Tree aging does not affect the ranking for water use efficiency recorded from δ13C in three Populus deltoides × P. nigra genotypes
iForest - Biogeosciences and Forestry, Volume 12, Issue 3, Pages 272-278 (2019)
doi: https://doi.org/10.3832/ifor2896-012
Published: May 21, 2019 - Copyright © 2019 SISEF
Research Articles
Abstract
A large variability of water use efficiency (assessed from the carbon isotopic discrimination in leaves and leaf soluble sugars) has been detected among poplar genotypes. Checking whether such differences detected in young trees (1-2 years old) remain stable with tree age is a prerequisite to use this trait with confidence for breeding purposes. In this study, a synchronic approach was used to test the age-related stability of the genotypic ranking of carbon isotopic discrimination in wood (Δ13C) until tree maturity. We sampled 376 trees between 4 and 20 years from three Populus deltoides × P. nigra genotypes growing in 41 common-garden trials across France. Carbon and nitrogen percentages along with δ13C was measured in the bulk wood of the year 2009 and used to compute the Δ13C. Basal area increment between 2008 and 2009 was also measured. Results showed that Δ13C increased (i.e., water use efficiency decreased) between ages 4 to 6 and remained stable later on. Significant differences among genotypes were found but the ranking among genotypes remained stable with age during the assessed life span. Furthermore, basal area increment and Δ13C were positively correlated interannually. This large-scale survey shows that despite crossing over in the temporal trend, water use efficiency remained stable with age across 3 poplar genotypes. However, further studies with a large number of genotypes are required to confirm whether this trait can be used to maintain or even improve productivity of poplar plantations, while lowering water consumption.
Keywords
Water Use Efficiency, Age, Wood, Tree Ring, Populus × euramericana, Basal Area Increment, Synchronic Approach
Introduction
In the context of climate change and fluctuating precipitation patterns ([20]), water use efficiency (WUE, ratio between biomass accumulated and water transpired) has become an interesting trait for breeding new genotypes ([9], [41], [46]). Isotopic discrimination against 13C (Δ13C) during CO2 diffusion through stomata and photosynthesis is an indirect indicator of intrinsic water use efficiency (iWUE, ratio between net CO2 assimilation rate, A, and stomatal conductance to water vapour, gs - [14], [15], [13]). Indeed, this trait was found to be under tight genetic control in several tree species like poplar, pines, chestnut and oaks among others ([22], [5], [4], [40]). Furthermore, given the lack of correlation observed between WUE and productivity or, in some instances, the positive correlation between both traits ([46]), selecting tree species or genotypes based on their ability to express a high WUE is desirable. Such a strategy has been explored in some crops in a context of improving water management ([9], [41]) and might thus be particularly suitable for sustaining wood production under the incertitude of water availability.
Poplar genotypes are known to display large differences in growth performance and biomass production ([39]). A large genotypic variability of Δ13C was found among Populus deltoides × P. nigra genotypes in controlled environment ([26]) and open field experiments ([29], [10]). Using direct measurements, this variability in Δ13C is also found tightly related to variability in the whole plant water use efficiency in Populus deltoides × P. nigra and in Populus nigra ([39], [40]). Interestingly, no correlation was found between productivity and Δ13C ([29], [10]), giving opportunity to breed for enhanced WUE without compromising productivity in poplar genotypes. Nevertheless, previous studies on poplar genotypes were conducted on seedlings of 1-2 years and there are few studies investigating the stability of genotype ranking for WUE at tree scale.
Previous studies has demonstrated that Δ13C (recorded from tree ring cellulose and bulk wood) is severely affected by age, usually decreasing by a 1-3 per mil with increasing tree age ([16], [1], [12], [28], [6]). Such an age-related effect may be related to: (i) assimilation of respired CO2 near the forest floor which is already depleted in 13C ([45]); (ii) re-assimilation of respired CO2 retained within the plant canopy ([16]); (iii) decreasing hydraulic conductivity of xylem conduits with age ([31]); (iv) reduced total soil-to-leaf hydraulic conductance with tree size ([27]); or (v) tree height and light availability ([6]). Records of Δ13C in leaves and phloem sap of poplars with different ages revealed a rather stable ranking of the genotypic differences with tree age ([3]). More recently, using a diachronic approach, several genotypes of Populus deltoides × P. nigra growing in common gardens were compared and the effects of ageing (from 5 to 20 years) on the genotype ranking were studied using the Δ13C signals in tree rings as a surrogate for WUE ([38]). In the latter study, small changes in Δ13C with tree age and no significant interaction between genotype and tree age were found, reinforcing the pertinence of ranking genotypes using Δ13C. However, in a such diachronic study, the influence of long-term environmental variability and annual effects related to rainfall on Δ13C is assumed similar as the sampled trees were grown in common gardens that experienced the same environmental conditions.
In the present study we used a synchronic approach where Δ13C was recorded and compared in tree rings with same formation date, but from tree of different ages and contrasting environmental conditions. This sampling technique was used to minimise long-term environment effect and maximise age effect. Three different genotypes of Populus deltoides × P. nigra were used and carbon isotopic composition (δ13C) of the bulk wood from the year 2009 was measured and Δ13C was computed. Since cellulose and lignin are the major components of bulk wood, and due to different biochemical pathways involved in their formation, the carbon isotopic signals of these two components are also different, i.e., lignin is ~3‰ depleted than cellulose ([24], [27]). Any variation in cellulose:lignin ratio can potentially affect the overall δ13C signals of whole wood. Therefore, the variation in carbon percentage in the whole wood was investigated, that potentially elucidates the variation of lignin in the bulk wood ([25]). Moreover, nitrogen percentage in the bulk wood that can give insight regarding the tree response to the variation of available soil nitrogen resource ([17], [2]) was also measured.
