Soil fungal communities across land use types
iForest - Biogeosciences and Forestry, Volume 13, Issue 6, Pages 548-558 (2020)
doi: https://doi.org/10.3832/ifor3231-013
Published: Nov 23, 2020 - Copyright © 2020 SISEF
Review Papers
Abstract
Land use change is one of the major causes of biodiversity loss, mostly due to habitat change and fragmentation. Belowground fungal diversity is very important in terrestrial ecosystems, however, the effect of land use change on soil fungal community is poorly understood. In this review, a total of 190 studies worldwide were analyzed. To monitor the effect of land use change, different fungal parameters such as richness, diversity, community composition, root colonization by arbuscular mycorrhizal (AM) and ectomycorrhizal (ECM) fungi, spore density, ergosterol, and phospholipid fatty acid (PLFA) content and AM fungal glomalin related soil protein (GRSP) were studied. In general, results from analyzed studies often showed a negative response of fungal quantitative parameters after land use change from less-intensive site management to intensive site management. Land use change mostly showed significant shifts in fungal community composition. Considering land use change types, only 18 out of 91 land use change types were included in more than 10 studies, conversion of primary and secondary forest to various, more intensive land use was most often represented. All these 18 types of land use change influenced fungal community composition, however, the effects on quantitative parameters were mostly inconsistent. Current knowledge is not sufficient to conclude general land use impacts on soil fungi as the reviewed studies are fragmented and limited by the local context of land use change. Unification of the methodology, detailed descriptions of environmental factors, more reference sequences in public databases, and especially data on ecology and quantitative parameters of key fungal species would significantly improve the understanding of this issue.
Keywords
Soil Fungi, Land Use Change, Fungal Diversity, Species Composition, Mycorrhizal Fungi
Introduction
Soil fungi represent one of the most diverse assemblages of organisms on the Earth. They inhabit a wide range of ecological niches and play an essential role in ecological and biogeochemical processes ([14]). Taxonomically, they are grouped mainly within the nine phyla -Opisthosporidia, Chytridiomycota, Neocallimastigomycota, Blastocladiomycota, Zoopagomycota, Mucoromycota, Glomeromycota, Basidiomycota and Ascomycota ([50]) - and categorized into four ecological guilds - endophytes, mycorrhizal fungi, saprotrophs, and pathogens ([100]). Endophytic fungi inhabit living plants and do not cause disease. Mycorrhizal fungi are symbionts that assist plants in nutrient and water uptake from soil and coping with stress conditions ([71]). Most terrestrial plants live in symbiosis with arbuscular mycorrhizal (AM) fungi, phylogenetically uniform groups within Glomeromycota and characterized by a low host specificity. Ectomycorrhizal (ECM) fungi are highly host specific symbionts of only 4.5% of terrestrial plants (mainly trees, often dominated in boreal and temperate forests) and evolved several times within Basidiomycota and Ascomycota ([9]). For this reason, there is no simple marker that would distinguish them from other trophic groups without identification to the genus. In contrast, pathotrophs cause diseases to plants and other biota ([1]). The saprotrophs serve as primary degraders of organic matter in the soil ([69]). Thus, being a component of soil microbiota, soil fungi are sensitive to any kind of alteration in the soil environment and any associated changes in the aboveground environment, such as human-mediated land use change.
The term “land use” can be defined as human employment of land cover type (biophysical state of the earth’s surface and immediate subsurface, e.g., forest, grassland, cropland, etc.) for certain purpose or function ([7]). It is often difficult to define the types of land use because it varies with land cover and the anthropogenic activities on it. In a broader sense, the changes in land use types from one state (e.g., primary forest, pasture) to other states (e.g., secondary forest, secondary grassland, cropland, etc.) can be defined as land use change. A particular land use type can be either intensive or extensive based on the degree of anthropogenic activities.
In general, land use change is a function of changes in vegetation structure, soil properties, and anthropogenic activities which are known to influence the soil fungal communities ([68]). Change in aboveground plant diversity, coverage, and community composition can influence soil fungi by regulating microclimatic conditions, nutrient addition to soil or exudation of allelochemicals, presence/absence of host plants for mycorrhizal fungi, etc. ([67], [85], [3]). Moreover, soil factors (soil porosity, soil pH, carbon, nitrogen, phosphorus content, etc.) can significantly shape soil fungal communities ([55], [89]). Likewise, anthropogenic activities like tillage, logging/mowing, fertilization, etc. have pronounced effects on soil fungal communities ([81]).
Although many studies have shown the land use change impact on fungal parameters ([51], [53], [59]), no review studies were published so far. For soil carbon, two meta-analyses were carried out on the effect of land use change on a global scale ([26]) and for tropical areas only ([15]). These two studies illustrate that the greatest loss in soil carbon occurs after the conversion of primary sites to cropland but pointed out the occurrence of local constraints that limit the formulation of a broad conclusion. Accordingly, in the case of soil fungi also, published studies differ in several parameters (geographic region, habitat, methods, and target fungal group) such that it can be very difficult to reach a clear conclusion. As land use change is so widespread in a global context, we aimed (i) to summarize the present knowledge on land use change impact on soil fungi, (ii) highlight the limits of the various methodological approaches, and (iii) describe the current knowledge gaps to encourage further research on this topic.
Current studies
A total of 190 published original research articles worldwide were identified (April 2020) using keywords “Land use AND Fungi”, “Land use AND Microorganisms”, “Restoration AND Fungi”, “Restoration AND Microorganisms” “Secondary succession AND Fungi”, “Secondary succession AND Microorganisms”, “Succession AND Fungi”, “Succession AND Microorganisms” through Web of Science®, Google Scholar® and from the references of the selected papers (Tab. S1 in Supplementary material). These studies were selected based on the following criteria: (i) at least two land use types were compared (Tab. 1); (ii) at least one soil fungal parameter was assessed (Tab. 2). Studies dealing with only one land use type with different management (forest clearing, logging, or thinning, fertilization, crop rotation, tillage system, etc.) and chronosequence in single land use were not included. We focused this study on rural land-use change, thus publications dealing with urban and mining areas were excluded. In such land use types severe disruption of spontaneous soil development, heavy metals, and other chemicals can seriously affect the soil fungal community. For each publication, the land use type, soil fungal parameters, the fungal group (ECM/AM/total fungi) under study, the identification method used, and an overview of results were summarized. Also, the place of study, publication date, soil types, and climate information were tabulated (Tab. S1, Tab. S2, Tab. S3, Tab. S4 in Supplementary material). Most of the studies were carried out after 2008 (Fig. 1).
