| Shoot height (SH, cm) | 7.505 | <0.001 | 10.628 | 0.001 | 1.809 | 0.753 |
| Stem diameter (SD, mm) | 6.841 | <0.001 | 153.327 | <0.001 | 35.852 | <0.001 |
| Shoot dry mass (SDM, g) | 55.070 | <0.001 | 12.797 | 0.001 | 1.441 | 0.724 |
| Total root length (total RL, m) | 8.492 | <0.001 | 65.499 | <0.001 | 2.034 | 0.113 |
| Total root volume (total RV, cm3) | 44.973 | <0.001 | 628.780 | <0.001 | 15.148 | <0.001 |
| Fine roots volume (fine RV, cm3) | 37.804 | <0.001 | 204.510 | <0.001 | 3.755 | 0.013 |
| Specific root length (SRL, cm g-1) | 22.507 | <0.001 | 58.911 | <0.001 | 3.072 | 0.031 |
| Root dry mass (RDM, g) | 4.088 | 0.009 | 222.983 | <0.001 | 14.753 | <0.001 |
| Shoot/root ratio (g g-1) | 18.519 | <0.001 | 97.853 | <0.001 | 15.080 | <0.001 |
| Pre-dawn xylem water potential (ΨW, MPa) | 4.791 | 0.007 | 2.260 | 0.143 | 3.484 | 0.027 |
| Maximal photochemical efficiency of PSII (Fv/Fm) | 3.252 | 0.024 | 38.806 | <0.001 | 6.442 | <0.001 |
| Stomatal conductance (gS, mmol m-2 s-1) | 3.227 | 0.035 | 10.358 | 0.003 | 1.001 | 0.680 |
Field seedling height (SH) was greater at 6F seedlings regardless of watering regime (Tab. 3). The 0F seedlings with W+ showed the highest SD value (Fig. 2A). SDM showed significant differences, for having the 12F seedlings the greater values and the 0F seedlings the lowest (Tab. 3).
Tab. 3 - Results of aboveground growth of 2-year old Quillaja saponaria seedlings under four fertilization doses in the nursery and two field watering regimes (W+: 2 L plant-1; W-: unwatered). Data obtained at the end of the dry season after outplanting (May 2012). Values are mean ± SE (n = 12); means with different letters indicate significant differences after Tukey’s HSD test (P <0.05). (a): No interaction of fertilizer dose × watering regime.
| Variables | Fertilization dose (F) | Watering regime (W) | ||||
|---|---|---|---|---|---|---|
| 0 g L-1 | 3 g L-1 | 6 g L-1 | 12 g L-1 | W+ | W- | |
| Shoot height (SH, cm)a | 36.06 ± 1.61 b | 36.36 ± 1.67 b | 41.46 ± 1.04 a | 36.21 ± 0.98 b | 39.09 ± 1.33 A | 35.96 ± 1.31 B |
| Shoot dry mass (SDM, g) a | 4.96 ± 0.22 c | 5.85 ± 0.19 b | 5.62 ± 0.21 bc | 7.29 ± 0.12 a | 6.17 ± 0.16 A | 5.69 ± 0.21 B |
Fig. 2 - Mean stem diameter (A), shoot/root ratio (B), specific root length (C) and root dry mass (D) of Quillaja saponaria seedlings grown under four nursery controlled-release fertilizer (CRF) doses two field watering regimes imposed after outplanting: watered 2 L plant-1 (W+, gray bars) and no watered (W-, white bars). The evaluation was made at the end of the first drought season after outplanting (May 2012). Mean values ± SE (n = 12) are reported. Different letters indicate significant differences after Tukey’s HSD test (P <0.05).
