
Three prescribed fire regimes on the restoration of flooded savannah grasslands under encroachment of Vochysia divergens Pohl, Pantanal, Brazil
iForest - Biogeosciences and Forestry, Volume 17, Issue 3, Pages 165-171 (2024)
doi: https://doi.org/10.3832/ifor4483-017
Published: Jun 17, 2024 - Copyright © 2024 SISEF
Research Articles
Abstract
The proliferation of woody plant species over grassland areas has been reported in different regions of the planet. This phenomenon has caused economic, social and environmental effects in the Brazilian Pantanal. Traditional knowledge based on scientific evidences technically supports the forms of management to control the process of colonization of woody plants in native grasslands, thus promoting the original ecological restoration of these ecosystems. Our study aimed to evaluate three management regimes with prescribed fire for controlling the invasion of Vochysia divergens Pohl. on grassland matrix areas in the Brazilian Pantanal. The results obtained showed that prescribed fire is an efficient tool in controlling V. divergens in the early stages of its establishment, and effectively contributes to ecological restoration processes in the savannah grasslands, aimed at controlling woody tree and shrub species invasions in the grasslands of the Brazilian Pantanal.
Keywords
Ecological Restoration, Wood-plant Encroachment, Prescribed Fire, Wetland, Natural Grassland
Introduction
The phenomenon of woody plant species proliferating over native grasslands is known worldwide ([30], [38], [15]), and is related to climate change and anthropogenic interventions, such as the conversion of natural areas for agriculture, mining and forest fires, and has caused economic damage with the reduction of grazing areas, as well as environmental damage, such as the increased risk of forest fires and the loss of biodiversity ([2], [1], [17]).
In the Brazilian Pantanal, the proliferation of Vochysia divergens Pohl. has occurred over areas of natural grassland, causing changes in the floristic composition and community structure of the grassland towards monodominant forests ([21]), resulting in a loss of biological diversity ([30], [23]). Economically, the livestock and tourism sectors are being particularly affected. The increased invasion of this woody species in pastures - as reported by farmers - results in productivity losses and management challenges, in addition to the loss of open environments and scenic landscapes, with negative impacts on ecotourism ([16]).
V. divergens is a tree species with a wide distribution in flooded areas in the Pantanal where multiannual flood pulses occur. After the long multi-year period of drought in the 1960s1970s, the super-humid period in the Pantanal from 1974 onwards favoured the expansion of woody plant species ([4]).
Although the effectiveness of prescribed fire in controlling woody vegetation in savannah ecosystems is known, this technique is only occasionally used in the ecological restoration of grasslands ([39]). Integrated fire management, which includes prescribed, planned, and monitored burning, has the central objective of a functional conservation of the ecosystem, and can promote the re-establishment of a native floristic composition ([27], [24], [8]).
Proactive control of invasive species in the early stages of intrusion is an important part of effective environmental management ([40]). However, the vast extension of Pantanal and the difficulty to access the areas due to traffic conditions, combined with the rapid dispersal of woody species, limit the capability of control, restore and re-establish the original functional identity in the grasslands.
Despite its potential, information on the use of fire as a management tool for restoring natural grasslands in the Pantanal is scarce, and the use of prescribed fire requires specific regulations and guidelines that are more suited to the reality of the Pantanal’s floodplain savannah ([27]). Indeed, regulations on the use of controlled fire in Brazil are still too general and unsuited to more complex ecosystems, such as seasonal wetlands, mainly because there are gaps in studies and research on the application of prescribed fire to these environments.
To fill these gaps, this study aimed at evaluating the efficiency of prescribed burning in controlling the intrusion of V. divergens as a tool to promote the ecological restoration of native grasslands in the Pantanal.
Materials and methods
The study was conducted in the Pantanal Floodplain, SE Brazil. Climate according to Koppen is Aw, tropical with a wet summer and dry winter season. The average temperature is 25.6 °C, ranging from a minimum of 15 °C to a maximum of 34 °C. Occasional extremes can reach 40 °C in summer and 0 °C in winter. Annual rainfall averages 2000 mm, concentrated during the summer ([11]), which is the period of flooding in the Pantanal ([22]).