Tab. 1 - Land use type change considered in the reviewed publications. The figure in each cell indicate the number of publications that considered corresponding land use change. If one paper dealt with several land use types, e.g., change of forest to pasture and then to cropland, score one was added to each land use change. Abbreviations of land use types: (PF) Primary forest; (SF) Secondary forest; (PL) Plantation forest; (PS) Primary shrubland; (SS) Secondary shrubland; (PG) Primary grassland; (SG) Secondary grassland; (WL/RL) Woodland or Rangeland; (PA) Pasture; (AL) Abandoned land; (AF) Agroforestry; (PC) Perennial cropland; (CR) Cropland; (Others) Bareland/Eroded land. Increase in land use intensity follows the order of: PF<SF<PL, PG<SG<CR, SG<PA<PC<CR, SF<AL<CR, etc.
| Land use types |
PF | SF | PL | PS | SS | PG | SG | WL/ RL |
PA | AL | AF | PC | CR |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SF | 24 | - | - | - | - | - | - | - | - | - | - | - | - |
| PL | 17 | 15 | - | - | - | - | - | - | - | - | - | - | - |
| PS | 0 | 1 | 0 | - | - | - | - | - | - | - | - | - | - |
| SS | 5 | 6 | 2 | 1 | - | - | - | - | - | - | - | - | - |
| PG | 0 | 2 | 2 | 1 | 1 | - | - | - | - | - | - | - | - |
| SG | 11 | 20 | 6 | 1 | 7 | 5 | - | - | - | - | - | - | - |
| WL/RL | 0 | 1 | 0 | 0 | 1 | 1 | 4 | - | - | - | - | - | - |
| PA | 13 | 11 | 8 | 1 | 1 | 1 | 10 | 1 | - | - | - | - | - |
| AL | 4 | 8 | 7 | 0 | 2 | 2 | 8 | 1 | 2 | - | - | - | - |
| AF | 2 | 4 | 1 | 0 | 0 | 0 | 1 | 0 | 2 | 1 | - | - | - |
| PC | 16 | 10 | 5 | 0 | 0 | 0 | 5 | 0 | 4 | 3 | 1 | - | - |
| CR | 23 | 27 | 15 | 1 | 6 | 13 | 26 | 5 | 18 | 19 | 3 | 10 | - |
| Others | 2 | 2 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 4 |
Tab. 2 - Effect of land use change (intensification) on different fungal parameters. The figure in each cell indicates the total number of papers dealing with fungal parameter. Publication list is given in Tab. S1 (Supplementary material).
| Fungal parameter | Variable | No difference |
Decrease | Increase | Varying | Change in community |
|---|---|---|---|---|---|---|
| 1. Species richness and diversity | (a) species / OTUs richness | 10 | 28 | 12 | 29 | - |
| (b) diversity | 14 | 23 | 9 | 20 | - | |
| 2. Community structure | (a) Higher taxa/guilds abundance | 4 | 5 | - | 37 | - |
| (b) Species/OTUs composition | 4 | - | - | - | 89 | |
| 3. Root colonization | - | 3 | 14 | 4 | 8 | - |
| 4. Spore density | - | 4 | 17 | 10 | 11 | - |
| 5. Fungal biomass | (a) Phospholipid fatty acid content | 3 | 17 | 4 | 17 | - |
| (b) Soil ergosterol content | - | 3 | - | - | - | |
| (c) ITS copy number/ Gene copy number | 1 | 8 | 1 | 1 | - | |
| 5. Glomalin related soluble protein | - | 1 | 7 | - | 4 | - |
| 6. Others | (a) % hyphal length | 1 | 3 | - | 1 | - |
| (b) Colony forming unit | 1 | 1 | 1 | 2 | - |
Fig. 1 - Number of publications based on years of appearance.
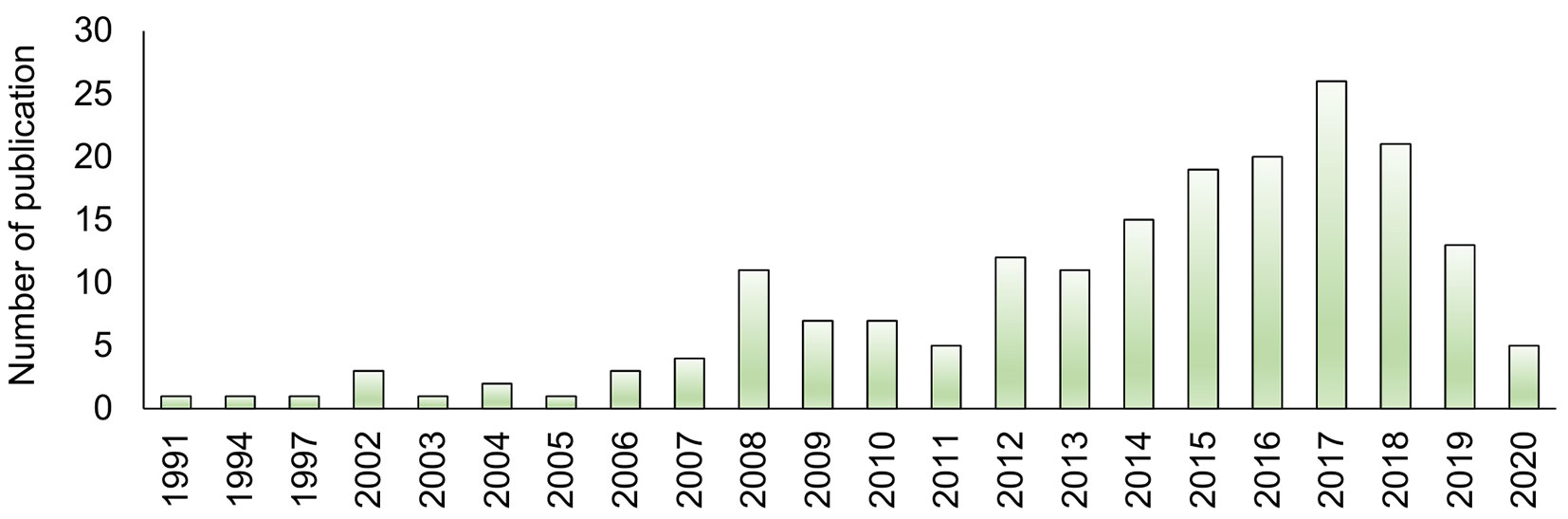
An increase in the number of studies in the last decade appears to be driven by an increasing concern of the effects of land use change, as well as by the use of next-generation sequencing methods (NGS) of environmental samples that has facilitated soil fungal research. A higher number of studies were carried out in Asia (73 studies) followed by Europe (49), South America (32), North America (17), Africa (12), Oceania (7). European and South American studies were distributed throughout the continent, whereas the Asian studies were mainly concentrated in the temperate region of China.
