
Energy production of poplar clones and their energy use efficiency
iForest - Biogeosciences and Forestry, Volume 7, Issue 3, Pages 150-155 (2014)
doi: https://doi.org/10.3832/ifor0978-007
Published: Jan 23, 2014 - Copyright © 2014 SISEF
Research Articles
Abstract
The poplar clones of Robusta and I-214 (Populus × euramericana) are characterized by fast growth and high quality biomass production. The calorific values for these poplar clones were determined, and modelling of calorific production of the entire poplar clone stands was performed. This research, dependent on age and site indexes, was conducted in four localities in Eastern Slovakia and seven in the western region of this country. Samples of bark, wood and small-wood with bark were excised from three tree stem areas and from two parts of the crown. No significant difference was observed between the two clones, with regard to either the biomass fraction or the location of the excised sample on the tree. The average calorific capacity of all biomass fractions was approximately 6.86-8.73 GJ m-3, with the lowest bark values emanating from the stem base. The mean annual calorific energy production for site indexes 40-20 after 17-26 years growth was 320-80 GJ ha-1 a-1, and this was calculable 2-3 years earlier in the I-214 clone than in Robusta. The energy use efficiency of these poplar clones was estimated at a maximum of 0.4-1.6% solar radiation utilization.
Keywords
Biomass, Calorific Value, Energy Use Efficiency, Poplar Clones, Populus × euramericana
Introduction
A fairly large portion of agricultural lands in Europe is suitable for plants of fast-growing tree species. Poplars have proven their efficacy in restoring functional and structural attributes of agro-ecosystems ([2], [16], [17]). Since this species produces valuable biomass for energy production ([3]), it is necessary not only to know the calorific value of its individual components, but also to understand the formation processes occurring throughout the tree existence. Poplar clones have a great advantage over other woody species because they produce a large amount of above-ground biomass during their relatively short life-cycle. Solar energy drives photosynthetic assimilation stored as chemical bonds in the biomass structural components, though the accumulated energy content can remarkably vary, primarily depending on species growth over a determined time period. Moreover, stand age and site index affect the biomass production of forest tree species, influencing the obtainable volume and yield.
Yield tables for “Robusta” and “I-214” poplar clones published by Petráš & Mecko ([12]) predict not only biomass volumes for whole trees, but also for their main parts including stem, large wood, under-bark components, etc. Such partition of the whole yield in its different fractions is essential for industrial processing purposes. The volume obtained from yield tables is then converted to weight units based on the density of its different components. Many studies have reported that bark has lower density than wood ([6], [14], [5]), and soft broadleaf species have registered the lowest wood density, followed by coniferous and hard broadleaf species. Petráš et al. ([13]) derived average densities of above-ground biomass fractions in the range of 350-470 kg m-3 for two clones of Populus × euramericana, and a similar wood density of 306-367 kg m-3 was recorded for 4 clones of Populus × deltoides by Kord & Samdaliri ([7]). In addition, wood density varies not only according to species considered but also to the excising location along the tree ([14], [4], [13]).
Besides the evaluation of a species’ calorific value, it is important to assess the relative efficiency of woody species in using the solar radiation. Pretzsch ([15]) named this characteristic “energy use efficiency”, defined as the ratio between the energy accumulated in the above-ground biomass and the incoming solar radiation. For example, the average annual sum of global radiation for Germany is approximately 36 000 GJ ha-1 y-1. Since net biomass production for Norway spruce, European beach, Scots pine, Sessile and Common oak is in the range 0.005-0.009, their energy use efficiency is less than 1% ([15]). Indeed, Larcher ([9]) and Körner ([8]) reported that 50-60% of incoming radiation is outside the photosynthetically useful spectrum, and a further 10-20 % is reflected or transmitted, while most of the remainder is lost in the photosynthetic process.
The aims of this analysis were: (i) to determine the calorific values of above-ground biomass for poplar clones “Robusta” and “I-214”; (ii) to examine the above-ground biomass of wood, bark and small-wood with bark in order to establish which fraction has the highest calorific capacity; (iii) to derive the calorific energy content and energy use efficiency of clones at the stand level; and (iv) to establish the most appropriate exploitation strategy of poplar clones for energy production purposes.
Materials and methods
Study sites
Eleven trees were sampled in poplar stands located in the southern area of western (7 stands) and eastern (4 stands) Slovakia, at elevations ranging from 100 to 300 m a.s.l. (Fig. 1). Average annual temperature in these regions is 9.0-10.0 °C, and vegetation period is approximately 180 days. Most poplar stands were affected by ground water from surrounding rivers.
Fig. 1 - Location of study sites in western and eastern Slovakia, indicated by black crosses.
