
A physiological approach for pre-selection of Eucalyptus clones resistant to drought
iForest - Biogeosciences and Forestry, Volume 13, Issue 1, Pages 16-23 (2020)
doi: https://doi.org/10.3832/ifor3185-012
Published: Jan 15, 2020 - Copyright © 2020 SISEF
Research Articles
Abstract
Water deficit is one of the abiotic stresses that most affects the growth and survival of Eucalyptus. Mechanisms used to tolerate water-limited environments influence the distribution of Eucalyptus species in their natural environment. Here, we take a physiological approach to pre-screen Eucalyptus plants for tolerance to drought. Ten different clones of E. urophylla and E. grandis × E. urophylla that are known to show contrasting responses to water deficit under field conditions, were grown in Clark’s nutrient solution (WW, well-watered) and with polyethylene glycol (-1.0 MPa) to simulate water deficit (WD). Clones responded differently to drought with differentiated photosynthetic limitations in drought-treated clones. Photosynthetic rates, stomatal conductance, transpiration and internal CO2 concentrations were reduced in all genotypes under stress conditions. Clone i144 had a smaller reduction in the evaluated physiological traits, also showing increased root growth in WD-treated plants. Clones 3367 and i224, thought to be moderately tolerant, also followed these patterns. Clones gg157, 1568 and 1641, all of which are moderately sensitive under field conditions, reduced most of the physiological characters evaluated. However, clone gg157 demonstrated increased root system growth, even during short periods of water stress. Clones i042 and i182 were deemed drought-susceptible, with large reductions in photosynthesis and growth, despite showing a high increase in abscisic acid content presumably as a defense mechanism. Interaction between A (photosyntetic rate), E (transpiration rate), ETR/A (electrons transport rate/photosynthetic rate) and SDM/ RDM (shoot dry matter/root dry matter) demonstrated the most significant differences between WD-treated clones and offer great potential for use as selection criterion for water deficit-tolerant genotypes.
Keywords
Clonal Variability, Genotypes, Cluster Analysis, Water Stress
Introduction
Eucalyptus spp. represent 72.4 % (5.7 millions ha) of the total commercial forest area in Brazil ([12]). Rapid development in the Brazil forestry sector and high demand for forest products have led to the expansion of new plantings to areas with limited water supply, resulting in reduced growth and survival rate of trees especially of the most drought-sensitive clones ([35]). An array of mechanisms to tolerate water limited environments exists among Eucalyptus species with the components of aridity thought to be a major differential characteristic among species of the genus ([21], [44]). Therefore, understanding the physiological, biochemical and hormonal responses to drought in various Eucalyptus genotypes is fundamental to select drought-tolerant clones to be cultivated in environments affected by water shortage ([42]).
Several mechanisms may participate in responses to water deficit, including processes that limit dehydration maintaining higher water potential, and/or processes that allow tissues to tolerate lower water potential. In the first case, plants use strategies that minimize water loss through regulation of transpiration via stomatal control ([16]). Photosynthesis is the physiological process most affected by water stress ([38]). Under these circumstances, plants induce hormone-mediated responses, including those governed by abscisic acid (ABA), often also improving water-use efficiency, thereby regulating plant-scale adaptive responses to restrictive conditions ([31]).
Under severe and long-term drought, reduction in net photosynthetic rate may also be the result of biochemical limitations in photosynthetic metabolism, including limitations in phosphorylation ([14]), ribulose-1,5-bisphosphate (RuBP) regeneration ([19]) and RuBisCO carboxylation ([43]). Both processes favor the reduction of carbon assimilation by plants and lead to super-excitation and accumulation of reducing power in the leaves. Protective mechanisms against excess reducing power are important strategies to combat drought-stress; they should therefore also be investigated to obtain a better understanding of drought tolerance by plants.
Drought stress elicits a range of concomitant effects that may limit growth and threaten survival, including photon flux in excess of that required for photoassimilation. To cope with these conditions, protective mechanisms possessed by drought-tolerant plants generally compete with the photochemical phase for absorbed energy, leading to a decrease in PS II quantum yield ([11]) and increasing thermal dissipation ([5]). These findings suggest that thermal dissipation helps to protect against oxidative damage and photoinhibition ([26]). Simultaneous measurement of gas exchange and chlorophyll a fluorescence is therefore an important tool for understanding the relationship between the use-efficiency of light, CO2 assimilation and photoinhibition, in addition to being an important indicator of water deficit in plants ([18]).
Several studies have been conducted to identify the physiological and morphological modifications caused by low water availability in Eucalyptus. Modifications such as reduced photosynthetic rate ([39]), changes in metabolite production ([20], [40]), growth, and productivity ([16], [22]) are the most common outcomes. Although many drought tolerance mechanisms have been studied, forest companies release new, relatively untested clones in each growth season with clone selection constrained by end-user specifications for timber quality. A fast primary screening for water deficit tolerance at the seedling stage assists identification and selection of superior genotypes to be taken to the field. In addition, it is noteworthy that each genotype has its own genetic characteristics and in general, several experiments have been only focused in physiological comparison between two contrasting genotypes. These make it more difficult to evaluate whether the analyses taken were suitable for pre-selection of drought tolerant genotypes.
Recently, early selection has been carried out in several Eucalyptus species. However, few studies have sought to complete a concurrent screen of multiple traits to investigate interactive and correlative properties of candidate characteristics contributing to water deficit tolerance ([29], [4]). Therefore, the purpose of the present study was to evaluate morphological, physiological and non-invasive traits that could potentially discriminate amongst clones for tolerance to water deficit. Using a fast aquaponics methodology on young seedlings, our objective was to provide quantitative physiological markers for initial screening of ten juvenile eucalypt different clones for their tolerance to the effects of water deficit.
Material and methods
Plant material and experimental design
Seedlings of ten Eucalyptus clones (E. grandis × E. urophylla hybrid: 1528, 3367 and VC865; Eucalyptus urophylla hybrid: clones 1641, gg157, i042, i144, i182, i224 and PL40) were used for the experiment. Clones i042 and i144 were previously characterized by the research group as drought-sensitive and -tolerant, respectively, and were used as reference for comparison with other genotypes in this study. Based on the previous observations, categorizations of sensitivity adopted throughout the manuscript are relative (not absolute). Terms adopted throughout this manuscript are therefore used in this context. Clones i042 and i144 were used in this study to ensure that a broad scope of field performance was reflected in the seedlings screened. Plants were grown in a greenhouse in plastic pots (8 L) and were acclimated in Clark nutrient solution ([3]). After 30 days of acclimation, the plants were exposed to two different water treatments: nutrient solution with water potential near 0 MPa (WW, well-watered) and subjected to water deficit (WD), initiated by the gradual addition of polyethylene glycol (PEG) 6000 (Vetec Quimica Fina LTDA, RJ, Brazil) to the nutrient solution. PEG 6000 doses were added every five days to gradually reduce the water potential of the solution (to -0.16, -0.32, -0.65, and -1.0 MPa - [23]). The experiment was arranged in a randomized block, using a 10 × 2 factorial design scheme (genotypes and water availability, respectively), with four biological replications and two plants per pot.
Water potential and gas exchange traits
Five days after application of a PEG 6000 dose to reduce the water potential of the solution to -1 MPa, predawn leaf water potential was measured using a pressure chamber (Scholander, Corvallis, USA) in order to evaluate the effective water deficiency imposed by PEG 6000.