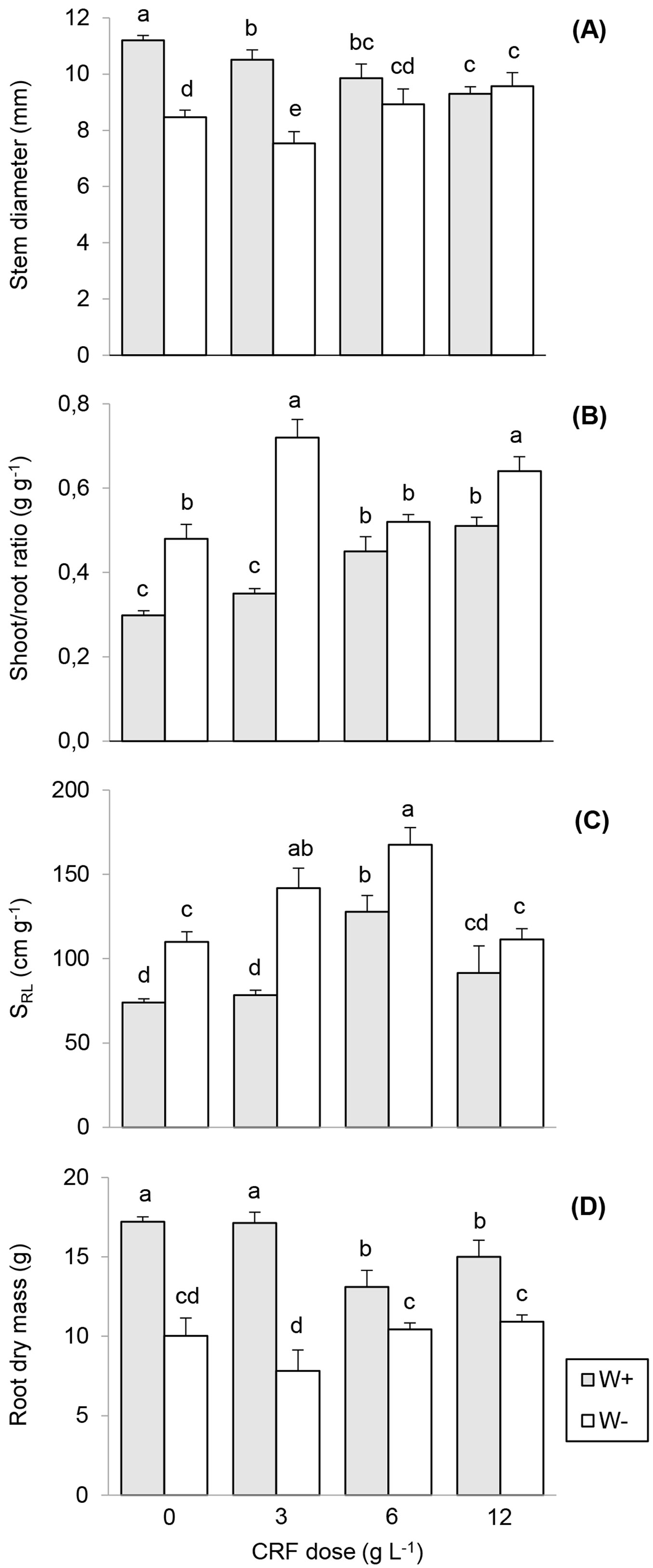
The shoot:root ratio under the W+ treatment was significantly lower in 0F seedlings and 3F. Biomass allocation drastically changed when seedlings were transplanted from the nursery to the field, decreasing from a general average of 2.4 before (Tab. 1) to 0.5 after transplant (Fig. 2B). After the first dry season since outplanting, the lowest shoot:root ratio was observed in 0F and 3F seedlings (Fig. 2B).