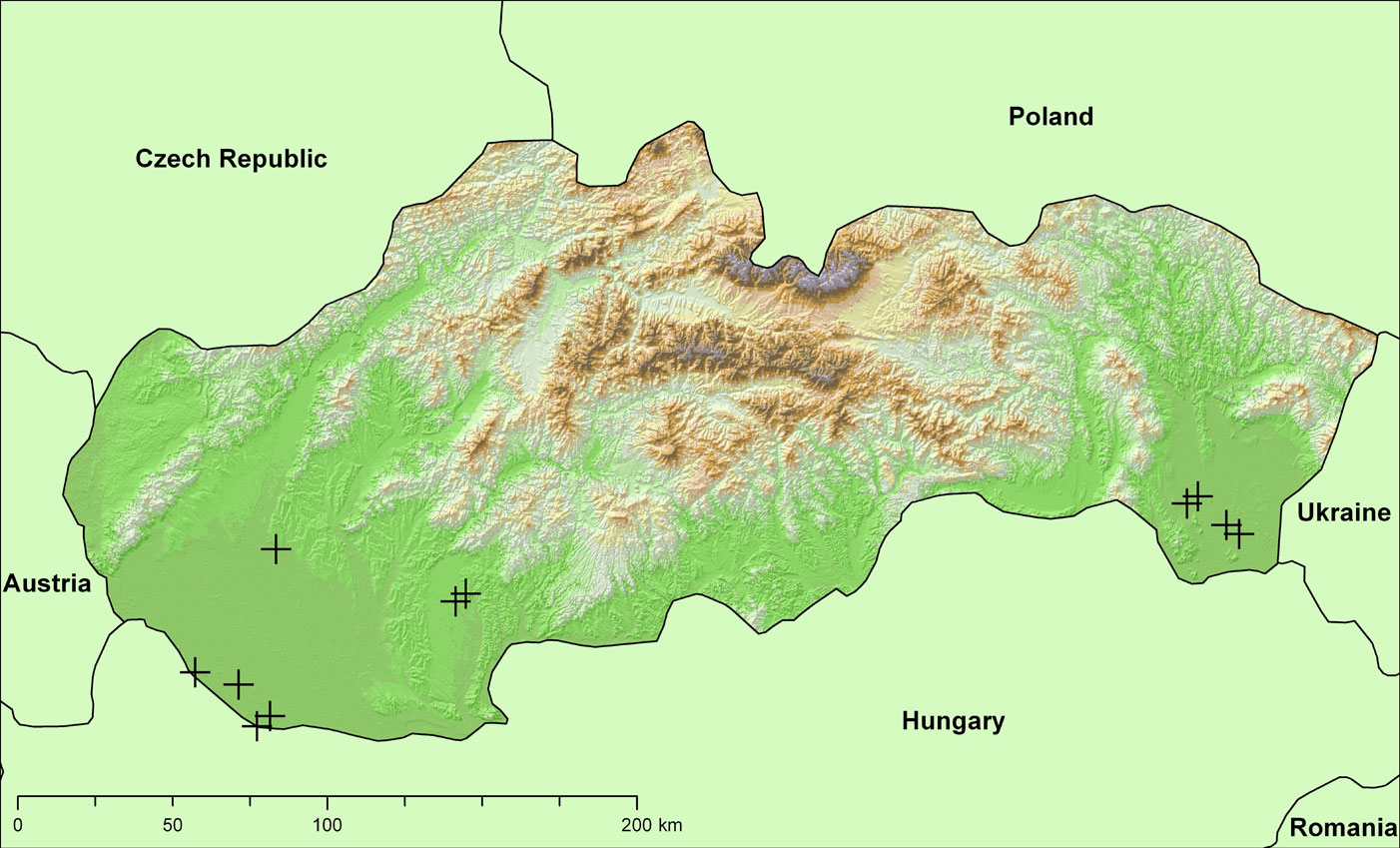
The site index q indicates the fertility of a location based on the mean tree height after 30 years since the establishment ([12]). For example, a site index of 40 indicates that a 30-years-old poplar stand attains a mean height of 40 meters. Additional characteristics of the sampled stands including age, diameter at breast height (DBH) and tree height are reported in Tab. 1.
Tab. 1 - Basic growth characteristics of the sampled poplar trees. (DBH): diameter at breast height.
| Clone | Number of trees | Age | DBH | Height | Site index |
|---|---|---|---|---|---|
| “Robusta” | 5 | 22-44 | 35-49 | 30-41 | 24-34 |
| “I-214” | 6 | 19-47 | 34-56 | 25-40 | 22-40 |
Sampling and analysis
Samples were taken from the following five tree segments: (1) the base, middle and top part of each stem provided three wood and bark samples; and (2) two small-wood samples were taken from the crown, one 4-6 cm in diameter and the other < 4 cm. A total of 88 samples were obtained, of which 33 wood, 33 bark and 22 small-wood samples with bark. Fresh volume of samples was determined to the nearest 1 ml in a calibrated cylinder. Dry weight was measured after oven-drying at 105 °C using a 0.01g precision scale. Samples were then ground with a Retsch cutting mill (Gmbh, Germany) and homogenized by thorough mixing. The calorific values (as Higher Heating Values) of dry matter (CVDM - [J g-1]) were determined by the C-200 calorimeter according to the Standard ISO 1928:2009 for solid mineral fuels. Four repetitions per sample were carried out.
Calculation procedures
The calorific production of the above-ground biomass for clones was derived by recalculation from its volume production. This was achieved using the yield tables’ model for the “Robusta” and “I-214” poplar clones ([11], [12]), their biomass densities ([13]) and the calorific value of the biomass dry matter.
Yield tables predict the biomass volume (VB - m3 ha-1) based on age t and site index q as follows (eqn. 1):
Biomass volume was thus obtained for whole-tree wood components, stem and large wood elements. All volumes (over-bark and under-bark) were given for mixed coppice, main stand, secondary crop, total production and also for total current annual and total mean periodic increments.
The calorific value capacity (CVC - GJ m-3) was calculated from measured density values (D - kg m3) and CVDM (J g-1) as follows (eqn. 2):
The calorific energy content of whole stand (CEC - GJ ha-1) was then derived by replacing in eqn. 2 the average value of calorific capacity (eqn. 3):
The final model then predicts the calorific energy production (CEP - GJ ha-1) of poplar clone stands based on age t and site index q (eqn. 4):
The energy use efficiency throughout the life-cycle (EUE - %) was derived according to Larcher ([9]) and Pretzsch ([15]), as the ratio of the energy accumulated in the above-ground biomass (AGTB - GJ ha-1) and the incoming solar energy as follows (eqn. 5):
where CEPAGTB is the calorific energy production in the above-ground trees biomass (GJ ha-1), and RMay-Sep is the incoming global radiation (GJ ha-1) over the vegetation period (May-September).
Data for annual global radiation were taken from the meteorological observatory at the Geophysical Institute of the Slovak Academy of Science in Mlynany, Slovakia (long. 18° 20’ E, 48° 19’ N), at an approximate elevation of 195 m a.s.l. Recorded average annual global radiation was 436 212 kJ cm-2 y-1 (43 621.2 GJ ha-1 y-1) for the period 1970-2002 ([10]). The incoming radiation for the period considered in this study (May to September) represents the 64.8% of the annual total radiation (Tab. 2).
Tab. 2 - Percentage distribution of annual global radiation in each months (1997-2002).
| Month | Jan | Feb | Mar | Apr | May | Jun | Jul | Aug | Sep | Oct | Nov | Dec |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| % | 2.5 | 4.3 | 7.6 | 10.6 | 13.9 | 14.8 | 14.7 | 12.8 | 8.6 | 5.7 | 2.6 | 1.9 |
Statistical analysis
Analysis of variance was applied to test for differences in calorific value capacity obtained from eqn. 2, using the following three factors: (1) the clone (“Robusta” and “I-214”); (2) the excising location on the tree (base, middle and top of the tree stem plus 4-6 cm and < 4 cm small wood); and (3) the biomass fraction (wood, bark and small-wood with bark). Tukey’s post-hoc HSD test was applied to test for differences in mean values (α = 0.05).
