
Forest growth and climate change: evidences from the ICP-Forests intensive monitoring in Italy
iForest - Biogeosciences and Forestry, Volume 4, Issue 6, Pages 262-267 (2011)
doi: https://doi.org/10.3832/ifor0596-004
Published: Dec 13, 2011 - Copyright © 2011 SISEF
Research Articles
Collection/Special Issue: COST Action FP0903 (2010) - Rome (Italy)
Research, monitoring and modelling in the study of climate change and air pollution impacts on forest ecosystems
Guest Editors: E Paoletti, J-P Tuovinen, N Clarke, G Matteucci, R Matyssek, G Wieser, R Fischer, P Cudlin, N Potocic
Abstract
A few concurrent and/or counteracting factors (increase of average air temperature, rainfall shortage, drought, CO2 enrichment, ozone, nitrogen fertilization, sulphate deposition) drive today the soil-tree-atmosphere relationships in the Mediterranean area. Radial stem growth measured within the ICP-Forests level II Italian network provides a sensitive response to these occurrences. Climate fluctuations and repeated anomalous seasons or extreme events are the major evidences of the change in progress. The 2003 heat wave is the main event occurred in this decade over large part of Europe. In Italy, it provoked a marked water deficit coupled to high air temperature, which resulted in a heavy water stress. The growth performance in different forest types was analyzed in this paper: growth rate in 2000-2004, compared with 1997-1999, showed reductions up to 50% on half of the plots examined. Most of them were: (i) in northern-central Italy within the southern continental border of the climatic deviation; (ii) at low elevation, which is more sensitive to high air temperature and drought; (iii) made up of deciduous broadleaved forests (beech and oaks), i.e., species showing growth effects also in the following year. Over the time-window 2005-2009, a significant growth decrease was vice-versa detected within the coniferous spruce forests located at medium-high elevation in the Alps. Repeated seasonal deviations in temperature and rainfall were recorded in the Alps in 2005-2009. Climate effects at local scale were examined in a site where two oak species with a different auto-ecology (sessile oak and Turkey oak) grow together. The on-going change produced heavy mortality and reduced the growth of the more demanding and less drought-tolerant sessile oak; in a few years Turkey oak became prevailing in the stand composition and structure.
Keywords
Climate fluctuations, Extreme events, Monitoring, Stem growth
Introduction
Trees react to environmental changes within their auto-ecological tolerance and search for a new equilibrium. Climate change, acts as a driving force on ecosystem processes with direct and indirect feedbacks ([3]). Further concurrent or counteracting factors, i.e., CO2 enrichment, ozone level, nitrogen fertilization, sulphates deposition, drive today the soil-tree-atmosphere relationships ([2], [25], [27], [32], [34], [35]) and the specific tree growth within each geographical area. As a feedback, changes in patterns of tree growth can have a huge impact on atmospheric and biogeochemical cycles, climate change, and biodiversity ([29]). The environmental changes also affect forests indirectly via the occurrence of diseases, pathogens and insect outbreaks ([17], [9], [4]).
Mediterranean forests are particularly sensitive to global change ([33], [6], [14]). Future scenarios predict an increase in drought stress (with increased temperature and decreased precipitation) throughout Europe (e.g., the 2003 heat wave - [31]), the Mediterranean region being particularly affected. The climatic trends in the area during the last 50 years have been characterized by a rise in mean temperature (2-4° C) and an increase in both frequency and intensity of severe droughts ([19], [20]). Drought is one of the most important factors triggering both temporary declines and mortality in temperate forests ([18]).
Current climate change is made by long-term variations in average parameters, as well as by the increased occurrence of anomalous years and extreme events. Whilst the long-term effects require extended data sets, the consequence of anomalies (usually one season, one year as a maximum) can be traced in the short-term. Tree growth, a quantitative indicator of tree vitality and ability to buffer environmental constraints ([7]), provides a sensitive response to impacts. Forest growth is among the parameters measured within the intensive (level II) forest monitoring of UN/ECE ICP-Forests (International Co-operative Programme on Assessment and Monitoring of Air pollution effects on Forests), aimed at analyzing the effects of air pollution and other stress factors on the conditions of forests in Europe ([26]). The programme, now focused on climate change, ozone effect on forests and biodiversity loss ([30]), was carried out (2009-2010) under the LIFE+ Futmon project.
Aims of this paper are to: (1) examine radial stem growth at ICP-Forests level II plots in Italy over the last 13 years (1997-2009); (2) find out the impact of the 2003 heat wave; (3) highlight the growth trend and occurrence of regional climate fluctuations over the period 2004-2009; (4) analyze the impact of climate deviations on stand structure and specific composition at a case study.
Materials and methods
The data set was made up of 31 plots, divided into four main types, i.e., beech high forests (A), spruce forests (B), beech and oak coppice forests (C), and transitory crops, i.e., coppice forests undergoing conversion into high forest (D). Detailed information on sites and stand characteristics are reported in Fabbio & Amorini ([10], [11]), Fabbio et al. ([12], [13]) and Petriccione ([30]). Status and change in key meteorological variables up to 2005 are in Amoriello & Costantini ([1]). The analysis of meteorological elements, i.e., mean air temperature and precipitation, was updated to 2009.
Individual tree measurements were carried out over the dormant season in 1996-97, 1999-2000, 2004-2005, and 2009-2010. Tree size and its change at plot level were used as response variable. Growth was expressed as basal area (b.a.) and basal area increment (b.a.i.); growth variation over time was computed as annual b.a.i. as percent of b.a. at the beginning of each growth period. The percentage ratio between subsequent b.a.i. (2000-2004 vs. 1997-1999 and 2009-2005 vs. 2000-2004) was used to assess evidence and magnitude of growth variation. Growth reduction was ranked as 1 to 15%, 15 to 30%, more than 30%.