The effect of CRF dose on root morphological variables was dependent on the level of soil water stress; water restriction (W-) resulted in a strong limitation of root growth. A significantly higher total RL (Tab. 2) was found for the 6F seedlings under W+ (15.36 ± 1.66 m) compared with the 0F seedlings (12.67 ± 0.50 m). For total RV, the 0F and the 12F seedlings had significantly highest values. Similar results were observed under the W+ treatment (15.50 ± 1.62 cm3 and 16.82 ± 1.17 cm3, respectively). Fine RV represented approximately 50% of the total RV, with a mean value of 6.32 cm3 for all treatments. For 6F and 12F seedlings with W+, fine RV was significantly greater (8.61 ± 0.24 cm3 and 8.45 ± 0.16 cm3, respectively) than for the other treatments (Tab. 2). The highest values of SRL were found for the 3F and 6F seedlings, both under W- (Fig. 2C). The W- condition improved the SRL by more than 30% as compared with the W+ condition. RDM had maximum values at the 0F and 3F seedlings, both under W+ (Fig. 2D).
Xylem water potential, chlorophyll fluorescence, and stomatal conductance
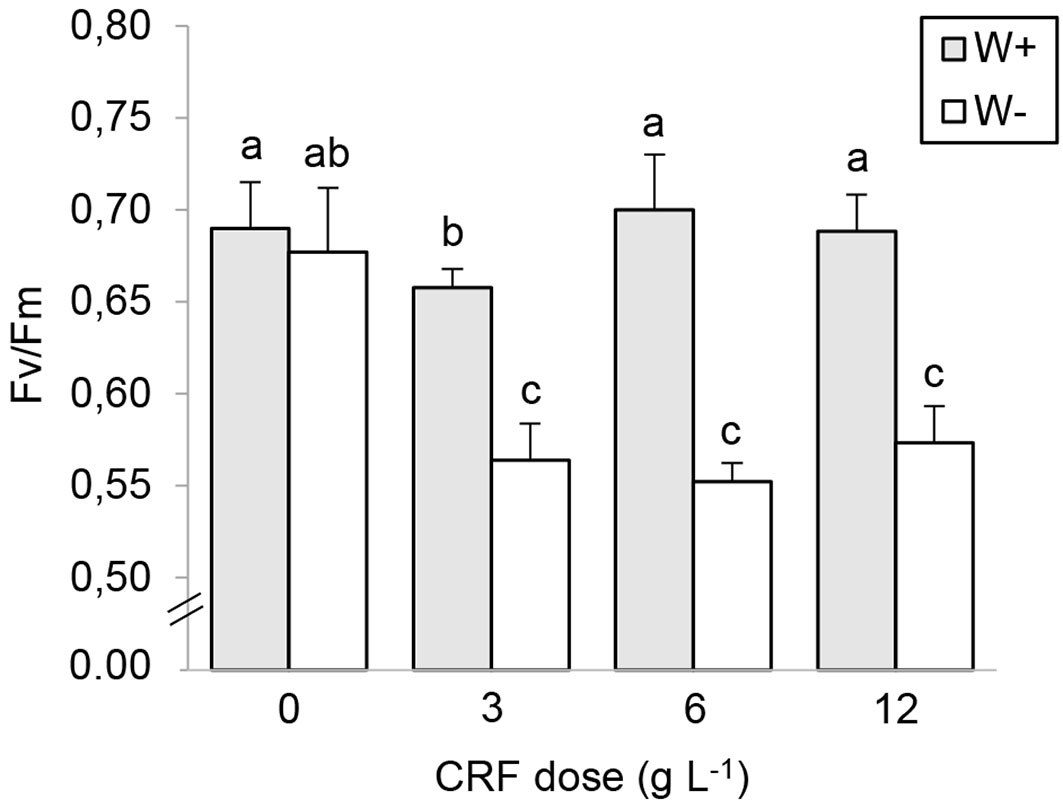
There was a significant effect of nursery fertilization and post-planting watering treatments on gS (Tab. 2), and a significant interaction effect on pre-dawn ΨW and Fv/Fm (Tab. 2). A large drop in pre-dawn ΨW was observed in all nursery fertilization and post-planting watering treatments during the dry period, with minimum values fluctuating from -4.43 to -5.60 MPa at the end of the season, but with no significant differences among treatments (Fig. 3). The combination of no fertilizer and W+ had significantly higher pre-dawn ΨW values during September (-2.25 ± 0.24 MPa) and November (-2.82 ± 0.16 Mpa - Fig. 3). The 0F seedlings had the highest average values of pre-dawn ΨW (-3.09 ± 0.23 MPa) during the driest months compared with the other treatments. Once the wet season started (second week of May 2012 - Fig. 1), the water status of all seedlings recovered quickly, with values of pre-dawn ΨW ranging from -1.05 to -1.50 MPa for that sampling date (Fig. 3), but with no differences among treatments. For midday Fv/Fm, the impact of W- was significant for the three fertilization doses (Fig. 4). The W+ seedlings showed significantly higher values of Fv/Fm compared with the W- seedlings. Seedlings of 0F, independent of the watering regime, had significantly higher values of Fv/Fm at the end of the dry season (April 2012) compared with seedlings in the fertilization treatments. For gS at the end of the dry season (April 2012), there were significant main effects of nursery fertilization and watering regimes (Tab. 2).
