Adjustment of photosynthetic carbon assimilation to higher growth irradiance in three-year-old seedlings of two Tunisian provenances of Cork Oak (Quercus suber L.)
iForest - Biogeosciences and Forestry, Volume 10, Issue 3, Pages 618-624 (2017)
doi: https://doi.org/10.3832/ifor2105-010
Published: May 17, 2017 - Copyright © 2017 SISEF
Research Articles
Abstract
Three-year-old seedlings of two Tunisian provenances of cork oak (Quercus suber L.) differing in climatic conditions at their geographical origin were subjected to increasing light intensities. Gaâfour was the provenance from the driest site and Feija from the wettest site. Low-light adapted seedlings from both provenances were exposed to two light treatments: full sunlight (HL) and low light (LL, 15% sunlight) for 40 days. The CO2-response curve of leaf net photosynthesis (An-Ci curve) established under saturated photon flux density was used to compare photosynthetic parameters between leaves subjected to continuous low light (LL leaves) and leaves transferred from low to high light (HL leaves). Transfer from low to high light significantly increased net photosynthesis (An) and dark respiration (Rd) in Gaâfour provenance but not in Feija. After transfer to high irradiance, specific leaf area (SLA) did not change in either provenance. This suggested that the increase in photosynthetic capacity on a leaf area basis in HL leaves of Gaâfour provenance was not due to increased leaf thickness. Only the seedlings from the Gaâfour provenance were able to acclimate to high light by increasing Vcmax and Jmax.
Keywords
Quercus suber, Photosynthesis, Vcmax, Jmax, Stomatal Limitation
Introduction
Quercus suber L. is an evergreen oak growing across a wide range of environmental conditions in the Mediterranean Basin. Due to these diverse conditions, Mediterranean forest tree species are exposed to a range of selective pressures that may lead to phenotypic plasticity and local adaptation ([30]). Light is one of the most heterogeneous factors affecting plant growth and development ([7]) and acclimation of leaf photosynthesis to different light environments is a key factor that maximizes carbon gain ([8], [18]). Acclimation to fluctuations in light conditions plays a crucial role in determining the competitive ability of forest tree species ([14]). Acclimation to environmental stresses is achieved through structural, morphological and physiological adjustments at both leaf and whole-plant level ([27], [33], [14]). The acclimation of plants to high light condition appeared in the ability to use high photosynthetic photon flux densities efficiently ([14], [35]).
Plants can form sun and shade leaves in response to the variation in growth irradiance. Sun leaves have a higher photosynthetic capacity than shade leaves. Higher photosynthetic capacities in sun leaves correlate with a greater leaf thickness and is supported by a greater investment of nitrogen in photosynthetic enzymes ([4], [9], [28], [37], [8], [23], [24], [25]). Furthermore, strong correlations were reported between photosynthetic capacity and surface area of mesophyll cells ([25]), between photosynthetic capacity and the chloroplast area facing the intercellular space ([23]) and between photosynthetic capacity and stomatal and mesophyll conductances to CO2 diffusion ([16]). Pandey & Kushwaha ([25]) studied the relationship of leaf anatomy with photosynthetic acclimation in Valeriana jatamansi under full irradiance [FI, 1600 μmol (PPFD) m-2 s-1] and shade conditions [SC, 650 μmol (PPFD) m-2 s-1]. FI plants had thicker leaves with lower specific leaf area (SLA), enabling to arrange all their chloroplasts along the mesophyll cell surface. Leaf thickness is determined by the irradiance during leaf development, and it changes little after leaf maturation ([32]). Nevertheless, when leaves are subjected to higher irradiance after maturation, their photosynthetic capacity often increases. However, high photosynthetic capacity is not always associated with thicker leaves.
The aim of this study was to test whether the habitat of origin induced an intraspecific variation in response to transfer of plants from low to high growth irradiance. To this purpose, seedlings of cork oak (Quercus suber L.) originating from two different natural habitats and representing a marked climate gradient were used. Changes in photosynthetic characteristics of mature leaves were examined when transferred from low to full sunlight condition. The photosynthetic capacity, the maximum carboxylation rate of Rubisco (Vcmax), the potential light-saturated electron transport rate (Jmax) and the specific leaf area were determined for leaves grown under different light conditions.
Materials and methods
Seed origin and experimental design
Acorns of two Quercus suber populations originating from contrasting environments in the northwestern provinces of Tunisia were collected in October 2010. The first site, the National Park of Feija (36° 30′ 00″ N, 08° 20′ 00″ E), is located in the North of the Kroumirie Mountains and is characterized by a cold and humid climate. The altitude varies between 800 and 1500 m a.s.l. and the average annual rainfall is 1217 mm, with precipitation increasing with altitude and reaching 1800 mm year-1. In January, the average temperature is 7 °C and can drop to 0 °C with snow which falls every year. In summer, the temperature rises notably and averages around 29 °C. This leads to a mean annual temperature of 14.3 °C. Cork oak trees are found in forest mosaics along with other tree species, including Zeen oaks (Quercus faginea), maritime pines (Pinus pinaster) and many shrub species.