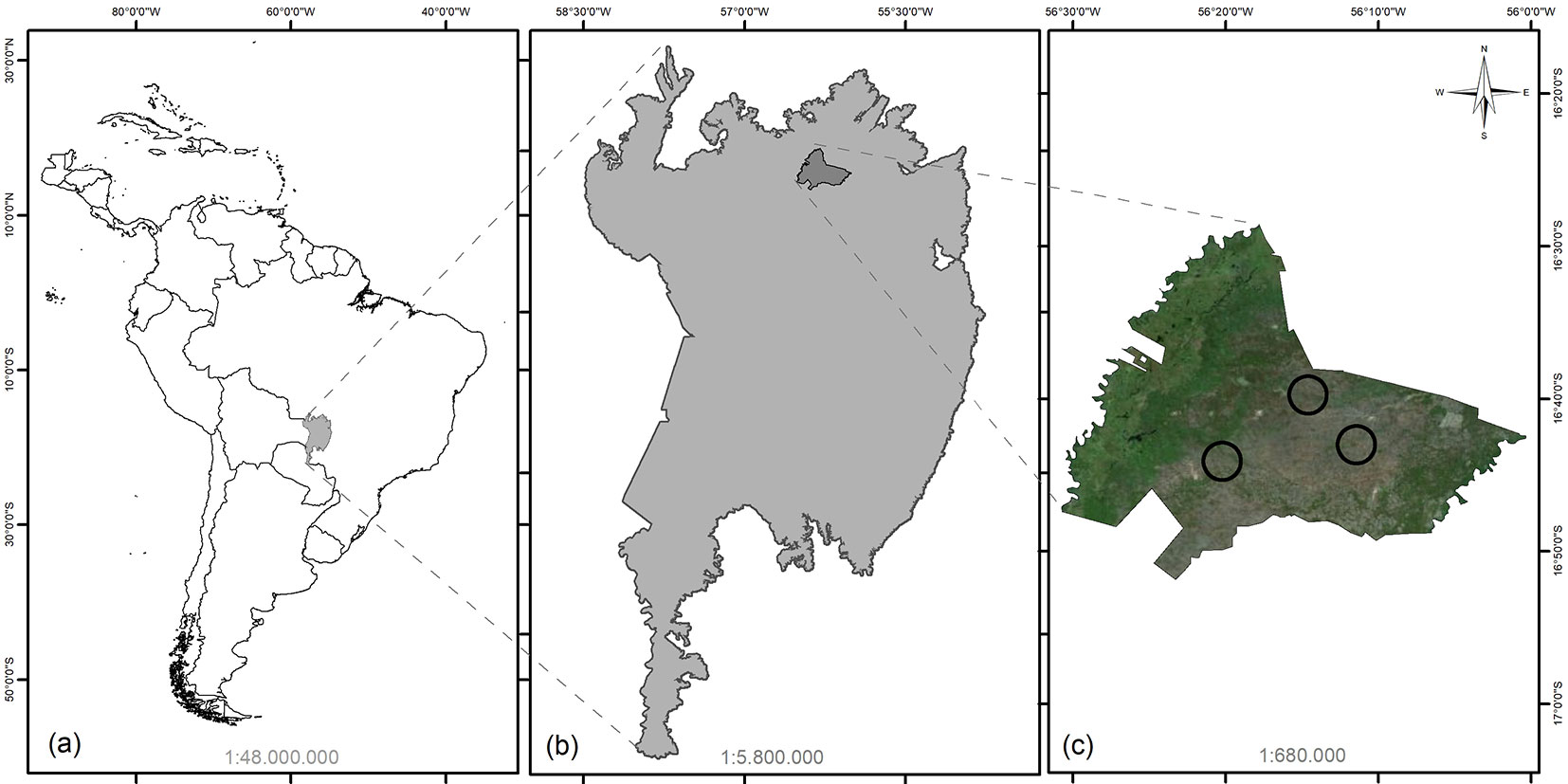
The study site is located in the SESC (Social Service of Commerce) Private Natural Heritage Reserve, in the Municipality of Barão de Melgaço, in the northern Pantanal of the State of Mato Grosso (Fig. 1). It is a Conservation Unit of Sustainable Use group, whose creation aims to conserve biological diversity.
Fig. 1 - Location of the study area, RPPN SESC Pantanal, in relation to South America, and inserted in the northern region of the Pantanal in Brazil. The black circles represent the regions where sampling was carried out.
Sample design
The experiment was set up in three blocks of savannah wetlands with V. divergens invasion. In each block, four 1-hectare plots (100 × 100 m) were delineated, with a minimum distance of 200 m each other. Each plot was subject to a different treatment: (i) Plot 1 - control plot, where no prescribed burning was applied; (ii) Plot 2 - early, burning applied in July; (iii) Plot 3 - modal, burning carried out in September; (iv) Plot 4 - late, burning carried out in November. This sampling design ensures 3 replications per treatment, summing up to 4 ha each and 12 ha of study area in total.
Pre-burn data collection
Quantitative data was collected by measuring only individuals of V. divergens with a height ≥ 1 m. Circumference at ground height was recorded for individuals with a total height ≤ 2.0 m and the circumference at breast height for trees with height > 2.0 m.
The total height of the largest individuals was taken using an electronic clinometer, and in cases where it was not possible to fully visualise the tree, the height was estimated by visual calibration ([37]). To validate the height estimates, we performed a correlation analysis between tree diameter and height using the linear regression model of the hypsometric relationship proposed by Curtis ([6]).
Qualitative data was obtained by analysing the individuals directly. Information was collected on stem shape (straight, slightly crooked and crooked) and health (healthy, slightly affected, and affected by fire).
