
Spatial and temporal variation of drought impact on black locust (Robinia pseudoacacia L.) water status and growth
iForest - Biogeosciences and Forestry, Volume 8, Issue 6, Pages 743-747 (2015)
doi: https://doi.org/10.3832/ifor1299-008
Published: Jun 18, 2015 - Copyright © 2015 SISEF
Research Articles
Abstract
Stimulated by the rising demand for bioenergy, forestry practices for energy production are of increasing importance worldwide. Black locust (Robinia pseudoacacia L.) is a suitable tree species for biomass production in short-rotation plantations in East Germany, especially on marginal land where insufficient water and nutrients are a limiting factor for tree growth. Our study aims to clarify the spatial and temporal variability of the black locust growth through the analysis of the plant water status, and to evaluate the effect of adverse edaphic conditions on growth performances, amplified by periods of summer drought. The study was carried out at two sites presenting comparable climatic but different edaphic conditions: (i) fertile agricultural soil; and (ii) heterogeneous unstructured soil from a reclaimed post-mining area. During the vegetation period, the growth rate decreased in both sites following the plant water status in terms of pre-dawn leaf water potential. Particularly in the post-mining area, due to the adverse edaphic conditions, below the critical pre-dawn water potential value of -0.5 MPa, the stem growth was drastically reduced during a period of summer drought. However, the trees could cope with the extreme soil and weather conditions in the post-mining site without perishing.
Keywords
Agroforestry, Drought Stress, Soil Water Availability, Soil Heterogeneity, Reclamation
Introduction
The interest in short-rotation coppice systems rose over the last decades, aiming to improve the sustainability of the second generation bioenergy crops. Besides their positive ecological functions, fast-growing trees like poplars and willows can be harvested and used for bioenergy production ([17]). Among the more common tree species used in northern Europe for bioenergy production, black locust (Robinia pseudoacacia L.) is planted with remarkable success for reclamation in lignite opencast mining areas in Brandenburg (NE Germany - [6], [4]). Here, due to the ongoing mining activities, the groundwater table is lowered, and therefore plant growth depends mostly on the amount of rainfall. In those areas characterized by unfavorable edaphic conditions ([7]), the soil is mostly an unstructured clayed-sandy material, extremely heterogeneous and poor in organic materials and nutrients. As a result ([3]), a decrease in productivity is to be expected, due to the scarcity of nutrients. Furthermore, the soil water availability during summer is more prone to water deficit than in agricultural soils under the same climatic conditions. Since water and nutrient availability are the main limiting factors for primary production, there is a need to evaluate the drought impact on the black locust growth performance, taking into account the response of the plant to different edaphic conditions. There are several research works carried out at field scale, describing the black locust biomass production and its relation to water constraint ([1]). The link between the black locust growth and the soil water availability has been recently clarified by Mantovani et al. ([11], [12]), however only in a controlled lysimeter experiment. Therefore, in our work we aimed at evaluating the impact of summer drought periods and nutrient paucity on black locust growth performance in the field, linking the plant water status to the radial increment of the trunk during the vegetation period. The spatial and temporal variability in leaf water potential and its implications for tree growth was taken into account. More specifically we carried out a critical comparison between black locust growth performance under water limitation in two sites with different edaphic but comparable climatic conditions.
Material and method
Study sites
The study area is located in southern Brandenburg, Germany, about 150 km southeast of Berlin and is characterized by an average annual precipitation of 556 mm and a mean annual temperature of 9.3 °C (1951-2003, nearest climatic station Cottbus, Deutscher Wetterdienst).
For the experiment two different sites were chosen: (i) Forst (51° 47′ 24″ N, 14° 37′ 57″ E), a fertile agricultural field characterized by sandy loam soil (Gleysol - [5]). While located close to the river Neiße, the annual variation of the groundwater table lies between 1.0 and 2.2 m below the soil surface. (ii) Welzow-Süd (51° 37′ 23″ N, 14° 19′ 34″ E), a reclaimed post-mining site with a groundwater table lower than 100 m below the soil surface due to the ongoing mining activity. The soil here is heterogeneous and consists mainly of sands and clayed-sands in an initial stage (Regosol - [5]), therefore it is still unstructured ([9]). The total organic carbon (TOC) measured in Forst (10.1 g kg-1) was higher than in Welzow-Süd (2.9 g kg-1), while the total nitrogen (TN) values amounted to 0.9 g kg-1 in both sites.