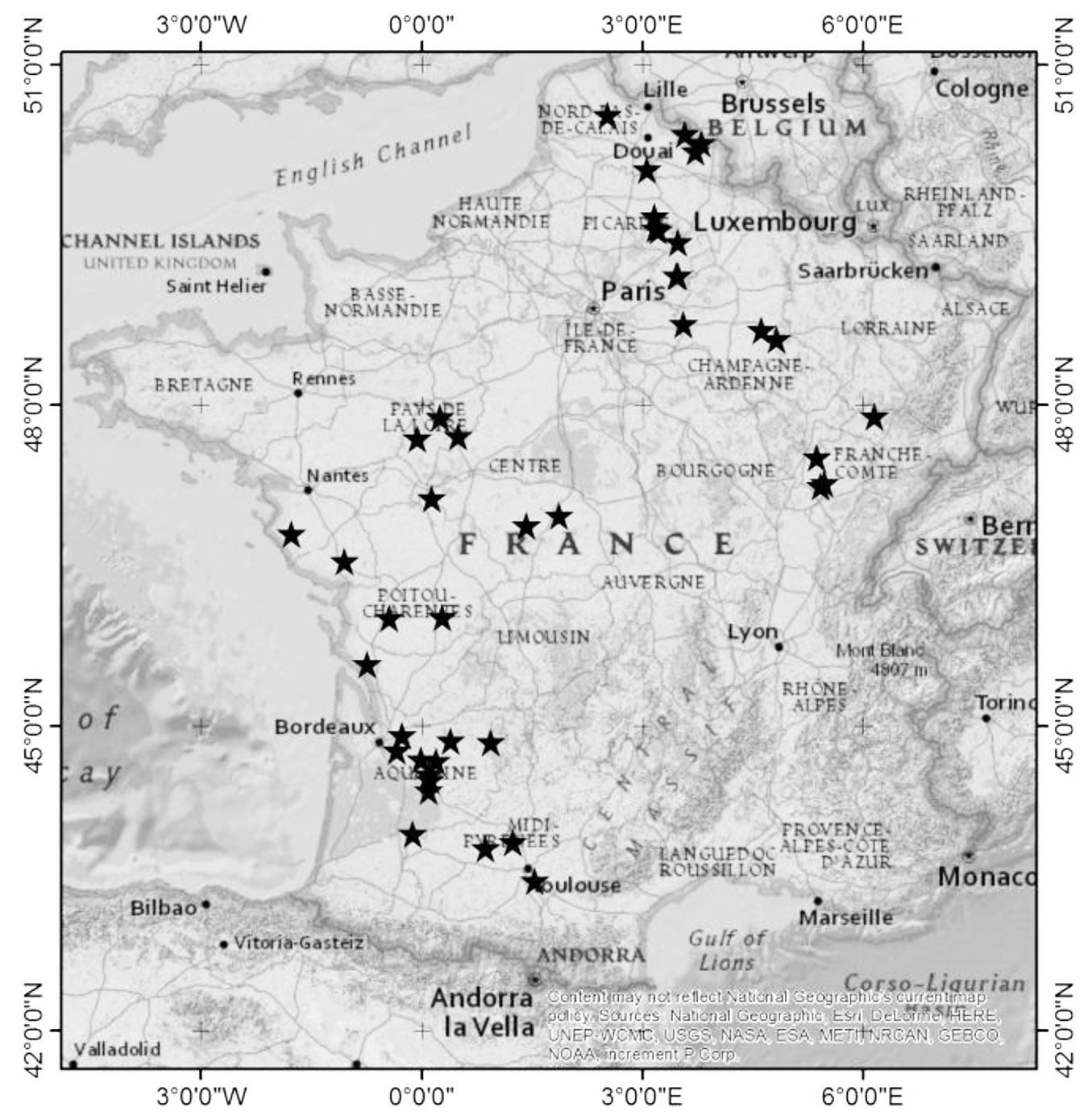
The main difficulty of the synchronic approach is the lack of large-scale common garden grouping a sufficient number of genotypes with different ages. We therefore used 41 small common gardens (Fig. 1) maintained in different locations in France by the Institut du Développement Forestier (IDF-CNPF) and gathered samples for three genotypes of Populus deltoides × P. nigra. Using this approach, we tested the hypotheses that: (i) water use efficiency estimated from Δ13C in wood is affected by tree age; and (ii) genotype ranking based on Δ13C does not change with tree age.
Fig. 1 - Location of 41 sampling sites across France. Each site is represented by a closed star.
Material and methods
The aim of this study was to evidence the effect of age on stability of genotype ranking for Δ13C. However, studying long-term physiological response in trees can be tricky because of age and environmental signals overlapping each other. Therefore, maximising age-related signals through appropriate sampling technique becomes a prerequisite. In this regard a synchronic technique, where carbon isotope discrimination was compared among tree rings with same formation date but from trees of different ages and environments, thereby maximising the age signal. A major constraint while applying such technique is to find a large number of poplar genotypes of different ages in a common garden. To obtain a satisfying sample of trees of different ages in different genotypes and environments, we sampled trees across a large number of plantations in France (Fig. 1). These small plantations were established by the Centre National de la Propriété Forestière - Institut pour le Développement Forestier (CNPF-IDF) to compare the local performance of genotypes.