All the qualitative and quantitative information obtained was recorded in the field using spreadsheets. Photographic and vector records were also made, such as the geographical location points of all the inventoried individuals, plot boundaries, and walks taken, using the Avenza Maps® app (Toronto, Canada), aided by pre-prepared maps with rasters and vectors georeferenced in the UTM Projection System to datum WGS84, thus enabling the geographical location of each individual plant ([5]).
Application of prescribed burning treatments
The prescribed burns were carried out in three periods during the year: (i) Treat_1 - at the end of the rainy season (July); (ii) Treat_2 - at the beginning of the dry season (September); and (iii) Treat_3 - at the beginning of the rainy season (November).
Each plot was burnt in a “U” shape, with the fires initially started against the wind, which is the method traditionally used for controlled burning in pastures. It provides greater safety during controlled burns and allows wildlife to escape.
Post-burn data collection
Thirty days after the burning, all the plots were revisited and a new qualitative survey was carried out, accessing all the geolocated individuals on a very high spatial resolution image map, which allowed to pinpoint all the inventoried individuals.
Fire impact was categorized by the degree to which the flames and heat had hit the V. divergens trees. The percentages of the crown affected and the fire damage to each stem were assessed ([32], [14]).
To assess crown damages, the percentage of burnt leaves, branches and shoots was estimated, taking a distance between the observation point and the individual tree so that the entire tree could be seen at a glance ([32], [25], [33]). We used an electronic clinometer to directly measure the percentage of the living crown in relation to the crown affected by burning. The condition of the crowns affected by flames is a direct indicator of the tree damage. The yellowing, reddening, or blackening of the leaves indicates the physiological effects of leaf senescence and loss of vigour, which can lead to mortality.
The extent of damage to the crown of a tree can be determined by comparing the green and the dead part after the exposure to flames. If the dead part exceeded 70% of the whole crown, this indicates a “severe” tree damage, from 50 to 70 per cent “medium” damage, and below 50 per cent “light” damage.
To assess the damage to tree trunks, the percentage of burned stem was estimated. We then focused on those trees that were most severely affected and proceeded to make precise cuts - or debarking - on the rhytidome. This allowed us to observe the tissues and evaluate the extent of damage in the neck region and at breast height. Following a macroscopic approach, we were able to verify the depth of combustion and ascertain the level of tissue damage. If combustion only affected the epidermis (outer bark), the injury was considered as “mild”; when combustion only affected the periderm (inner bark), the injury was “moderate”; and if combustion affected the meristematic cambium, it was considered as a “severe” injury.
We gauged the damage to the crown and stem based on the severity index, where (i) individuals with slight burning of the rhytidome and no burning of the crown were classified as having “mild severity” damage; (ii) individuals that suffered burning of up to 40% of the crown and burning of the stem slightly affecting the meristematic region of the cambium, as having “moderate severity” damage; (iii) when more than 40% of the crown was burnt, also affecting the tissues of the crown, as having “moderate severity” damage; and (iv) when burning affected more than 40% of the crown and the tissues of the meristematic region of the inner bark, as having “strong severity” damage.
Statistical analyses
The above data was recorded in electronic spreadsheets, generating a database that was statistically analysed using the software R version 3.0.1.
To validate the estimates of the total height of the largest individuals, we tested the correlation between the diameter and height by adjusting the hypsometric model of Curtis ([6]). The statistical criteria to adjust the coefficient of determination (R² adj), the standard error of the estimate (Syx%), and the regression coefficient were calculated.
To analyse the behaviour of the V. divergens population, we grouped the trees by size as described by Soares et al. ([37]). The size classes defined were: (i) class D_I, individuals with DBH < 0.5 cm; (ii) class D_II, individuals with DBH between 0.5 and 20.0 cm; and (iii) class D_III: individuals with DBH > 20.0 cm.
To check the statistical assumptions, we tested the absence of multicollinearity between the predictor variables (treatments, diameter classes, height classes) and between the dependent variables (the severity categories), using the “pairs.panels” function of the “psych” package in R software.
The severity categories were based on the three individual damage classes (see above): “mild severity”, “moderate severity”, and “strong severity”, with “mild severity” established as the reference category. We used the “multinom” function from the “nnet” package in R to build the statistical model, which was adjusted using an ANOVA against a null model (mod_0).
We obtained the coefficients and standard errors for the independent variables by carrying out two different regressions using the multinomial model: the first comparing “moderate severity” with the reference category “mild severity”, and the second comparing “high severity” with “mild severity”, generating two sets of coefficients and standard errors.
Results
A total of 822 individuals of V. divergens were inventoried in all the sampling areas. The absolute density recorded in the study area was 59.5 individuals per ha (Tab. 1).