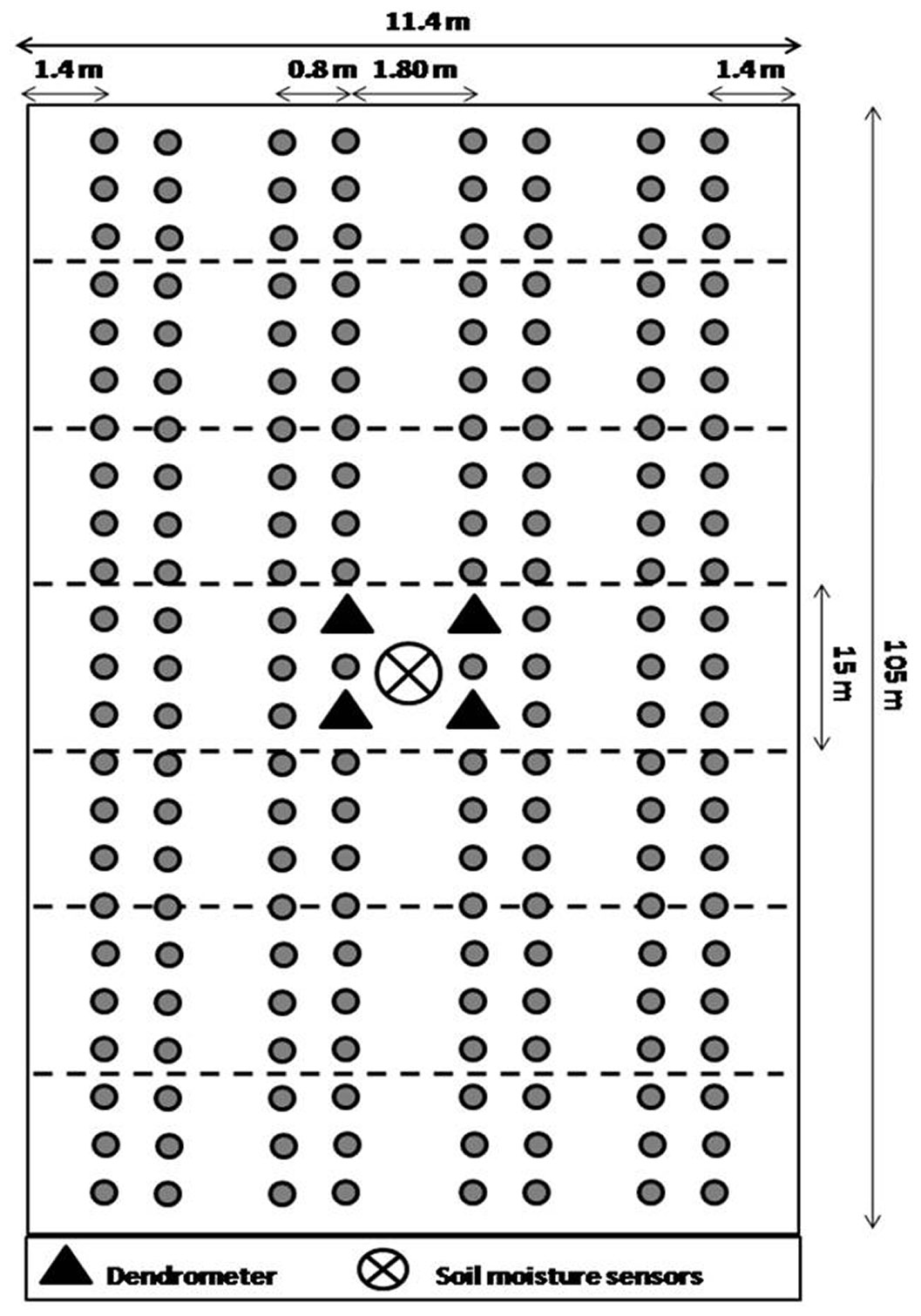
Black locust bare-root seedlings were planted in Forst in the spring of 2010 and in Welzow-Süd in the spring of 2007. The plants were established in form of an agroforestry system as a short-rotation coppice for bioenergy production. The design of the agroforestry system at both sites was comparable. The tree hedgerows consisted of four double rows. Along the row the trees were planted every 0.85 m. The tree distance within and between the double rows was 0.8 m and 1.8 m, respectively. Crop alleys in between the hedgerows had a width of 24 m. One plot of 11.4 × 105 m was selected at each site (Fig. 1), and split in 7 subplots of 11.4 × 15 m.
Fig. 1 - Schematic diagram of the tree plantation in Forst and Welzow-Süd. The figures are not in scale and the circles in the figure are only representative of the tree rows, not of the single trees.
Air temperature, relative humidity and precipitation values were measured using weather stations and stored on data loggers (Delta-T devices Ltd., Cambridge, UK) in 10 min intervals.
Hydrological soil characteristics
The saturated hydraulic conductivity (Ks) was determined in undisturbed soil cores (100 cm) sampled at both sites at 5, 20, and 40 cm depths in six replicates. We chose the permanent head method using a lab permeameter (Eijkelkamp, Giesbeek, Netherlands). After measuring the Ks, the bulk density of the same cores was determined by oven-drying at 105 °C until constant weight of cores was reached.
Plant growth and plant water status
For each subplot four fully developed trees were selected. The monthly tree growth was recorded by measuring the diameter at breast height (DBH) with a polyethylene caliper. At the beginning of the experiment, in the spring of 2012 the mean DBH was 18.4 ± 4 mm in Forst and 35.4 ± 8 mm in Welzow-Süd, which suggest that, despite the adverse edaphic conditions, tree growth can be considered relatively normal also in the latter site. In addition, the daily growth rate of the four selected trees was evaluated with dendrometers (DDL, Ecomatik, Dachau, Germany) at breast height at both sites. The selected trees were located in the center of the tree rows (in the subplots W4 and F4 - see Fig. 1). The plant water status of 56 selected trees was evaluated periodically, measuring the pre-dawn leaf water potential (Ψpd) of three leaves per tree, using the Scholander pressure chamber (Plant Water Status Console 3000, Soilmoisture Inc., Santa Barbara, CA, USA). The pre-dawn leaf water potential was measured starting two hours before sunrise. The trees were georeferenced by using a differential GPS system (Trimble R8 GNSS /R6 /5800, Dayton, Ohio, USA). The spatial information, together with the Ψpd values, was integrated into a Geographic Information System (ASDGIS Spatial Analyst software, ESRI, Redland, CA, USA) and used to create a dynamic map.
Statistical analysis
A non-parametric analysis Mann-Whitney U-Test (α < 0.05) was performed to compare the two sites in terms of: (i) bulk density; (ii) saturated hydraulic conductivity; (iii) mean cumulative daily diameter increment, measured in the subplots F4 and W4; (iv) mean Ψpd recorded during the same day in different subplots within the same site. The non-parametric Spearmans’ ρ correlation (α < 0.01) was applied to link the mean weekly growth rates to the Ψpd. The analyses were carried out using the IBM SPSS® version 21 (SPSS Inc. Chicago, IL, USA). For the geospatial analysis the ASDGIS Spatial Analyst software (ESRI) was used. Spatial and temporal variations of the Ψpd were evaluated by interpolating the Ψpd values obtained in the different subplots during the vegetation period with the inverse distance weighted interpolation method (IDW).