Results and discussion
Calorific values of biomass fractions
Density (kg m-3), calorific values of dry matter (J g-1) and calorific value capacity (GJ m-3) were obtained for each of the three biomass fractions considered for each clone (Tab. 3). Results of the analysis of variance carried out may be summarized as follows: (1) overall, the influence of the factors explored in the analysis was statistically significant (p<0.001 - Tab. 4); (2) the influence of the individual factors considered on biomass calorific value was statistically significant except for the clone (Tab. 5); (3) the post-hoc test applied confirmed that differences in the average calorific content do exist among the three biomass fractions analyzed (p<0.05 - Fig. 2); and (4) significant differences in the calorific value capacity were detected only between the three crown parts and the other two parts of the stem (the base and the middle). The above results indicate that stem wood from these poplar clones can be advantageously utilized for mechanical and chemical purposes.
Tab. 3 - Density, calorific dry matter and calorific capacity of the “Robusta” and “I-214” poplar clones analyzed. Mean values (± standard deviation) of calorific content are reported.
| Characteristics | Clone | Bark | Wood | Small-Wood |
|---|---|---|---|---|
| Density (kg m-3) | “Robusta” | 389.39 ± 29.99 | 427.47 ± 37.45 | 468.98 ± 39.36 |
| “I-214” | 385.25 ± 33.79 | 392.96 ± 58.49 | 456.72 ± 22.49 | |
| Calorific value of dry matter (J g-1) |
“Robusta” | 17975 ± 547 | 18350 ± 468 | 18589 ± 553 |
| “I-214” | 17801 ± 473 | 18492 ± 359 | 18307 ± 348 | |
| Calorific value capacity (GJ m-3) |
“Robusta” | 7.01 ± 0.65 | 7.85 ± 0.76 | 8.73 ± 0.93 |
| “I-214” | 6.86 ± 0.61 | 7.27 ± 1.10 | 8.36 ± 0.51 |
Tab. 4 - Results of the ANOVA on the poplar clones’ calorific value using the clone, the excising location on the tree and the biomass fraction as factors.
| Source of variability | Sum of Squares | F-statistic | p-value |
|---|---|---|---|
| Total | 89.66 | - | - |
| Explained | 51.74 | 2.36 | 3.72E-05 |
| Residual | 37.92 | - | - |
Tab. 5 - Results of the one-way ANOVA carried out to test the influence of individual factors on poplar clones’ calorific value.
| Factors | Sum of Squares | F-statistic | p-value |
|---|---|---|---|
| Biomass fraction | 34.12 | 26.11 | 1.32E-09 |
| Excising Location on the tree | 42.74 | 18.90 | 3.25E-11 |
| Clone | 2.90 | 2.87 | 0.094 |
Fig. 2 - Mean (± SE) calorific capacity of basic biomass fractions in “Robusta” and “I-214” poplar clones. Different letters indicate significant differences among the means (P < 0.05) according to the post-hoc Tukey HSD test.
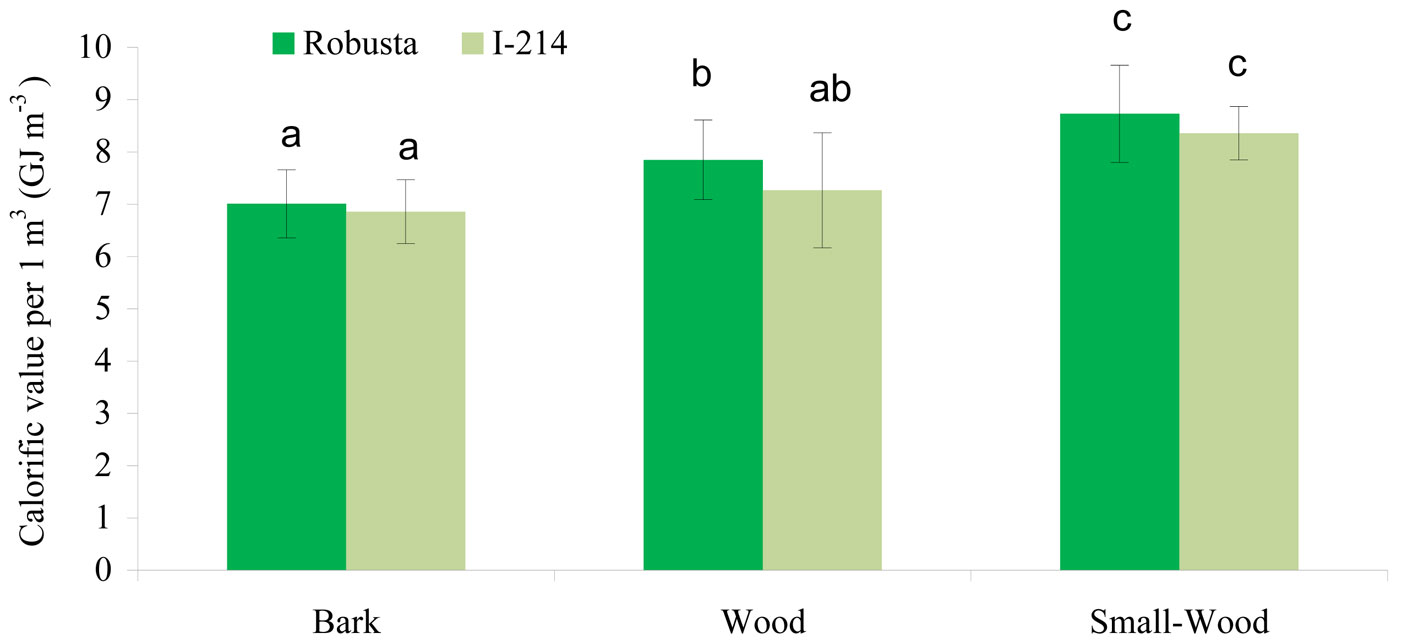
Energy content in plant tissues is proportional to their carbon content. Larcher ([9]) asserted that the greatest energy content in plant tissues is due to lignin (26.4 kJ g-1), lipids (38.9 kJ g-1), and terpenes (46.9 kJ g-1). Accordingly, in this study the highest calorific content was found for small-wood, likely depending on the higher abundance of the above-mentioned tissues in this biomass fraction.