Tab. 1 - Distribution of the inventoried individuals of V. divergens in the classes of the predictor variables (Treatment - Treat_1: July; Treat_2: September; Treat_3: November; Diameter - D_I: < 0.5 m; D_II: 0.5 to 20.0 cm; D_III: > 20.0 cm; Height - H_I: < 2.8 m; H_II: 2.8 to 5.0 m; H_III: > 5.0 m) and in the severity categories (response variable - see text).
| Treatment Class (=n) |
Diameter Class (=n) |
Height Class (=n) |
Severity Category (=n) |
|---|---|---|---|
| Control = 104 | D_I = 376 | H_I = 376 | Mild = 335 |
| Treat_1 = 106 | D_II = 271 | H_II = 361 | Moderate = 175 |
| Treat_2 = 356 | D_III = 175 | H_III = 85 | Strong = 312 |
| Treat_3 = 256 | - | - | - |
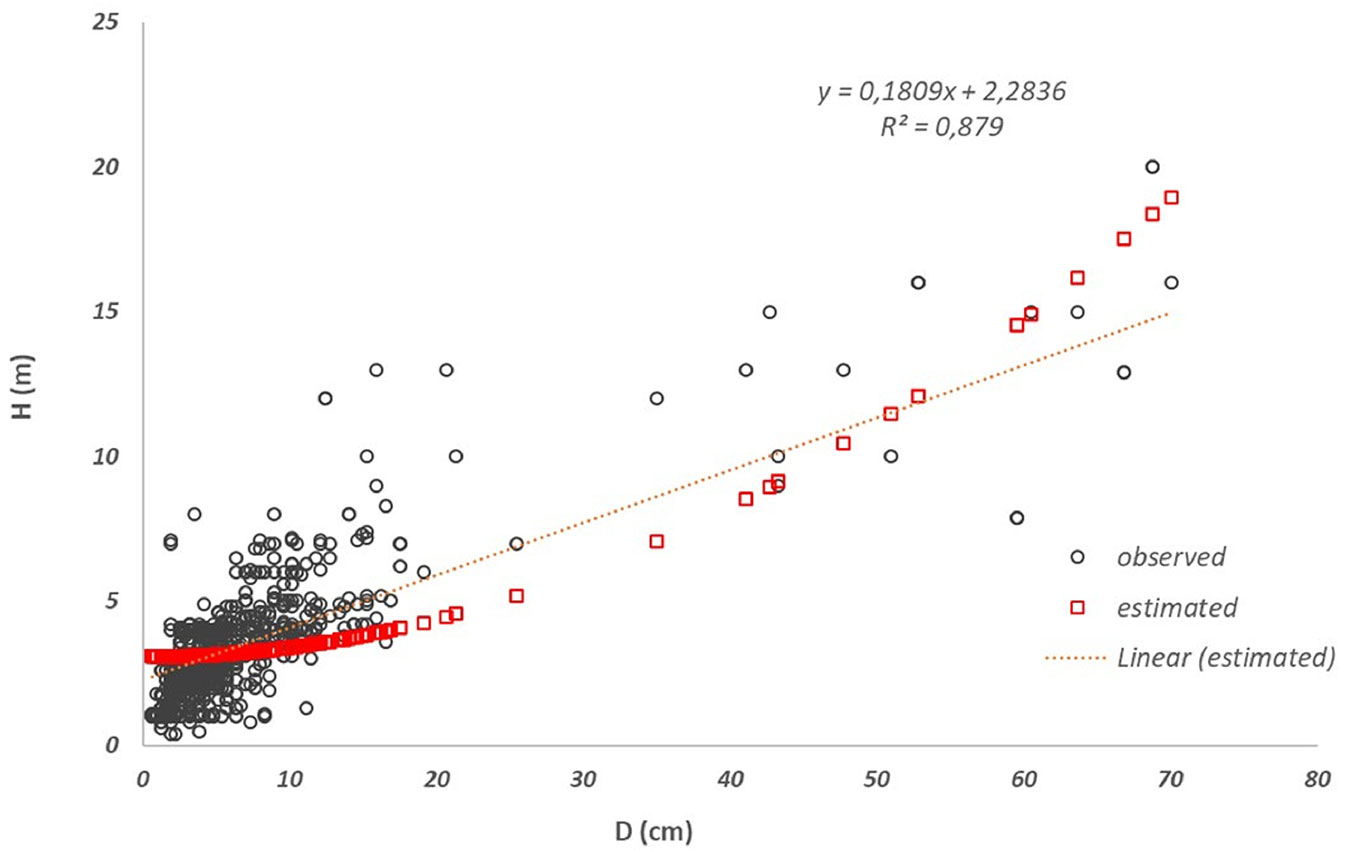
Height estimates using the hypsometric relationships showed a high correlation with the observed tree diameter and height (r = 0.7122). The coefficient of determination obtained for the regression was R² = 0.879 (Fig. 2).