Results
Climatic conditions
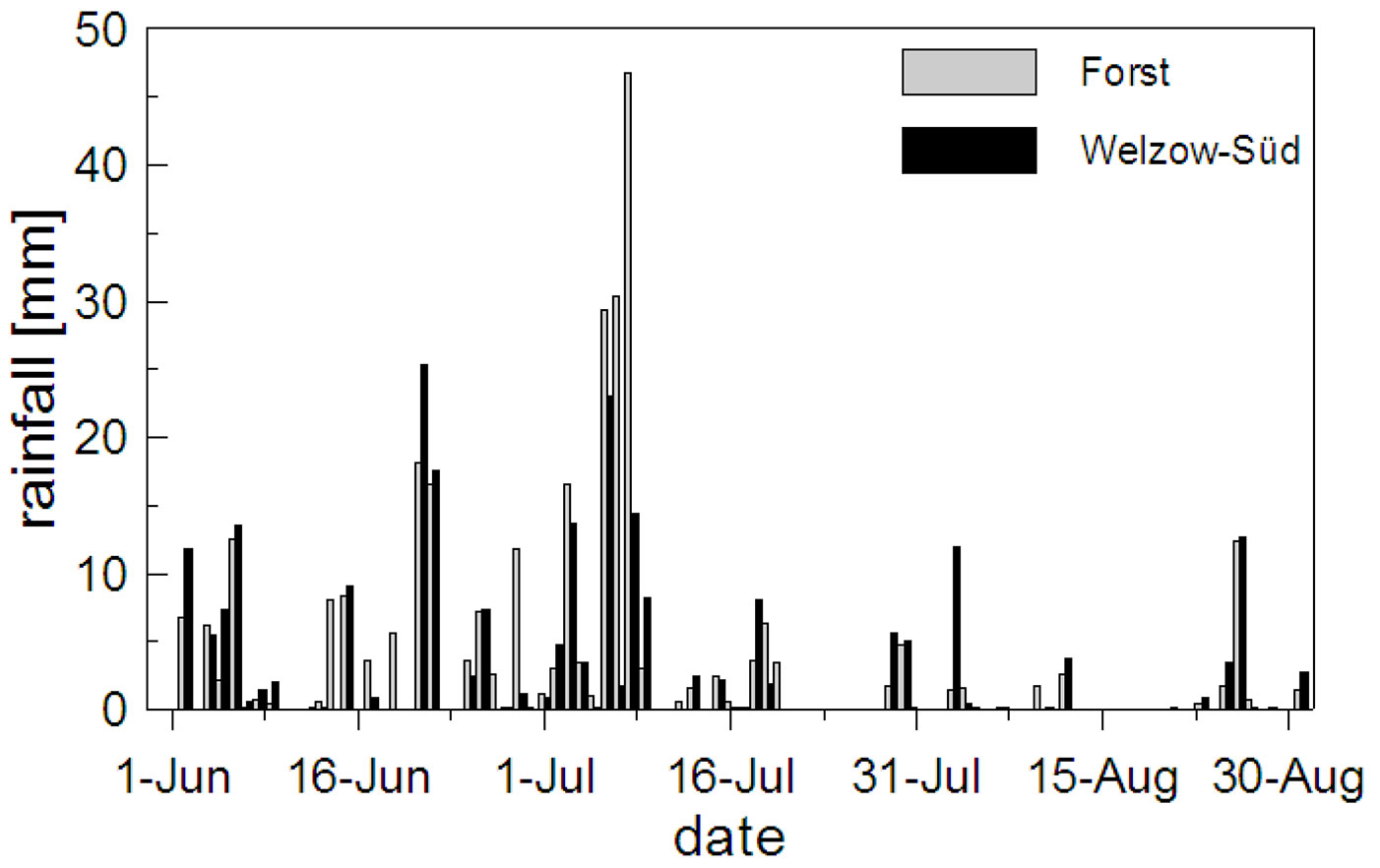
The mean daily air temperature and the vapor pressure deficit (VPD) at both study sites are shown in Fig. 2. In Welzow-Süd 30% and in Forst 25% of the measured days had a mean daily temperature lying between 20 and 25 °C. The VPD instead showed more variation between the two sites, with 33% and 16% of the days above 5 hPa Pa-1 for Welzow-Süd and Forst, respectively (Fig. 2b). The global radiation at both sites was comparable, with values higher than 200 Wm-2 for approximately 60% of the days (Fig. 3a). During the vegetation period the total rainfall in Welzow-Süd amounted to 238 mm, while in Forst 301 mm were recorded (Fig. 3b), the daily amount for each sites is shown in Fig. 4.
Fig. 2 - Daily mean air temperature (a) and daily mean vapor pressure deficit (b) measured at Forst and Welzow-Süd during the vegetation period (from June 1st to August 30th, 2012).