Modeling of calorific energy content
Since the yield tables used in this study predict only the volume for wood, bark and small-wood with bark, a simplification in the calculation of the calorific energy production was applied according to eqn. 4.
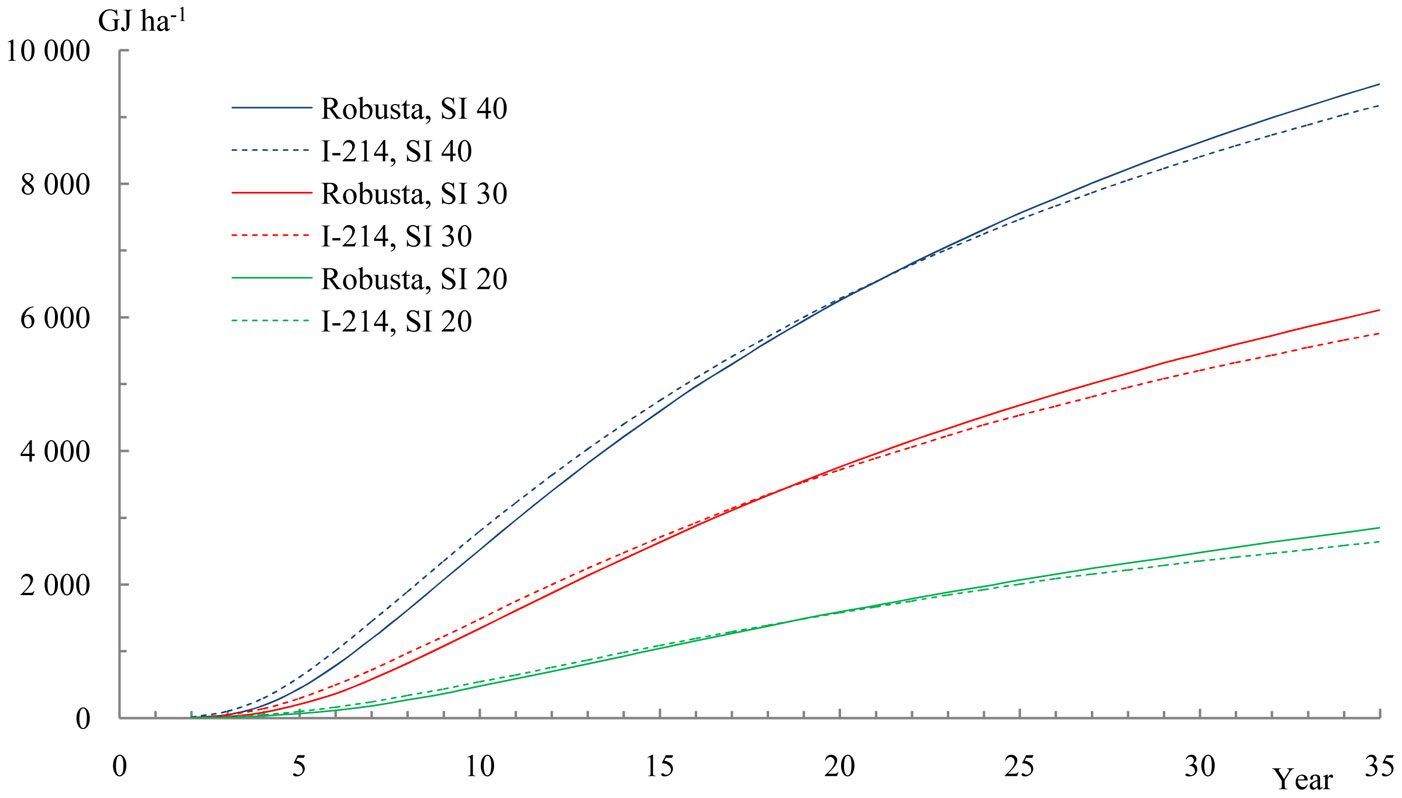
The life-time calorific energy production obtained from the model applied as a function of stand age and site index is displayed in Fig. 3. The growth curves starts with values accrued from the second year of the growth tables, given that values before that age are close to zero. However, after 35 years and at site indexes of 20, 30 and 40, the estimated calorific energy productions are approximately 2 700, 6 000 and 9 300 GJ ha-1, respectively. Clone “I-214” had a slightly higher calorific production than “Robusta” during the first half of its rotation time, the latter clone recording 7.8-8.0 GJ m-3 in the middle-late years and site indexes 40-20, while “I-214” clone had just 0.5 GJ m-3 less.
Fig. 3 - The calorific energy production in poplar clone biomass, dependent on stand age and site index (SI) 20, 30 and 40.