A case study (plot EMI1) located at the southern border of the Po Valley (northern Italy) was selected because of the presence of two deciduous oaks, Quercus cerris (Turkey oak) and Q. petraea (sessile oak), with a different auto-ecology. Climatic series and tree ring chronologies were available since 1977 (Bertini & Amoriello, internal report 2009). Ring widths were measured to the nearest 0.01 mm on cores from 20 dominant trees for each species, using LEGA Dendrochronograph SMIL 3 equipment and software package (SMIL 3.1 - [28]). Both the species being aged likewise, the analysis was performed on raw datasets without any indexing ([15]).
Results and discussion
Tree growth-climate relationships atregional level
The overall growth in terms of b.a.i. was between 0.5 and 3.5% (Fig. 1). Growth rate differed depending on forest type, i.e., coppice forests > transitory crops > spruce forests > beech high forests. Changes occurred even within each type at different inventories, with an overall trend toward growth reduction.
Fig. 1 - Mean and range of periodical basal area increment as percent of basal area at the beginning of each period per forest type (A-D) and inventory (1999-2004-2009).
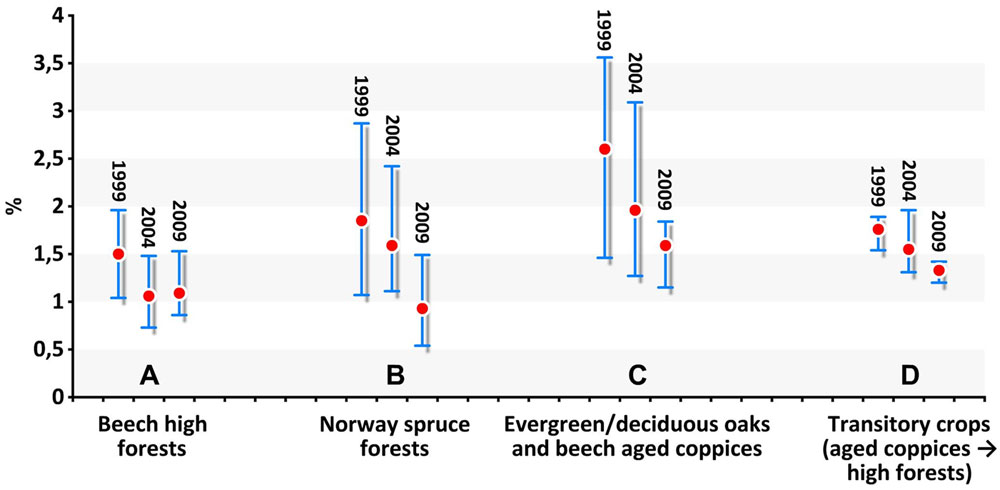
In central Europe, a number of studies have shown increasing forest growth over the last 15 years rather than a decline as was expected due to negative effects of air pollution ([36], [22]). In contrast, the extreme drought of 2003 led to a reduction of forest growth in low-land forests (up to 60%), whilst high altitude forests showed no growth change or increased growth ([21], [8]).
A growth reduction (Fig. 1) was observed in Italy for each forest type in the period 1999-2004; a further, minor decrease, not equally distributed over the types, was also detected in the period 2005-2009. Beech forests (A) showed a steady growth rate following the former reduction, whilst spruce forests (B) showed again a clear drop, with types C and D showing minor changes. In 2005-2009, the variability of growth was lower in all types than in the previous periods, suggesting a common disturbance.