The second site is located in Gaâfour (36° 32′ 190″ N, 09° 32′ 40″ E) in the southern hills and plains around the city of Siliana. It is characterized by a semi-arid climate (480 mm year-1) with moderate winters and hot dry summers. Cork oak trees at this site are found in agroforestry systems containing olive trees. The altitude is 560 m a.s.l. and the mean annual temperature is 17 °C.
Immediately after collection, acorns were planted in a common greenhouse at the National Research Institute for Rural Engineering, Waters, and Forestry under low light conditions (LL, 15% of full sunlight). On October 1st, 2013, low-light adapted seedlings were growing in 5 litres pots containing a mixture of equal proportions of soil and compost. Seedlings were randomly assigned to one of two light treatments for 40 days: (i) high light (100% natural incident irradiance, HL); and (ii) low light (LL). In the LL treatments, light levels were obtained through the use of layers of neutral shade-cloth, while in the HL treatment seedlings were left uncovered. On a sunny day, photosynthetically active radiation (PAR), measured using a Li-190® device (Li-Cor Bioscience, Lincoln, NE, USA) ranged between 1630-1810 μmol m-2 s-1 and 215-315 μmol m-2 s-1 in the HL and LL treatments, respectively. Irrigation to saturation was provided manually each day. Mean volumetric soil water contents of the pots containing seedlings in the two light treatments was monitored by a time domain refractometry (TDR, Trase system I, Soil moisture Equipment Corp., USA) and was approximately 25-30%. To ensure that all seedlings were exposed to a similar range of light throughout the experiment and minimize the possible effects of within-block light variability, each block containing twenty pots (ten for each provenance per light treatment) was periodically moved. All experiments were carried out using mature leaves that were fully expanded and developed prior to the light treatment.
CO2 response curves
Photosynthetic traits were measured in situ on mature leaves of four to six different seedlings per treatment (one leaf per plant) during the late morning (08:00-11:00 h) and early afternoon (13:00 -16:00 h). At the beginning of the experiment, leaf gas exchange was measured in three low light-adapted seedlings from each provenance. After 40 days, four plants from Gaâfour and Feija provenances were transferred from low to high light and used to carry out gas exchange measurements. Three additional LL adapted Gaâfour and Feija seedlings were used to ensure that there was no change of photosynthesis parameters during the experiment for plants kept under LL. Photosynthetic carbon dioxide response curves were recorded using a portable gas-exchange system (LI-6400®, Li-Cor Inc., Lincoln, NE, USA) equipped with a 2 × 3 cm light-source chamber (6400-02B LED®, Li-Cor). Each leaf was adapted to dark for approximately 30 minutes in the measurement chamber. When the gas exchange reached a steady state, net CO2 efflux was recorded as an estimate of dark respiration (Rd). At this point, incident PFD was set to 1600 µmol m-2 s-1 (saturating light). The leaf temperature was maintained at 25 °C and the humidity of the incoming air was kept at 50-60%. The external CO2 partial pressure (Ca) variation consisted of 12 steps, starting by inducing photosynthesis at ambient CO2 concentration of 400 µmol mol-1 until net photosynthesis (An) stabilized (An varied by less than ± 2%). This was important to ensure a steady-state activation of Rubisco ([19]). The external CO2 concentration (Ca) values were then decreased to 300, 200, 100 and 50 µmol CO2 mol-1 respectively. Upon completion of the measurement at 50 µmol CO2 mol-1, Ca was increased back to 400 ppm to check whether the original An could be restored. If this was achieved, Ca was increased stepwise to 600, 700, 1000, 1200, 1500 and 2000 ppm. Leaves were allowed to equilibrate for at least 5 minutes at each step before data logging. At the end of each A-Ci curve, the leaf area and mass enclosed in the chamber was measured with a leaf area meter portable laser (Model Cl-202). The specific leaf area (SLA) was determined as the ratio of the leaf area to leaf dry mass of individual leaves. To estimate the CO2-saturated rate of photosynthesis (Asat), CO2 response curves were fitted using a three components exponential function ([39]) according to the following equation (eqn. 1):
where An is leaf net photosynthetic rate and x is Ci. Using this equation, Asat was calculated as (a+c). The CO2 compensation point (Γ) was estimated from x-axis intercepts.
Estimation of Vcmax, Jmax and relative stomatal limitation
Non-linear regression techniques, based on the equations of Farquhar et al. ([11]) and later modified by Sharkey ([31]) and Harley & Sharkey ([17]), were used to estimate Vcmax (the maximum rate of carboxylation limited by Rubisco) and Jmax (the maximum rate of carboxylation limited by electron transport). In some cases, carboxylation may also be limited by triose phosphate availability ([31], [17]); however, this was not observed in any of the seedlings used in this study. Vcmax and Jmax describe the upper limit to biochemical processes (amount, activity, and kinetics of Rubisco and regeneration of RuBP), both parameters are derived from different portions of the An-Ci curve. Points in the lower portion (at low Ci) of the curve are used for Vcmax and points in the upper portion (at high Ci) for Jmax. The Michaelis-Menten constants for CO2 and O2 (Kc and Ko respectively) and the CO2 compensation point in the absence of mitochondrial respiration (Γ*), as well as their temperatures dependencies, were taken from Bernacchi et al. ([3]).
The stomatal limitation was calculated according to Farquhar & Sharkey ([10]) as follows (eqn. 2):
where Ao represents carbon assimilation under natural ambient CO2 in the absence of stomatal limitation.
Statistical analysis
Gas exchange measurements were repeated 4 and 6 times for HL and LL seedlings respectively. Mean values and standard errors were calculated. One-way analysis of variance (ANOVA) followed by the post-hoc Duncan’s test (α =0.05) were applied to test for differences among means of photosynthetic gas exchange parameters and specific leaf area.
Two-way analysis of variance was also conducted to test the interaction effect (provenance × light) on photosynthetic parameters. Pearson’s correlation between the photosynthetic parameters and SLA were also calculated. All ANOVA analyses were performed using the software package SigmaPlot® (Systat Software Inc., San Jose, CA, USA). Pearson’s correlation analysis was done using XLSTAT v. 2014 (⇒ http://www.xlstat.com) after data standardization.
Results
Photosynthetic responses to transfer from low to high light
Gas exchange parameters between LL and HL leaves in Gaâfour and Feija seedlings at an ambient CO2 concentration of 400 ppm were compared (Fig. 1). Stomatal conductance (gs) and net carbon assimilation (An) measured under saturating light in LL leaves were found to be similar in both Feija and Gaâfour provenances.
Fig. 1 - Leaf physiological traits of leaves maintained continuously in low light (black) or transferred from low to high light (white): net photosynthesis (An, µmol m-2 s-1); dark respiration (Rd, µmol m-2 s-1); stomatal conductance (gs, mmol m-2 s-1); intercellular CO2 concentration (Ci, µmol mol-1); relative stomatal limitation (Ls, %); and specific leaf area (SLA, cm2 g-1). Different uppercase letters indicate a significant difference between provenances in the same light environment, whereas different lowercase letters indicate significant difference between light environments in the same provenance (P ≤ 0.05).
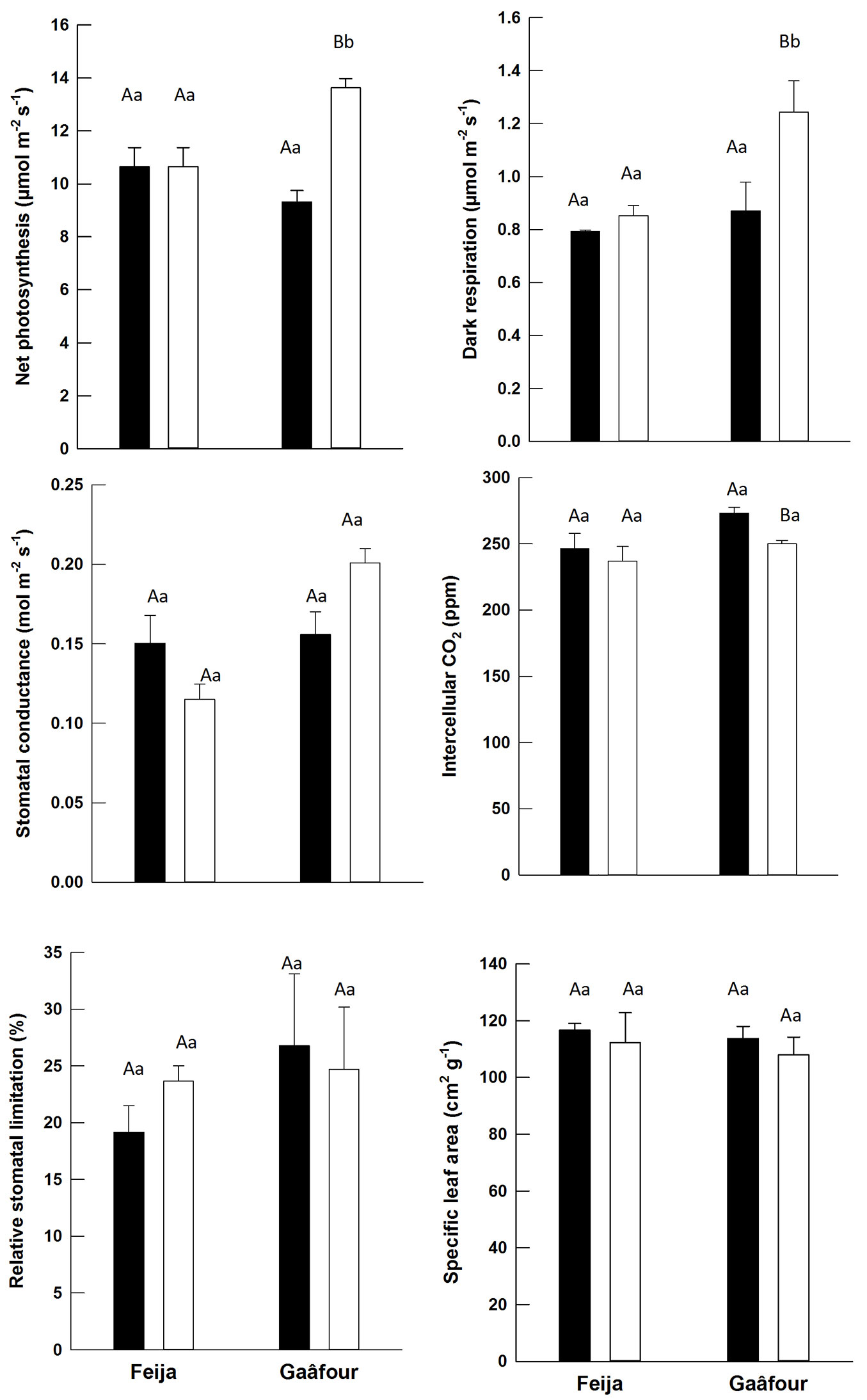
In HL leaves from the Gaâfour provenance, a significant increase in An was observed when compared to LL leaves (P < 0.01). While the gs in this environment also increased, it was not significant (P > 0.05). No notable variations in these parameters (An and gs) were detected in Feija leaves under different light condition treatments. At HL, An and gs were significantly higher in Gaâfour leaves than in Feija leaves (P = 0.027 and 0.003 for An and gs, respectively).