Net photosynthetic assimilation rate (A), stomatal conductance (gS), transpiration rate (E), and the ratio between internal and external CO2 concentration (Ci/Ca) were measured using an infrared gas analyzer (IRGA®, LI-6400xt, LI-COR Bioscience Inc., Lincoln, NE, US). Measurements were performed in fully-expanded leaves, between 8:00 am and 12:30 pm, under constant photosynthetically active radiation (PAR, 1400 μmol photons m-2 s-1), atmospheric CO2 concentration (Ca) of ~372 μmol mol-1, at 22-28 °C and 48-66% relative humidity. Carboxylation efficiency was calculated using A and Ci values (A/Ci) and the intrinsic water use efficiency (WUE) was calculated as the ratio between A and gs.
Chlorophyll a fluorescence
Variables of chlorophyll a fluorescence were measured using a fluorometer coupled to an IRGA (LI-6400xt®, Li-Cor Biosciences, NE, USA). Leaves were initially dark-adapted to obtain the initial fluorescence (F0) and maximal fluorescence (Fm). From this data, the potential quantum yield of photosystem II (PSII; Fv/Fm) was obtained. Leaf tissues were then exposed to actinic light and a saturating pulse to obtain the steady-state fluorescence (F) and the maximum fluorescence in a light-adapted state (Fm′), respectively. This allowed determination the minimum fluorescence of the illuminated plant tissue (F0′), the coefficient photochemical quenching (qL), the effective quantum yield of photochemical energy conversion in PSII (YII), the quantum yield of non-regulated energy (YNPQ) and the apparent electron transport rate (ETR), as detailed by Müller et al. ([25]).
Quantification of abscisic acid (ABA)
ABA was extracted from ground frozen leaf material (100 mg) using methanol (80%, 1 mL) as extraction solvent, according to Durgbanshi et al. ([6]), with modifications. Two hundred microliters of the sample extract had the methanol removed in a SpeedVac at 30 °C and the sample was resuspended in water with the pH corrected to 3.0 (100 μl of 10% v/v acetic acid). Diethyl-ether (500 μl) was added to allow phase separation. The organic phase was collected in a tube and the extract was washed again in diethyl-ether, repeating the process. The extract was dried using a heat block and the samples dissolved in LC-MS buffer for analysis. Leaf extracts containing ABA were analyzed using a Triple Quadrupole LC-MS 6430® (Agilent Technologies, Santa Clara, CA, USA) with the following settings: column (Agilent Eclipse Plus®, RRHD, 1.8 µm, 2.1 × 50 mm with guard column), solvents (A: Acetonitrile + 0.1% formic acid, B: Water + 0.1% formic acid), flow (0.3 mL min-1) for 7 min (gradient time/B%: 0/81, 3/50, 4/10, 4.25/10, 4.5/81). Deuterated ABA (ABAd4) standard was added during the extraction process. Detection and quantification of ABA in the samples was made by multiple reaction monitoring (MRM) by means of selecting the transition density of the molecule of interest (ABA 263 → 153; ABAd4 267 → 156), using the following Mass spectrometric parameters: Dwell 200, Fragmentor 60, CE 6, Accelerator voltage 7, Negative polarity. Data were processed using the software MassHunter workstation VB 06.00 (Agilent Technologies).
Chloroplast pigments
Pigments were extracted from leaf discs (0.6 cm2) using dimethyl sulfoxide (5 mL) saturated with calcium carbonate, under dark conditions at 65 °C. After four hours, absorbances were measured at 480, 649.1 at 665.1 nm in a spectrophotometer. Pure DMSO was used as a blank. Absorbance values were used to estimate chlorophyll a (Chla), chlorophyll b (Chlb) and total carotenoids (Carot) as described by Wellburn ([41]). Pigment concentration was expressed on the basis of leaf area.
Natural abundance carbon isotope composition (δ13C)
The isotopic composition was analyzed in fully expanded-leaves for abundance of 13C (δ13C, ‰), with an isotope ratio mass spectrometer (IRMS, Delta V Plus® model, Thermo Scientific, Waltham, MA, USA). The relationship between the stable isotopes of C was calculated by the formula: δ13C = (Rsample/Rstandard)-1, where R is the molar ratio of the heavy (13C) and light isotopes (12C). The isotope discrimination was calculated by (eqn. 1):
where δ13Catm corresponded to 8 ‰ ([7]).
Morphological traits
The height (cm) and stem diameter (mm) of the plants were measured using a ruler and a caliper, respectively. Leaves, stem and roots were wrapped in paper bags, oven-dried in a forced air circulation oven (65 °C) for 72h and weighed to determine the shoot dry matter (SDM), root dry matter (RDM), total dry matter (TDM) and the shoot and root dry matter ratio (SDM/ RDM) and root and total dry matter ratio (RDM/TDM).
Statistical analysis
Data were subjected to factorial ANOVA using Scott-Knott clustering (p <0.01 and p <0.05). The relationships between variables were evaluated using principal component analysis (PCA). The relative data for PCA and hierarchical clustering analysis were obtained from the normalization of the original data set as log of the ratio between the water stress treatment and the control values. Clones were grouped according to the tolerant or sensitive characters by the dendrogram using Ward’s linkage with Bray-Curtis dissimilarity measure. All statistical analysis were developed using the “vegan” package implemented in the R software ver. 3.2.0 (R Development Core Team, AKL, New Zealand).
Results
Water relations and gas exchange traits
A gradual increase in PEG concentration in the hydroponic solutions resulted in lower predawn leaf water potentials (Ψw), suggesting that plants were under water stress. The WW and WD plants showed Ψw an average of -0.26 and -1.35 MPa, respectively (Tab. 1). The net photosynthetic assimilation rate (A) of drought-stressed Eucalyptus plants was affected in all genotypes (p<0.05 - Tab. 1). The largest percentage reductions in A were observed for clones i042 (4.02 μmol m-2 s-1, 82%), gg157 (5.94 μmol m-2 s-1, 75%) and i182 (7.45 μmol m-2 s-1, 63%) under drought stress compared to plants under normal water availability. In clones i144, 3367 and i244, the reduction of A, 31% (12.72 μmol m-2 s-1), 40% (10.97 μmol m-2 s-1) and 48% (9.17 μmol m-2 s-1), respectively, was less pronounced when the plants were subjected to WD treatment, compared to WW plants. Similarly, water deficit caused a reduction in stomatal conductance (gS) and transpiration rate (E). The carboxylation efficiency (A/Ci) was reduced in all clones under WD, except for clone i144 (Tab. 1) and the water use efficiency (WUE) increased in all clones up to three-fold in i042 (110.7 µmol CO2 mol-1 H2O) and i182 (118.8 µmol CO2 mol-1 H2O) in WD plants compared to WW treatment (Tab. 1).
Tab. 1 - Predawn water potential (Ψw, -MPa); net photosynthetic assimilation rate (A, µmol CO2 m-2 s-1); stomatal conductance (gS, mol H2O m-2 s-1); transpiration rate (E, mmol H2O m-2 s-1), carboxylation efficiency (A/Ci) and intrinsic water use efficiency (WUE, µmol CO2 mol-1 H2O) in Eucalyptus clones grown in well-watered (WW) or water deficit (WD) conditions. Data are presented as the mean ± SE (n = 4). Capital letters compare different clones under well-watered (WW) treatment, and lower letters compare different clones under water deficit (WD) treatment using the Scott-Knott clustering (p <0.05). Asterisk compares WW and WD treatments within each clone. (**): p<0.01; (*): p<0.05.