In the period 2000-2004 (Fig. 2), a marked growth reduction (more than 30 and up to 50%) was observed on about half of the plots (red-labelled in the figure), all of them being located within the southern border of the 2003 heat wave. Most of plots were at low elevations and made up of deciduous broadleaved forests (oaks and beech), i.e., tree species characterized by a predetermined early growth. This feature means that an anomalous event in the growing season (ith) will also affect the early growth of the following year (ith+1). The growth reduction was likely a consequence of heat and drought in 2003 ([13]). Similar responses occurred in other regions of Europe ([21], [8], [5], [24]).
Fig. 2 - ICP-Forests level II network in Italy and growth reductions measured over 2000-2004 at each plot.
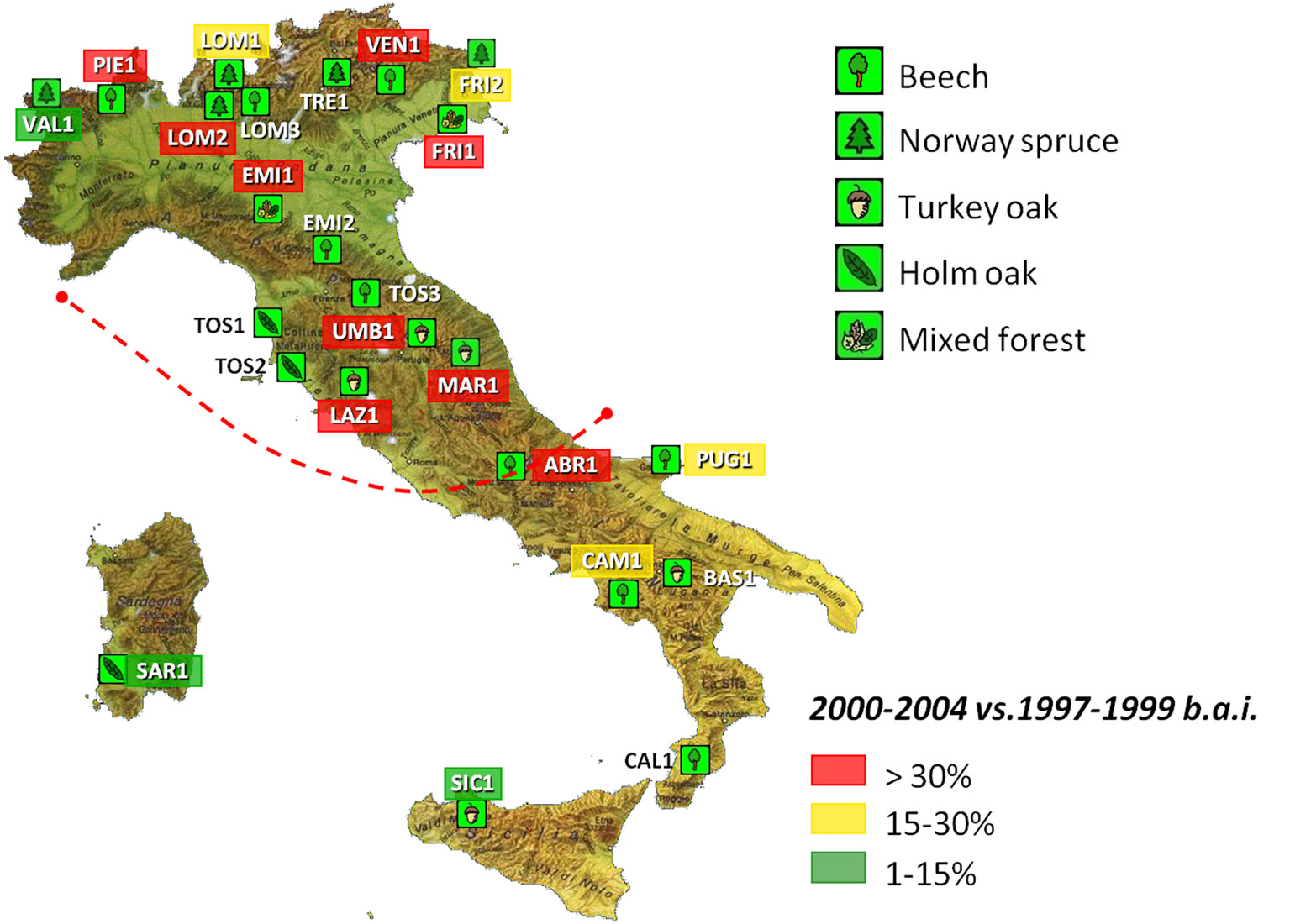
Climate anomalies occurred also before 2003; both 2000 and 2002 were very rainy years; a long dry period occurred in 2000 in northern Italy (January-February) followed by a cool and rainy summer in 2002 all over the country ([1]).
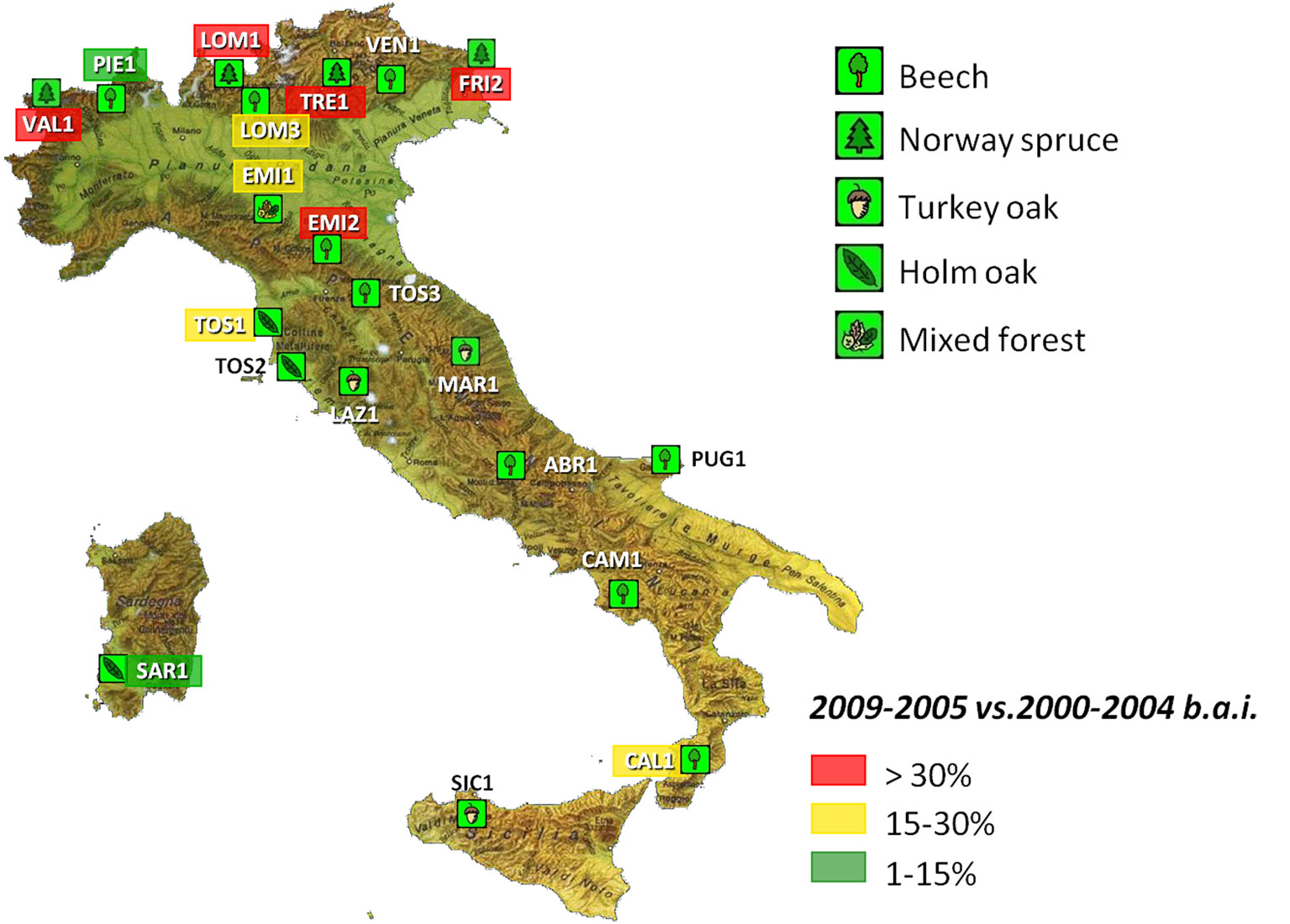
Over the last time-window (2005-2009), the more significant growth decrease was detected in the Alpine area where quite all the locations were spruce forests at medium-high elevations (Fig. 3). The analysis of climate regime over the Alpine area and northern Italy highlighted a few anomalies: a very cold winter in 2004-2005; a warm winter in 2006-2007 with the persistence of high air temperature up to late spring; a dry autumn in 2007; an unusual rise of air temperature in January-February 2008; and a rainy winter in 2008-2009 with heavy rainfalls up to mid spring. The repeated occurrence of seasonal deviations seems therefore the rule over the last decade in the Alps, in accordance with climate models of IPCC for the area (Bertini et al. unpublished).