It was expected that the variations of gs may explain significant differences of An in HL leaves of the two provenances. However, gs did not show such differences, since stomatal limitation to carbon assimilation (Ls) remained similar in both provenances at HL conditions. Similarly, the intercellular CO2 concentration (Ci) did not change in the HL leaves of Gaâfour and Feija seedlings when compared with LL leaves. A significant contribution of gs to the lower An in Feija leaves at HL is therefore not supported. Although gs did not differ between light treatments in the same provenance, there was a highly significant treatment × provenance interaction effect on both gs and An (P < 0.01 - Tab. 1).
Tab. 1 - Summary of two-way analyses of variance (F-values) for differences in gas exchange parameters between treatments and provenances. (ns): non-significant; (**): P< 0.01; (*): P <0.05.
| Factors | A n | Rd | g s | C i | L s | Γ | V cmax | J max | A sat |
|---|---|---|---|---|---|---|---|---|---|
| Provenance | 2.36 ns | 8.16* | 12.44** | 4.92* | 0.8 ns | 2.8 ns | 32.52** | 7.35* | 11.68* |
| Sunlight | 11.98** | 6.9* | 0.13 ns | 3.33 ns | 0.06 ns | 2.27 ns | 85.94** | 1.77 | 0.15 ns |
| Provenance × sunlight | 14.09** | 3.61ns | 9.63** | 0.57 ns | 0.46 ns | 1.37 ns | 95.07** | 2.35 ns | 0.45 ns |
There were no significant changes to the leaf dark respiration in the seedlings of Feija transferred from LL to HL in comparison to those maintained at LL (P > 0.05). Dark respiration was highest in HL leaves of Gaâfour provenance (Tab. 1) but did not show a significant treatment × provenance interaction effect (Tab. 1). No significant changes in specific leaf area were measured between leaves of seedlings maintained at LL and those transferred to a HL condition in either provenance (Fig. 1).
Photosynthesis in response to Ci
In order to characterize the photosynthetic activity in the leaves of Gaâfour and Feija provenances under the two light conditions, An was measured as a function of the intercellular CO2 concentration (Ci). As shown in Fig. 2 (insert), An of LL leaves in Gaâfour provenance was similar to Feija provenance at low Ci but it was higher at high Ci. In the HL leaves of Gaâfour, An as a function of Ci increased in comparison to LL leaves. No differences were observed between Feija seedlings grown in either light condition.
Fig. 2 - Net photosynthesis (An) as a function of intercellular CO2 concentration (Ci) of leaves maintained continuously in low light (solid line) or transferred from low to high light (dotted line) in Feija and Gaâfour provenances. Inserts: Feija (closed symbols) and Gaâfour (open symbols).