| Clone | Treat | Ψw | A | g S | E | A/Ci | WUE |
|---|---|---|---|---|---|---|---|
| 1528 | WW | 0.26 ± 0.02 A | 18.96 ± 2.03 A | 0.379 ± 0.06 B | 5.08 ± 0.77 B | 0.071 ± 0.005 A | 52.7 ± 5.3 A |
| WD | 1.58 ± 0.13 a** | 6.36 ± 0.88 b** | 0.063 ± 0.01 a** | 1.28 ± 0.20 a* | 0.032 ± 0.004 b | 100.8 ± 8.3 a** | |
| 1641 | WW | 0.20 ± 0.04 B | 21.27 ± 1.46 A | 0.354 ± 0.08 B | 4.90 ± 0.73 B | 0.087 ± 0.012 A | 81.6 ± 33.9 A |
| WD | 1.25 ± 0.09 c** | 9.29 ± 0.85 a** | 0.085 ± 0.01 a** | 1.59 ± 0.08 a** | 0.050 ± 0.006 a | 109.1 ± 4.1 a | |
| 367 | WW | 0.25 ± 0.08 A | 18.40 ± 0.66 A | 0.384 ± 0.06 B | 4.97 ± 0.53 B | 0.069 ± 0.006 A | 51.7 ± 8.3 A |
| WD | 1.38 ± 0.22 c** | 10.97 ± 1.80 b** | 0.120 ± 0.02 a** | 2.14 ± 0.24 a** | 0.051 ± 0.007 a | 91.2 ± 1.6 a** | |
| gg157 | WW | 0.15 ± 0.07 B | 23.61 ± 0.79 A | 0.502 ± 0.04 A | 5.86 ± 0.27 A | 0.087 ± 0.002 A | 47.7 ± 3.3 A |
| WD | 0.96 ± 0.16 b** | 5.94 ± 1.00 b** | 0.063 ± 0.01 a** | 1.23 ± 0.13 a** | 0.030 ± 0.008 b | 92.5 ± 9.1 a** | |
| i042 | WW | 0.18 ± 0.09 A | 21.36 ± 1.27 A | 0.543 ± 0.03 A | 6.25 ± 0.39 A | 0.076 ± 0.007 A | 39.7 ± 2.8 A |
| WD | 1.59 ± 0.07 c** | 4.02 ± 0.37 b** | 0.036 ± 0.00 a** | 0.75 ± 0.06 a** | 0.023 ± 0.005 b | 110.7 ± 11.7 a** | |
| i144 | WW | 0.30 ± 0.12 A | 18.39 ± 0.53 A | 0.506 ± 0.01 A | 6.58 ± 0.28 A | 0.062 ± 0.002 A | 36.4 ± 1.3 A |
| WD | 1.27 ± 0.10 b** | 12.72 ± 2.15 a** | 0.140 ± 0.03 a** | 2.81 ± 0.52 a** | 0.063 ± 0.010 a | 93.4 ± 5.0 a** | |
| i182 | WW | 0.44 ± 0.06 B | 20.16 ± 0.85 A | 0.546 ± 0.03 A | 6.38 ± 0.43 A | 0.070 ± 0.004 A | 37.4 ± 3.06 A |
| WD | 1.53 ± 0.14 c** | 7.45 ± 0.90 b** | 0.067 ± 0.01 a** | 1.34 ± 0.12 a** | 0.043 ± 0.007 a | 111.8 ± 11.5 a** | |
| I224 | WW | 0.30 ± 0.06 A | 17.75 ± 2.03 A | 0.376 ± 0.08 B | 4.55 ± 0.78 B | 0.066 ± 0.005 A | 56.7 ± 14.5 A |
| WD | 1.23 ± 0.03 c* | 9.17 ± 1.04 a** | 0.102 ± 0.01 a** | 2.02 ± 0.30 a** | 0.044 ± 0.007 a | 90.7 ± 6.9 a** | |
| PL40 | WW | 0.36 ± 0.04 B | 19.80 ± 1.06 A | 0.360 ± 0.07 B | 4.88 ± 0.64 B | 0.078 ± 0.003 A | 59.8 ± 8.5 A |
| WD | 1.36 ± 0.12 c** | 8.28 ± 1.09 b** | 0.071 ± 0.01 a** | 1.28 ± 0.14 a** | 0.046 ± 0.005 a | 117.1 ± 4.0 a** | |
| VC865 | WW | 0.20 ± 0.07 B | 19.49 ± 1.4 A | 0.470 ± 0.07 A | 5.81 ± 0.52 A | 0.071 ± 0.006 A | 44.5 ± 7.4 A |
| WD | 1.40 ± 0.12 c** | 8.01 ± 0.59 b** | 0.077 ± 0.01 a** | 1.39 ± 0.06 a** | 0.041 ± 0.003 a | 106.0 ± 5.6 a** |
Chlorophyll a fluorescence
Chlorophyll a fluorescence was affected by water stress and genotype (p <0.05). Initial chlorophyll a fluorescence (F0 - data not shown) and the potential quantum efficiency of PSII (Fv/Fm) showed minor variations in WD plants (Tab. 2). Responses in the electron transport rate (ETR - data not shown) and the ratio ETR/A were reduced dramatically by the onset of water deficit, with the largest reduction for clones i042 (77%, 0.04) and i182 (60%, 0.006) and the lowest for clone i144 (28%, 0.010) compared to WW plants (p <0.05 - Tab. 2). The estimate of PSII open centers (qL) increased significantly (p <0.05) in clone i144 (45%, 0.08) and decreased in i042 (42%, 0.049) compared to well-watered plants (Tab. 2). The quantum yield of non-regulated energy (YNPQ) of WD plants, increased around 25% for the clones gg157 (0.55), i042 (0.59), 1528 (0.62), 1641 (0.59) and i182 (0.58), compared to WW plants (p <0.05 - Tab. 2).
Tab. 2 - Potential quantum yield of PSII (Fv/Fm), the ratio between electron transport rate and photosynthesis (ETR/A), coefficient photochemical quenching (qL), effective quantum yield of photochemical energy conversion in PSII (YII) and the quantum yield of non-regulated energy (YNPQ) in Eucalyptus clones grown in well-watered (WW) or water deficit (WD) conditions. Data are presented as the mean ± SE (n = 4). Capital letters compare different clones under well-watered (WW) treatment, and lower letters compare different clones under water deficit (WD) treatment using the Scott-Knott clustering (p <0.05). Asterisk compares WW and WD treatments within each clone. (**): p< 0.01; (*): p<0.05.