Fig. 3 - ICP-Forests level II network in Italy and growth reductions measured over 2005-2009 at each plot.
Tree growth-climate relationships at level of a case-study
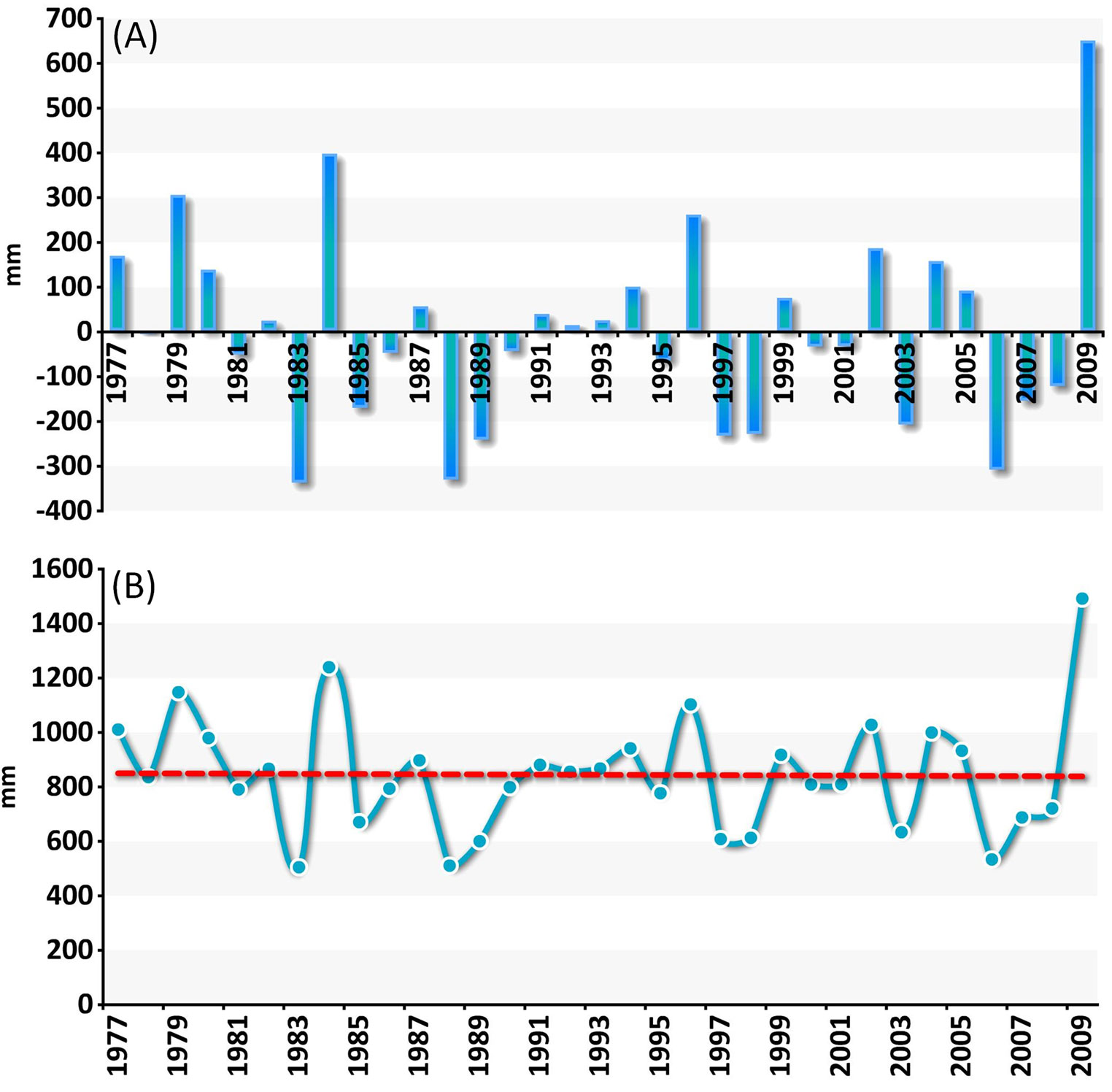
The plot EMI1, facing the southern border of the Po valley, is an interesting case study because two deciduous species with different ecological requirements - Turkey oak and sessile oak - grow together at the same site. The presence of sessile oak is not residual, as both over mature, large-sized trees and regeneration cohorts occur in the neighbourhood. No limiting factors to this oak are therefore evident at the site. Sessile oak, a mesophytic species, needs relatively high moisture content in the air and in the soil and at least a rainfall of 120 mm over the growing season. Turkey oak, a relatively drought-tolerant species, stands up to 2-4 consecutive months with reduced rainfall (< 40 mm) and up to 4 months of drought. A 30-year climatic series (1977-2009) at this site showed repeated occurrence of deviations from the mean annual air temperature and rainfall, and an increase of mean air temperature over time (Fig. 4 and Fig. 5).
Fig. 4 - Deviations and trend of mean annual air temperature at a case study plot (EMI1). (A): deviation from mean AT (1977-2009); (B): Mean AT (1977-2009).