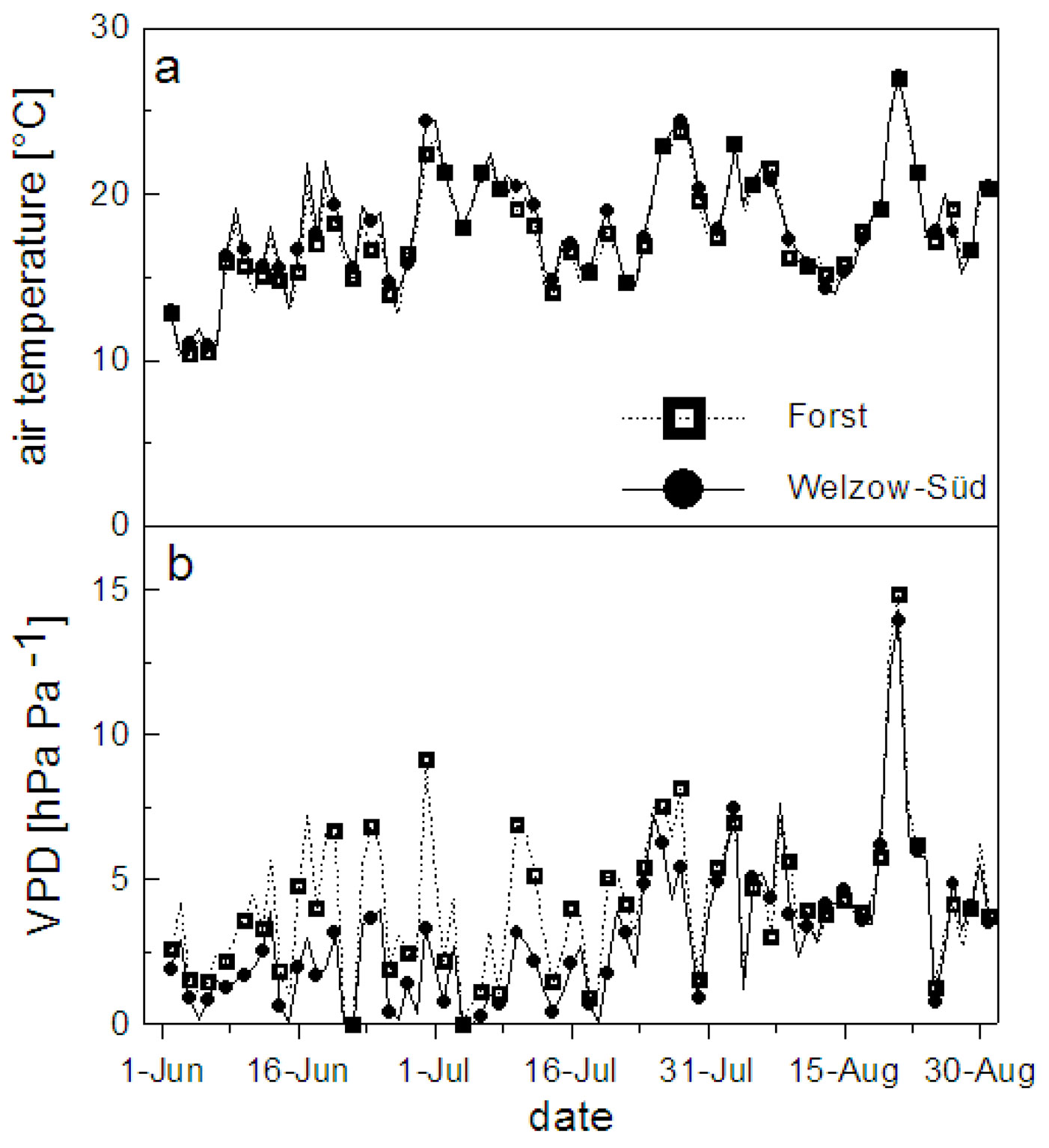
Fig. 3 - Daily mean global radiation (a) and cumulative rainfall (b) measured at Forst and Welzow-Süd during the vegetation period (from June 1st to August 30th, 2012).
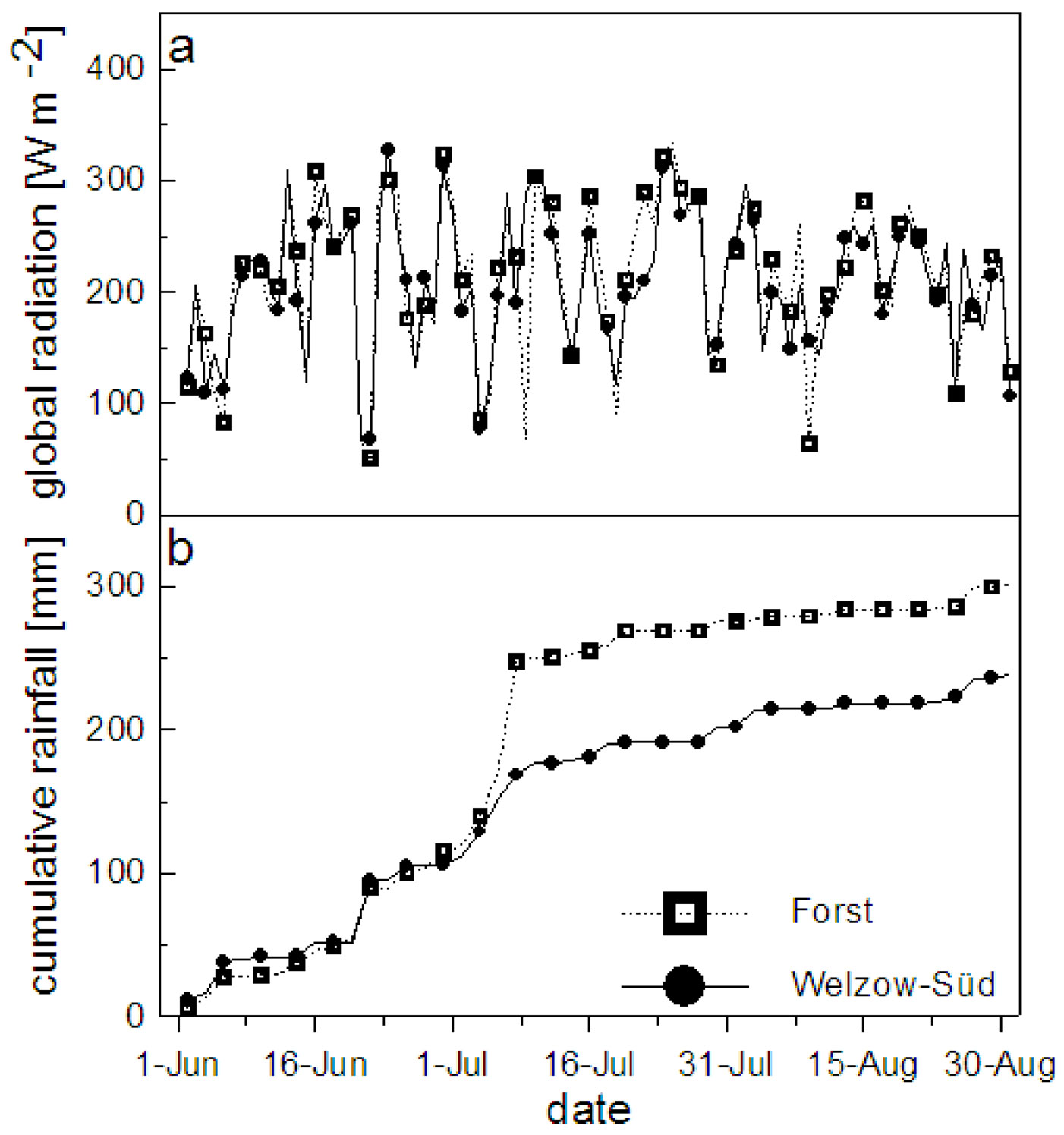
Fig. 4 - Daily rainfall amount at Forst (a) and Welzow-Süd (b) recorded during the vegetation period (from June 1st to August 30th 2012).