| Clone | Treat | Fv/Fm | ETR/A | qL | YII | YNPQ |
|---|---|---|---|---|---|---|
| 1528 | WW | 0.83 ± 0.001 B | 0.015 ± 0.002 A | 0.087 ± 0.013 A | 0.149 ± 0.012 A | 0.49 ± 0.02 A |
| WD | 0.83 ± 0.001 a | 0.006 ± 0.001 a** | 0.078 ± 0.008 a | 0.107 ± 0.009 b** | 0.62 ± 0.01 a** | |
| 1641 | WW | 0.84 ± 0.002 A | 0.009 ± 0.002 B | 0.051 ± 0.007 B | 0.106 ± 0.009 B | 0.48 ± 0.04 A |
| WD | 0.84 ± 0.003 a | 0.005 ± 0.000 a** | 0.069 ± 0.007 a | 0.107 ± 0.010 b | 0.59 ± 0.01 a* | |
| 3367 | WW | 0.83 ± 0.007 B | 0.012 ± 0.002 B | 0.073 ± 0.009 A | 0.121 ± 0.011 B | 0.54 ± 0.01 A |
| WD | 0.83 ± 0.002 a | 0.006 ± 0.000 a** | 0.063 ± 0.002 a | 0.110 ± 0.005 b | 0.54 ± 0.03 a | |
| gg157 | WW | 0.84 ± 0.003 A | 0.013 ± 0.001 A | 0.060 ± 0.006 B | 0.136 ± 0.005 A | 0.42 ± 0.02 A |
| WD | 0.83 ± 0.004 a* | 0.005 ± 0.001 a** | 0.054 ± 0.011 a | 0.082 ± 0.014 b** | 0.55 ± 0.03 a** | |
| i042 | WW | 0.83 ± 0.002 B | 0.018 ± 0.001 A | 0.085 ± 0.008 A | 0.159 ± 0.014 A | 0.46 ± 0.03 A |
| WD | 0.83 ± 0.004 a | 0.004 ± 0.000 a** | 0.049 ± 0.001 a** | 0.076 ± 0.003 b** | 0.59 ± 0.02 a** | |
| i144 | WW | 0.83 ± 0.004 B | 0.014 ± 0.001 A | 0.044 ± 0.003 B | 0.096 ± 0.005 B | 0.44 ± 0.03 A |
| WD | 0.84 ± 0.001 a | 0.010 ± 0.001 a** | 0.080 ± 0.005 a** | 0.153 ± 0.020 a** | 0.47 ± 0.07 a | |
| i182 | WW | 0.83 ± 0.002 B | 0.015 ± 0.001 A | 0.062 ± 0.007 B | 0.116 ± 0.010 B | 0.48 ± 0.04 A |
| WD | 0.83 ± 0.001 a | 0.006 ± 0.002 a** | 0.075 ± 0.005 a | 0.113 ± 0.007 b | 0.58 ± 0.02 a* | |
| i224 | WW | 0.83 ± 0.003 B | 0.011 ± 0.002 B | 0.062 ± 0.010 B | 0.121 ± 0.010 B | 0.45 ± 0.04 A |
| WD | 0.83 ± 0.004 a | 0.007 ± 0.002 a* | 0.057 ± 0.007 a | 0.104 ± 0.017 b | 0.53 ± 0.03 a | |
| PL40 | WW | 0.84 ± 0.001 A | 0.011 ± 0.002 B | 0.058 ± 0.007 B | 0.118 ± 0.008 B | 0.48 ± 0.05 A |
| WD | 0.83 ± 0.001 a | 0.004 ± 0.001 a** | 0.059 ± 0.006 a | 0.107 ± 0.012 b | 0.52 ± 0.03 a | |
| VC865 | WW | 0.83 ± 0.003 B | 0.014 ± 0.002 A | 0.072 ± 0.007 A | 0.117 ± 0.012 B | 0.55 ± 0.02 A |
| WD | 0.83 ± 0.007 a | 0.005 ± 0.001 a** | 0.067 ± 0.014 a | 0.100 ± 0.013 b | 0.56 ± 0.02 a |
ABA, pigment content and isotopic discrimination
Water deficit increased ABA levels in the leaves of plants subjected to drought-stress, especially in clones i224 (153%, 18 pmol mg-1 ), i144 (76%, 10.7 pmol mg-1) and i042 (74%, 8.1 pmol mg-1), compared to WW plants (p <0.05). Similarly, chlorophyll a concentration increased in most of the clones, especially in i182 (37%, 48.7 μg cm-2) and i144 (35%, 50.5 μg cm-2) when subjected to WD (p <0.05 - Tab. 3). The greatest increment of chlorophyll b concentration in WD plants was observed in clone i144 (39%, 17.4 μg cm-2), i182 (28%, 15.7 μg cm-2) and 1641 (28%, 18.5 μg cm-2). Significant increases in the total carotenoid concentrations was observed in clones i182 (9.89 μg cm-2), vc865 (10.52 μg cm-2) and 1528 (11.05 μg cm-2), on average, 25% in plants subjected to WD compared to WW (p <0.05 - Tab. 3). The carbon isotope discrimination (Δ13C) was significantly reduced in clone i042 (17.99‰) and i182 (18.57‰) subjected to WD, in 14% when compared to WW plants (p <0.05). No differences were found in carbon isotope discrimination in control plants of clones 1528, 1641, i144, i224, and PL040 compared to the drought-stressed plants.
Tab. 3 - Abscisic acid content (ABA, pmol mg-1 FW), chlorophyll a (Chl a, μg cm-2), chlorophyll b (Chl b, μg cm-2), total carotenoids (Carot, μg cm-2) and isotopic discrimination (Δ13C, ‰) in Eucalyptus clones grown in well-watered (WW) or water deficit (WD) conditions. Data are presented as the mean ± SE (n = 4). Capital letters compare different clones under well-watered (WW) treatment, and lower letters compare different clones under water deficit (WD) treatment using the Scott-Knott clustering (p <0.05). Asterisk compares WW and WD treatments within each clone. (**): p< 0.01; (*): p<0.05.
| Clone | Treat | ABA | Chl a | Chl b | Carot | Δ13C |
|---|---|---|---|---|---|---|
| 1528 | WW | 6.2 ± 1.0 C | 44.1 ± 2.4 A | 15.5 ± 0.58 A | 10.05 ± 0.50 A | 19.19 ± 1.06 B |
| WD | 8.7 ± 0.6 d | 51.8 ± 4.8 a* | 19.7 ± 2.62 a** | 11.04 ± 0.63 a | 17.69 ± 0.64 b | |
| 1641 | WW | 13.3 ± 1.1 A | 42.0 ± 1.8 A | 14.4 ± 0.81 A | 8.16 ± 0.92 A | 17.64 ± 0.97 B |
| WD | 13.7 ± 0.5 b | 52.8 ± 1.3 a** | 18.5 ± 0.90 a** | 10.07 ± 0.92 a* | 17.80 ± 0.34 b | |
| 3367 | WW | 10.9 ± 1.6 B | 39.3 ± 4.5 A | 13.0 ± 1.17 A | 8.62 ± 0.80 A | 19.58 ± 0.75 A |
| WD | 11.3 ± 1.8 c | 48.5 ± 2.7 a* | 16.6 ± 1.02 b** | 9.93 ± 0.63 a | 17.52 ± 0.24 b* | |
| gg157 | WW | 9.0 ± 0.2 B | 41.0 ± 2.2 A | 13.1 ± 0.63 A | 9.11 ± 0.50 A | 20.16 ± 0.30 A |
| WD | 15.7 ± 2.3 b** | 46.1 ± 2.9 a | 15.0 ± 0.86 b | 9.68 ± 0.44 a | 18.41 ± 0.40 b* | |
| i042 | WW | 4.7 ± 0.5 C | 39.3 ± 0.9 A | 13.6 ± 0.36 A | 8.92 ± 0.26 A | 20.85 ± 0.43 A |
| WD | 8.1 ± 1.5 d* | 49.6 ± 2.7 a** | 16.9 ± 0.86 b* | 10.51 ± 0.93 a | 17.99 ± 0.45 b** | |
| i144 | WW | 5.4 ± 1.1 C | 37.5 ± 0.6 A | 12.5 ± 0.22 A | 8.41 ± 0.22 A | 20.72 ± 0.50 A |
| WD | 10.7 ± 0.6 c** | 50.5 ± 1.5 a** | 17.4 ± 0.36 a** | 10.22 ± 0.64 a | 20.17 ± 0.04 a | |
| i182 | WW | 5.8 ± 0.8 C | 35.5 ± 1.1 A | 12.2 ± 0.09 A | 7.57 ± 0.36 A | 21.47 ± 0.24 A |
| WD | 7.7 ± 0.9 d | 48.7 ± 0.9 a | 15.7 ± 0.47 b* | 9.89 ± 0.27 a* | 18.57 ± 0.19 b** | |
| i224 | WW | 6.2 ± 1.2 C | 45.8 ± 3.9 A | 15.6 ± 1.31 A | 10.01 ± 0.73 A | 19.49 ± 0.60 B |
| WD | 18.0 ± 1.0 a** | 49.6 ± 5.1 a* | 17.6 ± 0.89 a | 10.17 ± 1.44 a | 18.07 ± 0.48 b | |
| PL40 | WW | 6.8 ± 0.3 C | 40.4 ± 0.4 A | 14.0 ± 0.45 A | 8.76 ± 0.21 A | 19.12 ± 0.28 B |
| WD | 10.2 ± 0.7 c* | 48.1 ± 0.8 a* | 16.2 ± 0.72 b | 10.29 ± 0.47 a | 17.50 ± 0.57 b | |
| VC865 | WW | 7.4 ± 1.1 C | 38.1 ± 2.3 A | 12.6 ± 0.62 A | 8.51 ± 0.36 A | 19.47 ± 1.17 B |
| WD | 12.0 ± 0.2 c** | 46.5 ± 1.3 a* | 15.0 ± 0.69 b | 10.52 ± 0.25 a* | 17.48 ± 0.51 b* |
Growth and SDM/RDM ratio
We observed for the clone PL40 a greater reduction in height (H - p <0.05), shoot dry matter (SDM - p <0.05) and total dry matter (TDM - p <0.05) in WD as compared to WW, with TDM reductions up to 64% (5.91 g plant-1), followed by 3367 (54%, 5.97 cm plant-1) and i042 (48%, 6.13 cm plant-1 - Tab. 4). The lowest reduction in SDM was observed in clone i224 (5.51 cm plant-1, 32%) compared to WW plants (Tab. 4). Root dry matter (RDM) was not affected by WD conditions in clones 1528 and gg157. However, the lowest ratios of reduction in shoot to root dry matter (SDM/RDM - data not shown) and of the root to total dry matter ratio (RDM/TDM) were found for clone 3367 (13.2 and 8.6%, respectively) under water-deficit conditions (p <0.05 - Tab. 4). No significant changes in the TDM of plants of clones 1528, and i224 were detected under WD (Tab. 4), and the lowest reduction was found by i144 (5.01 cm plant-1; 33%), a drought-tolerant genotype (p <0.05).