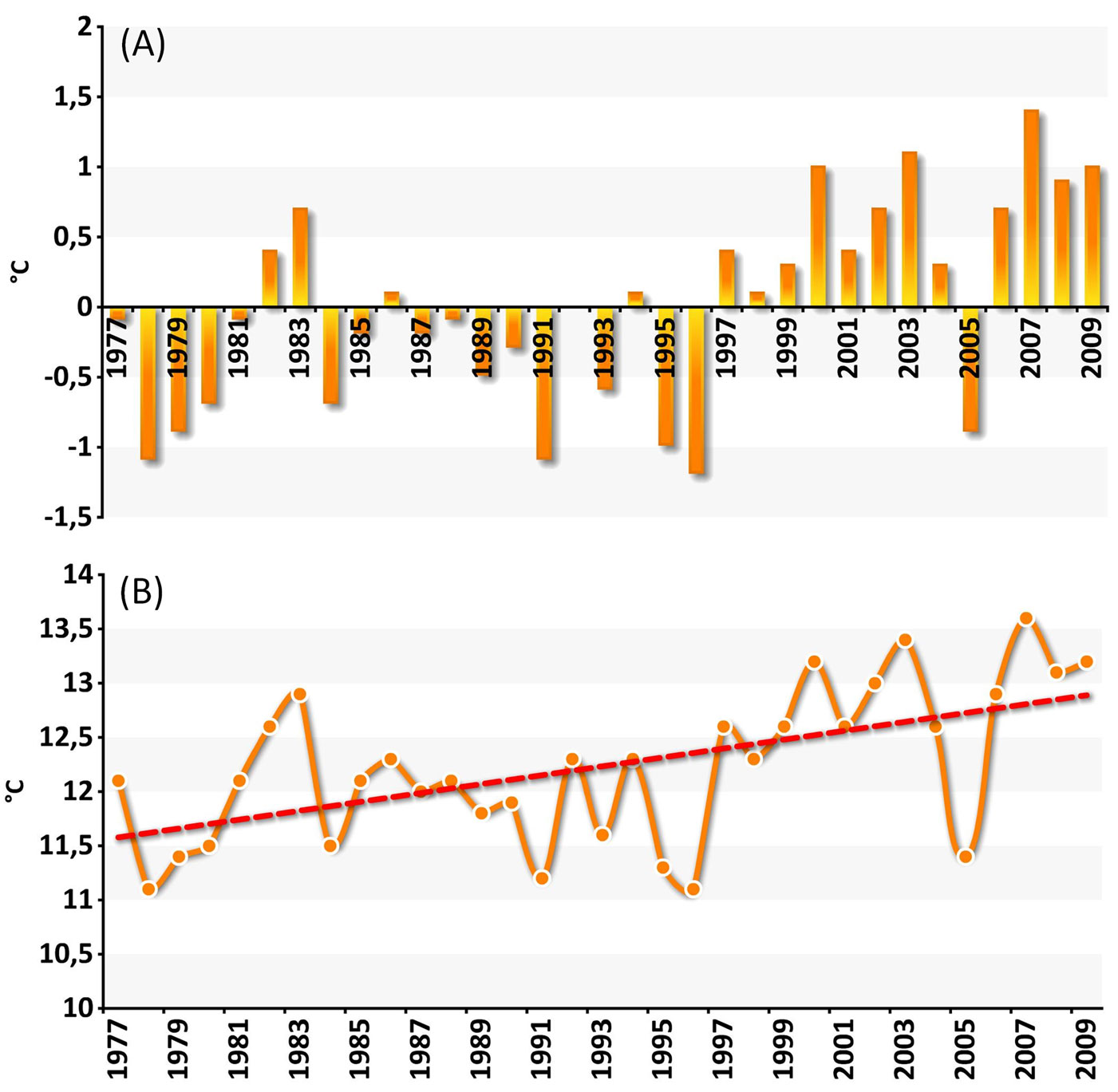
Fig. 5 - Deviations and course of mean annual rainfall at EMI1. (A): deviation from mean PR (1977-2009); (B): mean PR (1977-2009).
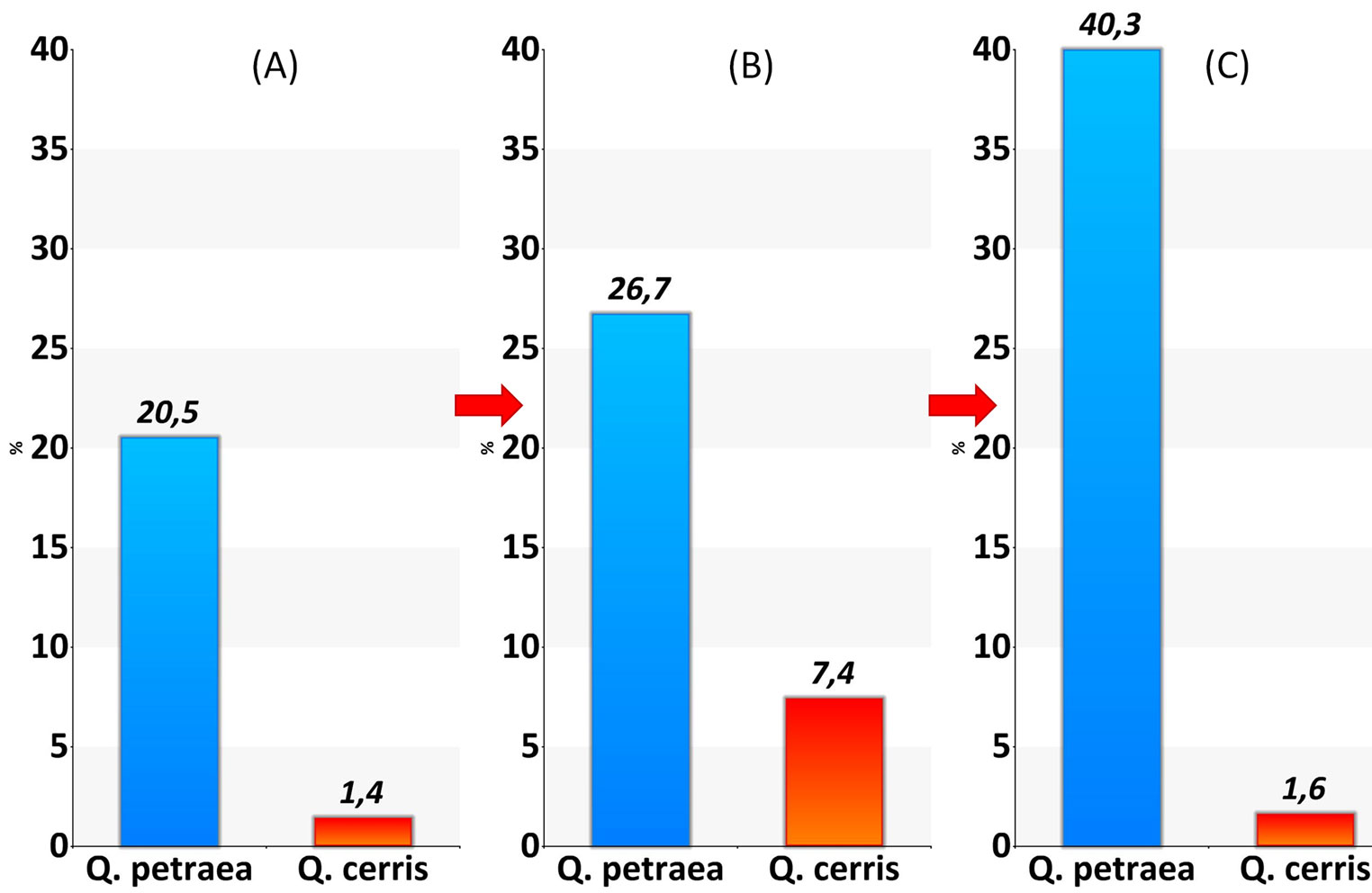
The raw mean chronologies of tree ring widths for the two same-age oak species showed a declining course in sessile oak (Fig. 6), i.e., the species that is more affected by drought and increased air temperature, suggesting a