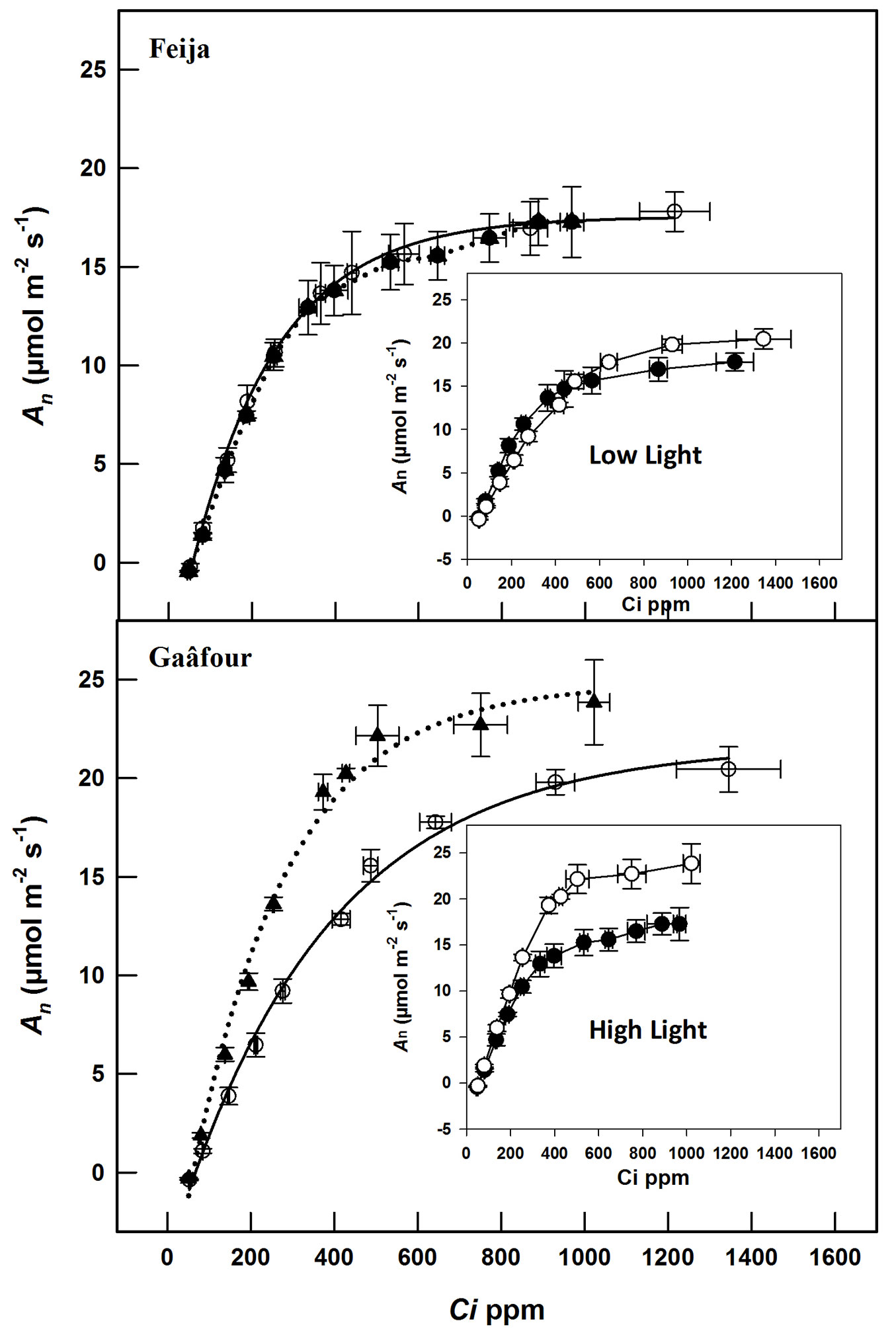
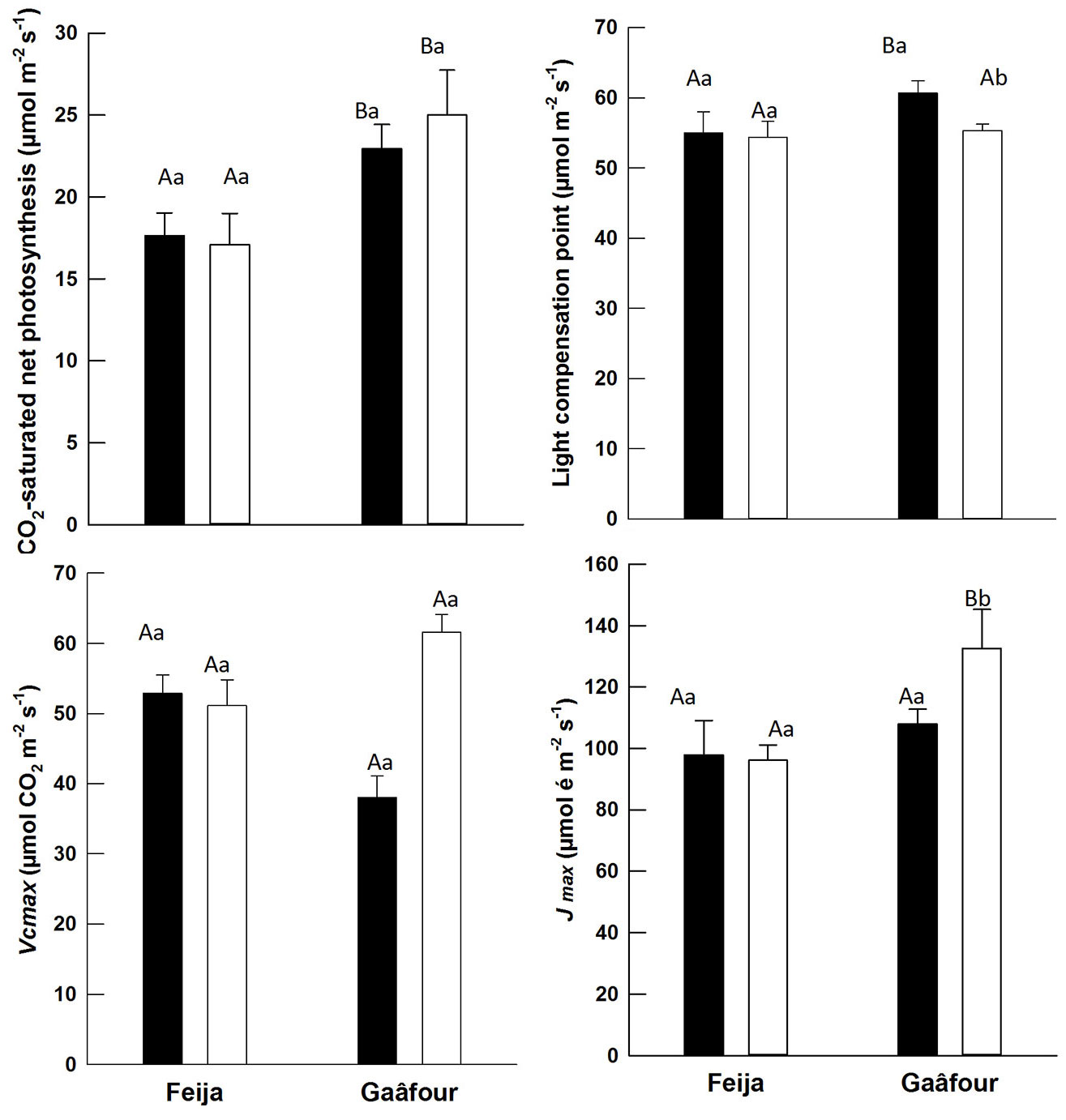
Fig. 3 shows that the CO2 compensation point (Γ) remained constant in leaves of both provenances when the leaves were transferred to full sunlight, despite a negligible decrease detected in Gaâfour leaves. Under both light regimes, leaves of Gaâfour seedlings had significantly higher CO2-saturated photosynthesis rates (Asat) than those of Feija (P < 0.05). The statistical analysis (two-way ANOVA), did not show any significant treatment × provenance interaction effect on either Γ or Asat (Tab. 1).
Fig. 3 - Photosynthetic parameters estimated from An/Ci curves of leaves maintained continuously in low light (black) or transferred from low to high light (white): the CO2-saturated net photosynthesis (µmol CO2 m-2 s-1), the CO2 compensation point Γ (ppm), the maximum ribulose 1.5- bisphosphate carboxylase/oxygenase (Rubisco) carboxylation Vcmax (µmol CO2 m-2 s-1) and the potential light-saturated electron transport rate Jmax (µmol m-2 s-1). Different uppercase letters indicate a significant difference between provenances in the same light environment, whereas different lowercase letters indicate significant difference between light environments in the same provenance (P ≤ 0.05).
Estimation of Vcmax and Jmax