A higher proportion of studies compared natural and semi-natural habitats (primary/secondary forest, grassland, shrubland, and woodland/rangeland) with croplands, which represent a typical example of extensive land use and intensive land use types, respectively. In addition, extensive land use types resulting from agricultural land abandonment, abandonment of pasture, plantation in disturbed areas, or natural successional changes, are also commonly compared (33 studies). Tab. 1shows the number of studies analyzed for various types of land use change. The majority of studies (27) compared secondary forest with cropland, 26 studies croplands with secondary grassland, 24 studies primary with secondary forests, and 23 studies primary forest with cropland.
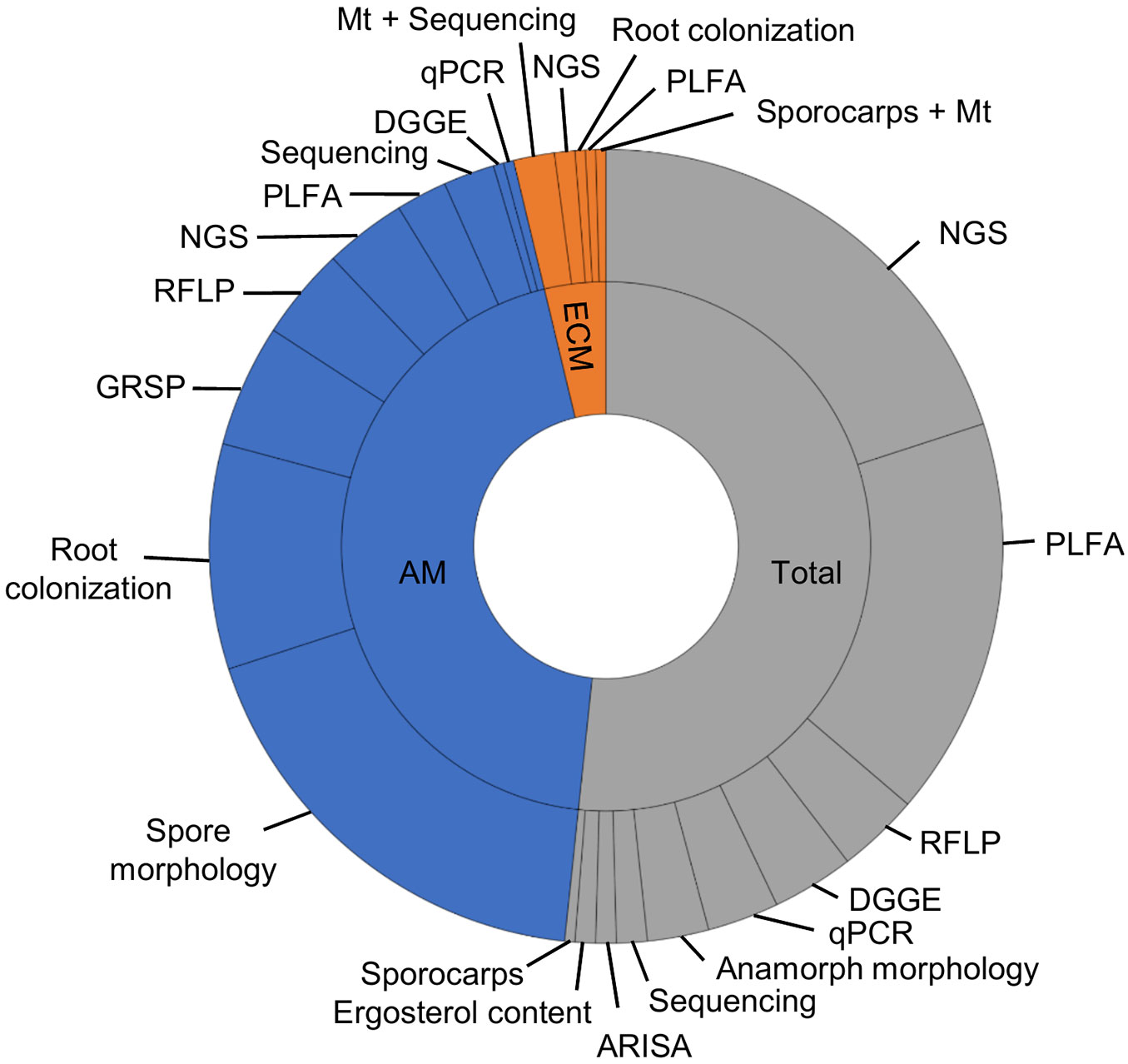
In general, all fungal groups were assessed in studies after the introduction of the NGS methods (ca. 2008). Before this date, most studies were focused on AM fungi and considerably fewer studies dealt with ECM fungi (9) or saprotrophic fungi (micromycetes) assessed by cultivation methods (6 - Fig. 2). The low numbers of studies on ECM fungi could be due to the absence of ECM trees for comparative evaluation across the land use types. In the earlier studies identification of the fungi was based mostly on morphology.
Fig. 2 - Proportion of published papers dealing with monitoring the impact of land use change according to the fungal group studied (AM: arbuscular mycorrhizal fungi; ECM: ectomycorrhizal fungi; and total fungi) and the different methods used to measure fungal parameters. If a particular paper deals with more than one group of fungi (total, AM, or ECM fungi), each group is scored with 1. (PLFA): phospholipid fatty acid analysis; (RFLP): restriction fragment length polymorphism; (DGGE): denaturation gradient gel electrophoresis; (Mt): morphotyping; (ARISA): automated ribosomal intergenic spacer analysis; (GRSP): glomalin related soil protein; (qPCR): quantitative PCR; (Sequencing): Sanger sequencing; (NGS): next generation sequencing.
Land use change impact on soil fungi
Following a land use change, different fungal parameters showed a negative response to land use intensification (e.g., primary forest < secondary forest < plantation forest < cropland). However, positive response or varying/no-difference in observed fungal parameters were also found. The clear result was found in the case of fungal community composition. Eighty-nine out of 93 studies focusing on changes in community composition showed a significant change in fungal community composition (Tab. 2). The majority of the studies also showed a decrease in quantitative fungal parameters following land use intensification. In the following subsections, some representative results have been described briefly.
Species richness and diversity
Studies on the effects of land use change on soil fungal richness and diversity (measured using different indices) used direct analysis of soils, or used soils incubated in a greenhouse with trap plants, such as Sorghum sp., Plantago sp., Allium sp. ([51]). In early studies, identification of species to quantify species richness and diversity was based on morphological characteristics, such as spore morphology of AM fungi ([5]), ECM morphotypes, sporulation of anamorphs (saprotrophs - [41]), and sporocarp morphology ([78]). Along with these methods, molecular fingerprint methods, such as denaturing gradient gel electrophoresis (DGGE - [16]), restriction fragment length polymorphisms (RFLPs - [34]), and automated ribosomal intergenic spacer analysis (ARISA - [54]) were also used. Later, next-generation sequencing methods (454 pyrosequencing, Illumina sequencing, and Ion Torrent platform) have been introduced ([76]). For DNA based analysis, the SSU ([43]), LSU ([85]) or ITS region ([86]) were usually amplified using appropriate primers. The estimates were then expressed in terms of actual species number or operational taxonomic unit (OTUs).
Generally, soil fungal richness decreased with the conversion of primary forest/grassland/shrubland to the more intensive land use sites, like secondary forest, plantation forest, perennial cropland, cropland (monoculture or crop rotation system -[51], [67]). For instance, during the conversion of forested areas to cropland, or to perennial cropland and fallow land ([77]), a reduction in the AM fungal richness was found. Likewise, when analyzing ECM fungal richness in roots of poplar plants in different land use (former cropland, former grassland, and former spruce stand), lower ECM fungal richness was found in former cropland sites compared to that found in the former spruce forest or former grassland ([23]). Although a decrease in species/OTUs richness values were common after increasing land use intensity, some contrasting results were also observed. The richness value either increases with increasing land use intensity (10 studies) or there is no clear direction of change (29 studies - Tab. 2). For example, higher AM fungal species richness was found in the secondary forest compared to the primary forest in the Amazon ([74]). Higher observed OTUs richness was found in Jungle Rubber than in the rain forest in Sumatra, Indonesia ([8]). While comparing perennial cropland (vineyard) to the adjacent sclerophyllous forest in the Mediterranean, there was no significant difference in total fungal species richness ([11]).
Similar to the total species richness estimates, diversity estimates based on the Shannon index of AM fungi ([30]), ECM fungi and total fungi were found to decrease significantly with land use intensification ([23]). The comparison of perennial cropland sites with the secondary forest and fallow land showed a lower value of the Shannon diversity index of AM fungi for cropland ([72]). Likewise, higher OTUs diversity of AM fungi was observed by comparing grassland sites with agricultural sites ([39]). Lumini et al. ([37]) found that the sites with low inputs (pasture and covered vineyard) showed a higher AM fungal diversity than those subjected to human inputs (managed meadow and tilled vineyard). But there were some equivocal results also. For instance, the conversion to intensive land use sites (primary forest to secondary forest) did not necessarily lead to the decrease in species richness, and/or a change in the diversity index of soil fungi ([35]). Comparable diversity was reported while comparing the soybean field with plantation forests in temperate China ([75]). A higher Shannon diversity index of AM fungi was found in cropland sites compared to Atlantic forest sites ([60]). Conversion of the pristine forest into other land uses (in tropical humid Amazon) did not reduce AM fungal diversity ([74]). Similarly, McGuire et al. ([40]) did not found significant differences in total fungal diversity between primary forest, regenerating forest, and oil palm plantation.
The maintenance of species richness may be due to the increase in plant diversity, density/coverage, as well as abiotic factors among land uses that promote diversity even in intensive land use systems. A pattern of increased AM fungal richness in cropland, agroforestry, and secondary forest sites compared to mature tropical forest sites has been explained by an increasing number of plant species that harbor different AM fungi growing in these land use types ([74]). It has been observed that some mature tropical forests with climax plant communities are associated with the low richness of AM fungi ([70]). However, unaltered richness and diversity estimates do not mean that species composition has not changed. Using species richness as a parameter, replacement of species can result in a similar level of diversity and is indistinguishable from no impacts of land use change ([96]).
Community structure
Modification of soil fungal species/OTU composition (estimated by the same methods mentioned in the previous chapter) after land use change is quite common and most obvious while comparing forest sites with cropland sites. As expected, various native forest types showed different fungal species composition compared to secondary forest ([45]), plantation forest ([34], [43], [8]), pasture ([87]), grassland ([97]), and perennial cropland ([36]). After the conversion of rain forest sites to rubber plantation, Kerfahi et al. ([35]) observed changes from a Basidiomycota dominated to an Ascomycota dominated community. However, a study from China’s Loess Plateau showed increasing abundance of Ascomycota and decreasing abundance of Basidiomycota while comparing cropland to plantation ([94]). During the conversion of primary forest to oil palm monoculture (in lowland evergreen rain forest of peninsular Malaysia dominated by Dipterocarpaceae), representatives of Archaeorhizomycetales, Hypocreales, and Saccharomycetales were found more abundant, whereas Cantharellales and Mortierellales dominated the primary forest sites. For ECM genera Amanita and Lactarius were more abundant in the regenerating forest, whereas Craterellus and Russula were more abundant in the primary forest ([40]). Replacement of grassland with willow and poplar plantation leads to the enrichment of putative pathogenic and ECM fungi associated with trees ([93]). Significant differences in fungal communities were found in different land use types (bog, rough grazing site, and forest plantation sites) when investigating ericoid mycorrhizal diversity in two different ericaceous plant species, Vaccinium macrocarpon and Calluna vulgaris ([28]).
Comparing AM composition of forests, croplands, and grasslands in the southern part of Tibetan Plateau, Glomeraceae were less abundant in forests, Gigasporacae in grasslands, and Acaulosporaceae in croplands than in other land use types ([92]). In agricultural systems, the occurrence of Acaulospora denticulata, Glomus ambisporum, and Claroideoglomus etunicatum was significantly affected by cropping and non-cropping systems in Taita Hills, Kenya ([33]). A study in Terceira Island (Azores, Atlantic Ocean) showed that semi-natural pastures were dominated by species of Acaulospora and Scutellospora, particularly S. calospora and A. cf. myriocarpa. Whereas, for intensively farmed pastures, species with glomoid spores, and members of the two genera Claroideoglomus and Paraglomus were found most frequent and abundant ([42]).
Root colonization by AM and ECM fungi
Mycorrhizas on roots have also been quantified to evaluate the land use change impacts on AM and ECM communities. In ECM fungi, the percentage of root colonization (proportion of mycorrhizal and non-mycorrhizal root tips) and for AM fungi the percentage of arbuscules, vesicles, and hyphae were quantified. For both types of mycorrhizas, the extent of colonization showed variation with land use change and could be explained by alteration of soil properties after land use change ([47]).