persistent water stress over the growing season. The resulting tree mortality (in terms of number of stems) over the following inventories affected sessile oak much more than Turkey oak (Fig. 7).
Fig. 6 - Tree ring widths mean chronologies of oak species at EMI1.
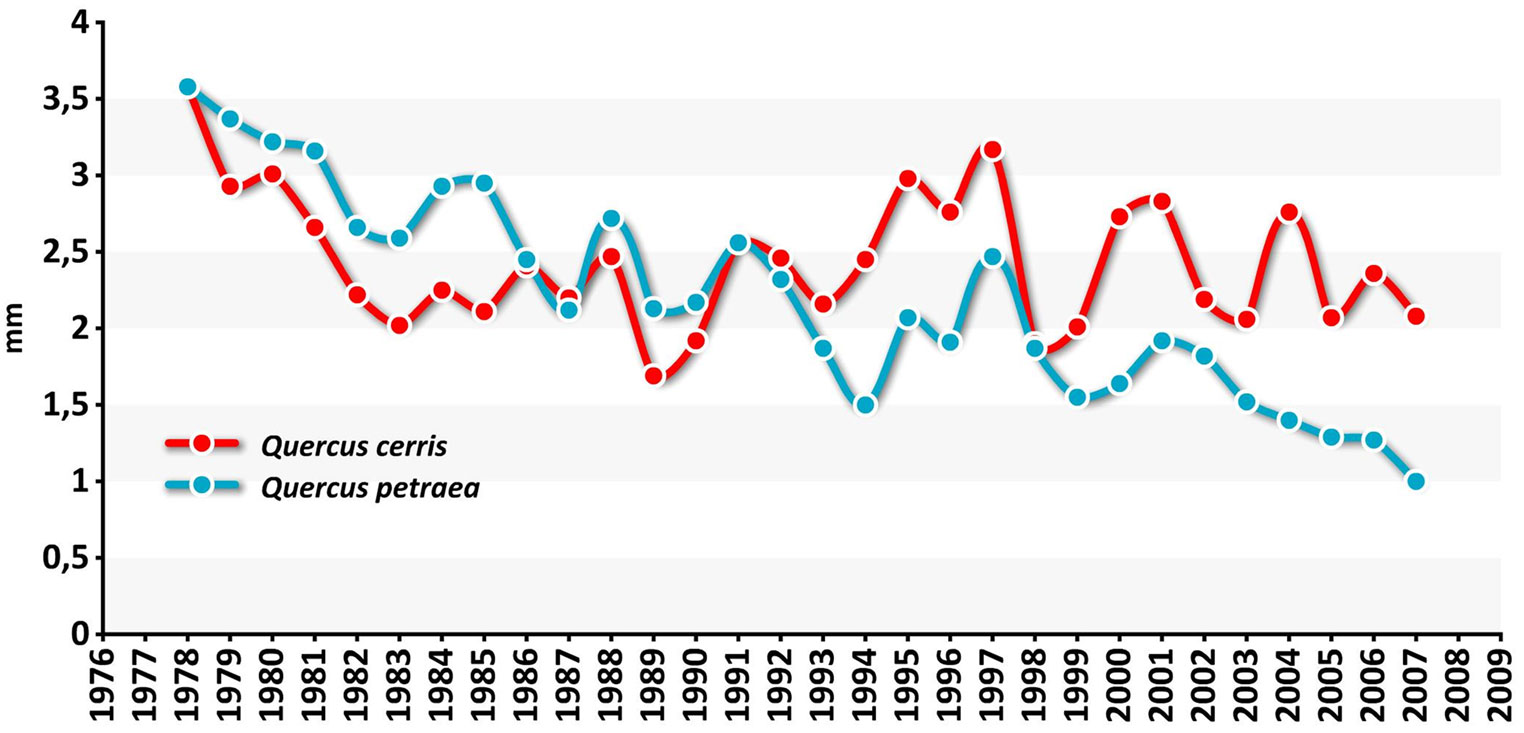
Fig. 7 - Tree mortality (% of stem number) of oak species at EMI1. (A): years 1997-2000; (B): years 2000-2005; (C): years 2005-2010.