Fig. 2 - Scatter plot of the observed heights and diameters of the V. divergens trees (black dots), and the height estimates (red) obtained using the linear hypsometric model by Curtis ([6]).
The calibration of the height estimates carried out using an electronic clinometer showed an accuracy of 95% and a sampling error of 4.6%.
Analysing qualitative data
The diameter class D_II (DBH = 0.5-20 cm) showed all the three levels of mild, moderate and severe damage caused by burning, and therefore served as a reference standard for damage to the stems.
The highest severity was recorded in the smallest individuals belonging to the diameter class D_I, resulting in a total of 249 individuals with strong damages. Class D_II followed with a total of 111 individuals with moderate severity and 97 with strong severity damages, and D_III (larger trees) with 92 individuals showing moderate severity damages (Fig. 3).
Fig. 3 - Examples of injuries after prescribed burning. Individuals in Diameter Class II were established as reference standards for stem injury analysis: (a) mild injury; (b) moderate injury; (c) severe injury. Crown damage, evaluated as percentage of leaves affected by the heat: (d) mild damage (<30% of leaves affected); (e) moderate damage (30-70%); (f) severe damage (≥70%).

When the relationship between the degree of severity and the period of prescribed burning is considered, the results showed that the treatment Treat_2 (modal burning - September, beginning of the dry season) had the largest number of V. divergens individuals in the “strong severity” damage class (195 individuals), followed by Treat_3 (late burning, November - 111 individuals) and Treat_1 (early burning, July - 54 individuals). As for the “moderate severity” damage class, again the highest number of individuals was found in the class Treat_2 (modal burning: 133), followed by Treat_3 (late burning: 101) and Treat_1 (early burning, 44 individuals).
The result for the fit of the multinomial model showed an associated p-value of 0.0000015, indicating a strong statistical difference from the null model (mod_0). Indeed, the results show that there was a significant effect of the predictor variables on the response variable. For the model tested, we obtained a pseudo R² = 0.6111 using the Nabelkerke method ([20]).
To adjust the model developed using the response variable “Severity”, and the predictors “Treatments”, “Diameter Class” and “Height Class”, we created a null model (M0) without independent variables, which was compared with the constructed model (Mod) using an ANOVA, in order to analyse the overall effects of the independent variables. We found that all the predictors (i.e., the independent variables) were statistically significant. To analyse the specific effects of each independent variable on the dependent variable, we applied the z-test and obtained the statistical coefficients reported in Tab. 2.
Tab. 2 - Analysis of the specific effects of the independent variables (predictors) in relation to the response variable, expressed through the statistical coefficients (Z-test). (*): p<0.05; (**): p<0.01; (***): p<0.001.
| Group | Predictors | Estimate | Std. Error | Z value | Pr (>|Z|) |
|---|---|---|---|---|---|
| II | (Intercept) | -1196.501 | 4392.569 | -0.2724 | 0.785321 |
| Treat_1 | 1467.076 | 4392.575 | 0.3440 | 0.738387 | |
| Treat_2 | 1495.084 | 4392.484 | 0.3404 | 0.733575 | |
| Treat_3 | 1271.196 | 4392.373 | 0.2894 | 0.772268 | |
| D_II | -1.28918 | 0.39790 | -32.399 | 0.001196 ** | |
| D_III | -224.354 | 0.50656 | -44.290 | 9.469e-06 *** | |
| H_I | 0.39509 | 0.50073 | 0.7890 | 0.430094 | |
| H_II | 0.95822 | 0.40941 | 23.405 | 0.019258 * | |
| III | (Intercept) | -1666.131 | 0.51021 | -326.555 | < 2.2e-16 *** |
| Treat_1 | 2047.299 | 0.41020 | 499.096 | <2.2e-16 *** | |
| Treat_2 | 2127.255 | 0.28644 | 742.650 | <2.2e-16 *** | |
| Treat_3 | 1756.625 | 0.25716 | 683.074 | <2.2e-16 *** | |
| D_II | -308.714 | 0.41756 | -73.933 | 1.433e-13 *** | |
| D_III | -586.076 | 0.59825 | -97.965 | < 2.2e-16 *** | |
| H_I | 0.85534 | 0.62226 | 13.746 | 0.169263 | |
| H_II | 142.637 | 0.55660 | 25.627 | 0.010387 * |
The independent variables marked as Group II in Tab. 2 were interpreted by comparing the values of the coefficients obtained for moderate severity with the reference value for mild severity, while the variables marked as Group III were interpreted by comparing the coefficients obtained for severe severity with mild severity. The coefficients showing p-values > 0.05 were not statistically significant (Tab. 3). The diametric classes D_II and D_III had negative coefficients (-1.28 and -224.35, respectively), indicating a lower chance of showing strong severity damages when compared to mild severity category. On the other hand, the height classes showed positive coefficients, with the low (H_I) and medium (H_II) height classes being respectively 0.39 and 0.95 times more likely to show moderate severity damages compared to mild severity class. While the low (H_I) and the medium (H_II) height classes were 0.85 and 142.637 times more likely to show strong severity damages compared to the mild severity category.