Fig. 3 - Mean pre-dawn xylem water potential (ΨW) of Quillaja saponaria seedlings grown under four nursery controlled-release fertilizer (CRF) doses and two field watering regimes imposed after outplanting: watered 2 L plant-1 (W+, solid line) and not watered (W-, dashed line). Mean values ± SE (n = 5); bars with asterisks indicate significant differences (P <0.05, Tukey’s HSD test) in measurements in September and November 2011, indicating greater drought resistance because of a smaller seedling size in relation to the other fertilization treatments.
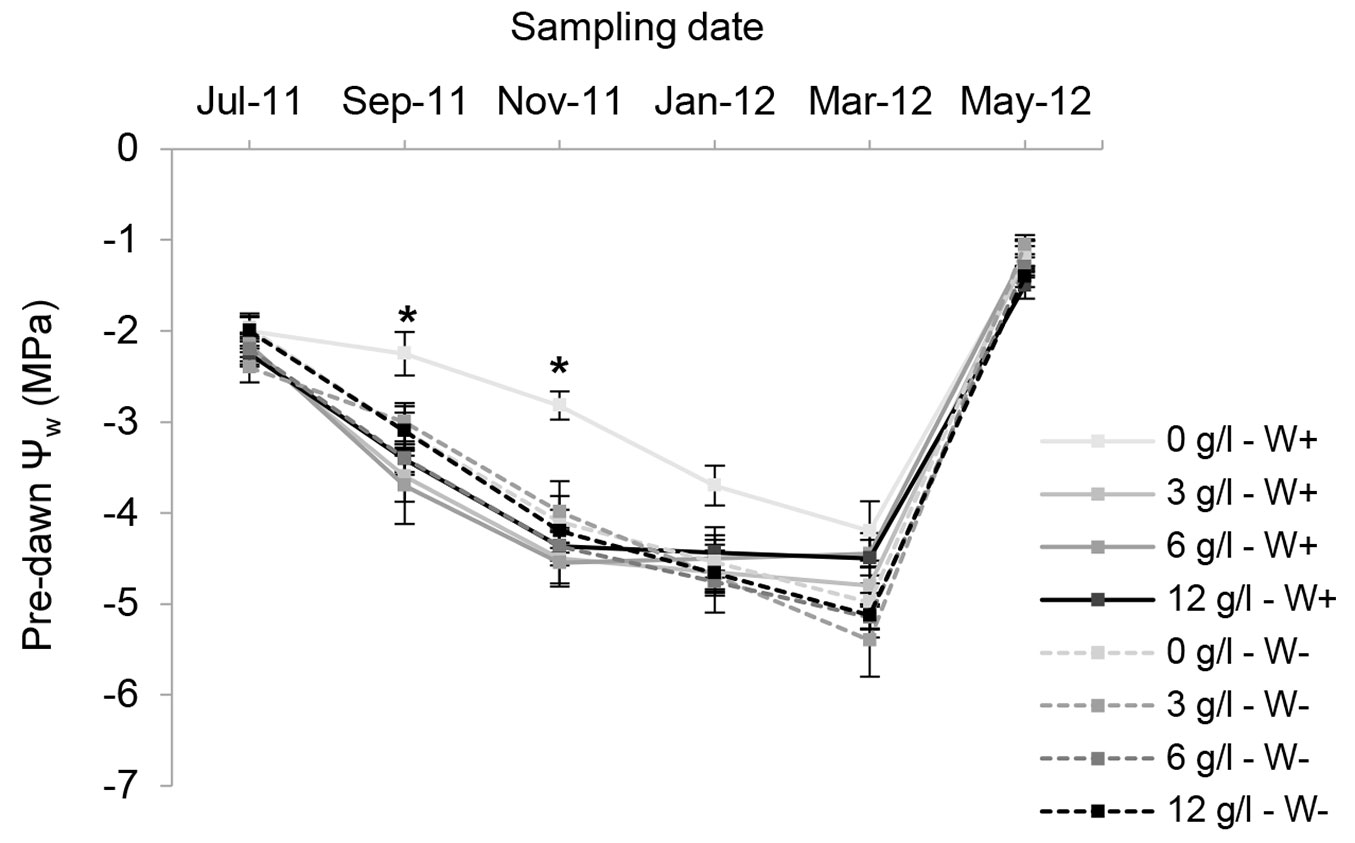
Fig. 4 - Mean Fv/Fm of Quillaja saponaria cultivated under four nursery controlled-release fertilizer (CRF) doses and two field watering regimes imposed after outplanting: watered 2 L plant-1 (W+, grey bars) and no watered (W-, white bars). The evaluation was made at the end of the first drought season after outplanting (May 2012). Mean values ± SE (n = 12); bars with different letters indicate significant differences after Tukey’s HSD test (P <0.05).
Discussion
Morphological responses of seedlings
Nutrient loading at the nursery affected the morphological traits of seedlings under different watering regimes after the first growing season in the field. However, these differences did not condition seedling survival. Under the high dose treatment (12F) applied in the nursery, there was an improvement in shoot productivity after outplanting, but this did not represent a significant contribution to the development of drought-avoidance traits in Q. saponaria when no water was applied for 7 months. The survival of this group of seedlings was higher compared with other reforestation studies that reported early severe mortality for this species ([13], [1]). However, none of these previous studies considered early water application or nursery fertilization, or completely controlled herbivory.
