
Linking patterns of forest dieback to triggering climatic and weather events: an overview on Mediterranean forests
iForest - Biogeosciences and Forestry, Volume 17, Issue 5, Pages 309-316 (2024)
doi: https://doi.org/10.3832/ifor4545-017
Published: Sep 30, 2024 - Copyright © 2024 SISEF
Review Papers
Collection/Special Issue: Project LIFE MODERn(NEC)
Workshop "Climate change and forest health monitoring in Italy" - Rome (Italy) May 5, 2023
Guest Editors: Bussotti F, Pollastrini M
Abstract
Mediterranean forest ecosystems are highly vulnerable to climate extremes and anomalies induced by climate change, as evidenced by the increase in forest tree decline and death over the past few decades. Forest dieback may occur with different spatial patterns and in response to many climatic and weather events (i.e., drought spells, heat waves, compound events or hot drought, cold spells, and windstorms). This paper describes the physiological behaviour of Mediterranean tree species (namely evergreen sclerophyllous trees) under extreme climatic events, with a focus on embolism formation and carbon depletion. Additionally, we describe three widely recognised patterns of forest dieback: die-off, patchy dieback, and salt and pepper dieback. Overall, drought spells, heat waves, and compound events have been reported to induce all the mortality patterns described, revealing the great vulnerability of Mediterranean forests to rising air temperature and vapour pressure deficit trends. Many intrinsic factors of plant and structural and contingent factors of the forest can interact, favouring one mortality pattern over another. Patterns of dieback and mortality are the product of the interaction between the complexity of the landscape, the species assemblage, the kind and intensity of the disturbance, and biotic factors. Different patterns have different ecological consequences, and implications for monitoring and management. The combination of terrestrial and remote sensing (RS) is recommended.
Keywords
Mediterranean Forest, Forest Mortality, Climate Change, Drought-induced Dieback
Introduction
Mediterranean ecosystems have prolonged summer droughts and rocky soils, with poor nutrient content, especially nitrogen and phosphorous. They are biodiversity hotspots and host many plant and animal species. The species richness is the result of different migration pathways during the glacial age. Many species evolved under different conditions and were subsequently adapted to the Mediterranean conditions. Differently from temperate and boreal forests, the Mediterranean forest landscape consists of several isolated and usually small-sized patches of woody plant communities ([67]). Mediterranean forests are subjected to many anthropic pressures, such as grazing, extraction of charcoal wood, forest fires, fragmentation, urbanization, air and water pollution. Ongoing climate change is an additional factor able to exacerbate the effects of natural and anthropic disturbances. This region was subjected to large climate changes in the past and has been identified as one of the most prominent hot spots in future climate change projections ([50]). Indeed, the Mediterranean Basin has warmed faster than the global mean (0.28 °C/decade, with an observed trend of about 0.4 °C/decade) and other Mediterranean regions (0.15-0.28 °C/decade - [28]). Warmer conditions are accompanied by the decrease of relative humidity (RH). Additionally, this trend is expected to continue over the 21st century ([75]).
Mediterranean forests and shrublands are characterized by sclerophyllous evergreen species, that are supposed to be resistant to drought. Sclerophylly, however, is related to the scarcity of soil nutrients rather than climatic factors ([8]). Sclerophyllous evergreen species are adapted to mild temperature seasonal fluctuations and to the absence of pronounced drought periods ([4]). For these reasons, Mediterranean forests are sensitive to climate change and even moderate changes in the current climatic parameters may produce significant impacts on their vitality and composition.