Tab. 4 - Height (cm plant-1), stem diameter (mm plant-1), shoot dry matter (SDM, g plant-1), root dry matter (RDM, g plant-1), total dry matter (TDM, g plant-1) and the ratio RDM/TDM in Eucalyptus clones grown in well-watered (WW) or water deficit (WD) conditions. Data are presented as the mean ± SE (n = 4). Capital letters compare different clones under well-watered (WW) treatment, and lower letters compare different clones under water deficit (WD) treatment using the Scott-Knott clustering (p <0.05). Asterisk compares WW and WD treatments within each clone. (**): p< 0.01; (*): p<0.05.
| Clone | Treat | Height | Stem diameter | SDM | RDM | TDM | RDM/TDM |
|---|---|---|---|---|---|---|---|
| 1528 | WW | 39.38 ± 4.17 C | 5.77 ± 0.48 A | 6.46 ± 0.90 B | 2.48 ± 0.18 A | 8.94 ± 1.07 B | 0.28 ± 0.02 A |
| WD | 29.25 ± 3.85 c* | 4.42 ± 0.55 a** | 4.29 ± 0.39 b | 2.67 ± 0.20 a | 6.96 ± 0.58 b | 0.38 ± 0.01 a** | |
| 1641 | WW | 60.63 ± 1.71 A | 6.12 ± 0.45 A | 12.73 ± 1.42 A | 3.56 ± 0.55 A | 16.29 ± 1.93 A | 0.22 ± 0.01 C |
| WD | 49.31 ± 1.37 a* | 4.38 ± 0.19 a** | 6.52 ± 0.72 a** | 2.40 ± 0.21 a* | 8.93 ± 0.90 a** | 0.27 ± 0.01 b** | |
| 3367 | WW | 48.00 ± 5.05 B | 5.31 ± 0.16 A | 8.44 ± 0.71 B | 2.59 ± 0.30 A | 11.03 ± 0.83 B | 0.24 ± 0.02 B |
| WD | 37.25 ± 4.54 c* | 3.80 ± 0.16 a** | 4.46 ± 0.43 b* | 1.51 ± 0.05 b* | 5.97 ± 0.47 b* | 0.26 ± 0.01 b | |
| gg157 | WW | 42.50 ± 1.17 C | 5.04 ± 0.23 A | 6.42 ± 0.35 B | 1.55 ± 0.13 B | 7.96 ± 0.48 B | 0.19 ± 0.01 C |
| WD | 33.31 ± 1.49 c* | 3.87 ± 0.25 a* | 3.18 ± 0.25 b* | 1.67 ± 0.12 b | 4.84 ± 0.33 b | 0.35 ± 0.02 a** | |
| i042 | WW | 58.00 ± 3.92 A | 5.65 ± 0.41 A | 9.09 ± 1.25 B | 2.69 ± 0.23 A | 11.78 ± 1.45 B | 0.23 ± 0.01 B |
| WD | 42.81 ± 6.55 b** | 4.69 ± 0.29 a | 4.30 ± 0.78 b** | 1.83 ± 0.32 b | 6.13 ± 1.08 b** | 0.30 ± 0.02 b** | |
| i144 | WW | 39.00 ± 1.21 C | 4.69 ± 0.24 A | 5.66 ± 0.32 B | 1.87 ± 0.05 B | 7.53 ± 0.33 B | 0.25 ± 0.01 B |
| WD | 30.42 ± 1.72 c* | 4.01 ± 0.51 a | 3.36 ± 0.77 b | 1.65 ± 0.24 b | 5.01 ± 1.02 b | 0.34 ± 0.02 a** | |
| i182 | WW | 48.69 ± 1.57 B | 5.94 ± 0.23 A | 11.73 ± 1.04 A | 2.72 ± 0.19 A | 14.45 ± 1.23 A | 0.19 ± 0.01 C |
| WD | 37.19 ± 2.37 c* | 4.44 ± 0.06 a** | 5.97 ± 0.79 a** | 2.33 ± 0.34 a | 8.30 ± 1.09 a** | 0.28 ± 0.01 b** | |
| i224 | WW | 48.63 ± 3.31 B | 5.48 ± 0.28 A | 8.04 ± 1.58 B | 2.49 ± 0.46 A | 10.54 ± 2.01 B | 0.24 ± 0.01 B |
| WD | 37.69 ± 0.95 c* | 4.55 ± 0.30 a | 5.51 ± 0.87 b | 2.24 ± 0.48 a | 7.75 ± 1.32 b | 0.28 ± 0.02 b** | |
| PL40 | WW | 52.75 ± 3.55 A | 5.24 ± 0.74 A | 13.33 ± 2.57 A | 3.37 ± 0.63 A | 16.70 ± 3.19 A | 0.20 ± 0.01 C |
| WD | 43.13 ± 4.71 b* | 4.06 ± 0.26 a* | 4.26 ± 0.44 b** | 1.65 ± 0.26 b** | 5.91 ± 0.69 b** | 0.28 ± 0.02 b** | |
| VC865 | WW | 54.38 ± 1.43 A | 5.83 ± 0.39 A | 11.79 ± 2.52 A | 2.81 ± 0.42 A | 14.60 ± 2.93 A | 0.20 ± 0.01 C |
| WD | 54.50 ± 2.25 a | 5.51 ± 0.21 a | 9.12 ± 0.78 a | 3.36 ± 0.35 a | 12.48 ± 1.03 a | 0.27 ± 0.02 b** |
PCA and hierarchical clustering
Four principal components (PCs), based on relative values, explained 79% of the total variation in the analyzed data, and the first two PCs explained 33% and 19%, respectively (Fig. 1). The first PC was characterized by high positive scores for gas exchange traits (A/Ci, A, ETR/A, ETR, E, gS and YII) and negatively associated with root characteristics (RDM, RDM/TDM), Ψw and YII (Fig. 1). The second PC had a strong positive correlation with pigments and WUE and negative correlation with Ci/Ca. Other variables such as ABA, SD, Δ13C had minor effects on the first two PCs evaluated (Fig. 1).