Hydraulic conditions and soil moisture
The mean bulk density of the Gleysol in Forst was 1.6 g cm-3 and the Ks values measured at 5, 20, and 40 cm depth were 59, 17, and 27 cm day-1 respectively. Although the structureless Regosol sampled in Welzow-Süd presented a comparable bulk density (1.7 g cm-3), the Ks was significantly higher, varying between values of 559 (p = 0.032), 223 (p = 0.036) and 73 (p = 0.42) cm day-1, measured at 5, 20, and 40 cm depth respectively.
Daily diameter increment
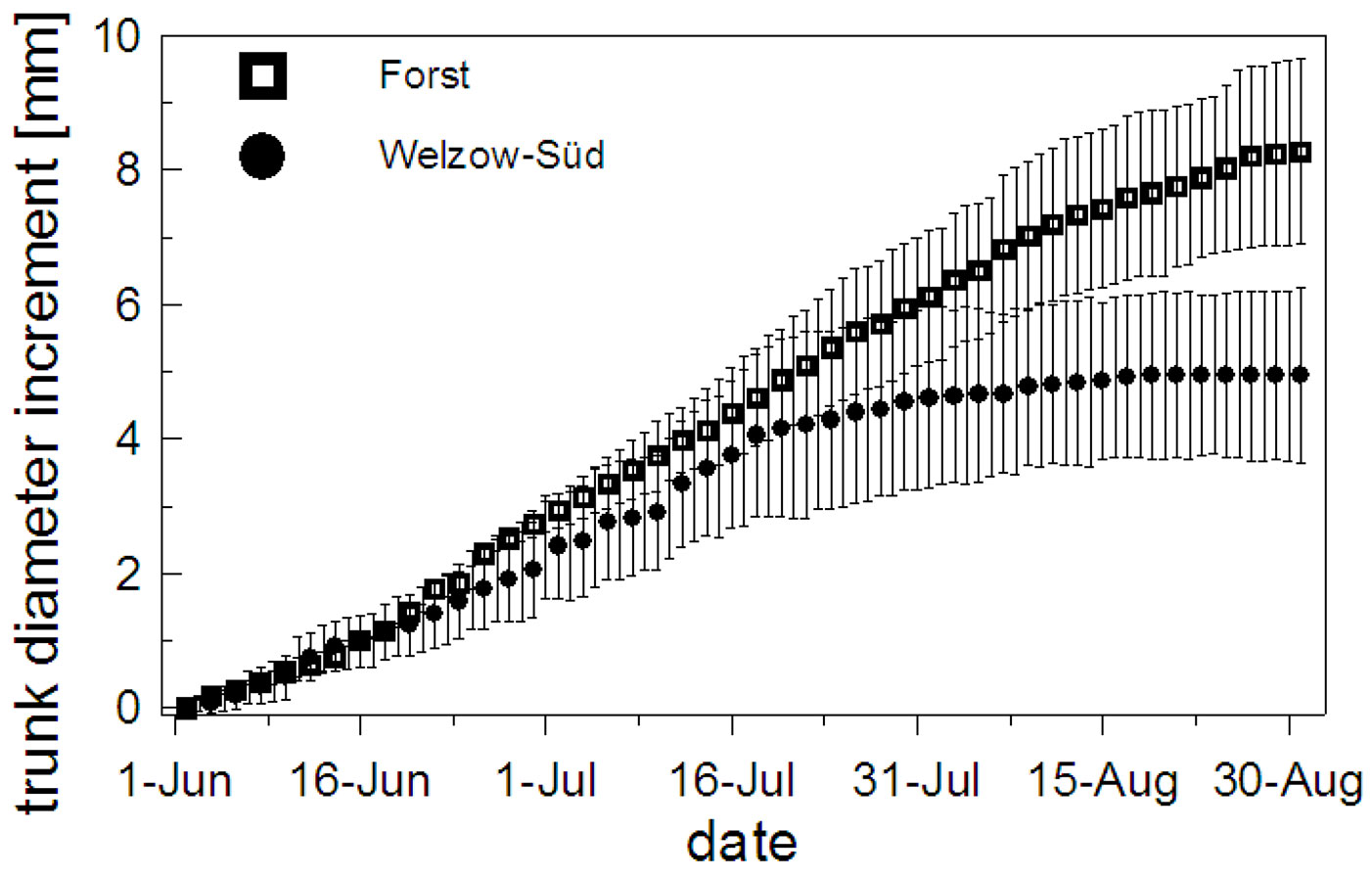
The dendrometers installed in the subplots F4 and W4 recorded the daily diameter growth and we found no significant difference (p = 0.067) between the sites until July 25, 2012 (Fig. 5). The difference only became significant (p = 0.038) towards the end of the experiment when in Welzow-Süd the plant diameter increased only 52% of the plants established in Forst.
Fig. 5 - Mean and standard deviation of the trunk diameter increment measured in the F4 subplot (Forst) (a) and W4 subplot (Welzow-Süd) (b) during the vegetation period (from June 1st to August 30th, 2012).
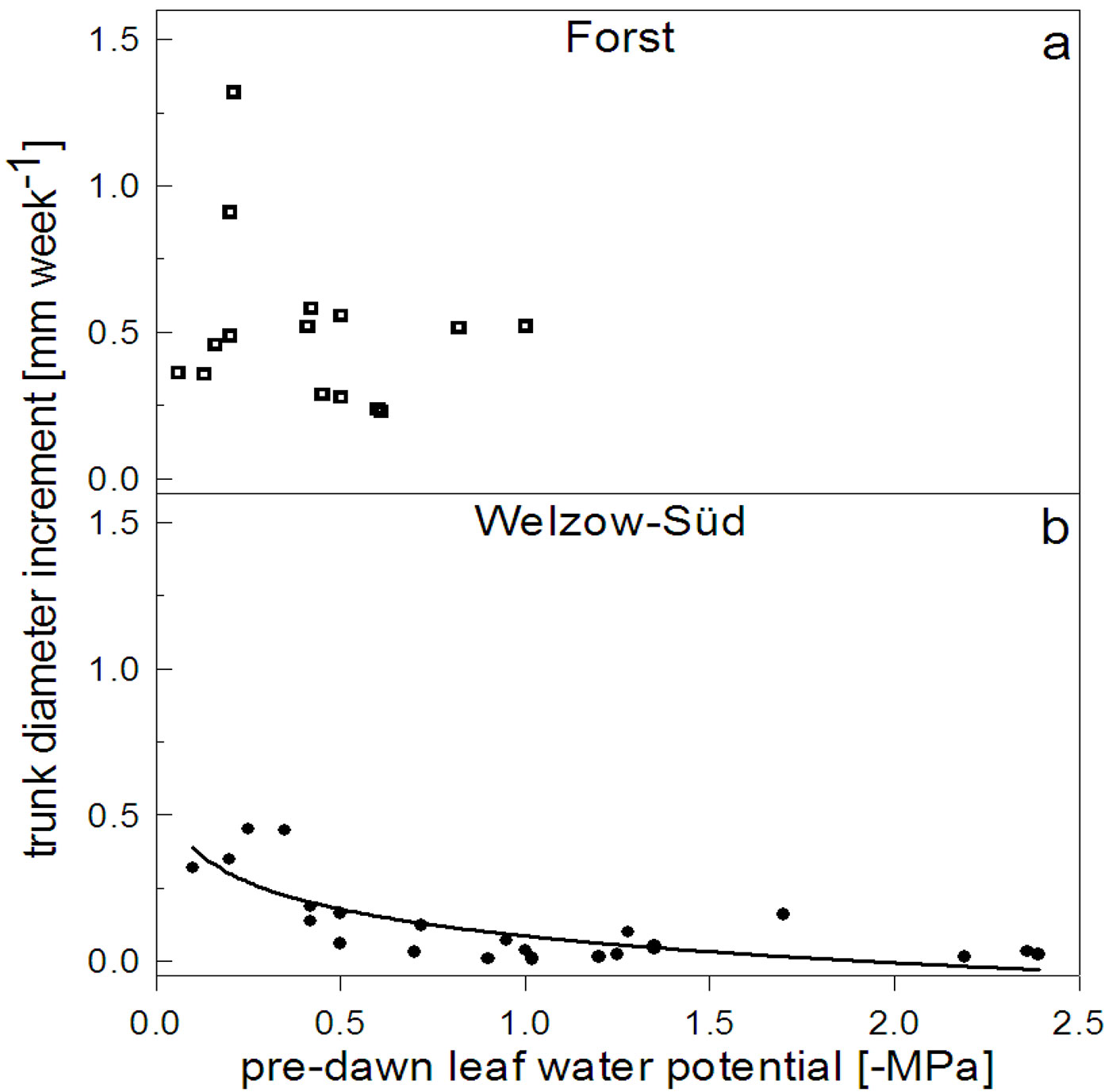
Weekly growth and plant water status
The plant water status (Ψpd) in the subplots F4 and W4 became more negative in the final stages of the experiment (Fig. 6). In August both sites faced a summer drought period, which affected the tree growth. The correlation between the weekly diameter increment rate and the Ψpd was significant (p = 0.689) only in Welzow-Süd, and their relation was better described by a logarithmic function (Y = -0.13·log(X) + 0.09, r2 = 0.60) with a drastic decrease of the diametric increment following the Ψpd reduction (Fig. 7b).
Fig. 6 - Pre-dawn leaf water potential and weekly diameter increase of trees in Forst (a) and Welzow-Süd (b) measured during the vegetation period (from June 1st to August 30th, 2012).

Fig. 7 - Weekly trunk diameter increase in relation to the pre-dawn leaf water potential of trees at the F4 subplot (Forst) (a) and W4 subplot (Welzow-Süd) (b) measured during the vegetation period (from June 1st to August 30th, 2012).