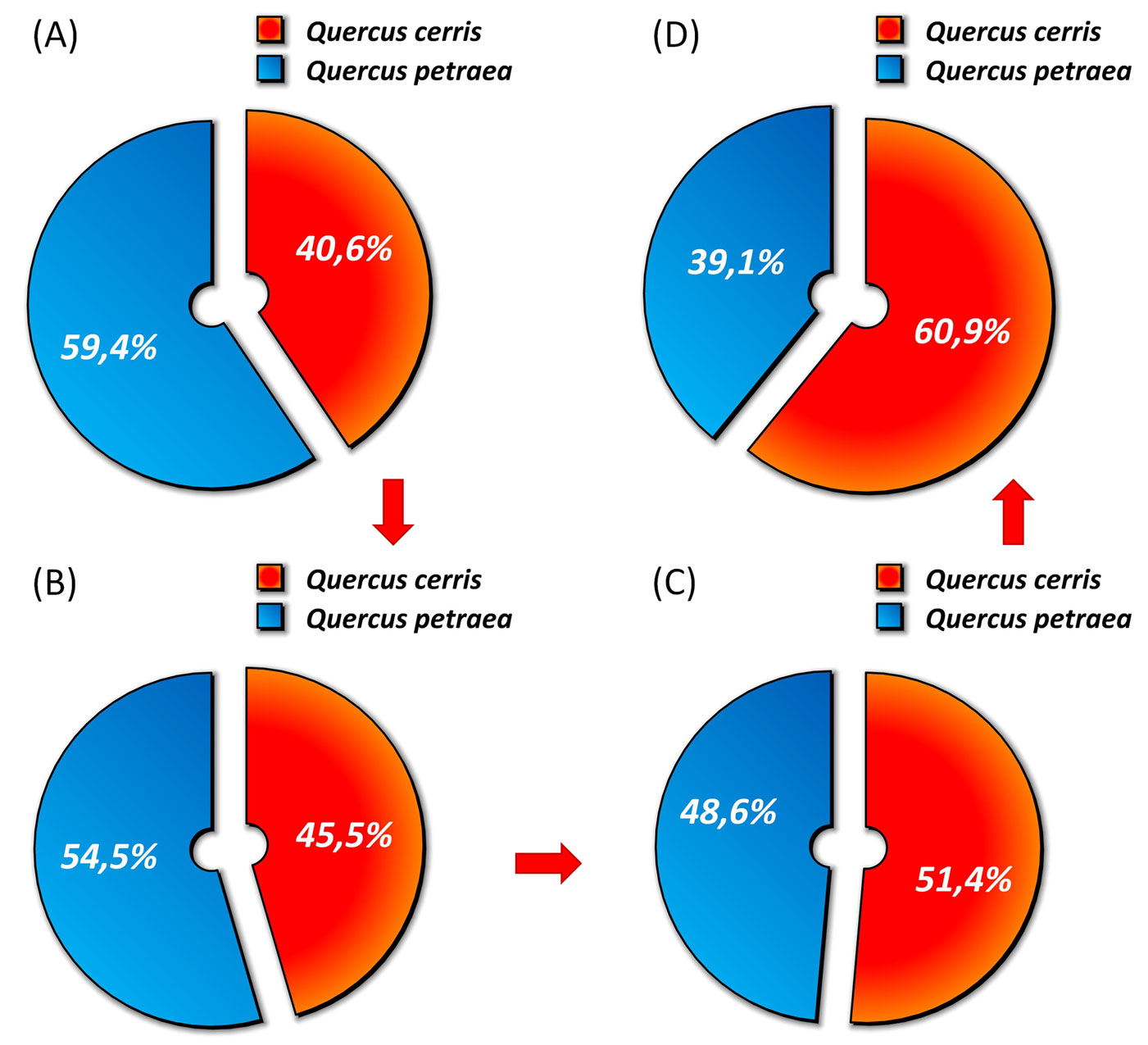
The shift towards a less favourable growth environment for the more demanding species resulted in a heavy change in structure and species composition as well. Sessile oak was the most abundant species in the stand at the beginning of the monitoring period i.e., in 1997 (59% b.a.), while in 2009 it was less abundant (39%) than Turkey oak, because the latter species was much less affected by the ongoing drier conditions (Fig. 8).
Fig. 8 - Variation of species composition in terms of % basal area over the inventories at EMI1. (A): year1997; (B): year 1999; (C): year 2004; (D) year 2009.
Intra-annual stem growth measurements by permanent tree-girth tapes highlight the radial growth pattern of each species (Fig. 9). The seasonal course was synchronous, but the growth rate was lower in sessile oak than in Turkey oak over the whole spring-summer period.
Fig. 9 - Intra-annual radial growth (2010) of the oak species at EMI1.