Study sites
The genotypes were planted in plots of 25 trees at a distance of 7 × 7 m, i.e., at a density of 204 stems ha-1. Forty-one sites were selected (Fig. 1) which were located all over France (see Tab. S1 in Supplementary material for site information and spatial coordinates). Sites covered a large range of climates with mean annual temperature from 10 to 14 °C and precipitation from 600 to 1200 mm. Sampling sites were subsequently categorised as water available (WA, annual rainfall from 1000 to 1200 mm) or water limiting (WL, annual rainfall from 600 to 800 mm; precipitation record maintained by the Centre National de la Propriété Forestière - Institut pour le Développement Forestier, CNPF-IDF).
Sample collection and preparation
We focused on 3 Populus deltoides × P. nigra genotypes: Koster, I-214 and Dorskamp, as these genotypes are known to display contrasting Δ13C at leaf level ([29]). Moreover, they were present in a large number of the surveyed plantations and covered the different age classes. The last annual ring corresponding to year 2009 was sampled in February 2010 with an increment borer (0.5 cm2) on ~5 trees site-1 genotype-1, yielding 376 samples. All cores were divided into seven age classes, i.e., 4, 7, 9, 11, 13, 15 and 18 years. In order to minimise the environmental effect on Δ13C: (i) date effect was discarded as all cores corresponded to year 2009; and (ii) site effect was averaged in each age class by ensuring the representation of trees from both site types, i.e., WA and WL sites.
The ring formed in 2009 was carefully separated from the adjacent one and from bark with a sharp razor blade. After drying at 70 °C for 48h, each ring was ground separately into fine homogeneous powder using a ring grinder (SODEMI, CEP Industries Department, Cergy-Pontoise. France). One mg of the resulting wood powder was weighed in tin capsules for δ13C analysis.
Carbon and nitrogen percentage and carbon isotope analysis
Wood powder was combusted at 1050 °C in sealed evacuated quartz tubes containing cobalt oxide and chromium oxide as catalyst, and an amount of pure oxygen. The gases produced during combustion, CO2, H2O and NOx were passed through a reduction tube where N2 was produced and excess of oxygen was removed. Water was trapped by using anhydrous magnesium perchlorate and after reversible absorption, carbon and nitrogen percentages were measured in an elemental analyser (NA 1500-NC®, Carlo Erba, Milan, Italy - [38]). Finally, combusted products were separated by gas chromatography and the CO2 was delivered to an isotope ratio mass spectrometer (Delta-S®, Finnigan, Bremen, Germany). Carbon isotope composition was expressed as δ13C (eqn. 1):
where Rsample and Rstandard are the 13C/12C ratios in a sample and the standard (Vienna- Pee Dee Belemnite) respectively. Accuracy of the measurements was ± 0.1‰. Carbon isotope discrimination between atmosphere and wood was calculated as (eqn. 2):
where δ13Cair is the carbon isotope composition of CO2 in the atmosphere and δ13Cwood is the carbon isotope composition of the wood powder. Given that δ13Cair was -8.07‰ during 2003 and an annual decrease of 0.0281‰ ([27]), δ13Cair was estimated at -8.24‰ for 2009. We assumed that the mean value of δ13Cair was similar across sites and did only marginally change with the season.
Basal area increment
The data corresponding to annual circumference (cm) of each individual tree was acquired through the annual growth records from the Centre National de la Propriété Forestière-Insitut pour le Développement Forestier. Assuming trunks were circular in section, circumference was used to calculate the radius and eventually basal area for the years 2008 and 2009. BAI (cm2 y-1) for year 2009 was calculated as the difference between the total area corresponding to years 2009 and 2008.
Statistical analysis
Normality and homoscedasticity of data were checked graphically with residual vs. predicted and normal quantile-to-quantile plots. The data set covering the seven age classes was analysed using linear mixed models fitted with genotype, age and their interaction as fixed effects and site as a random effect. Multiple comparison tests (post-hoc Tukey HSD test) were used to evaluate pairwise differences between age classes within each genotype. Furthermore, each age class was tested for genotype effect and post-hoc Tukey HSD test were used to evaluate pair-wise differences between genotypes within each age class. All tests were performed with R ([37]) and R packages “nlme” ([35]) and “multcomp” ([19]). All tests and correlations were declared significant at P < 0.05.