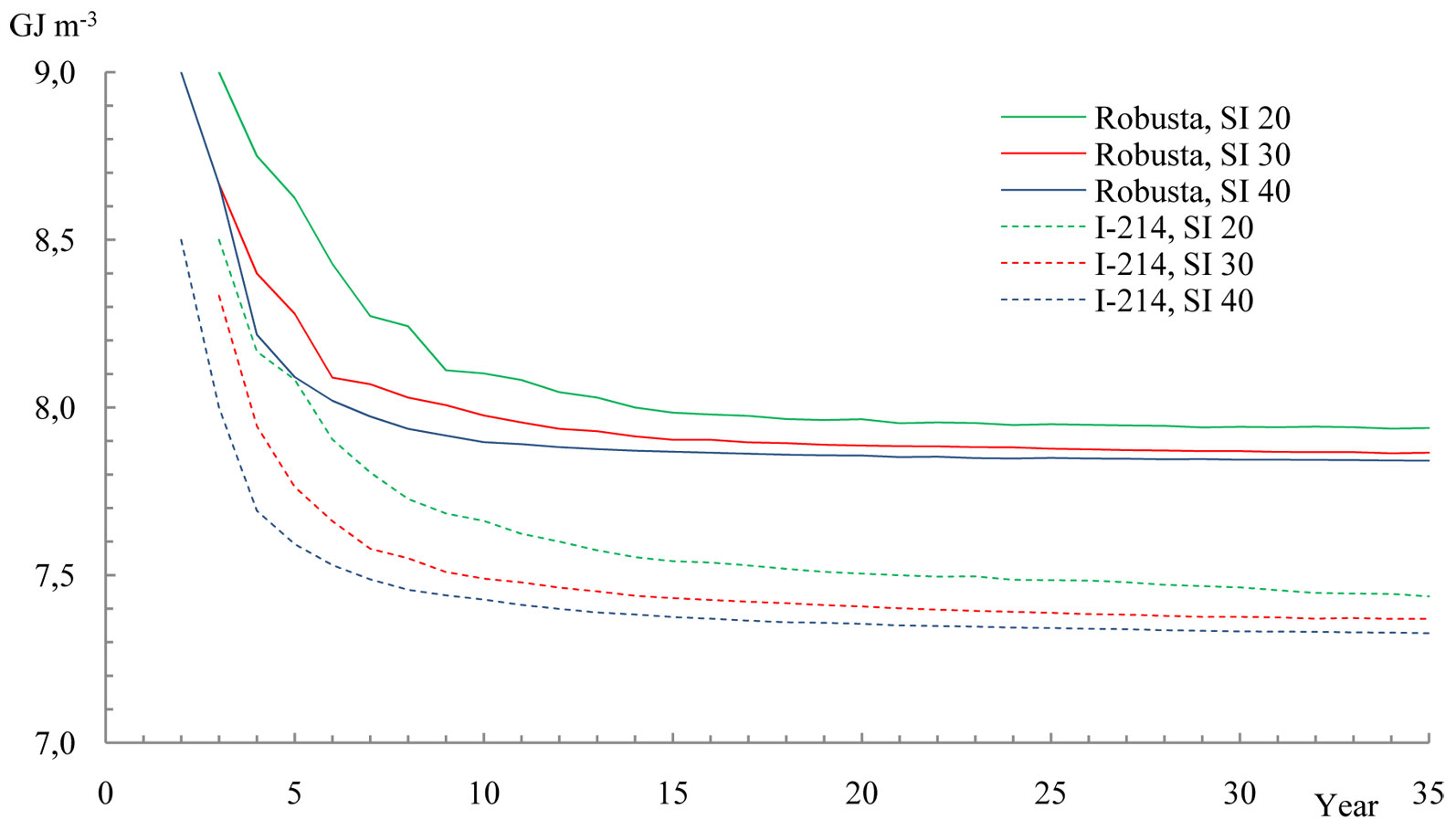
However, larger differences between the clones analyzed were observed when the calorific value per m3 of biomass at lower site indexes was considered (Fig. 4). Indeed, a significant reduction in the calorific value from the 2nd to the 5th year of growth was observed.
Fig. 4 - The average calorific capacity in 1 m3 of poplar clone biomass, dependent on stand age and site index (SI) 20, 30 and 40.
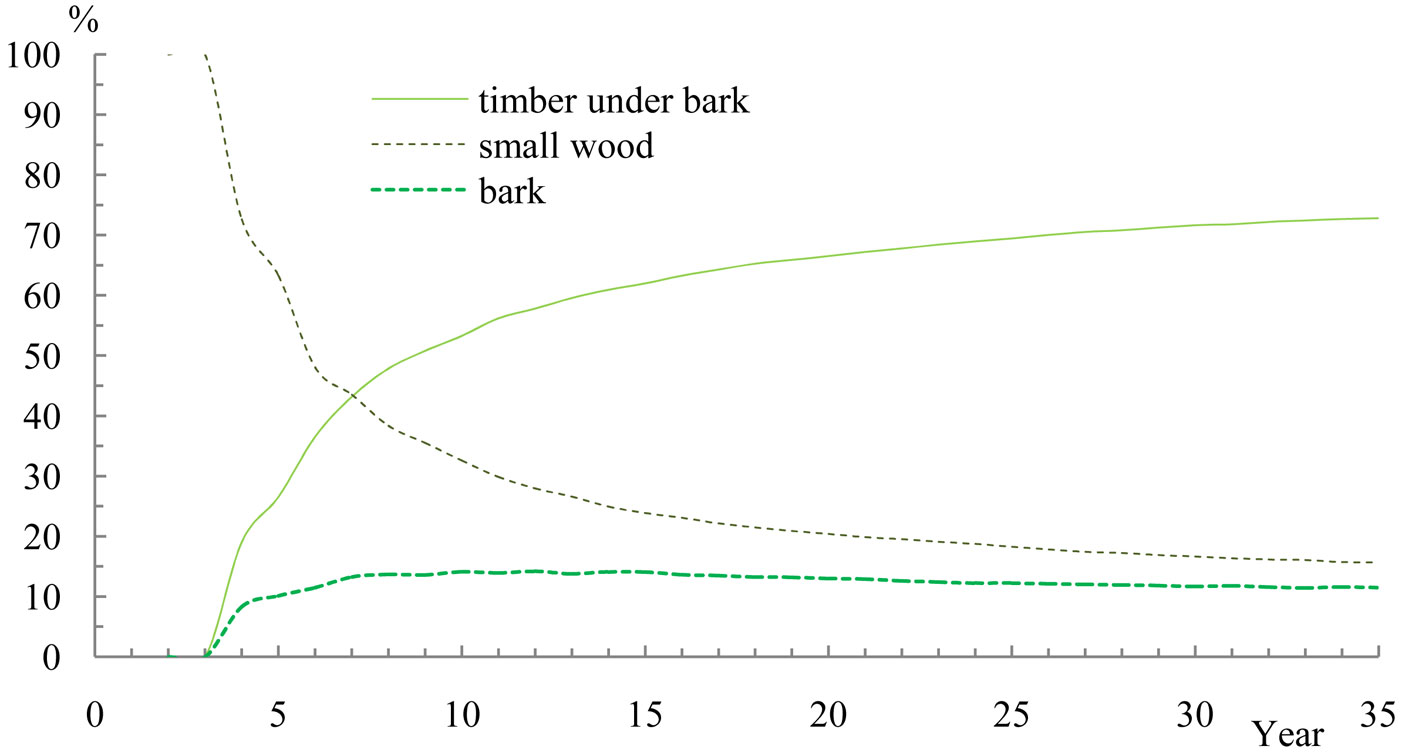
As mentioned above, calorific capacity accumulated differently in the different biomass fractions as a function of a varying proportion of lignin, terpenes and essential oils contained in the tissues (Fig. 5). After 35 years since plantation, 73% of the calorific value was stored in the wood, 16% in the small-wood component, and 11% in the bark. It is should be noted here that the decrease in bark calorific capacity was quite low, reducing only from 15% to 11% over a twenty year period.