The percentage of colonization by ECM fungi of Salix sp. was found to be higher in the natural forest compared to short rotation coppice ([31]). The difference was explained by soil water content, an initial inoculum of ECM fungi, host genotype, and soil pH. In a study of edaphic effects on symbiosis during soil development, mycorrhizal colonization shifted from AM fungi to ECM fungi (the plants had both types of mycorrhizal association) in response to decreasing phosphorus content in the soil. Such a shift in mycorrhizal association enables the acquisition of organic phosphorus via extracellular phosphatase activity ([2]).
The effect of increasing intensity of land use on AM fungal colonization is less clear than that for ECM fungi, being this possibly due to lower host specificity of AM fungi ([25]). In the investigation of Islas et al. ([32]), no relationship between the degree of land use intensification (pristine soil to agricultural soil) and root colonization of AM fungi was observed. On the other hand, the conversion of natural forested areas to grassland/pasture increased AM fungal colonization ([57]). Analysis of AM colonization in experimental plants established in degraded forest and in planted forest sites, showed lower root colonization in planted forest sites than in degraded forest ([67]). This difference was explained by the increased canopy of the planted forest reducing light and temperature at the forest floor, and thus plant density and hence lower root colonization.
Such increases or decreases in root colonization by ECM and AM fungi after land use intensification can be explained also with soil nutrient availability and perturbation in soil ([57]). Often the increase in soil nutrient availability has a negative impact on mycorrhizal colonization ([82]). Perturbation of the soil (e.g., soil compaction) reduces the porosity of soil, thereby reducing passages for organic nutrients and decreasing fungal mycelium ramification ([18]). On the other hand, the tillage in agricultural land breaks the extraradical mycelium into smaller units, and thus it increases the infectivity of fungal mycelium ([6]).
Spore density
AM fungi produce thick-walled spores as their propagules, and morphological variations in such spores were traditionally employed for genus identification ([66]). Quantifying spore number per volume of soil by direct microscopic counting ([12]) and expressing this count as a spore density/abundance, was used to observe land use effects.
Undisturbed areas, like natural grasslands and rangelands, were reported to harbor higher spore density in comparison to fallow land, pasture, plantation sites, and cropland ([33], [36]). The effect of land use intensification on spore density was observed also at the species level ([53]). For instance, in a comparison of natural sites with pasture, the spore density of Glomus sp. was higher in natural sites while Acaulospora sp. was higher in pasture sites ([55]). Alternatively, spore density was also expressed as infective propagule numbers (the number of spores that can infect the test host plant, determined by the Most Probable Number technique). The total number of infective propagules was found to be significantly higher in pasture compared to traditionally managed cropland ([73]). However, other studies reported comparable spore density of AM fungi in forested sites and fallow sites ([77]). They reported that apart from the overall land use impacts, soil organic carbon and pH levels affect the sporulation of AM fungi. While comparing short fallow land with the perennial cropland and secondary forest, significantly higher spore density was observed in short fallow and found to be related with C:N ratio in soil ([72]). Also, the impact of vegetation has been highlighted. It is known that grassland species have a more abundant fine-root system than trees in a mature forest, and this correlates positively with AM fungal colonization and sporulation ([46]).
Fungal biomass
Quantification of fungal biomass using lipid components of fungi, namely soil ergosterol content and phospholipid fatty acids (PLFA), including neutral fatty acids (NLFA), have also been employed to monitor land use impacts on soil fungi. The unsaturated fatty acid 18:2ω-6.9 was used as a marker of fungal biomass in the community profile ([56]). For AM fungi, 16:1ω-5c ([73]), and for ECM and saprotrophic fungi, 18:2ω-6.9c and 18:1ω-9c ([4]) were also applied. Additionally, estimates of ITS copy number (a proxy estimate of fungal biomass) from quantitative PCR were also used to show land use change effects on fungi ([24]).
In a comparison of farmland and forest sites, soil ergosterol contents were found to follow the order of the intensity of land use, i.e., intensively cropland < pasture < forest soils ([83]). The plantation in mangrove forest sites with coconut, rubber, etc. was also found to reduce the soil ergosterol content by up to 90% ([13]).
The decreasing value of fungal PLFA (18:2ω-6.9) content with increasing land use intensity was reported by Hamer et al. ([27]). Similarly, higher fungal PLFA content was found in forest sites compared to perennial cropland ([16]). The content of PLFAs of AM fungi was lower in forest soils compared to abandoned and heathland soils, whereas the PLFAs of saprophytic and ECM fungi were more abundant in ancient forest soils ([56]). Such trends shown by AM and ECM fungi plus saprophytic fungi have been explained by soil nutrient conditions and differences in host plants. Higher amounts and structural complexity of carbon sources in forests form a bigger niche availability for ECM and saprophytic fungi, whereas higher concentrations of N and P in the former arable site support the AM fungi. However, some contrasting results have also been observed. With some exceptions, higher fungal biomass measured with the PLFA (18:2ω-6.9) was observed in perennial agricultural soil compared to coastal grassland ([17]). Similarly, PLFA content (18:2ω-6.9) was found to be increased in the transition from forest to pasture to an abandoned field, due to the availability of nutrients as grass litter decomposed faster compared to the forest ([61]). However, a study of the conversion of native grassland to pasture showed no significant difference in fungal PLFAs ([91]). It was suggested that no significant differences in soil properties (e.g., available P, soil pH, and vegetation cover) maintained an equal amount of fungal PLFAs in native grassland and pasture. Wang et al. ([89]) also found no differences in fungal PLFA content with the conversion of natural forest to rubber plantations, but the AM fungi PLFA marker (16:1ω-5c) was found to be higher in the natural forest than in the rubber plantation. It has been explained that soil acidification and higher phosphorus content reduced AM fungal biomass in rubber plantation sites.
Using a qPCR approach, no difference in the log number of copies of gene/gm of soil was found in abandoned land compared with plantation forest, but its values were significantly lower than mixed forest ([36]). Similarly, no significant change in the log number of copies of gene/gm of soil between secondary grassland, pine plantation, and cropland was found ([96]). A study by Grantina et al. ([24]), showed the order of log number of copies/gm of soil as former forest > agriculture > pasture > agricultural land.
Glomalin related soil protein (GRSP) of AM fungi
Glomalin is a proteinaceous substance produced by AM fungi which is released to the soil during hyphal turnover and after the death of the fungus ([19]). Different fractions of this protein, namely immunoreactive soil protein (IRSP) and easily extractable immunoreactive soil protein (EE-IRSP) were quantified in soil to assess land use change impacts ([65]).