Results
Carbon and nitrogen percentage
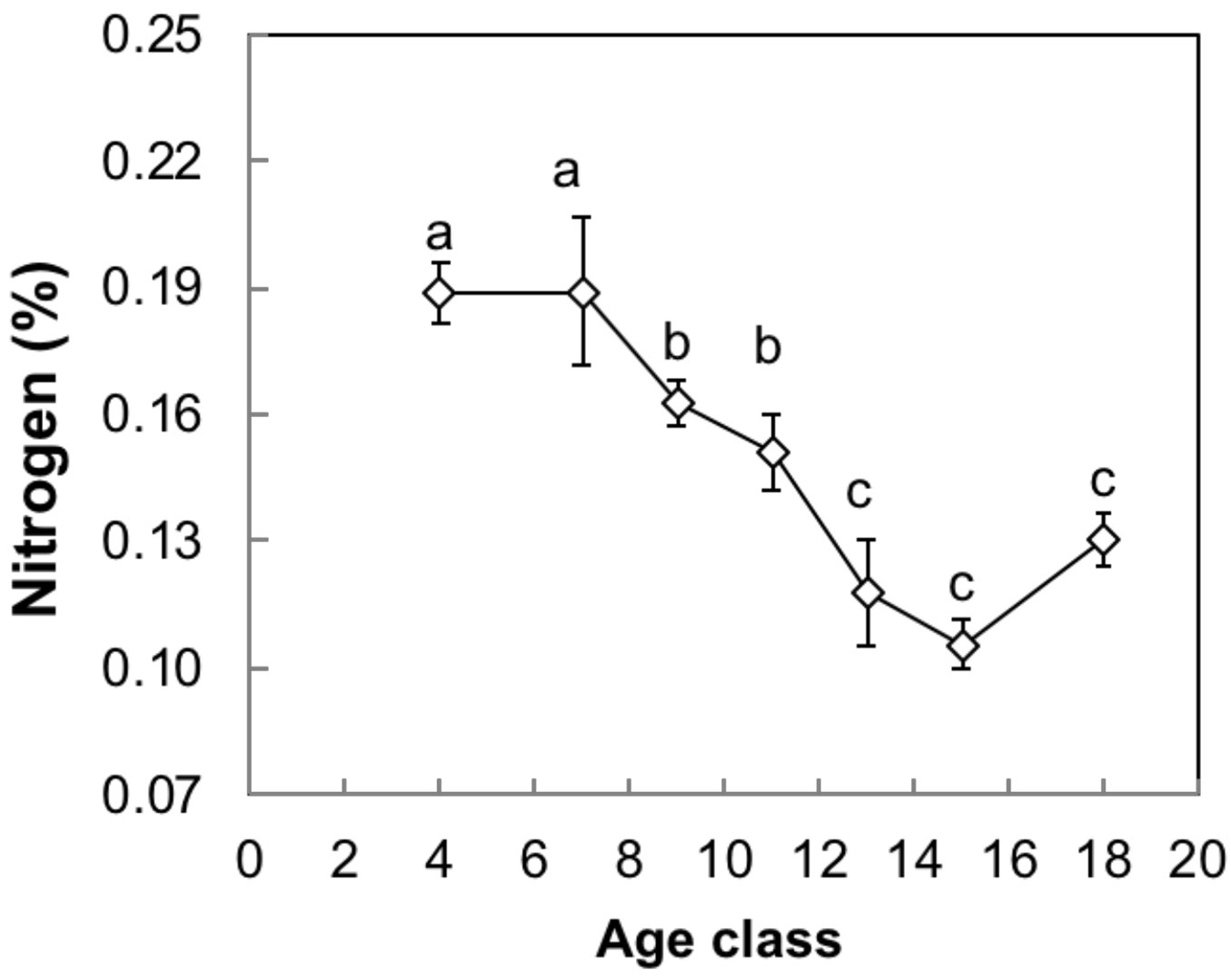
Variance between sites was smaller than residual variance (intercept = 0.731; residuals = 1.84) and model R2 resulted at 0.547 for carbon percentage. No genotype-age interaction was detected in the carbon percentage of bulk wood. It differed significantly among genotypes (P < 0.001 - for means, see Tab. 1). A non-significant trend with age was detected due to the slightly lower carbon percentage values at the age of 13 yrs. Nevertheless, carbon percentage did not display any significant trend related to age (Fig. 2). Variance between sites was smaller than residual variance (intercept = 0.009; residuals = 0.071) and model R2 resulted at 0.525 for nitrogen percentage. For the nitrogen percentage, no interaction between genotype and age was found. Nitrogen percentage did not differ among genotypes but declined significantly with age (P < 0.001); it dropped from 0.19 down to 0.11% over the tested period (Fig. 3).
Tab. 1 - Carbon and nitrogen percentage, and Δ13C measured on the bulk wood from the year 2009 from three genotypes of Populus deltoides × P. nigra aged from 4 to 18 years. Data was analyzed using linear mixed model for the effects of genotype (G), age (A) and interaction (G × A). Table shows the mean values (± SE) combining all age classes. Different letters indicate significant genotype differences after multi-comparison test (Tukey HSD, P < 0.05).
| Variable | Clone | Effect (Prob.) | ||||
|---|---|---|---|---|---|---|
| Dorskamp | I-214 | Koster | G | A | G × A | |
| C % | 46.8 ± 0.28 a | 47.7 ± 0.097 b | 47.7 ± 0.10 b | <0.001 | 0.051 | 0.305 |
| N % | 0.154 ± 0.004 | 0.14 ± 0.007 | 0.168 ± 0.007 | 0.248 | <0.001 | 0.734 |
| Δ13C (‰) | 20.3 ± 0.076 a | 19.9 ± 0.053 b | 19.4 ± 0.078 c | <0.001 | <0.001 | <0.001 |
| BAI (cm2 yr-1) | 88.4 ± 9.54 | 103 ± 11.0 | 107.3 ± 13.4 | 0.467 | 0.023 | 0.167 |
Fig. 2 - Age-related variation of Carbon percentage in the bulk wood of the year 2009, sampled during winter 2010 in three Populus deltoides × P. nigra genotypes over seven age classes. Each point represents the mean of several sites. Error bars represent ± SE.