Fig. 5 - The proportion of calorific energy production in wood, bark and small-wood for the “Robusta” clone at site index 30.
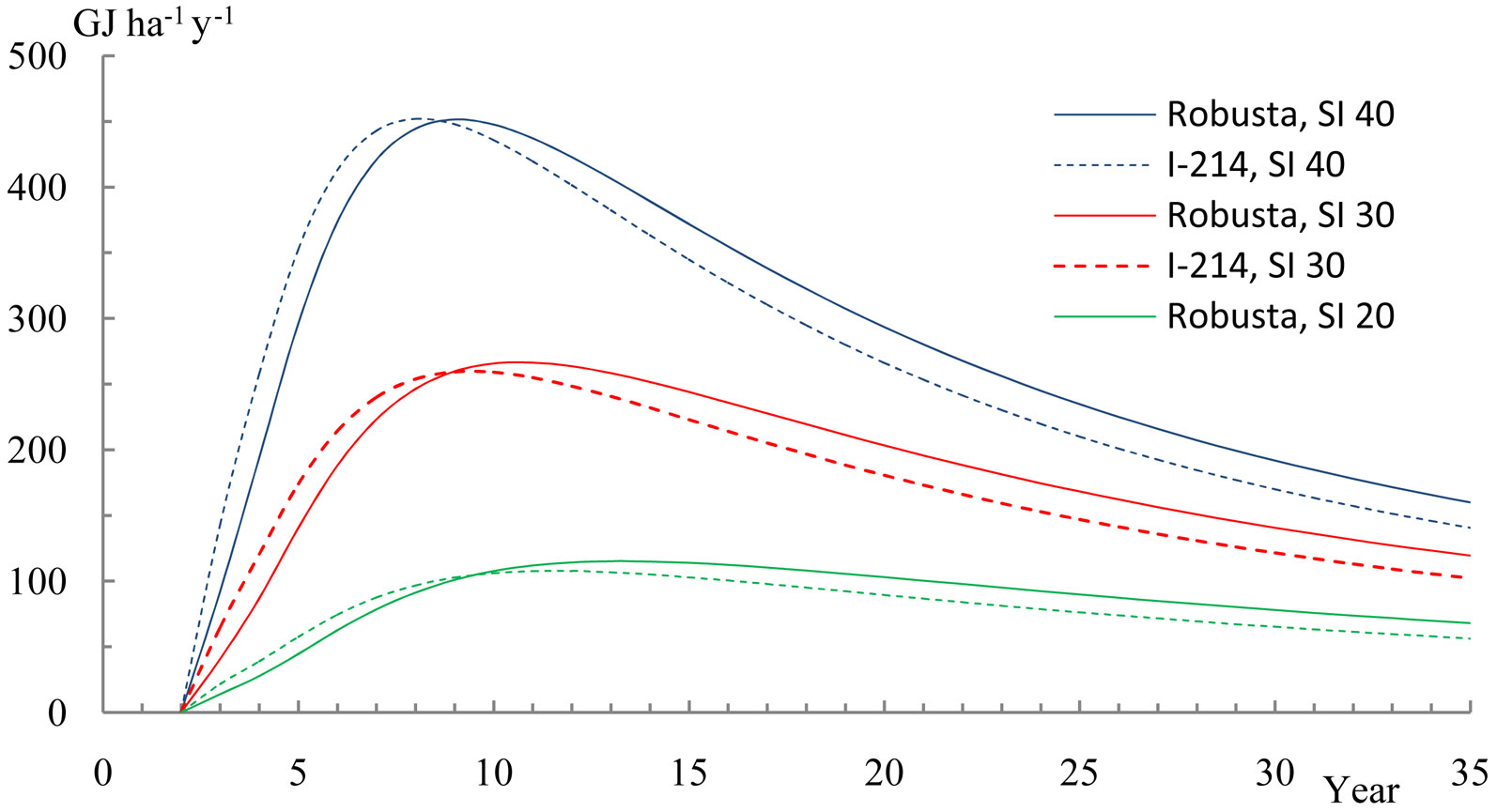
Annual increment estimates in calorific energy content culminated approximately at age 8-13 in the 20-40 site indexes, with a value of 110-450 GJ ha-1 y-1 (Fig. 6), the “I-214” clone culminating 1-2 years earlier than “Robusta”. The calorific value then markedly decreased with increasing age to approximately 60-160 GJ ha-1 y-1 after 35 years. As a comparison to hard broadleaf species, Bublinec et al. ([1]) found that the average annual increment of energy content in aboveground biomass in a 75 years-old beech stand was approximately 176 GJ ha-1 y-1.
Fig. 6 - Annual increments in poplar biomass calorific energy, dependent on stand age and site index (SI) 20, 30 and 40.