Stomatal constraints on photosynthesis were similar in the Feija and Gaâfour provenances, indicating that photosynthetic acclimation was curtailed at the biochemical level. The parameters describing the maximum rate of carboxylation by Rubisco (Vcmax) and the maximum rate of electron transport (Jmax) are shown in Fig. 3. In the LL leaves, Vcmax was higher in Feija than Gaâfour leaves (P< 0.05). When transferred to HL conditions, Vcmax increased significantly in the Gaâfour seedlings only. This increase of Vcmax in HL leaves of Gaâfour showed approximately a 60% enhancement compared to leaves that remained in LL conditions.
Although Jmax in LL leaves were similar in Feija and Gaâfour provenances, Feija seedlings displayed significantly lower Jmax after 40 days of growth under HL conditions. This suggests a lower capacity of electron transport in Feija leaves, leading to the limitation of RuBP regeneration. The statistical analysis showed a highly significant treatment × provenance interaction effect on Vcmax, but this interaction was non-significant on Jmax (Tab. 1).
Discussion
We studied the effect of increasing light intensity on photosynthesis in cork oak (Quercus suber L.) seedlings of two provenances differing in climatic conditions at their geographical origin. Three-year-old cork oak seedlings of both provenances, grown in pots in a common greenhouse, were randomly assigned to one of two light treatments (HL, full sunlight and LL, 15% of sunlight).
The HL leaves of Gaâfour provenance proved to have a significantly higher photosynthetic capacity (P < 0.05) when compared to the Feija leaves. Different studies showed a possible intraspecific variability and phenotypic plasticity in Cork oak as an adaptation to contrasting regional climate conditions ([2], [1], [34], [30], [21], [15]). Investigating the influence of full-sun on seedling hardiness, Calzavara et al. ([6]) suggested that the acclimation process can induce changes in physiological, anatomical, and morphological traits of plants, favoring their establishment after transplantation to the field. The results of the present study indicate that the lower An in the HL leaves of Feija were not related to the limited stomatal conductance. Furthermore, the higher An in Gaâfour leaves at full sunlight as compared to those of Feija, was consistently observed at all Ci values applied (Fig. 2). These findings do not exclude that diffusional limitations are important factors affecting photosynthesis. However, it does highlight that additional, non-diffusive limitations affect photosynthesis in Feija leaves compared to those of Gaâfour for seedlings transferred to HL condition. It is widely assumed that Jmax and Vcmax parameters represent the major limitations to light-saturated photosynthesis ([19]) and that the decrease in biochemical capacity (Jmax and Vcmax) can limit An at high light, possibly by affecting nitrogen partitioning to Rubisco with increasing relative irradiance ([14], [18], [12]). Mechanistic photosynthetic models may be used to determine the impact of varying environmental conditions - including those predicted to be affected by climate change - on the biochemistry of photosynthesis and carbon acquisition at the leaf and plant levels ([26]). The model by Farquhar et al. ([11]) describes limitation processes in photosynthesis.
In this study Vcmax and Jmax estimated from An/Ci curves were within the range observed for other Mediterranean species ([13], [1], [22], [38]). Correlations between the photosyntheticgas exchange parameters and SLA are shown in Tab. 2. The Rd, gs, Vcmax and Jmax were significantly and positively correlated with the An. The SLA and other gas exchange parameters were not significantly correlated with the An. This correlation analysis elucidates that Vcmax and Jmax are the major contributors in the photosynthetic acclimation to HL conditions in cork oak.
Tab. 2 - Correlations between photosynthetic gas exchange parameters. (*): P ≤0.05; (**): P ≤0.01.
| Variables | A n | Rd | g s | C i | L s | Γ | V cmax | J max | A sat | SLA |
|---|---|---|---|---|---|---|---|---|---|---|
| A n | 1 | 0.5895* | 0.6205* | -0.1450 | -0.0213 | -0.4866 | 0.8207** | 0.6425* | 0.4425 | -0.2974 |
| Rd | - | 1 | 0.6044* | 0.1031 | 0.0754 | -0.0648 | 0.7062* | 0.6369* | 0.3974 | -0.4165 |
| g s | - | - | 1 | 0.2751 | -0.0041 | -0.2484 | 0.5637 | 0.4559 | 0.4237 | -0.3551 |
| C i | - | - | - | 1 | -0.0387 | 0.8235** | -0.2873 | -0.0079 | 0.3514 | 0.2816 |
| L s | - | - | - | - | 1 | 0.3311 | 0.0144 | 0.6661* | 0.0988 | 0.0550 |
| Γ | - | - | - | - | - | 1 | -0.3731 | -0.0082 | 0.3580 | 0.3768 |
| V cmax | - | - | - | - | - | - | 1 | 0.6889* | 0.4536 | -0.4946 |
| J max | - | - | - | - | - | - | - | 1 | 0.7077* | -0.3083 |
| A sat | - | - | - | - | - | - | - | - | 1 | 0.0186 |
| SLA | - | - | - | - | - | - | - | - | - | 1 |
The results from this study indicate that only Gaâfour seedlings were able to acclimate physiologically to a high light environment. This is in accordance with previous results indicating that the increase in Vcmax and Jmax reflects a physiological plasticity at the cell levels caused by an increase in the photosynthetic capacity per unit of leaf tissue ([14]). The high photosynthetic capacity of HL leaves of Gaâfour was accompanied by a significant increase (P < 0.05) of the dark respiration.
A previous study showed that a large investment of nitrogen in photosynthetic enzymes (especially Rubisco) supports the increase of photosynthetic capacity in spinach leaves transferred from low to high growth irradiance ([36]). It also increases the respiration rate in wheat leaves ([20]). [23] suggest that this increase in respiration rate in Chenopodium album is related to the maintenance processes of leaves, e.g., protein turnover which consumes respiratory energy. The high dark respiration in HL leaves confirms the acclimation ability of Gaâfour seedlings to full sunlight, since variations in respiration have been widely proposed as a component of acclimation to photon availability ([25]). Similar to An, the Rd was significantly and positively correlated to gs, Vcmax and Jmax.
A remarkable finding of the experiment was that the SLA did not differ among provenances or light treatments (Fig. 1), suggesting that leaf function can acclimate to changing light conditions despite no structural or morphological adjustments. A very common morphological response to high light is a lower SLA (or high leaf mass area), which is generally due to increased leaf thickness ([5]). However, high photosynthetic capacity is not always accompanied by thicker leaves ([23]). Oguchi et al. ([23], [24]) explained the mechanism by showing that the transfer from low to high light increases the area of chloroplasts facing the intercellular space. The mesophyll cells of LL leaves had opened spaces along cell walls where chloroplasts were absent, which enabled the leaves to increase the photosynthetic maximum rate when they were exposed to high light. Leaf thickness determines an upper limit to the photosynthetic maximum rate of leaves subjected to a change from low to high light conditions. Shade leaves would only increase the photosynthetic maximum rate when they have open space to accommodate chloroplasts which elongate after light conditions improve, which possibly was not the case in Feija leaves. No significant correlation was observed between SLA and the gas exchange parameters (Tab. 2).