Tab. 3 - Odd ratios for the coefficients obtained using the exponential function, with confidence interval, showing that the odd ratios are statistically significant when they differ from 1. The results are shown for two regressions using the multinomial model, one comparing “moderate severity” with the reference category “mild severity” (upper section), and a second comparing “high severity” with “mild severity” (lower section), generating two sets of coefficients and standard errors, with the p-values showing the significance of the variables. Model: (Multinomial = Severity ~ Treatments + ClassD + ClassH).
| Severity | Classes | Odds Ratio | 95% Confidence Interval |
p-value | |
|---|---|---|---|---|---|
| Moderate | Treatments | Control | - | - | - |
| Treat_1 | 2,351,953 | 0.00; 57.69 · 1042 | 0.7 | ||
| Treat_2 | 3,112,192 | 0.00; 76.20 · 1042 | 0.7 | ||
| Treat_3 | 331,692 | 0.00; 8.10 · 1042 | 0.8 | ||
| Diameter | D_I | - | - | - | |
| D_II | 0.28 | 0.13; 0.60 | 0.001 | ||
| D_III | 0.11 | 0.04; 0.29 | <0.001 | ||
| Height | H_I | 1.48 | 0.56; 3.96 | 0.4 | |
| H_II | 2.61 | 1.17; 5.82 | 0.019 | ||
| H_III | - | - | - | ||
| High | Treatments | Control | - | - | - |
| Treat_1 | 1,732.020,979 | 987,947,439; 3,036,494,200 | <0.001 | ||
| Treat_2 | 778,533,565 | 348,429,002; 1,739,563,893 | <0.001 | ||
| Treat_3 | 42,552,684 | 25,705,604; 70,441,095 | <0.001 | ||
| Diameter | D_I | - | - | - | |
| D_II | 0.05 | 0.02; 0.10 | <0.001 | ||
| D_III | 0.00 | 0.00; 0.01 | <0.001 | ||
| Height | H_I | 2.35 | 0.69; 7.96 | 0.2 | |
| H_II | - | - | - | ||
| H_III | 4.16 | 1.40; 12.4 | 0.010 | ||
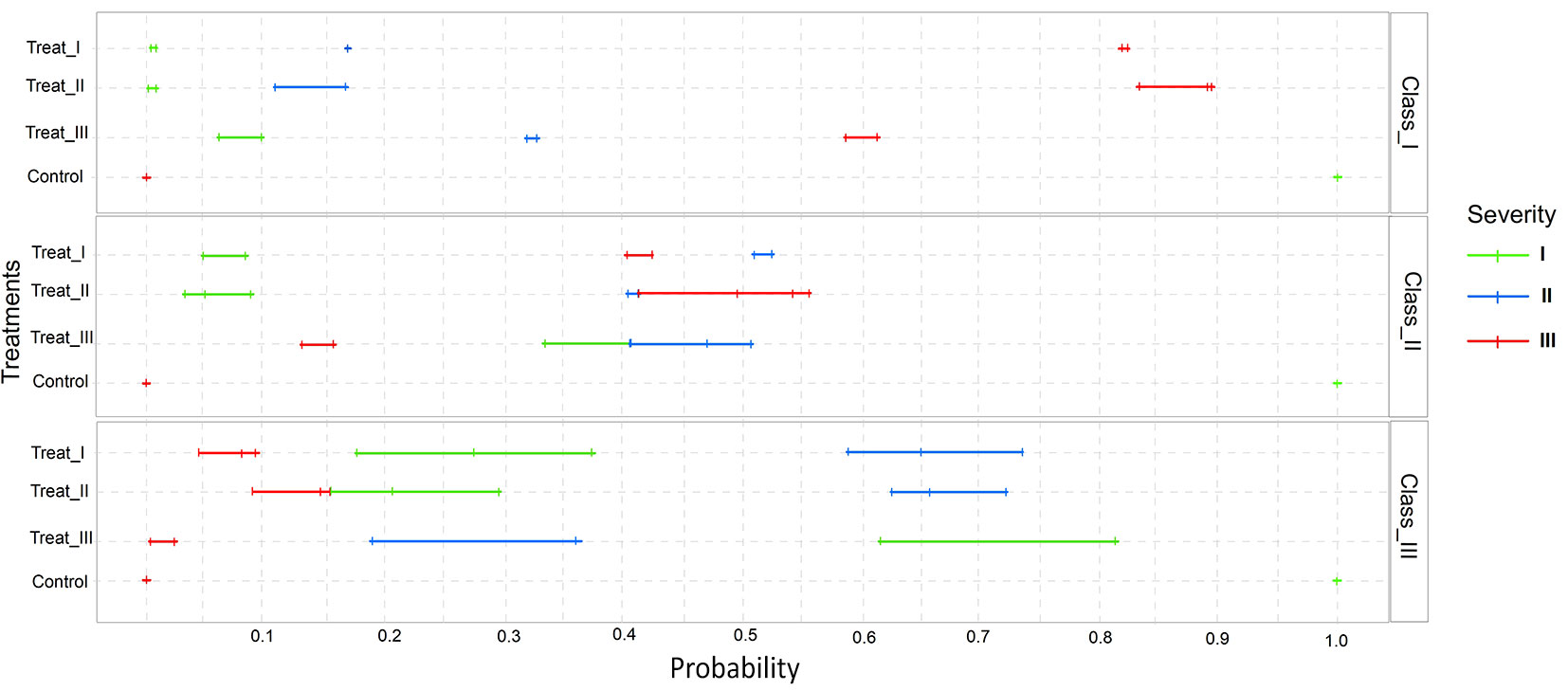
We illustrate our results with a graph from the multinomial logistic regression analysis (Fig. 4).