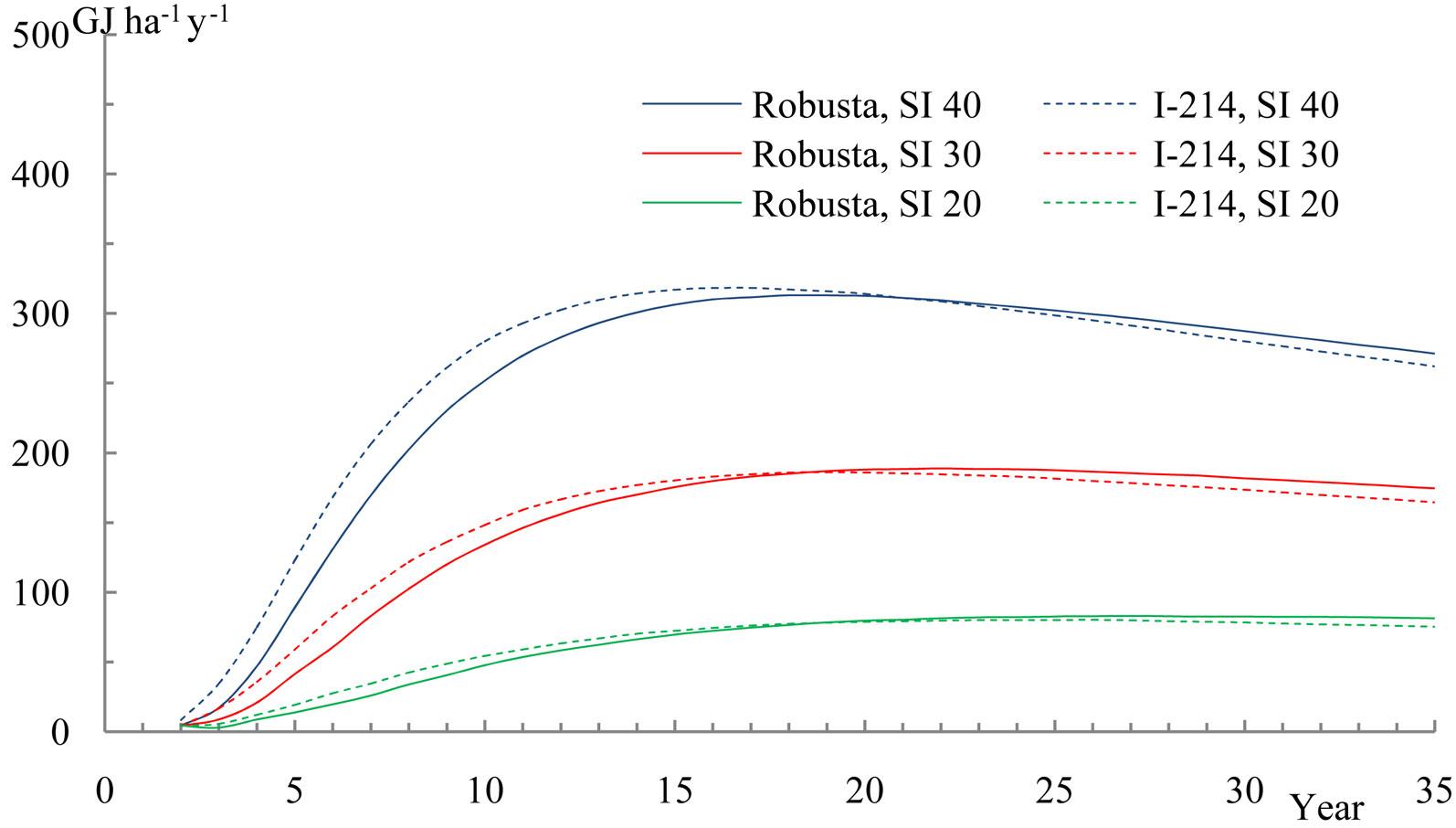
The estimated average annual production of calorific energy content (Fig. 7) for site indexes 20-40 culminated approximately 17-26 years after plantation, with values of 80-320 GJ ha-1 y-1, the “I-214” clone culminating again 2-3 years earlier than “Robusta”. This value can be compared with the highest annual increment of 450 GJ ha-1 y-1 recorded for 8-9 years-old poplar stands on above-average site indexes in the temperate climatic zone in Central Europe. According to the above evidence, it is recommended to harvest the poplar clone stands at this age, since the average annual energy production under these conditions is lower both before and after this time.
Fig. 7 - Average annual biomass calorific energy production, dependent on stand age and site index (SI) 20, 30 and 40.
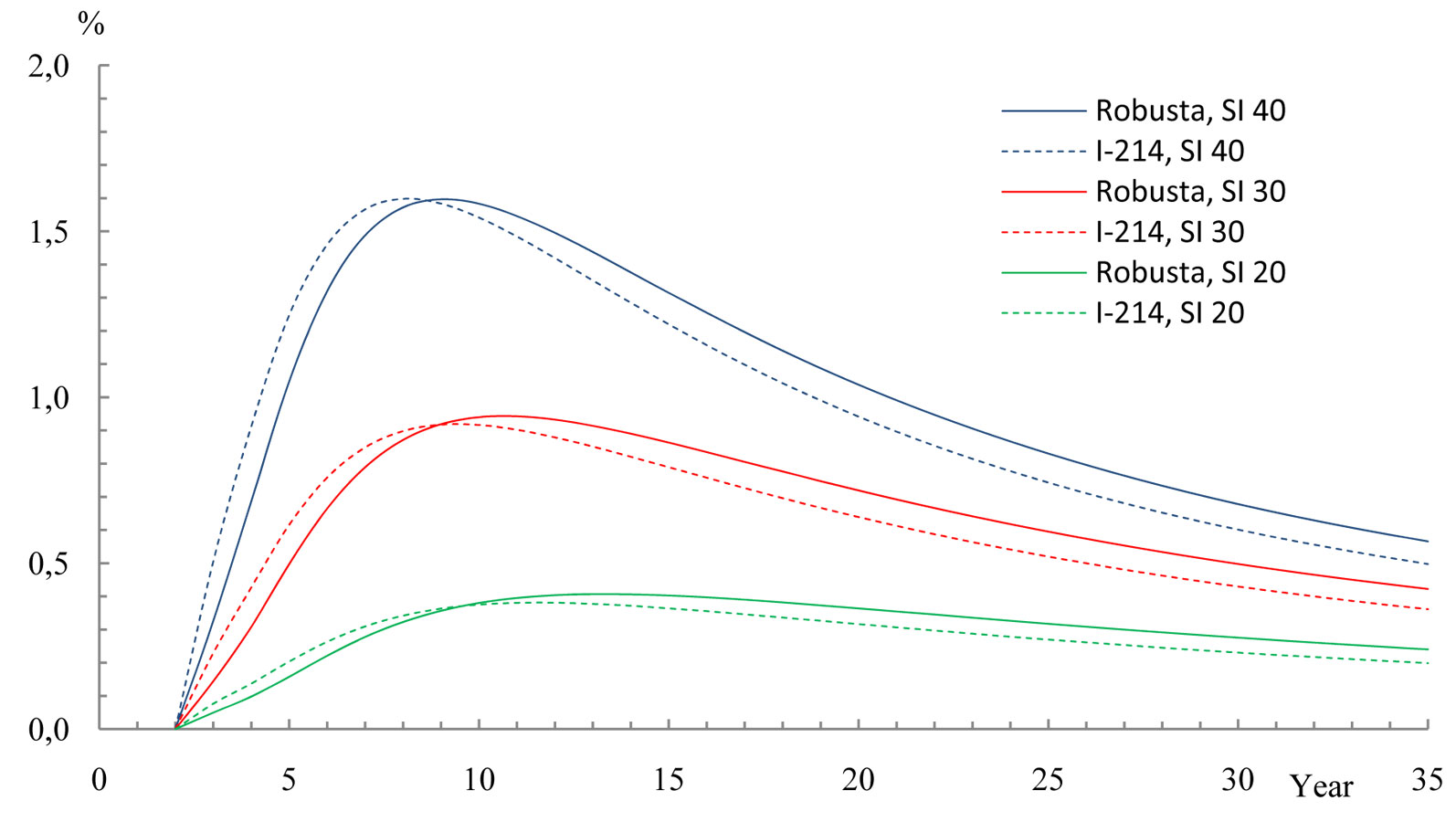
Energy use efficiency
The energy use efficiency calculated by eqn. 5 for “Robusta” and “I-214” poplar clones has a similar relationship to their annual calorific production increments (Fig. 8). Poplar clones stands achieve their maximum solar radiation usage (approximately 1.6%) at 8-9 years after plantation at the highest site indexes. This value, however, markedly decreases with lower site index and higher age, attaining only 0.2-0.6% by age 35. As for comparison, Bublinec et al. ([1]) report that beech ecosystems in Central Europe effectively utilize less than 1% during the photosynthetic activity period (May to September). Moreover, Pretzsch ([15]) reported energy use efficiency values of 0.8-0.9% for spruce and pine in Germany and 0.5-0.6% for broadleaf species.