Fig. 3 - Age-related variation of mean nitrogen percentage in the bulk wood of the year 2009, sampled during winter 2010. The curve represents the means of three Populus deltoides × P. nigra genotypes over seven age classes. Error bars represent ± SE and different letters indicate significant differences between age classes after Tukey HSD test (P < 0.05).
Age and genotype effects for Δ13C
Variance between sites was smaller than residual variance (intercept = 0.38; residuals = 0.51) and model R2 resulted at 0.99. Δ13C variation was significantly explained by an interaction between genotype and age (P < 0.001 - Tab. 1). A detailed post-hoc analysis revealed that an age effect was detectable in all three genotypes (Fig. 4). There was a significant increase of Δ13C by 2‰ from 4 to 9 yrs in Dorskamp and Koster (Fig. 4a, Fig. 4c) and a less visible one in I-214 (Fig. 4b) where it increased from 4 to 7 yrs and remained almost stable afterwards.
Fig. 4 - Age-related variation in carbon isotope discrimination (Δ13C) in the 2009 year-ring for three Populus deltoides × P. nigra genotypes (a) Dorskamp, (b) I 214 and (c) Koster. Each point represents a mean value from several sites. Error bars represent ± SE and different letters indicate significant differences between age classes for each genotype after Tukey HSD test (P < 0.05).
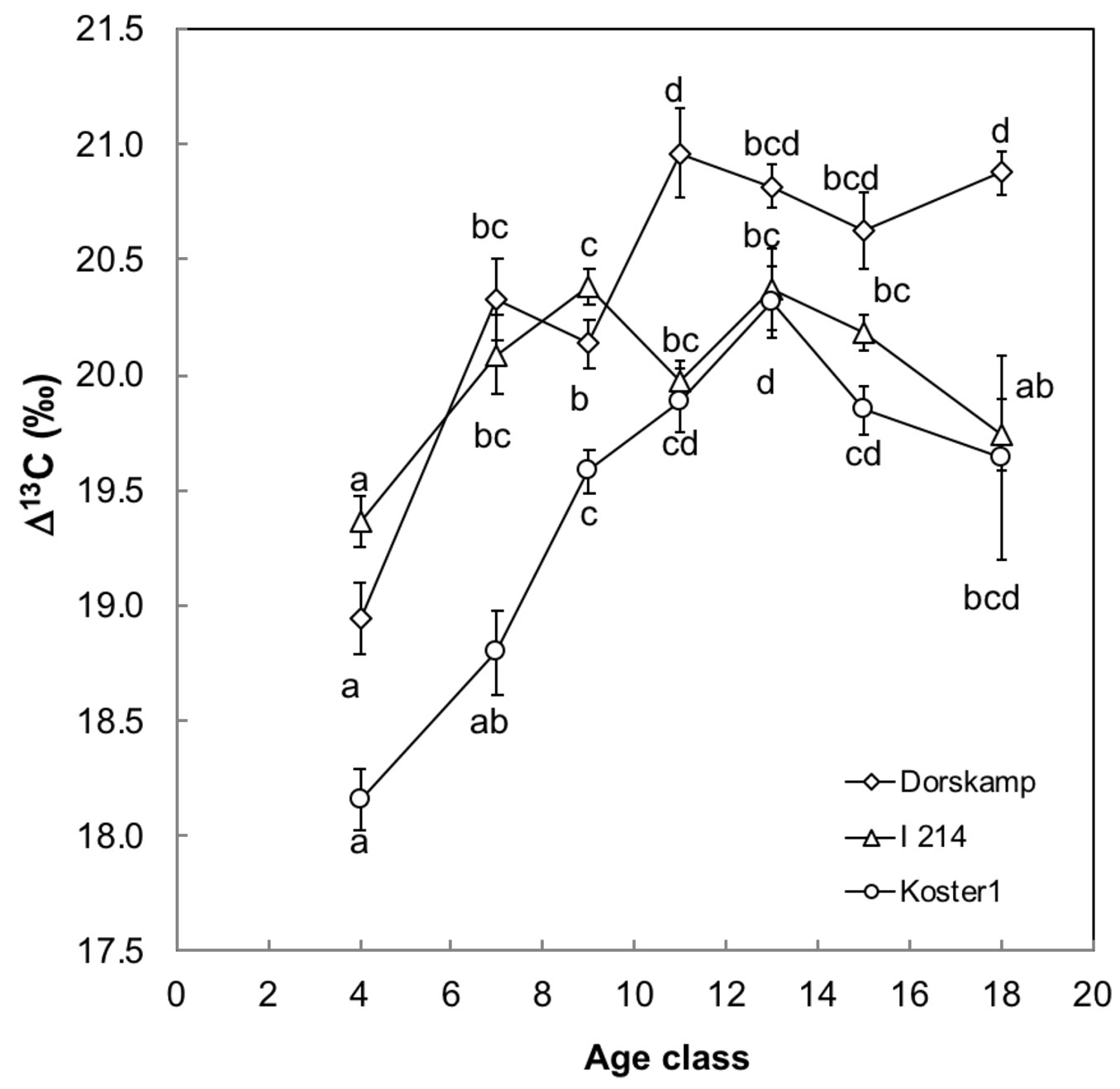
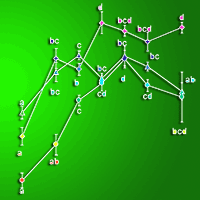
Genotype effect within each age group was found significant and was assessed with post-hoc Tukey HSD test (Tab. 2). In the young age classes (4 to 9 yrs), Koster displayed the lowest mean Δ13C followed by Dorskamp and I-214 that displayed similar values. At higher ages, Koster and I-214 displayed the lowest and Dorskamp displayed the highest mean Δ13C, with the exception of age class of 13 yrs, where no significant difference could be detected.