Fig. 1 - Principal Component Analysis (PCA). Biplot between PC1 and PC2 showing the contribution of 29 physiological and morphological traits in variability and the segregation of the 10 Eucalyptus clones under drought stress conditions.
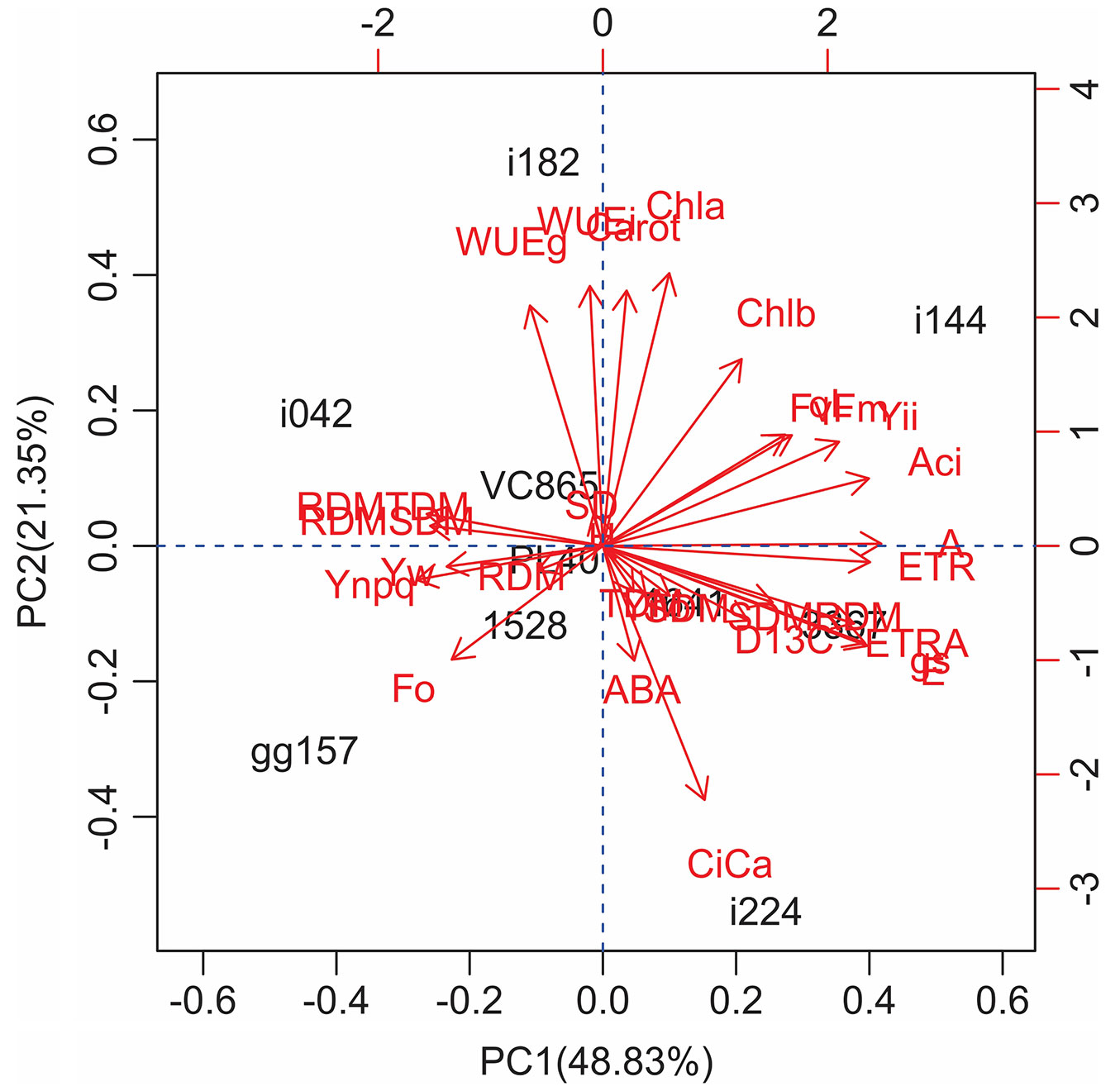
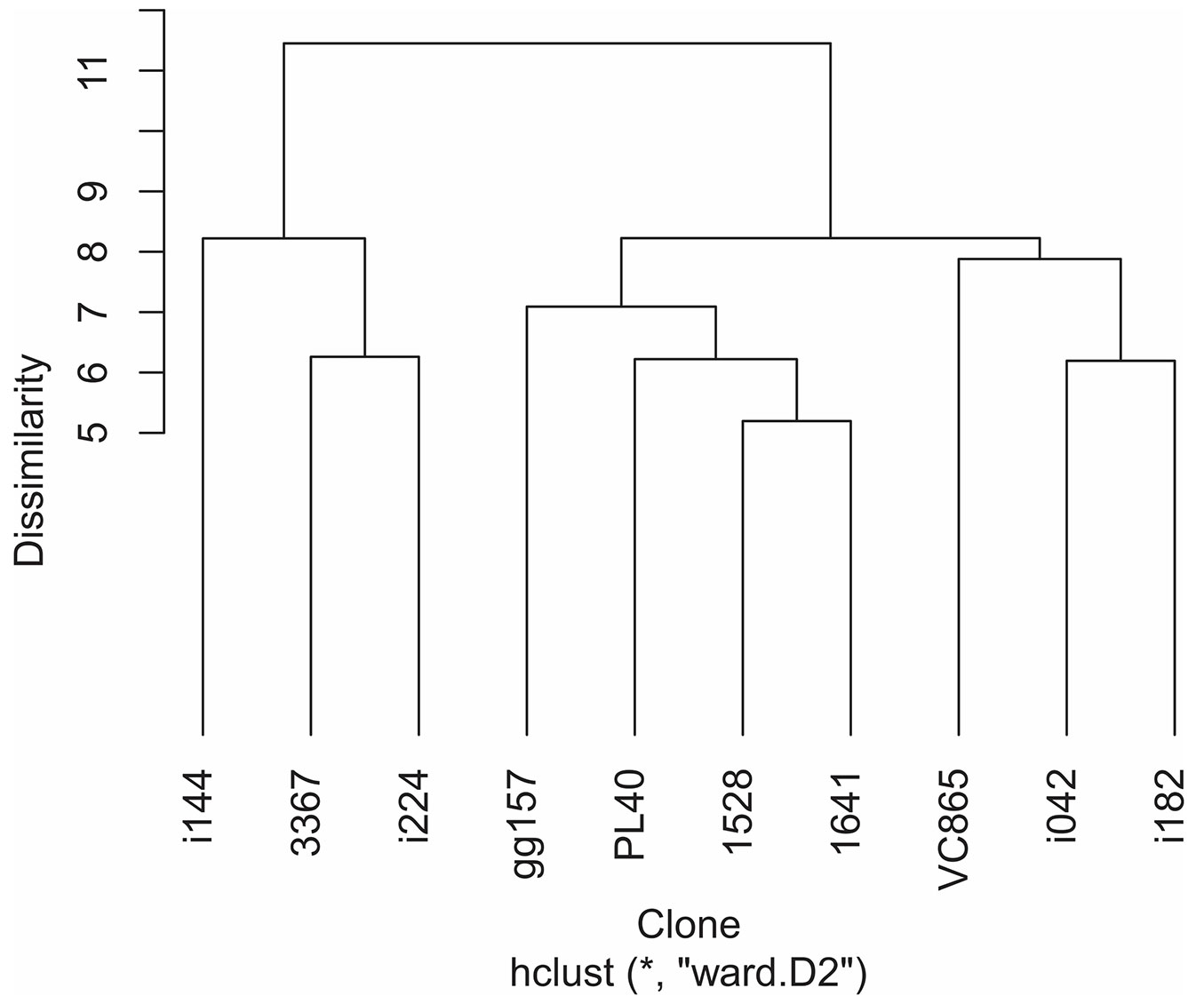
Analyzing all physiological and morphological evaluated traits among various genotypes and the dendrogram using Ward’s linkage allowed to categorize the Eucalyptus clones into four groups based on the relative magnitude of the measured responses to treatment: (i) Clone i144 as resistant and (ii) 3367 and i224 as moderately resistant to the effects of water deficit. All were characterized by the lowest reduction in gas exchange traits, indicating continued carbon gain (Fig. 2). Clone i144 also demonstrated no change in A/Ci and showed the highest ABA levels under drought. The other two groups characterized as follows: (iii) clones gg157, pl40, 1528 and 1641, moderately-sensitive, where 1528 was not affected in RDM, gg157 showed the highest increases in RDM and YNPQ variables; and (iv) vc865, i182 and i042, characterized as sensitive clones to drought stress. Clones vc865 and i182 invested in shoot growth, to the detriment of the root system. Clone i182 had an increment of WUE and reduced Δ13C; however, these alterations were not reflected in drought-tolerance during water stress.