Fig. 8 - Energy use efficiency of poplar clone biomass calorific production, dependent on stand age and site index (SI) 20, 30 and 40.
Based on our estimates, poplar clones received 28 267 GJ ha-1 solar radiation during the period May to September, while Pretzsch ([15]) calculated 36 000 GJ ha-1 over the entire year. The above difference may be due to the fact that the latter estimate is based only on the marketable wood quantities from the national forest inventory. However, it is widely accepted that total above-ground biomass volume is much larger, exceeding 8-10% for coniferous species and 12-15% for broadleaves. Although Larcher ([9]) reported similar energy use efficiency values, with a 2% limit for forest stands, the solar radiation usage reported for most woody species was less than 1%.
Conclusions
The poplar clones “Robusta” and “I-214” poplar clones have high above-ground biomass production, eminently suitable for bio-fuel energy. Crown small-wood provided 0.8 GJ m-3 higher calorific value capacity than stem wood, and wood from the base and the middle of the poplar stems had 1 GJ m-3 less than the crown parts. Moreover, the bark registered only approximately a 0.61 GJ m-3 lower calorific capacity than the wood. The above findings are especially valuable in the light of industrial exploitation of the poplar clones’ biomass, since industrial energy utilization efficiency must include costs for both production and transport to the consumption point.
A calorific energy content of approximately 5 000 GJ ha-1 is available from the burned biomass of poplar clones at above-average site indexes, with a 25% decrease below-average sites. Therefore, the energy content of a 30 years-old poplar stand is similar to the 5 137 GJ ha-1 reported by Bublinec et al. ([1]) for a 75 years-old beech stand.
In addition to their fast growth, large biomass and high calorific production, poplar clones “Robusta” and “I-214” also exhibited a particularly high energy use efficiency compared to other wood species. Moreover, most poplar stands in this study accumulated more than 1% of the solar radiation.
Acknowledgment
This research was supported by grants from the Slovak Research and Development Agency, Project No. APVV-0131-07, and from the Slovak Grant Agency VEGA No. 2/0006/11.
References
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Slovak Academy of Sciences-Institute of Forest Ecology, Štúrova 2, SK-96053 Zvolen (Slovakia)
Slovak University of Agriculture in Nitra, A.Hlinku 2, SK-94976 Nitra (Slovakia)
Julian Mecko
National Forest Centre,Forest Research Institute, T.G.Masaryka 22, Zvolen (Slovakia)
Slovak Academy of Sciences, Institute of Landscape Ecology, Štefánikova 3, SK-81499 Bratislava (Slovakia)
Corresponding author
Paper Info
Citation
Jamnická G, Petrášová V, Petráš R, Mecko J, Oszlányi J (2014). Energy production of poplar clones and their energy use efficiency. iForest 7: 150-155. - doi: 10.3832/ifor0978-007
Academic Editor
Francesco Ripullone
Paper history
Received: Feb 21, 2013
Accepted: Nov 21, 2013
First online: Jan 23, 2014
Publication Date: Jun 02, 2014
Publication Time: 2.10 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2014
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 56067
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 46814
Abstract Page Views: 3113
PDF Downloads: 4591
Citation/Reference Downloads: 20
XML Downloads: 1529
Web Metrics
Days since publication: 4553
Overall contacts: 56067
Avg. contacts per week: 86.20
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2014): 10
Average cites per year: 0.83
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Growth performance and nitrogen use efficiency of two Populus hybrid clones (P. nigra × P. maximowiczii and P. trichocarpa × P. maximowiczii) in relation to soil depth in a young plantation
vol. 9, pp. 847-854 (online: 22 September 2016)
Research Articles
Tree aging does not affect the ranking for water use efficiency recorded from δ13C in three Populus deltoides × P. nigra genotypes
vol. 12, pp. 272-278 (online: 21 May 2019)
Research Articles
Climate-wise models of biomass productivity for hybrid poplar clones in Europe
vol. 16, pp. 188-194 (online: 30 June 2023)
Research Articles
Revealing the physiological basis of forester’s choice of poplar clones (Populus spp.)
vol. 17, pp. 156-164 (online: 14 June 2024)
Research Articles
Assessment of cadmium tolerance and phytoextraction ability in young Populus deltoides L. and Populus × euramericana plants through morpho-anatomical and physiological responses to growth in cadmium enriched soil
vol. 10, pp. 635-644 (online: 01 June 2017)
Research Articles
Effective woody biomass estimation in poplar short-rotation coppices - Populus nigra × P. maximowiczii
vol. 16, pp. 202-209 (online: 25 July 2023)
Research Articles
Nursery screening of poplar and willow clones for biofuel application in Ukraine
vol. 15, pp. 401-410 (online: 06 October 2022)
Research Articles
First vs. second rotation of a poplar short rotation coppice: leaf area development, light interception and radiation use efficiency
vol. 8, pp. 565-573 (online: 27 April 2015)
Research Articles
Verification of new Populus nigra L. clone improvement based on their performance over three rotations
vol. 13, pp. 185-193 (online: 12 May 2020)
Research Articles
Nutritional, carbon and energy evaluation of Eucalyptus nitens short rotation bioenergy plantations in northwestern Spain
vol. 9, pp. 303-310 (online: 05 October 2015)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