Fig. 4 - Multinomial logistic regression analysis graph for the proposed model. The predictor variables are Treatment, Diametric Class and Height Class, and the response variable is Prescribed Burning Severity. The probability of the effects of the predictor variables on the response variable is expressed on the x-axis.
Discussion
The studied population of Vochysia divergens shows the classic model of establishment of woody plants through the native grasslands in the Pantanal, with a steadily growing abundance in ecosystems such as grasslands and savannas in both terrestrial and wetland environments ([7], [34]).
The observed diametric distribution of V. divergens in this study is fairly typical of tree populations colonizing an inhospitable environment. According to Silva Junior ([36]), this can be observed in self-regenerating species and occurs when smaller individuals successively replace adult individuals in the population. Theoretically, controlling and eradicating the invasive species while the population is still in the initial establishment process is an effective way of avoiding dominant monospecific formations in native grasslands.
Studies on the effects of prescribed fire on woody species has been carried out in different regions of the world, with a predominance of studies on coniferous species in temperate regions ([26]). In the tropical region, the effects of fire on hardwood species have been investigated in forest ecosystems in the Amazon ([29], [18]) and in the Cerrado ([19], [31]). However, due to the great diversity of species in tropical zones - and especially in the Pantanal -there are still major gaps in understanding the effects of fire.
We found that young individuals of V. divergens were especially affected by severe damages when prescribed fire is applied in September (Treat_2), which is the driest period in the study area, indicating the effectiveness of this practice in the control and eradication of that species. Indeed, Nunes da Cunha & Junk ([22]), Santos et al. ([35]), and Pott et al. ([28]) mentioned that V. divergens is a wetland species especially sensitive to fire.
According to our results, both tops and stems of the smallest individuals were severely damaged by all treatments. Fire application in September (Treat_2) showed the greatest severity, resulting in a high mortality rate of saplings and seedlings of V. divergens, confirming the findings by Franco et al. ([10]) and Hoffmann ([12]). The impact on reproductive efforts due to the mortality of the aerial part of trees restricts the entry of individuals into the population ([13]).
The greater severity of burns in smaller individuals is associated with more intense exposure to flames and heat in the lowest layer, where the combustible load accumulates. Even with controlled fires, where heat production is not as intense as in wildfires, serious burns and/or total combustion of individual trees often occur. This was expected, as controlled fires were aimed to reduce the biomass of trees and the fuel load, thus favouring the growth of native herbaceous species.
We also observed that severe injuries in the neck region, where the rhytidome is thicker, resulted in higher mortality of individuals in all treatments. Bark thickness is an important factor in predicting the effects of fire on tree populations ([41]), and there is a direct correlation between diameter and bark thickness. The larger the individual, the thicker the bark, and consequently the greater the barrier to heat transfer. In the case of large individuals with burning in the form of ringing (i.e., deep lesions that encircled the stem), the mortality rate was higher. Bova & Dickinson ([3]) report that the meristematic region of the vascular cambium in angiosperms is ring-shaped, and if the fire reaches the rhytidome in such a way as to encircle the trunk, it can cause ringing which result either in the death of trees or the partial necrosis of the cambium. The mortality of larger trees, due to the direct effect of heat, can occur up to three years after the fire ([32], [14]).
Another important factor observed is the burning of the tips, the region of the apical meristem of the individuals. However, basal aerial shoots appeared at different heights on the stems of survivors. These shoots end up altering the architecture of the tree canopy, which can lead to a larger crown shading the understory, thus hampering the growth of native herbaceous species. Moreover, the colonization of meadows by woody species like V. divergens leads to a reduction in herbaceous biomass and a consequent reduction in combustible material, making burning less subsequent and less intense ([30]).