In the soil of grasslands, total glomalin related soil protein (GRSP) content was higher compared to a plantation site and was even lower in a maize monoculture ([5]). Similarly, native mixed deciduous forest (Acer, Alnus, Juglans, Carya, etc.) contained a higher concentration of GRSP compared to agricultural and afforested sites ([73]). Total GRSP was lower in cropland and fallow land when compared to forest soils covered by the AM-forming timber species Combretodendron macrocarpum, Piptadeniastrum africanum and Disternonanthus benthamianus ([20]). Likewise, glomalin concentration was found lower in cropland than the native forest of temperate Himalaya ([48]). Here, it has been explained that an increase in the root network of trees in the forest leads to an increase in AMF colonization which contributed to higher glomalin concentration. Prolonged arable land use of former grassland soils reduced the content of GRSP further ([62]). However, some studies showed no changes in GRSP content after land use change. For example, when poplar plantation was compared with cropland (maize field) no significant change in GRSP content was found ([88]).
Summary of the individual land use change impacts on soil fungal parameters
Generally, conclusions on the impact of land use change on soil fungi are limited by the number of replications. However, 18 types of land use change were considered in ≥10 replicated studies (Tab. 1). Thus, to extract generalized information needed for stakeholders, a summary of these replicated studies was made. Fig. 3 shows the overall impacts of these 18 types of land use change on qualitative (species composition) and quantitative fungal parameters (richness, diversity, root colonization, spore density of AM fungi, and fungal biomass - Tab. S5 and Tab. S6 in Supplementary material). If we set ≥50% of the results as a boundary of increase/decrease or change, all 18 land use types showed changes in the community composition of all fungal groups (Tab. S5, Tab. S6). When comparing primary forest to secondary forest, secondary grassland, pasture, and cropland, no fungal quantitative parameters were changed. AM fungal quantitative parameters decreased when the primary forest was changed to plantation forest, perennial cropland, or cropland. Surprisingly, total fungal quantitative parameters increased when the primary forest was changed for plantation forests. Brinkmann et al. ([8]) explained this pattern based on the intermediate disturbance hypothesis, which proposed that the highest diversities will occur in ecological systems with moderate disturbances.
Fig. 3 - Summary of the individual land use change impacts on quantitative parameters and species composition of total, AM, and ECM fungi. Bar graph represents the number of studies showing various impacts positive/negative/varying or no impacts on quantitative parameters and the asterisk “*” sign represents the change in species composition <50% of the studies. (PF): Primary forest; (SF): Secondary forest; (PL): Plantation forest; (PG): Primary grassland; (SG): Secondary grassland; (PA): Pasture; (AL): Abandoned land; (AF): Agroforestry; (PC): Perennial cropland; (CR): Cropland.
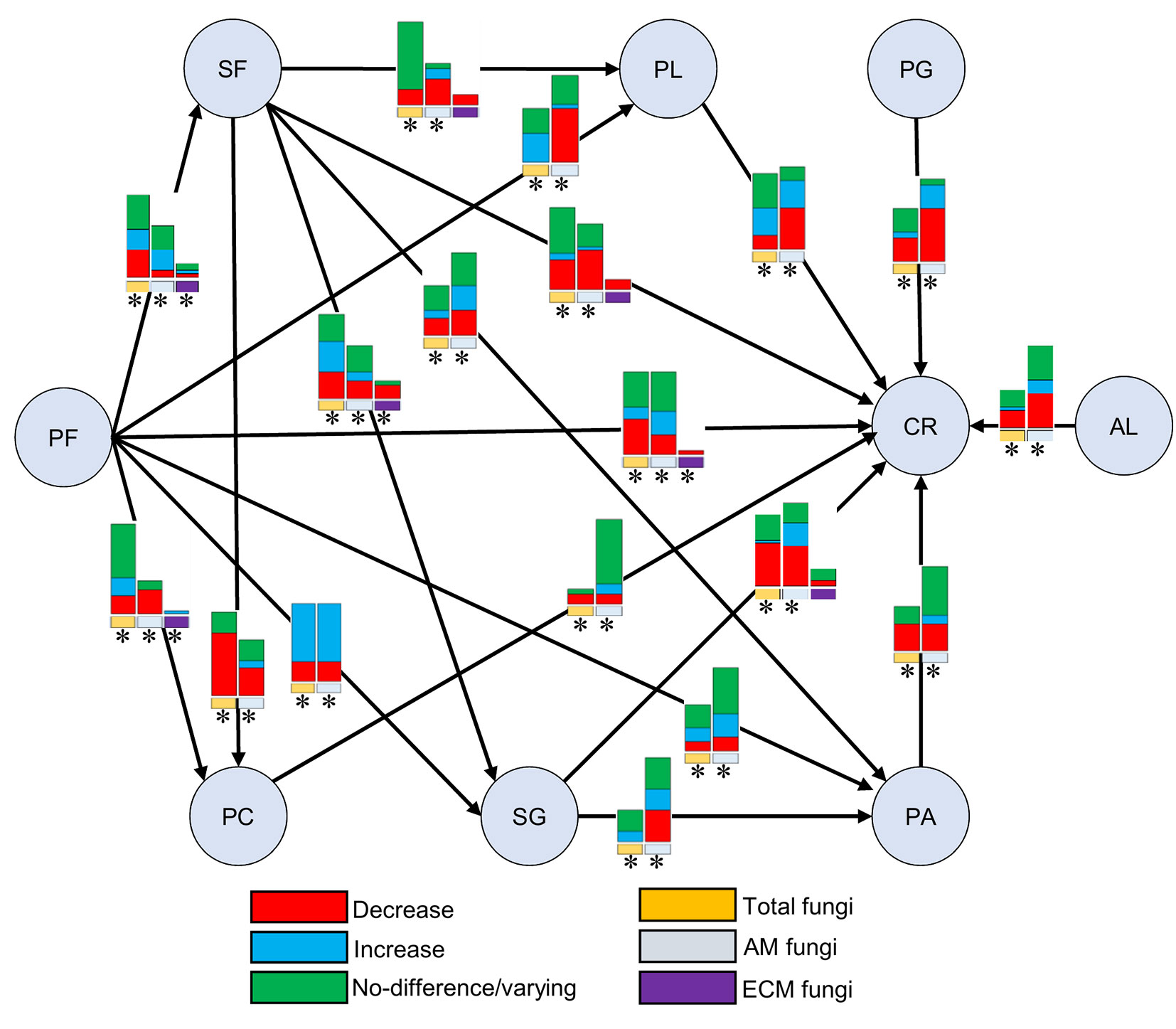
There are no clear changes in fungal parameters when the secondary forest is compared with secondary grassland and pasture. But converting of secondary forest to plantation forest, or perennial cropland or cropland, showed decreases in AM fungal parameters. There are also negative impacts on total fungal parameters following changes of secondary forest to perennial cropland. Only AM fungal parameters decreased when the plantation forest is changed to cropland.
These results demonstrate that it is very difficult to evaluate the response of soil fungal communities based on quantitative parameters. The fact that AM fungi provide a clearer response is probably due to their similar life strategy and low specificity. In contrast, total fungi contain many groups with so many different life strategies that their responses overlap, and the response is very much context-dependent.
Limitations of methodological approaches
The major limitation in evaluating the effect of the land use change on soil fungal communities is a poor consensus in land use type categorization. For instance, some studies defined land use types through the climatic zone ([80]), some by plant cover type ([73]), and some considered parent bedrock composition in single land use types ([51]). In addition, some studies poorly described the vegetation, soil types, and climate of their experimental plots ([38]). So, the effect of land use change cannot be correctly interpreted, because these factors could surpass land use change impact.
The other difficulty in the interpretation of land use change effects is a sampling design. As the experimental plots are distributed at a landscape level, homogeneous replicate plots selection is often difficult. However, there is also the limitation of selecting all studied land use types at one locality. Consequently, plot replicates in a distant location could influence soil fungi due to changes in environmental conditions. For instance, in fragmented agricultural fields, the management of soil fertility may be different, which affects soil fungi. Similarly, during the abandonment of the agricultural field, less productive fields are abandoned earlier, such that it can be presumed that the soil conditions are already different compared to previous land use. Therefore, as much information as possible should be gathered about land use history by interviewing local people.
Discrepancies in sampling is also another important factor. Some studies sampled soil along a transect with plot size as small as 1 m2 ([3]), whereas others sampled 1 ha plots with replicates of soil cores ([60]). The preparation of composite soil samples from replicates of soil cores was a common method among studies. A few studies also considered fractions of different soil horizons ([64]). The preparation of composite soil samples may overlay the vertical differences, i.e., the land use change effect might be only in the topsoil layer, and deeper soil layer remains unaffected. In the case of predictor variables, many studies considered land use types as a typical predictor and soil factors as additional predictors. However, a comprehensive description of vegetation (as a next predictor) was missing in most studies ([98], [8]).
In early studies, delimitation of species (evaluation of species richness and diversity) was based on morphological characteristics such as AM fungal spore morphology ([5]), ECM morphotypes, ([41]), and sporocarps morphology ([78]) that relied on expert knowledge and could differ between studies. Molecular fingerprinting methods like denaturing gradient gel electrophoresis (DGGE) can overlook less abundant species. Introduction of NGS methods (454 pyrosequencing, Ion Torrent platform, Illumina sequencing - [76]) has solved the aforementioned issue. However, it has the limitation that appropriate primers for all fungal groups during PCR amplification must be used ([79]) and several other technical issues ([99]). The species-level identification could be also limited due to missing reference sequences in public databases. Moreover, knowledge of the ecological requirements of many taxa remains unknown ([49]) that arises difficulties in the interpretation of obtained data ([58]). During root colonization assays, obtaining the identity of fine roots from particular plants could be difficult in a field-based investigation. Although the use of trap plants solves this problem, trap plants may not reflect the exact field conditions. The age of plants and the proportion of fine roots could also affect root colonization by AM and ECM fungi. Further, studying spore density in soil could be limited in the sporulation capacity of a particular taxon. Sporulation of AM fungi could differ between AM species and could be influenced by differences in seasonal dynamics ([52]). Assessment of fungal and AM biomass was also employed in several land use change studies. PLFA method has its advantages of being rapid and inexpensive, sensitive, and reproducible but the interpretation of PLFAs as indicative of different groups of organisms is not straightforward. The specificity of different fatty acid markers (e.g. 16:1ω-5c for AM fungi, 18:2ω-6.9c for ECM fungi and 18:1ω-9c for saprophytic fungi) has been questioned ([22]). In the case of AM fungi, GRSP is reputed to be a protein of AM fungi, but its presence in AM fungi only is unclear ([90]). Humic substances present in the soil could also interfere with the measurement of GRSP using colorimetric assays ([44]). Consequently, origin of the soil will influence the results. For instance, GRSP represents a good indicator of land use change effect in herb dominated ecosystems, being more concentrated in permanent than temporal systems. Its content in forests depends probably on mycorrhizal types of dominant trees (AM/ECM). As the amount of GRSP content in soil depends upon hyphal growth and spore production, individual species that remains unaffected by land use change may also have roles in determining the differences in concentrations of GRSP across sites ([63]). To obtain more robust and reliable data usage of several methods, i.e., NGS method accompanied by any morphological method (sporocarps, morphotypes, spores, root colonization) and biomass measurement (PLFA content, GRSP content), is highly recommended.
Conclusion and future perspectives
We found 190 relevant studies dealing with land use impact on soil fungi. The studies came mostly from Asia and Europe and were based mainly on next-generation sequencing, PFLA analysis, or spore morphology of AM fungi. Around 80% of these studies were published in the last decade. Land use type classification in these studies was mainly dependent on the local context. The most studied land use change was a conversion from primary forest to cropland/secondary forests, cropland to plantation forest/secondary grassland/pasture. From 91 types of land use change, only 18 were included in more than 10 studies.
Most of the results showed that species richness and diversity were negatively impacted by land use change, but the results are not unequivocal. This means land use change does not necessarily lead to the changes in the soil fungal diversity. Generally, land use change has greater impacts on community composition. Root colonization of AM and ECM decreases in intensive land use sites. For AM fungi, several studies showed decreases in spore density with increasing land use intensity. Soil ergosterol, PFLA, and glomalin contents were found to follow the order of the intensity of land use, however, also equivocal results were published.
Our study suggests that current knowledge is not sufficient to reach conclusions on the effect of single land use change on soil fungi. This is due to the low number of replicates, extreme variability of local conditions worldwide, different methodological approaches and their limitations, the high species richness of fungi, and different responses of various fungal guilds, dependent on plant cover and soil conditions.
A proper description of land use history and environmental factors and a sufficient number of replicates is required for good comparisons. During soil sampling, it should be taken into account that larger plots are limited by sufficient replication and smaller plots by the representativeness of samples. Thus, widely accepted soil sampling strategies can be devised to have at least some uniformity of plot size, soil core size, and the number of replications, etc. Controlled long-term experiments can be one possible option on how to disentangle loss of diversity due to disturbance and management from natural changes caused by a change of associated host plants or soil properties ([29]).
As NGS methods provide a new level of information in soil fungal research, it should be a basic method to assess community composition. However, it is often unclear whether DNA sequences originate from hyphae, spores, or relic DNA ([10]). Thus, non-molecular methods, such as biomass estimation, provide an important cross-check and additional information. This is especially useful in studies of AM fungi because, due to redundancy caused by a high diversity and many life strategies, quantitative parameters of total fungi often do not change.
Although different parts of the world are covered to date, studies in tropical Asia, temperate areas of the Hindukush region, East Asia, Eurasia, and Africa are very few. Forest conversion to agricultural lands is increasing in tropics, Asia, and Africa ([21]). Because these areas are also an important habitat for biodiversity protection ([95]), more studies are needed to fill this gap.
Apart from methodological issues, the main problem of the reviewed studies is in the interpretation of data obtained. In the case of quantitative parameters, there are no generally accepted average values for a particular land use type. Due to the above-mentioned variability, such values can be obtained only in a local context, with sufficient replicates.
A promising tool for species data interpretation is a trait approach that has been widely used in vegetation science ([84]). However, data on fungal ecology are limited to main guilds assessment only ([49]) without any details about biology and niche requirements of selected species. In a European context, some information could be assessed based on red lists, indicator species lists, or monographies, but in other parts of the world, these sources are mostly not yet available.
Moreover, many fungal species are undescribed for science, and if so, have no reference sequence in public databases (e.g., Dothideomycetes, Leotiomycetes). Data on the properties of higher taxa are very rough and often contrasting life strategies are present within one genus. Therefore, we strongly recommend building the database of key species characteristics to move our knowledge from description and comparison to functionality. It is crucial to obtain more data on the ecology of key species ([58]) to develop ideas on the functional significance of changes in species composition.
This review reveals that soil fungal response to land use change needs further evaluation. Soil fungal community dynamics can be reflected as an indicator of land use change. This understanding will help in combating land change impacts on other biotic communities and assist land-management strategies.
List of abbreviations
(AM): Arbuscular Mycorrhiza; (ARISA): Automated Ribosomal Intergenic Spacer Analysis; (DGGE): Denaturation Gradient Gel Electrophoresis; (ECM): Ectomycorrhiza; (GRSP): Glomalin Related Soil Protein; (ITS): Internal Transcribed Spacer; (LSU): Large Subunit; (NGS): Next Generation Sequencing; (OTU): Operational Taxonomic Unit; (PLFA): Phospholipid Fatty Acid Analysis; (RFLPs): Random Amplified Fragment Length Polymorphism; (SSU); Small Subunit.
Acknowledgement
This work was supported by the Ministry of Education, Youth and Sports of Czech Republic within the CzeCOS program, grant no. LM2018123. We thank all the authors who contributed to the original research articles.
References
Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Authors’ Info
Authors’ Affiliation
Martina Vašutová 0000-0003-0128-874X
Department of Botany, Faculty of Science, University of South Bohemia, Na Zlaté stoce 1, Ceské Budejovice 37005 (Czech Republic)
Douglas Godbold 0000-0001-5607-5800
Pavel Cudlín 0000-0003-1464-5160
Department of Carbon Storage in the Landscape, Czech Academy of Sciences, Lipová 1789/9, Ceské Budejovice 37005 (Czech Republic)
Institute of Forest Ecology, University of Natural Resources and Life Sciences, Peter-Jordan-Straße 82, Wien 1190, Vienna (Austria)
Department of Ecosystem Biology, Faculty of Science, University of South Bohemia, Branišovská 1760, Ceské Budejovice 37005 (Czech Republic)
Corresponding author
Paper Info
Citation
Balami S, Vašutová M, Godbold D, Kotas P, Cudlín P (2020). Soil fungal communities across land use types. iForest 13: 548-558. - doi: 10.3832/ifor3231-013
Academic Editor
Alberto Santini
Paper history
Received: Sep 08, 2019
Accepted: Sep 17, 2020
First online: Nov 23, 2020
Publication Date: Dec 31, 2020
Publication Time: 2.23 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2020
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 49733
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 36810
Abstract Page Views: 7732
PDF Downloads: 4313
Citation/Reference Downloads: 21
XML Downloads: 857
Web Metrics
Days since publication: 2081
Overall contacts: 49733
Avg. contacts per week: 167.29
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Jul 2026)
Total number of cites (since 2020): 31
Average cites per year: 4.43
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Potential spread of forest soil-borne fungi through earthworm consumption and casting
vol. 8, pp. 295-301 (online: 26 August 2014)
Research Articles
Shifts in the arbuscular mycorrhizal fungal community composition of Betula alnoides along young, middle-aged plantation and adjacent natural forest
vol. 13, pp. 447-455 (online: 07 October 2020)
Research Articles
Effect of plant species on P cycle-related microorganisms associated with litter decomposition and P soil availability: implications for agroforestry management
vol. 9, pp. 294-302 (online: 05 October 2015)
Research Articles
Effects of arbuscular mycorrhizal fungi on microbial activity and nutrient release are sensitive to acid deposition during litter decomposition in a subtropical Cinnamomum camphora forest
vol. 16, pp. 314-324 (online: 13 November 2023)
Research Articles
Fungal and bacterial communities in a forest relict of Pinus pseudostrobus var. coatepecensis
vol. 16, pp. 299-306 (online: 09 November 2023)
Short Communications
Culturable fungi associated with wood decay of Picea abies in subalpine forest soils: a field-mesocosm case study
vol. 11, pp. 781-785 (online: 28 November 2018)
Research Articles
Mapping fungi from below ground: online genetic resources and ectomycorrhizal geographic distributions
vol. 4, pp. 252-255 (online: 13 December 2011)
Research Articles
Ectomycorrhizal diversity in a mature pedunculate oak stand near Morović, Serbia
vol. 16, pp. 345-351 (online: 22 November 2023)
Research Articles
Effects of tree species, stand age and land-use change on soil carbon and nitrogen stock rates in northwestern Turkey
vol. 9, pp. 165-170 (online: 18 June 2015)
Research Articles
Stand structure and deadwood amount influences saproxylic fungal biodiversity in Mediterranean mountain unmanaged forests
vol. 9, pp. 115-124 (online: 08 September 2015)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