The drastic changes in shoot:root ratio experienced by Q. saponaria in the transition from the nursery to the field indicated its ability to partition biomass to optimize its water balance. Different studies have suggested that reforestation success in Mediterranean systems is affected by the ability of tree seedlings to partition their above- and belowground biomass in a way that optimizes their water balance ([18], [26]). In fact, in the current study, an adequate biomass balance for severe drought conditions might have been achieved without any influence of fertilization. Similar responses were also reported for Mediterranean species when they were fertilized with low doses of CRF (Osmocote®, N:P:K 14:8:14) under controlled conditions ([12]).
Higher doses of CRF (12F) resulted in the greatest RDM on seedlings grown in the nursery. However, this positive response was not maintained after outplanting and contrasted with other recent studies conducted in Mediterranean conditions that suggested that a high nutrient supply generates early root growth and a positive carbon balance that helps sustain the demand for resources to maintain growth biomass ([20], [5], [40]). The low RDM could be attributed to the effects of residual nutrients from the fertilizer contained within the polymer-coated fertilizer, and the root plug, which tends to inhibit the growth ability of roots in the planting hole under low soil water available ([14], [15]). By contrast, in more temperate and wet environments, CRF promoted root development ([32], [11]). The latter was corroborated by the control (0F) and low dose (3F) treatments, both with W+, which had higher growth in RDM after outplanting.