Tab. 2 - Carbon isotopic discrimination (± SE) between atmosphere and bulk wood (Δ13C) in the year 2009 for three Populus deltoides × P. nigra genotypes (Dorskamp, I-214 and Koster). Each age class was tested for genotype effect (F-value and P-value) and different letters indicate a significant genotype difference for each age class after Tukey HSD test (P < 0.05).
| Age class (yrs) |
df | Dorskamp | I-214 | Koster | F-value | P-value |
|---|---|---|---|---|---|---|
| 4 | 43 | 18.9 ± 0.156 a | 19.3 ± 0.107 a | 18.1 ± 0.132 b | 22.3 | <0.001 |
| 7 | 47 | 20.3 ± 0.179 a | 20.0 ± 0.172 a | 18.7 ± 0.179 b | 18.4 | <0.001 |
| 9 | 88 | 20.1 ± 0.103 a | 20.3 ± 0.079 a | 19.5 ± 0.096 b | 17.9 | <0.001 |
| 11 | 80 | 20.9 ± 0.191 a | 19.9 ± 0.095 b | 19.8 ± 0.136 b | 13.8 | <0.001 |
| 13 | 24 | 20.8 ± 0.095 | 20.3 ± 0.175 | 20.3 ± 0.150 | 3.03 | 0.069 |
| 15 | 34 | 20.6 ± 0.161 a | 20.1 ± 0.077 b | 19.8 ± 0.103 b | 9.26 | <0.001 |
| 18 | 39 | 20.8 ± 0.092 a | 19.7 ± 0.151 b | 19.6 ± 0.443 b | 19.6 | <0.001 |
Inter annual and genotype correlation between Δ13C and BAI
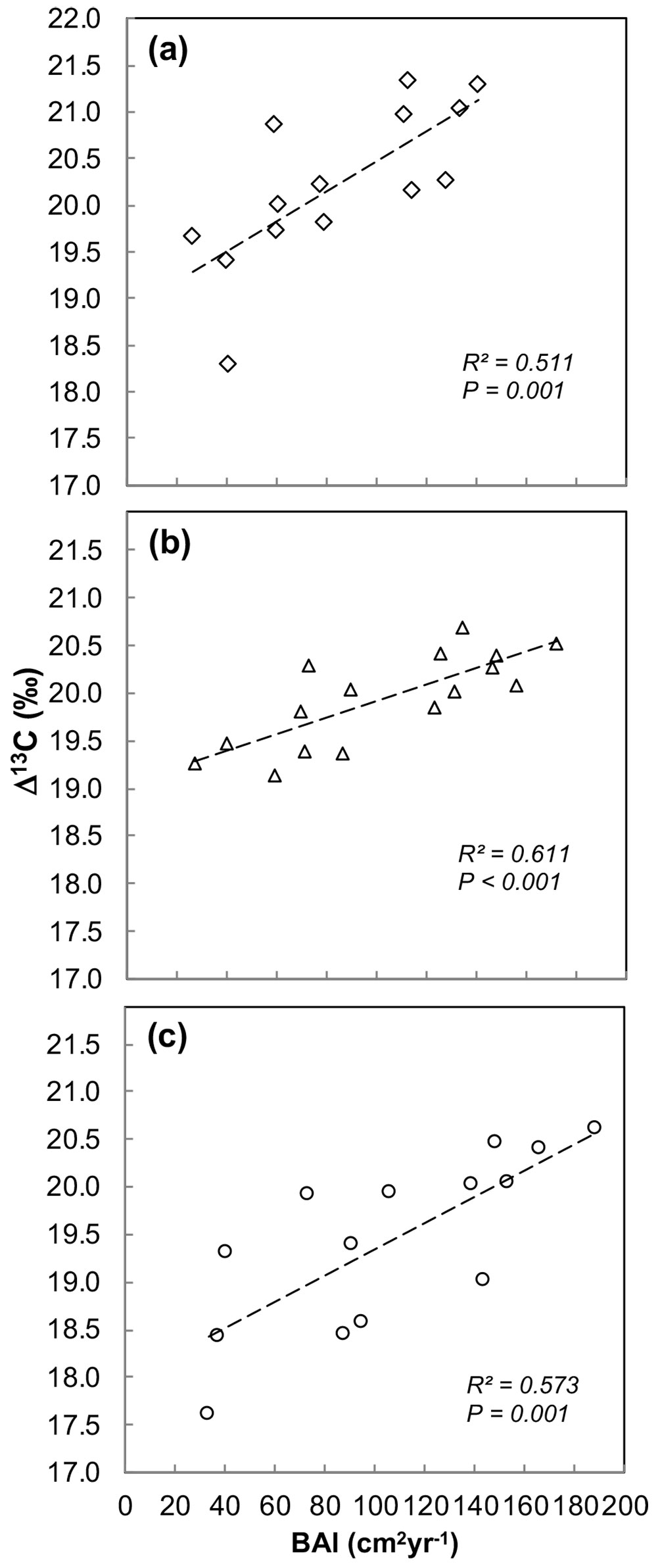
The average BAI of the year 2009 was homogenous among the three genotypes over the tested period (Tab. 1), and therefore the genotypic differences in Δ13C were independent from any difference in radial growth rate. The inter annual variation of BAI due to the age effect was positively correlated to Δ13C in the three genotypes (Fig. 5)
Fig. 5 - Correlation between carbon isotopic discrimination (Δ13C) and basal area increment (BAI) in three Populus deltoides × P. nigra genotypes (a) Dorskamp: Δ13C = 0.0161BAI + 18.9; (b) I-214: Δ13C = 0.0087 BAI + 19.0; and (c) Koster: Δ13C = 0.0138 BAI + 18.0. Each point represents values measured at a given site.