For tropical regions, tree mortality assessed through severity of damage in the cambial region of the stems or in combination with crown damage requires precise assessments, taking into account the anatomical, morphological and ecophysiological variability of the species. The methodology adopted in this study sought references in methodologies that have been used to quantify cambial death, ranging from direct sampling of cambial tissue ([32], [25], [33]) to the thickness of charring of the rhytidome ([9]).
Conclusions
Based on our findings, we conclude that the colonization of the invasive species Vochysia divergent in flooded savannah areas in the Pantanal can be effectively controlled through periodical prescribed fires, which eliminate young individuals and help preserve and restore this type of native grassland.
The most effective period for prescribed fire to control the proliferation of V. divergens was during the driest part of the year, in September. However, controlled burning must be exclusively used during periods permitted by law and always supported by an efficient fire prevention and firefighting infrastructure.
The application of prescribed fire as a tool for restoring savannah grasslands should be carefully evaluated in an ecological context, looking at the impacts of its use on the local biological diversity. Controlling invasive seedlings, saplings and trees may be the best way of preserving and restoring the Pantanal’s natural grasslands, and this can include the use of burning, as long as it is used in a correct manner, taking into account the technical parameters of integrated management and the level of stabilisation of the species’ population.
Acknowledgements
This work was supported by the project “Assessment of the effect of fire on biodiversity and soil, contributions to the establishment of Integrated Fire Management in the Pantanal” (012/2020 SEMA-MIF PANTANAL), and the project “Pantanal Research Network” from the Ministry of Science and Technology of Brazil (FINEP 01.20.0201.00).
Special thanks to: National Wetlands Institute; SESC - Pantanal Socio-Environmental Pole; Mato Grosso State Secretariat for the Environment; Chico Mendes Institute for Biodiversity Conservation.
References
CrossRef | Gscholar
CrossRef | Gscholar
Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Authors’ Info
Authors’ Affiliation
Postgraduate Programme in Ecology and Biodiversity Conservation - PPGECB, Federal University of Mato Grosso - UFMT, Cuiabá - Mato Grosso (Brazil)
National Centre for Research and Conservation of Carnivorous Mammals, Chico Mendes Institute for Biodiversity Conservation, Municipal Road Hisaichi Takebayashi, 8600, Usina neighbourhood, Atibaia 12952-011, SP (Brazil)
National Institute of Science and Technology in Wetlands - INCT/INAU, Cuiabá - Mato Grosso (Brasil)
Corresponding author
Paper Info
Citation
Ebert A, Berlinck CN, Nunes da Cunha C (2024). Three prescribed fire regimes on the restoration of flooded savannah grasslands under encroachment of Vochysia divergens Pohl, Pantanal, Brazil. iForest 17: 165-171. - doi: 10.3832/ifor4483-017
Academic Editor
Giorgio Vacchiano
Paper history
Received: Oct 08, 2023
Accepted: Jun 05, 2024
First online: Jun 17, 2024
Publication Date: Jun 30, 2024
Publication Time: 0.40 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2024
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 13765
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 8607
Abstract Page Views: 2484
PDF Downloads: 2343
Citation/Reference Downloads: 1
XML Downloads: 330
Web Metrics
Days since publication: 737
Overall contacts: 13765
Avg. contacts per week: 130.74
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
(No citations were found up to date. Please come back later)
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Tree encroachment dynamics in heathlands of north-west Italy: the fire regime hypothesis
vol. 3, pp. 137-143 (online: 27 September 2010)
Research Articles
Post-fire recovery of the plant community in Pinus brutia forests: active vs. indirect restoration techniques after salvage logging
vol. 11, pp. 635-642 (online: 04 October 2018)
Review Papers
Prescribed burning in Italy: issues, advances and challenges
vol. 6, pp. 79-89 (online: 07 February 2013)
Review Papers
Post-fire soil hydrology, water erosion and restoration strategies in Andosols: a review of evidence from the Canary Islands (Spain)
vol. 9, pp. 583-592 (online: 17 March 2016)
Research Articles
Post-fire effects and short-term regeneration dynamics following high-severity crown fires in a Mediterranean forest
vol. 5, pp. 93-100 (online: 30 May 2012)
Research Articles
Historical fire ecology and its effect on vegetation dynamics of the Lagunas de Montebello National Park, Chiapas, México
vol. 14, pp. 548-559 (online: 01 December 2021)
Research Articles
Differences of fire activity and their underlying factors among vegetation formations in Greece
vol. 6, pp. 132-140 (online: 08 April 2013)
Technical Reports
Post-fire forest management in southern Europe: a COST action for gathering and disseminating scientific knowledge
vol. 3, pp. 5-7 (online: 22 January 2010)
Research Articles
Near zero mortality in juvenile Pinus hartwegii Lindl. after a prescribed burn and comparison with mortality after a wildfire
vol. 12, pp. 397-402 (online: 31 July 2019)
Technical Notes
Goat grazing as a wildfire prevention tool: a basic review
vol. 7, pp. 260-268 (online: 26 March 2014)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