Fig. 2 - Hierarchical clustering analysis. Dendrogram of 10 Eucalyptus clones using Ward’s linkage with Bray-Curtis dissimilarity measure. Data consists in relative values of 29 physiological and morphological traits in eucalypt plants subjected to drought stress conditions.
Discussion
Drought-tolerant genotypes had lower negative effects on gas exchange and carbon isotope composition during water stress
Photosynthesis is one of the primary processes initially affected by water restriction in plants ([2]), mainly in isohydric species ([36]). The imposition of water deficit clones resulted in reductions in A, Ci, E and gS that were more pronounced in clones i042 (sensitive), gg157 and i182, considered within the context of our study as moderately sensitive or sensitive to drought stress.
The reduction in stomatal conductance observed here may be partially mediated by higher ABA concentrations ([31]) and osmotic adjustment ([20], [1]), leading to increased water-use efficiency ([28]) and avoiding unnecessary water loss. Clones i042, i144 and i224 contained higher ABA levels in the leaves and greatly reduced stomatal conductance; nevertheless, no significant correlation was obtained between ABA and gS (data not shown). For genotypes 1528, gg157, 3367 and i182 however, there were no differences in ABA leaf concentrations; however, stomatal conductance was greatly reduced under drought. Taken together, these results suggest that stomatal closure in these genotypes is not mediated by ABA. Alternatively, ABA regulation of stomatal aperture may be masked by rapid metabolism during the first stages of drought, therefore it does not accumulate in the leaves ([13]). Some studies have shown the relationship of higher ABA accumulation and drought tolerance in Eucalyptus genotypes ([37], [17]). In this experiment, clone i144 had an approximate 40% increase in ABA concentration, which possibly contributed to the increase (~156%) in water use efficiency (WUE - Tab. 1), confirming the drought-tolerant characteristics of this genotype.
Increase in water-use efficiency is thought to be an important property that contributes to the resilience and growth of genotypes under drought conditions ([15]). Our results suggest that while significant variability exists in the parameters measured in this study, it is clear that factors governing leaf gas exchange did not delineate the genotypes under the various treatment conditions. Surrogate measures of leaf gas exchange, including carbon isotope abundance, may offer significant insight into the tolerance of individual genotypes to the effects of water deficit ([16], [32]). Nevertheless, the tolerant genotype (i144), and other clones considered to be moderately sensitive genotypes (1528, 1641, i224, and PL040) did not show differences in leaf Δ13C, suggesting an absence of pattern to this trait to drought tolerance selection in early stages of Eucalyptus growth. One possible explanation for this may be that carbon acquisition was not of a sufficient magnitude under the stress conditions; therefore, a change in isotope abundance was not observed. This represents a significant limitation in the use of Δ13C as an indicator of plant stress tolerance and, at the very least, needs to be considered in the context of net carbon assimilation.
Non-stomatal limitations to photosynthesis were observed among Eucalyptus clones
Differential limitations to photosynthesis were observed among clones under the effects of water deficit. In long-term and moderate stress, the reduction of gS can induce a decrease in CO2 concentration at the carboxylation sites of RuBisCO ([8], [9]), with consequent stomatal limitation that reduces photosynthetic rates. However, in this study, internal CO2 concentrations (Ci) and the corresponding A/Ci were not proportionately reduced for most clones (1528, gg157, i042, i182 - moderately-sensitive or sensitive to drought), suggesting that, under the conditions tested here, changes or damage to photosynthetic metabolism ([14]) resulted in non-stomatal limitation of A.
RUBP regeneration capacity, a non-stomatal limitation, is limited by drought, possibly due to reduction in the electron transport rate and, consequently, in NADPH ([8]). These authors found that the decrease in photochemical efficiency was evidenced by a high reduction in the electron transport rate (ETR), accompanied by a reduction in photochemical quenching (qL), as observed in the present study, especially in the drought-sensitive clone i042 under water-deficit conditions. Decreases in A and ETR are generally proportional, suggesting a strong link between the photosynthetic processes ([10]). Alternatively, a decrease in A and maintenance of ETR, leading to an increased ETR/A ratio, suggest that water deficit causes no inhibition of photochemical reactions ([34]). In the present study, for almost all genotypes, the ETR/A and A/Ci ratios decreased, especially for clones i042 and i182, characterized here as sensitive to water deficit. These responses suggest that the reduction in the electron transport rate was as high as the drop in the photosynthetic rate during water deficit. Although the possible biological limitations were not measured in this study, drastic changes in the above-mentioned processes suggest predominance of non-stomatal limitations in these genotypes.
Conversely, in the drought-tolerant genotype (i144), slight reductions in ETR and increases in qL, associated with unchanged A/Ci and ETR/A ratios, suggest a decrease in A by stomatal limitations. Clones 3367 and i224 were similar to i144 in terms of these characteristics and were characterized as moderately tolerant genotypes. The results suggest that these genotypes have a greater potential for post-stress recovery, because the photosynthetic apparatus remains functional during water-deficit periods. Alternatively, higher photosynthetic rates during the early drought stages also increase plant survival and dry matter accumulation ([28]); in genotypes with higher A under water-deficit conditions, photoinhibition of photosynthesis is reduced ([34]).
Excess energy dissipation may also result from the accumulation of reducing power (NADPH) and ATP produced in the photochemical phase. This is subsequently released in the form of thermal energy, measured by NPQ. The dissipation of absorbed energy through thermal dissipation (NPQ) increased most in clones 1528 and i182. In these genotypes, the thermal dissipation may have been important to avoid photoinhibition. This is because dissipation of excess energy, either photochemically or non-photochemically (e.g., thermal dissipation) controls excess excitation energy and electron fluxes to O2, that can prevent photooxidative damage ([26]) and consequently photoinhibition ([30]).
Factors governing gas exchange and chlorophyll a fluorescence undoubtedly played a major role in the stress response of individual clones, highlighting the need for integrated measures of plant scale physiological measurements to be part of the pre-breeding toolbox. We also showed that non-invasive parameters reflecting non-stomatal factors such as ETR/A may be used as part of the suite of tools for assessment of performance under drought conditions.
Genotype growth and classification
Clone i144, considered drought-tolerant in the field and confirmed in this study as tolerant during seedling stage and greenhouse controlled-conditions, performed well compared to other clones under water stress. This performance may have been the result of minor changes in A, gS, Ci/Ca, and minor damage to the photosynthetic apparatus (lower ETR/A) as well as the lower SDM/RDM ratio. The lower SDM/ RDM ratio indicates expansion of the root system at the expense of shoot growth that allows for better recovery after water deficit ([24]). To increase water uptake, many plants increase root growth, laterally or in terms of depth. In Eucalyptus species, the difference in drought tolerance was attributed to differences in root depth ([24]) and the hydraulic conductivity of the root system ([33]). Therefore, the above-mentioned characteristics may have led to a greater increase in WUE and continued growth despite the effects of water deficit. The performance of the clones 3367 and i224 was similar to that of the tolerant clone (i144) in terms of most characteristics evaluated, and were characterized as moderately tolerant to drought.
The largest reductions in growth and photosynthetic rate observed for clone i042 reinforce the classification of its susceptibility to water deficit and drought under field conditions ([27]). It is noteworthy that although clones i042, vc865 and i182 were in closed groups by multivariate analysis, the last genotype was moderately drought-tolerant in the field. However, this potential was not expressed under greenhouse conditions in this study. According to the distribution of the dendrograms and univariate analysis of studied traits, the genotypes i224 and 3367 were classified as moderately drought-tolerant, and other genotypes were classified as moderately sensitive.
Conclusions
Assessing drought tolerance under field conditions faces significant challenges because of the scale and accessibility of leaf and root tissues, canopy and soil heterogeneity and the dynamics of soil/water/ plant interactions. Developing selection traits at the seedling stage prior to investment in field planting allows selection to take place early in the developmental cycle as well as providing a relatively consistent imposition of stress to standardize the severity of stress conditions. For the present study, it is important to emphasize that we were not able to identify a “stand-alone” variable characterizing drought-tolerance in the Eucalyptus clones tested. It is far more likely that a combination of such tools, including A, ETR/A and SDM/RDM, will yield the most effective form of pre-breeding assessment, to provide informed analysis of drought-tolerant genotypes.
Factors governing gas exchange undoubtedly played a major role in the stress response of individual clones, highlighting the need for integrated measures of plant scale gas exchange to be part of the pre-breeding toolbox. We also showed that non-invasive parameters reflecting non-stomatal factors such as ETR/A may be used as part of the suite of tools for assessment of performance under drought conditions. The assessment of these traits, under the conditions imposed in this study, highlight the substantial potential for rapid assessment of physiological and chemical parameters to improve plant breeding efforts while recognizing the need to validate these tools under field conditions. Improved insight into such relations will not only inform breeding programs of the mechanistic relationships with stress tolerance, but will also provide candidate tools for use in plantation management under varying environmental and climatic conditions.
List of abbreviations
(A): net photosynthetic assimilation rate; (A/Ci): carboxylation efficiency; (ABA): abscisic acid; (Carot): total carotenoids; (Chla): chlorophyll a; (Chlb): chlorophyll b; (Ci/Ca): ratio between internal and external CO2 concentration; (E): transpiration rate; (ETR): apparent electron transport rate; (Fv/Fm): potential quantum yield of PSII; (gS): stomatal conductance; (PEG): polyethylene glycol; (PSII): photosystem II; (qL): coefficient photochemical quenching; (RDM): root dry matter; (SDM): shoot dry matter; (TDM): total dry matter (TDM); (WD): water deficit; (WUE): water use efficiency; (WW): well-watered; (YII): effective quantum yield of photochemical energy conversion in PSII; (YNPQ): quantum yield of non-regulated energy; (Δ13C): isotope discrimination; (Ψw): predawn water potential.
Acknowledgments
This work was supported by the Minas Gerais State Agency for Research and Development (FAPEMIG), the Coordination of Improvement of Higher Education Personnel (CAPES/PRODOC, grant number 00113/ 010-0) and the National Council for Scientific and Technological Development (CNPq/Universal, grant number 475787/ 2012-9). CM and BERH are grateful to the CAPES, CNPq and Science without borders (CsF) for scholarships. AM acknowledges the Australian Research Council Future Fellowship Scheme (FT120100200).
Author contributions
CM and NFB designed the study; CM conducted the experiments and performed the physiological measurements; BERH performed the carbon isotope composition analysis; CM, BERH and AM wrote the manuscript. All authors discussed the data, read and approved the final version of the manuscript.
Disclosure statement
No potential conflict of interest was reported by the authors.
References
Authors’ Info
Authors’ Affiliation
Bárbara Elias Reis Hodecker 0000-0003-2653-5583
Nairam Félix De Barros
Department of Soil Science, Federal University of Viçosa, Viçosa MG, 36571-000 (Brazil)
Bárbara Elias Reis Hodecker 0000-0003-2653-5583
Andrew Merchant 0000-0001-8979-6786
School of Life and Environmental Sciences, Centre for Carbon, Water and Food, The University of Sydney, Camden NSW, 2570 (Australia)
Corresponding author
Paper Info
Citation
Müller C, Hodecker BER, De Barros NF, Merchant A (2020). A physiological approach for pre-selection of Eucalyptus clones resistant to drought. iForest 13: 16-23. - doi: 10.3832/ifor3185-012
Academic Editor
Claudia Cocozza
Paper history
Received: Jul 04, 2019
Accepted: Nov 21, 2019
First online: Jan 15, 2020
Publication Date: Feb 29, 2020
Publication Time: 1.83 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2020
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 48085
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 38272
Abstract Page Views: 4600
PDF Downloads: 4325
Citation/Reference Downloads: 6
XML Downloads: 882
Web Metrics
Days since publication: 2389
Overall contacts: 48085
Avg. contacts per week: 140.89
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Jul 2026)
Total number of cites (since 2020): 15
Average cites per year: 2.14
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Response of Chinese sea buckthorn clonal growth and photosynthetic physiological mechanisms toward a soil moisture gradient
vol. 14, pp. 337-343 (online: 15 July 2021)
Short Communications
Stomata morphological traits in two different genotypes of Populus nigra L.
vol. 8, pp. 547-551 (online: 16 September 2014)
Research Articles
Two Populus deltoides W.Bartram ex Marshall clones cope differentially with sodium salinity stress
vol. 18, pp. 259-266 (online: 10 October 2025)
Research Articles
Clonal structure and dynamics of peripheral Populus tremula L. populations
vol. 7, pp. 140-149 (online: 13 January 2014)
Research Articles
Links between phenology and ecophysiology in a European beech forest
vol. 8, pp. 438-447 (online: 15 December 2014)
Research Articles
Use of δ13C as water stress indicator and potential silvicultural decision support tool in Pinus radiata stand management in South Africa
vol. 12, pp. 51-60 (online: 24 January 2019)
Research Articles
Tree aging does not affect the ranking for water use efficiency recorded from δ13C in three Populus deltoides × P. nigra genotypes
vol. 12, pp. 272-278 (online: 21 May 2019)
Research Articles
Variability of tolerance of Wild cherry clones to PEG-induced osmotic stress in vitro
vol. 15, pp. 265-272 (online: 25 July 2022)
Research Articles
Contribution of environmental variability and ecosystem functional changes to interannual variability of carbon and water fluxes in a subtropical coniferous plantation
vol. 9, pp. 452-460 (online: 25 January 2016)
Research Articles
Spatial and temporal variation of drought impact on black locust (Robinia pseudoacacia L.) water status and growth
vol. 8, pp. 743-747 (online: 18 June 2015)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