Discussion
The present research was aimed to assess the stability of genotype differences in water use efficiency (WUE) with tree age; we compared three cultivars of Populus deltoides × P. nigra for this trait in plantation in situ across France and used a synchronic approach, based on Δ13C recorded from the year 2009 of individuals with different ages.
Genotype and age effect on Δ13C and relationship with BAI
Whole wood consists of complex mixtures of molecules like cellulose, hemicellulose and lignin with variable isotopic signatures. Early studies used whole wood for the isotopic analysis on tree rings. However, the isotopic composition of individual wood components differs largely ([47]). Thus, we suspected that the observed significant genotypic difference in mean Δ13C could potentially be due to changes in cellulose vs. lignin ratios with contrasting isotopic signals. Although previous studies have shown that using whole wood is better than inducing extraction error while using cellulose to measure δ13C ([18], [38]), in this study we minimised this effect by using the last annual ring (2009, with same cambial age) for isotope analysis.
Our results showed the existence of an age effect on Δ13C, where Δ13C increased to a larger extent during first age classes of 4-11 years in two genotypes (Dorskamp and Koster). This positive trend in Δ13C with age is contrary to several previous studies reporting age-related decreases in Δ13C using different dendrochronological approaches ([1], [12], [28], [34]). Δ13C has been expected to be an estimator of water use efficiency ([15]). Several studies have shown a clear negative correlation between Δ13C and A/gs (intrinsic water use efficiency, iWUE) and WUE (whole-plant water use efficiency - [43], [7], [39]). Thus, increase in Δ13C with age in our study reflects a decrease in iWUE with age. This decrease in iWUE (A/gs) can either be due to decreased CO2 assimilation rate (A) or increased stomatal conductance (gs - [14], [9]). Furthermore, Δ13C and tree growth are often correlated depending upon the growth environment. This correlation was found to be positive for some species, i.e., Eucalyptus globulus Labill ([33]. [36]), Fagus sylvatica L. ([11]) and Pinus radiata ([44]). On the contrary, some other conifer species displayed a negative correlation between Δ13C and growth, i.e., Larix occidentalis Nutt. ([49]), Pinus pinaster ([32], [5]), Picea mariana Mill ([21]) and Pinus caribaea Morelet ([48]). However, no correlation between Δ13C and tree growth was found in P. × euramericana in common garden ([29]) and poplar plantation ([3]). Within each genotype, we found a positive correlation between BAI and Δ13C with age, which is in line with previous findings in different Populus deltoides × P. nigra genotypes ([38]). Similar positive correlation between BAI and Δ13C has been reported in Pinus radiata ([44]). The positive correlation between Δ13C and BAI suggests that the inter-annual variation of Δ13C is controlled to a larger extent by stomatal conductance than by photosynthetic capacity ([21], [48]). In poplar, mostly variation in stomatal conductance rather than photosynthetic capacity seems to control carbon gain and growth ([8], [30], [40]). Therefore, based on the significant positive correlation between BAI and Δ13C across tested genotypes, we may conclude that the observed increase in Δ13C with age and interannual variation of Δ13C was largely controlled by the variation in stomatal conductance rather photosynthetic capacity.
Stability of the genotypic ranking for Δ13C
Checking genotype rank stability with age is of central importance for selecting genotypes for higher water use efficiency. Therefore, genotype ranking in Populus deltoides × P. nigra was tested for Δ13C in controlled vs. open field conditions ([29]) and subsequently, in well irrigated vs. water stress under field conditions ([30]). In both studies, genotypic ranking remained stable with no correlation found between Δ13C and productivity traits. However, these studies were done on young plants. Parallel to that, many previous studies evidenced an age effect on Δ13C and demonstrated that duration and extent of age effect is variable according to many species ([23]). In this context, genotype ranking made on young plants were susceptible to change with age. Our results showed that genotype ranking shuffled across first age class (4 years) to seventh age class (18 years), i.e., I-214 displayed highest and Koster the lowest Δ13C values in the first age class (4 years), whereas I-214 was ranking second in the seventh age class (18 years). In spite of rank shuffling with age, genotypic ranking for mean Δ13C values matched with that found by Marron et al. ([26]) and Monclus et al. ([29]), where Koster had the lowest Δ13C and Dorskamp had the highest, and with Rasheed et al. ([38]) in Begaar for Dorskamp and I-214. Thus, we may conclude that based on a relatively small number of cultivars, a stability of genotype ranking was observed with age in two cultivars, at least during the approx. 20 years from planting to harvest. This conclusion is based on a limited range of cultivars. Unfortunately, extension to a larger number of genotypes was impossible given the lack of suitable even-aged common garden plantation of poplar cultivars at suitable age, not to speak of common gardens with different tree ages.
Conclusion
In this study, Δ13C assessed from the year 2009 increased with tree age in the three genotypes, indicating a decrease in water use efficiency with age at whole tree level. Furthermore, inter-annual variation of Δ13C was found positively correlated to BAI in all genotypes which shows that Δ13C was largely controlled by stomatal conductance rather photosynthetic capacity. Significant genotypic effect was detected for mean Δ13C over the tested period. The genotypic ranking of Δ13C was: (i) maintained among the three Populus deltoides × P. nigra genotypes, despite of crossing detected with tree age; (ii) found consistent with the previous ranking for Δ13C. Finally, Koster genotype was found highly water use efficient and productive with age among the tested genotypes. However, further studies are required to test other genotypes as well.
Acknowledgements
The authors are very grateful to Cyril Buré (INRA, UMR EEF) for helping in arranging the field visits and in collecting samples together with the different technicians of CRPF. Christian Hossann and Claude Bréchet (INRA, UMR EEF), for spending time doing all isotope analyses. Finally, we thank WFJ Parsons (CEF) for English editing. We are thankful to two anonymous reviewers for their suggestions to improve the manuscript. The research was supported by the European Union FP7 Project “Novel Tree Breeding Strategies” Project no. FP7 - 211868. F. Rasheed was supported by a grant from the Higher Education Commission of Pakistan.
References
CrossRef | Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Authors’ Info
Authors’ Affiliation
Erwin Dreyer 0000-0003-4999-5072
Didier Le Thiec 0000-0002-4204-551X
INRA, UMR1137 Ecologie et Ecophysiologie Forestières, IFR 110 EFABA, F-54280 Champenoux (France)
Zikria Zafar 0000-0001-8881-7653
Department of Forestry & Range Management, University of Agriculture Faisalabad, P.O. Box 38000 (Pakistan)
Didier Le Thiec 0000-0002-4204-551X
Université de Lorraine, UMR1137 Ecologie et Ecophysiologie Forestières, IFR 110 EFABA, F-54500 Vandoeuvre-lès-Nancy (France)
Department of Natural Sciences, Institute of Temperate Forest Sciences (ISFORT), University of Quebec in Outaouais (UQO), 58 Main St, Ripon, Quebec J0V 1V0 (Canada)
Corresponding author
Paper Info
Citation
Rasheed F, Dreyer E, Le Thiec D, Zafar Z, Delagrange S (2019). Tree aging does not affect the ranking for water use efficiency recorded from δ13C in three Populus deltoides × P. nigra genotypes. iForest 12: 272-278. - doi: 10.3832/ifor2896-012
Academic Editor
Rossella Guerrieri
Paper history
Received: Jun 15, 2018
Accepted: Mar 18, 2019
First online: May 21, 2019
Publication Date: Jun 30, 2019
Publication Time: 2.13 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2019
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 51329
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 40244
Abstract Page Views: 3653
PDF Downloads: 6561
Citation/Reference Downloads: 3
XML Downloads: 868
Web Metrics
Days since publication: 2593
Overall contacts: 51329
Avg. contacts per week: 138.57
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2019): 1
Average cites per year: 0.14
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Preliminary study on genetic variation of growth traits and wood properties and superior clones selection of Populus ussuriensis Kom.
vol. 12, pp. 459-466 (online: 29 September 2019)
Research Articles
Energy production of poplar clones and their energy use efficiency
vol. 7, pp. 150-155 (online: 23 January 2014)
Research Articles
Genetic variation and heritability estimates of Ulmus minor and Ulmus pumila hybrids for budburst, growth and tolerance to Ophiostoma novo-ulmi
vol. 8, pp. 422-430 (online: 15 December 2014)
Research Articles
Seedling emergence capacity and morphological traits are under strong genetic control in the resin tree Pinus oocarpa
vol. 17, pp. 245-251 (online: 16 August 2024)
Research Articles
Role of photosynthesis and stomatal conductance on the long-term rising of intrinsic water use efficiency in dominant trees in three old-growth forests in Bosnia-Herzegovina and Montenegro
vol. 14, pp. 53-60 (online: 28 January 2021)
Research Articles
Age trends in genetic parameters for growth and quality traits in Abies alba
vol. 9, pp. 954-959 (online: 07 July 2016)
Research Articles
Genetic diversity of core vs. peripheral Norway spruce native populations at a local scale in Slovenia
vol. 11, pp. 104-110 (online: 31 January 2018)
Research Articles
Earlywood vessel features in Quercus faginea: relationship between ring width and wood density at two sites in Portugal
vol. 8, pp. 866-873 (online: 27 April 2015)
Research Articles
Genetic control of intra-annual height growth in 6-year-old Norway spruce progenies in Latvia
vol. 12, pp. 214-219 (online: 25 April 2019)
Research Articles
Patterns of genetic variation in bud flushing of Abies alba populations
vol. 11, pp. 284-290 (online: 13 April 2018)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