In conclusion, this finding corroborates previous results ([1], [29]) showing a large provenance-level differentiation in cork oak with provenance from dry places exhibiting the higher tolerance.
List of abbreviations
Asat: CO2-saturated photosynthesis; HL: high light; LL: low light; PAR: photosynthetically active radiation; An: net photosynthesis; PPFD: photosynthetic photon flux density; LED: light emitting diode; Ca and Ci: external and intercellular CO2 molar fractions; gs: stomatal conductance; Γ: CO2 compensation point; Ls: stomatal limitation; Jmax: maximum electron transport rate; Vcmax: ribulose 1-5 biphosphate carboxylation.
Acknowledgments
This work was supported by the National Research Institute for Rural Engineering, Waters, and Forestry in Tunis, Tunisia. We wish to thank Dr. Peter Streb (UniversitéParis-Sud 11, Ecologie, Systematique et Evolution, UMR-CNRS 8079, Orsay, France) for carefully reading the manuscript. We sincerely thank Mr. Ahmed Al-Fatlawi for his review of the English language of this manuscript.
References
Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Authors’ Info
Authors’ Affiliation
Abdelhamid Khaldi
Zouheir Nasr
Laboratoire de Gestion et de Valorisation de Produits Forestiers (LGVPF), Institut National de Recherche en Génie Rural, Eaux et forêts (INRGREF, Tunisia)
Laboratoire des Ressources Sylvo-pastorales, Institut Sylvo-pastoral de Tabarka, Université de Jendouba (Tunisia)
Laboratoire de Biophysique et Technologies Médicales, Institut Supérieur des Technologies Médicales de Tunis, Université de Tunis El-Manar (Tunisia)
Laboratoire des Plantes Extrêmophiles (LPE), Centre de Biotechnologie de Borj Cédria (CBBC), BP 901, Hammam-Lif 2050 (Tunisia)
Corresponding author
Paper Info
Citation
Rzigui T, Cherif J, Zorrig W, Khaldi A, Nasr Z (2017). Adjustment of photosynthetic carbon assimilation to higher growth irradiance in three-year-old seedlings of two Tunisian provenances of Cork Oak (Quercus suber L.). iForest 10: 618-624. - doi: 10.3832/ifor2105-010
Academic Editor
Silvano Fares
Paper history
Received: May 07, 2016
Accepted: Mar 06, 2017
First online: May 17, 2017
Publication Date: Jun 30, 2017
Publication Time: 2.40 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2017
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 52544
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 43464
Abstract Page Views: 3396
PDF Downloads: 4222
Citation/Reference Downloads: 62
XML Downloads: 1400
Web Metrics
Days since publication: 3332
Overall contacts: 52544
Avg. contacts per week: 110.39
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2017): 8
Average cites per year: 0.89
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Photosynthesis of three evergreen broad-leaved tree species, Castanopsis sieboldii, Quercus glauca, and Q. myrsinaefolia, under elevated ozone
vol. 11, pp. 360-366 (online: 04 May 2018)
Research Articles
Light acclimation of leaf gas exchange in two Tunisian cork oak populations from contrasting environmental conditions
vol. 8, pp. 700-706 (online: 08 January 2015)
Research Articles
Drought tolerance in cork oak is associated with low leaf stomatal and hydraulic conductances
vol. 11, pp. 728-733 (online: 06 November 2018)
Research Articles
Links between phenology and ecophysiology in a European beech forest
vol. 8, pp. 438-447 (online: 15 December 2014)
Research Articles
SimHyb: a simulation software for the study of the evolution of hybridizing populations. Application to Quercus ilex and Q. suber suggests hybridization could be underestimated
vol. 11, pp. 99-103 (online: 31 January 2018)
Research Articles
Stomatal morphometry of Andean species and their relationship with spatial variation
vol. 18, pp. 327-334 (online: 03 November 2025)
Research Articles
A new approach to ozone plant fumigation: The Web-O3-Fumigation. Isoprene response to a gradient of ozone stress in leaves of Quercus pubescens
vol. 1, pp. 22-26 (online: 28 February 2008)
Research Articles
Carbon storage in degraded cork oak (Quercus suber) forests on flat lowlands in Morocco
vol. 9, pp. 125-137 (online: 08 August 2015)
Research Articles
The use of branch enclosures to assess direct and indirect effects of elevated CO2 on photosynthesis, respiration and isoprene emission of Populus alba leaves
vol. 1, pp. 49-54 (online: 28 February 2008)
Research Articles
Delineation of seed collection zones based on environmental and genetic characteristics for Quercus suber L. in Sardinia, Italy
vol. 11, pp. 651-659 (online: 04 October 2018)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