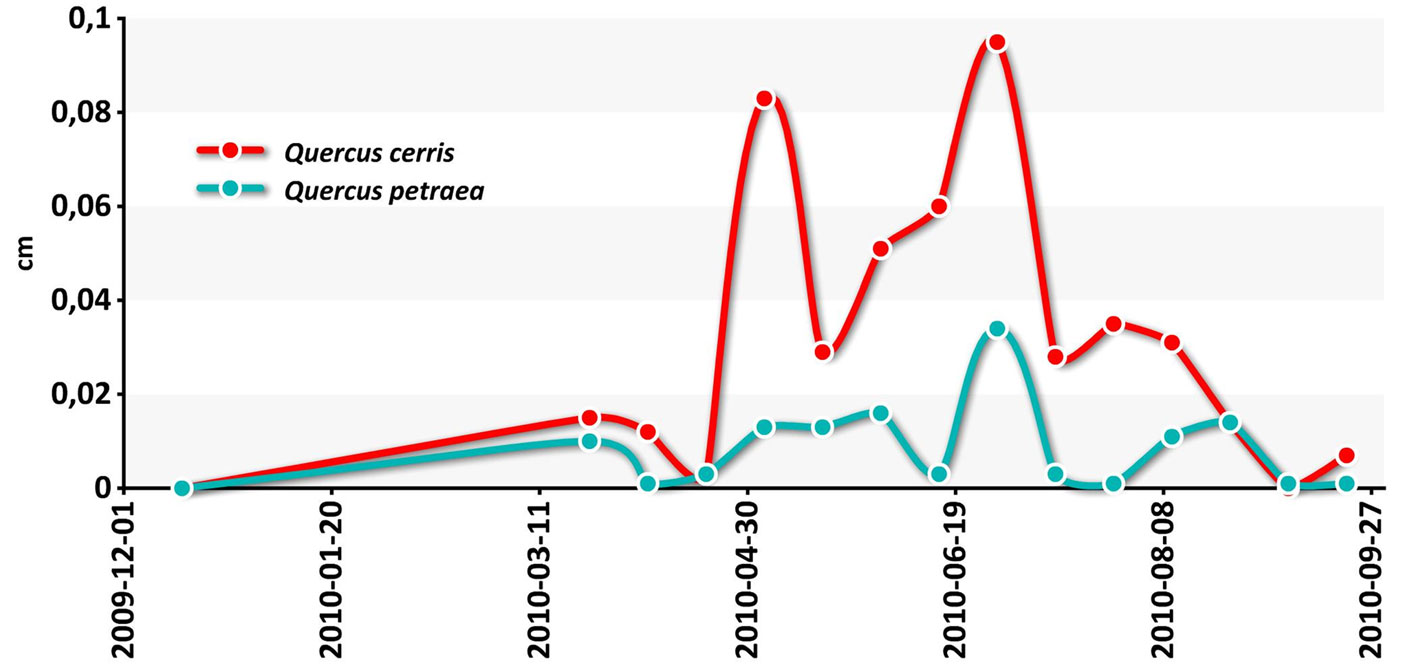
These results confirm an increased frequency of extreme drought events, potentially leading to dramatic reductions in wood productivity and changes in species composition and structure ([23], [16], [5]).
Conclusions
In addition to atmospheric chemistry, current climate fluctuations, recurrent anomalous years and extreme events throughout Europe are the main driving forces acting on forest health and vitality. The effects are registered in tree growth, a sensitive signal of tree condition. Unravelling and quantifying these complex interactions on forests is further complicated by changes at different latitudes, elevations and according to the species sensitivity. Recognizing the effect of each pressure, identifying possible feedbacks and the onset of limiting factors seem to be the priority issues today. Monitoring activity can play this role and highlight different impacts at different scales, as in the present case.
Acknowledgements
The authors wish to thank Bruno Petriccione and Enrico Pompei, National Forest Service (CFS), the technical staff of the National Forest Service, the territorial administrations and Regional parks working at the plots, Angelo Vanini, Antonia Cavalieri and Nicola Donetti at Parco Regionale “Boschi di Carrega” (EMI1), for their professional and helpful field assistance.
The monitoring activity is currently funded within the project LIFE+ ENV/DE/000218 “FUTMON”.
References
Gscholar
Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Authors’ Info
Authors’ Affiliation
Consiglio per la Ricerca e la Sperimentazione in Agricoltura, Roma (Italy)
Corresponding author
Paper Info
Citation
Bertini G, Amoriello T, Fabbio G, Piovosi M (2011). Forest growth and climate change: evidences from the ICP-Forests intensive monitoring in Italy. iForest 4: 262-267. - doi: 10.3832/ifor0596-004
Paper history
Received: Nov 26, 2010
Accepted: Aug 12, 2011
First online: Dec 13, 2011
Publication Date: Dec 13, 2011
Publication Time: 4.10 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2011
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 66891
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 55110
Abstract Page Views: 4914
PDF Downloads: 5234
Citation/Reference Downloads: 44
XML Downloads: 1589
Web Metrics
Days since publication: 5307
Overall contacts: 66891
Avg. contacts per week: 88.23
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2011): 50
Average cites per year: 3.33
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Short Communications
Climate effects on growth differ according to height and diameter along the stem in Pinus pinaster Ait.
vol. 11, pp. 237-242 (online: 12 March 2018)
Research Articles
Extreme climatic events, biotic interactions and species-specific responses drive tree crown defoliation and mortality in Italian forests
vol. 17, pp. 300-308 (online: 30 September 2024)
Review Papers
Should the silviculture of Aleppo pine (Pinus halepensis Mill.) stands in northern Africa be oriented towards wood or seed and cone production? Diagnosis and current potentiality
vol. 12, pp. 297-305 (online: 27 May 2019)
Research Articles
Tree-oriented silviculture: a new approach for coppice stands
vol. 9, pp. 791-800 (online: 04 August 2016)
Research Articles
Large wildland fires and extreme temperatures in Sardinia (Italy)
vol. 7, pp. 162-169 (online: 14 February 2014)
Review Papers
Linking patterns of forest dieback to triggering climatic and weather events: an overview on Mediterranean forests
vol. 17, pp. 309-316 (online: 30 September 2024)
Research Articles
Climatic fluctuations trigger false ring occurrence and radial-growth variation in teak (Tectona grandis L.f.)
vol. 9, pp. 286-293 (online: 28 September 2015)
Research Articles
Allometric relationships for predicting the stem volume in a Dalbergia sissoo Roxb. plantation in Bangladesh
vol. 3, pp. 153-158 (online: 15 November 2010)
Research Articles
A rapid method for estimating the median diameter of the stem profile of Norway spruce (Picea abies Karst) trees
vol. 10, pp. 328-333 (online: 11 February 2017)
Research Articles
Hearing nature’s heartbeat: towards large-scale real-time forest monitoring network in Italy
vol. 18, pp. 202-211 (online: 09 August 2025)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword