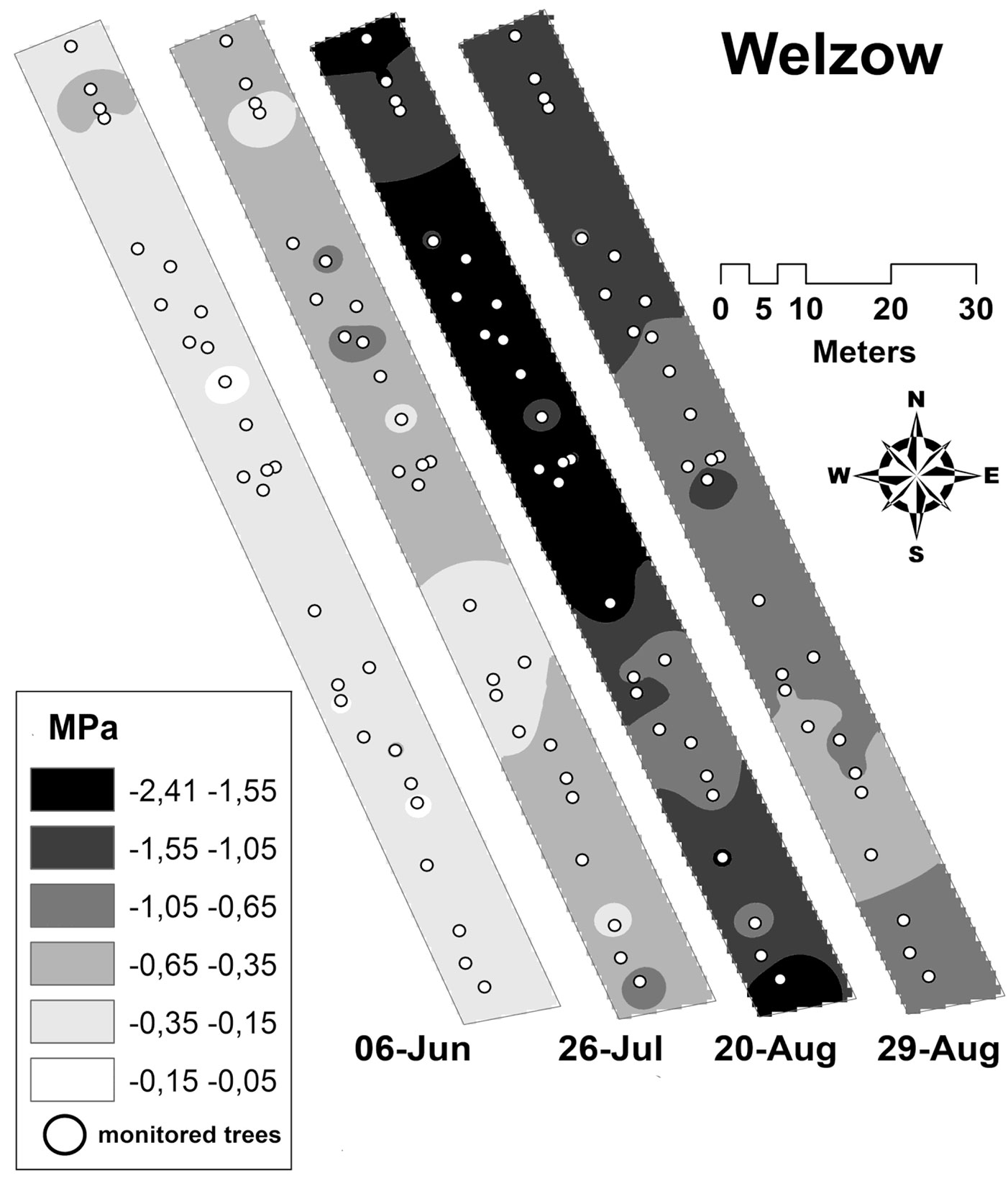
Spatial variation of pre-dawn water potential
Until July 25, 2015 for all the subplots measured at both sites the mean Ψpd values were > -0.5 MPa. At the Forst site, the mean Ψpd values decreased for all subplots until the mean minimum of -0.81 MPa was reached on August 24, and no significant differences between the subplots were found. Conversely, starting from August 14 the mean Ψpd decreased drastically at the Welzow-Süd site, reaching the minimum mean values of -1.56 MPa on August 24. Here, the differences between the subplots in terms of Ψpd became significant (p = 0.028), ranging from -2.06 MPa in subplot W4 to -0.96 MPa in subplot W6. The soil heterogeneity recorded at Welzow-Süd is well described by the spatial and temporal variation of the Ψpd evaluated at this site (Fig. 8).
Fig. 8 - Spatial interpolation of the pre-dawn leaf water potential values measured at Welzow-Süd using the Inverse Distance Weighted (IDW) technique. The values are measured periodically on single trees during the vegetation period (from June 1st to August 30th).