In our study, the highest SRL values were found in the W- treatment, suggesting that Q. saponaria has the ability to alter the morphological components of its root system under drought conditions. This response was maximized in the low (3F) and medium dose (6F) treatments applied in the nursery. Other studies carried out in the Iberian Peninsula with Mediterranean species grown under low soil nutrient status also found an increment in SRL ([35]). We had hypothesized that the effect of high seedling nutritional status on root growth would be reflected by an improvement of the ability to maintain high water potential. However, in our study, the greatest ability to maintain high ΨW during the first months (September to November 2011) of the dry season was only significant for the control (0F) treatment with watering (W+). This greater drought resistance might have been associated with the higher stem diameter, higher root dry mass, and lower shoot:root ratio of these seedlings compared with those subject to other treatments. It has been suggested that this phenotype is favored in environments with severe summer drought ([12], [4]). Our findings differed to those of other studies that reported a positive relation between nutrient availability and drought resistance ([5], [40]), with the exception of SDM, which had high values in the high dose treatment (12F) attributed to high leaf N and P nutrient load at the end of the drought period. However, there was no response in root biomass, which did not match what ex-pected. Under mild Mediterranean summer droughts, there is a positive effect of larger seedling size with high N concentration on water status and post-planting survival ([20], [24]). Differences in the results of these and the current study could be due to the length of the drought period (7 months) that occurred during this study, which was more severe than in other studies conducted in the Iberian Mediterranean region.
Seedling ecophysiological performance after outplanting
Following 7 months of drought, the control (0F) treatment with no water (W-) had the highest Fv/Fm, compared with the fertilized treatments. The latter suffered a drastic decrease in Fv/Fm values under water stress (W-), supporting the idea that small seedling phenotypes might exhibit a better resistance to an extended drought. These effects are further accentuated in semiarid Mediterranean environments, given the high light intensity and temperatures reaching the leaf surface ([8]). Vilagrosa et al. ([39]) reported an exponential decrease of Fv/Fm, to values close to 0.20 in an experiment conducted in a Mediterranean zone of eastern Spain, where Quercus coccifera seedlings were preconditioned to drought. Our study showed Fv/Fm values greater than 0.55 in all treatments, illustrating the high photochemical efficiency of Q. saponaria under conditions of low water availability.
The water status of seedlings was strongly altered by summer drought. It is likely that the intensity of water stress at this stage could overwhelm the fertilization effects on ΨW. However, following the start of the wet season (May 2012), seedlings rapidly recovered their plant water status, reaching higher ΨW values than before the onset of drought. A similar response in water potential was experienced by Quercus rubra seedlings as the water supply recovered ([16]). In addition, these authors also reported an increase in photosynthetic activity and stomatal conductance, which could indicate an adaptive response to drought. One of the key adaptive morphological characteristics that could explain the high resistance is the deep rooting habit of Q. saponaria. Giliberto & Estay ([9]) found a correlation between water status and ΨW during summer drought for Q. saponaria, contrasting to other South American Mediterranean species. This ability to control the water balance could be influenced by the ability of Q. saponaria to rapidly modify its biomass allocation of shoots and roots during the growing season ([7]). However, these studies were conducted in sites that were highly degraded, and more exposed to herbivores ([23]).
Conclusions
Our results suggest that high fertilization doses produced larger seedlings in the nursery, with high N and P foliar concentrations, which resulted in increased shoot dry mass after outplanting. However, unfertilized seedlings developed better drought-avoidance traits after outplanting, such as higher stem diameter, higher root dry mass, and lower shoot:root ratio, but these were associate with high water availability. Drought-avoidance traits influenced the ability to maintain higher xylem water potential during the first months of drought and chlorophyll fluorescence at the end of summer season. Therefore, our study demonstrated that higher fertilizer doses applied in nursery, although improving shoot productivity after outplanting, do not represent a significant contribution to the development of drought-avoidance traits in Q. saponaria under drought conditions. For dryland reforestation, a relevant nursery decision is the downward adjustment of fertilization doses to obtain seedlings better conditioning to anticipate field moisture stress.
Acknowledgements
This work was supported by CONICYT N°24121675-2012, MECESUP-UC0707, and Minera AngloAmerican Chile. The authors would like to thank Vivero y Jardín Pumahuida Ltda for providing seedlings and Germán Greve from the Universidad de Chile. We thank the Marie Curie Actions fellowships (7th European Community Framework Programme), ForEAdapt 269257, and the Center of Applied Ecology & Sustainability (CAPES) by CONICYT FB-0002-2014. We thank Julio Calderón, Mónica Escobar, and Rommy Oliva who provided technical support in the field and laboratory work. We are thankful to James Burger for his collaboration and constructive comments.
References
Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Authors’ Info
Authors’ Affiliation
Eduardo C Arellano
Pablo Becerra
Rosanna Ginocchio
Center of Applied Ecology & Sustainability (CAPES), Pontificia Universidad Católica de Chile, Avenida Libertador Bernardo O’Higgins 340, 8320000 Santiago (Chile)
Pablo Becerra
Rosanna Ginocchio
Departamento de Ecosistemas y Medio Ambiente, Facultad de Agronomía e Ingeniería Forestal, Pontificia Universidad Católica de Chile, Avenida Vicuña Mackenna 4860, 8320000 Santiago (Chile)
Department of Natural Systems and Resources, Universidad Politécnica de Madrid, Ciudad Universitaria s/n, 28040 Madrid (Spain)
Corresponding author
Paper Info
Citation
Ovalle JF, Arellano EC, Oliet JA, Becerra P, Ginocchio R (2016). Linking nursery nutritional status and water availability post-planting under intense summer drought: the case of a South American Mediterranean tree species. iForest 9: 758-765. - doi: 10.3832/ifor1905-009
Academic Editor
Claudia Cocozza
Paper history
Received: Oct 27, 2015
Accepted: Jan 22, 2016
First online: Jun 03, 2016
Publication Date: Oct 13, 2016
Publication Time: 4.43 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2016
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 55500
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 45775
Abstract Page Views: 3967
PDF Downloads: 4331
Citation/Reference Downloads: 37
XML Downloads: 1390
Web Metrics
Days since publication: 3719
Overall contacts: 55500
Avg. contacts per week: 104.46
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Jul 2026)
Total number of cites (since 2016): 19
Average cites per year: 1.73
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Use of alternative containers for promoting deep rooting of native forest species used for dryland restoration: the case of Acacia caven
vol. 10, pp. 776-782 (online: 02 September 2017)
Technical Reports
Nursery practices increase seedling performance on nutrient-poor soils in Swietenia humilis
vol. 8, pp. 552-557 (online: 09 December 2014)
Research Articles
Conservation of Betula oycoviensis, an endangered rare taxon, using vegetative propagation methods
vol. 13, pp. 107-113 (online: 23 March 2020)
Research Articles
Combined pre-hardening and fall fertilization facilitates N storage and field performance of Pinus tabulaeformis seedlings
vol. 9, pp. 483-489 (online: 07 January 2016)
Research Articles
Controlled-release fertilizers combined with Pseudomonas fluorescens rhizobacteria inoculum improve growth in Pinus halepensis seedlings
vol. 8, pp. 12-18 (online: 12 May 2014)
Research Articles
The effect of calcium on the growth of native species in a tropical forest hotspot
vol. 11, pp. 221-226 (online: 01 March 2018)
Research Articles
Seedling quality and short-term field performance of three Amazonian forest species as affected by site conditions
vol. 17, pp. 80-89 (online: 21 March 2024)
Research Articles
Size and age: intrinsic confounding factors affecting the responses to a water deficit in black spruce seedlings
vol. 8, pp. 401-409 (online: 09 December 2014)
Research Articles
Substrates and nutrient addition rates affect morphology and physiology of Pinus leiophylla seedlings in the nursery stage
vol. 10, pp. 115-120 (online: 02 October 2016)
Research Articles
Forecasting the field performance of Austrian pine seedlings using morphological attributes
vol. 10, pp. 99-107 (online: 13 October 2016)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