Discussion
Besides the difference in precipitation, which led to an increased water input in Forst, the different soil characteristics such as soil texture and TOC influenced the saturated hydraulic conductivity and the water holding capacity of the soil at both sites, enhancing its water storage capacity. The growth rate variation recorded at the two sites became relevant from July onwards, when the Ψpd reduction occurred in Welzow-Süd. Here the growth decrement was probably caused by the combined effects of the reduced precipitation and soil water storage, higher VPD values and low nutrient availability. Consequently, stomatal closure and the reduction of the leaf area under drought conditions limited the transpiration ([2], [11], [12]), and enabled the trees to tolerate long-term drought conditions due to its ecophysiological and morphological adaptations ([20], [14]). However, the reduction of the CO2 uptake had drastic effects on the plant growth and consequently on the biomass production, as reported by Mantovani et al. ([11], [12]), where the primary production was directly proportional to the water used, independently from the drought stress frequency and duration. Instead, water constraint caused changes in the soil-root and leaf-atmosphere interfaces. Consequently, CO2 uptake and transpiration decrease limited the tree growth, and in extreme cases the tree survival was at stake. The heterogeneity recorded in Welzow-Süd in terms of plant growth was related to the Ψpd and mirrored the heterogeneous soil property which caused the water availability variation in space and time, described by the dynamic maps in Fig. 8. During the night the water potential of a non-transpiring plant equilibrated with the wettest water potential of the soil around the root system ([13]). As recorded by Veste et al. ([15]), the spatial heterogeneity of soil water availability is linked with the pre-dawn water potential of trees under water limitation. In addition to water constraint, the indirect effect of the nutrient uptake reduction did occur, caused by the poor edaphic conditions at Welzow-Süd. However, since the diametric increment was higher in Forst than in Welzow-Süd at similar water potential values, other factors such as different tree ages and better soil conditions and nutrition may have contributed to increase the primary production in Forst. Robinia pseudoacacia is less dependent on the uptake of nutrients in comparison to other broad leaf trees because of its capability to fix biological nitrogen even under extreme edaphic conditions, as they are found in post-mining sites ([16]) or under drought conditions ([10], [18]). Nevertheless, Kanzler et al. ([8]) has observed for Robinia an increase of the growth rates after phosphate fertilization in Welzow-Süd. The fact that limited phosphate, in combination with low Molybdenum availability in the soil, has an impact on the biological nitrogen fixation was confirmed by Wurzburger et al. ([19]). Those differences in soil nutrients might partly explain the differences in the absolute growth rate between sites, while temporal and spatial differences within sites depend on the tree access to soil water.
Conclusions
The black locust growth performance is severely affected by the temporal and spatial variation of the leaf water potential, which varies according to the soil typology and water availability. Further nutrient deficiency may inhibit root expansion, causing the reduction of water uptake which indirectly influences the leaf water potential as well. If the water and nutrient uptake is limited due to critical edaphic conditions, which were site-specific in our experiment, the tree growth stops during a longer drought period. However, due to its morphological and ecophysiological adaptations R. pseudoacacia is able to cope with the extreme soil conditions in post-mining sites. Therefore, black locust could be an alternative species to be grown in post-mining areas, where the competition with food crops is reduced and soil protection and rehabilitation is the main goal. The spatial and temporal variation of the growth performance in such post-industrial areas can be simply quantified by using the pre-dawn leaf water potential method, evaluating at the same time the degree of soil heterogeneity in the area.
References
Gscholar
Online | Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Maik Veste
Christian Böhm
Dirk Freese
Brandenburg University of Technology Cottbus-Senftenberg, Institute of Soil Protection and Recultivation, Konrad-Wachsmann-Allee 6, D-03046 Cottbus (Germany)
CRA-OLI - Centro di Ricerca per l’Olivicoltura, v. Nursina 2, I-06049 Spoleto (PG - Italy)
CEBra - Centre for Energy Technology Brandenburg e.V., Friedlieb-Runge-Strasse 3, D-03046 Cottbus (Germany)
University of Bologna, Department of Agricultural Sciences, v. Fanin 44, I-40127, Bologna (Italy)
Corresponding author
Paper Info
Citation
Mantovani D, Veste M, Böhm C, Vignudelli M, Freese D (2015). Spatial and temporal variation of drought impact on black locust (Robinia pseudoacacia L.) water status and growth. iForest 8: 743-747. - doi: 10.3832/ifor1299-008
Academic Editor
Gianfranco Minotta
Paper history
Received: Mar 26, 2014
Accepted: May 15, 2015
First online: Jun 18, 2015
Publication Date: Dec 01, 2015
Publication Time: 1.13 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2015
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 56953
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 46627
Abstract Page Views: 4273
PDF Downloads: 4488
Citation/Reference Downloads: 26
XML Downloads: 1539
Web Metrics
Days since publication: 4058
Overall contacts: 56953
Avg. contacts per week: 98.24
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2015): 22
Average cites per year: 2.00
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Use of δ13C as water stress indicator and potential silvicultural decision support tool in Pinus radiata stand management in South Africa
vol. 12, pp. 51-60 (online: 24 January 2019)
Research Articles
Soil water deficit as a tool to measure water stress and inform silvicultural management in the Cape Forest Regions, South Africa
vol. 13, pp. 473-481 (online: 01 November 2020)
Research Articles
Effect of plant species on P cycle-related microorganisms associated with litter decomposition and P soil availability: implications for agroforestry management
vol. 9, pp. 294-302 (online: 05 October 2015)
Review Papers
Quantifying and modeling water availability in temperate forests: a review of drought and aridity indices
vol. 12, pp. 1-16 (online: 10 January 2019)
Research Articles
Spatial heterogeneity of soil respiration in a seasonal rainforest with complex terrain
vol. 6, pp. 65-72 (online: 07 February 2013)
Research Articles
Oak sprouts grow better than seedlings under drought stress
vol. 9, pp. 529-535 (online: 17 March 2016)
Research Articles
Effect of drought stress on some growth, morphological, physiological, and biochemical parameters of two different populations of Quercus brantii
vol. 11, pp. 212-220 (online: 01 March 2018)
Research Articles
Multi-temporal influence of vegetation on soil respiration in a drought-affected forest
vol. 11, pp. 189-198 (online: 01 March 2018)
Research Articles
Recovery of water potential and leaf gas exchange performance following drought stress in Quercus cerris populations
vol. 19, pp. 94-101 (online: 17 March 2026)
Research Articles
The responses of soil microbial community and enzyme activities of Phoebe zhennan cultivated under different soil moisture conditions to phosphorus addition
vol. 11, pp. 751-756 (online: 15 November 2018)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword