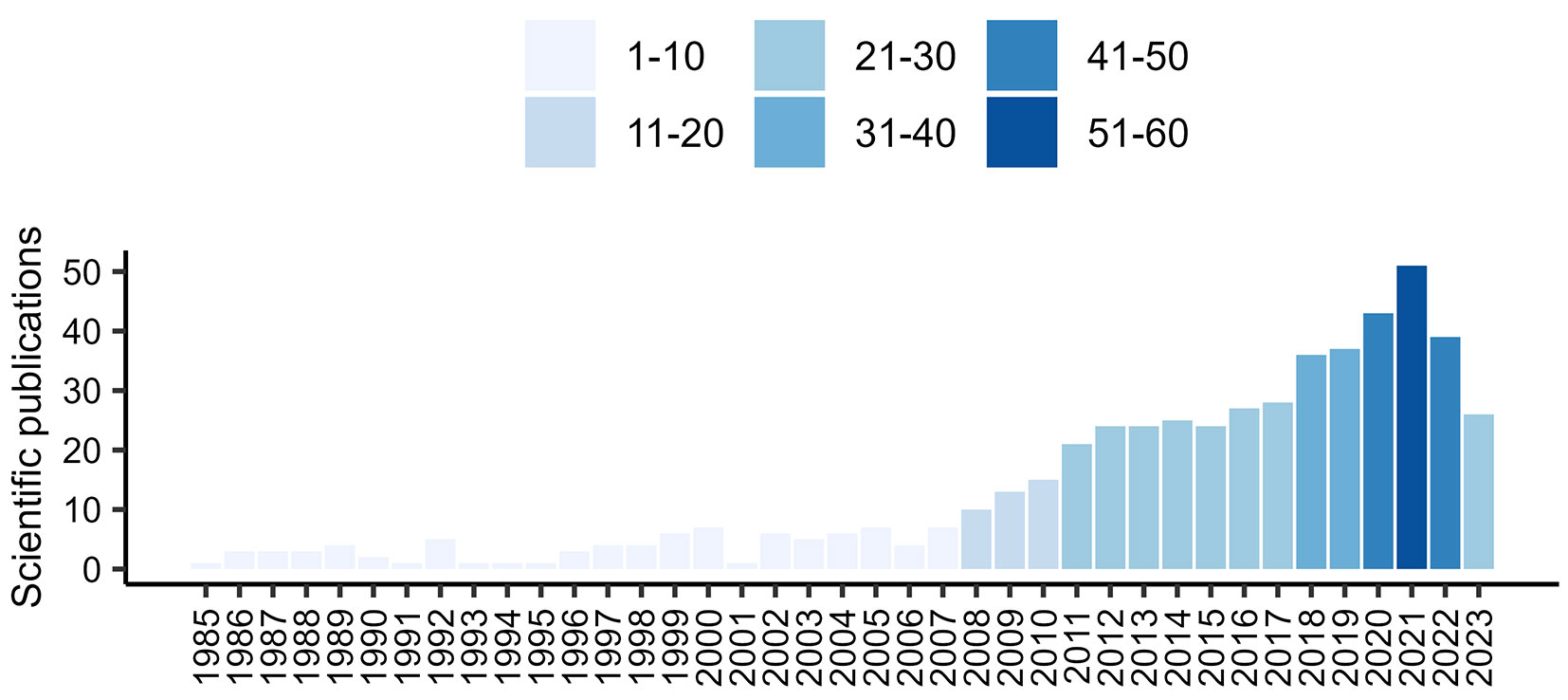
The scientific interest on Mediterranean forests dieback is rapidly rising as testified by the increasing number of scientific papers published on this topic. Searching the term “Mediterranean forest dieback” on the Clarivate Web of Science™ platform, the number of retrievable papers strongly surged over last forty years, surpassing the threshold of thirty publications per year after 2010 (Fig. 1).
Fig. 1 - Number of scientific papers published in the last years retrieved searching the term “Mediterranean forest dieback” on the Clarivate Web of Science™ platform.
In the Mediterranean Basin, the reduction in tree growth and mortality caused by extreme climatic events have been reported on thermophilus deciduous species and evergreen species, with special reference to evergreen oaks, Quercus ilex L. and Quercus suber L. ([51], [60]). The most relevant cases have been documented in Spain and Italy since 1980s, due to the extremely warm and dry summer seasons ([58], [41]), which also affected evergreen Mediterranean shrubs in some recent years (e.g., 2004, 2012 and 2017 - [14], [11]). Based on the European crown condition survey conducted within the International Co-operative Programme on Assessment and Monitoring of Air Pollution Effects on Forests (ICP Forests), the evergreen oaks displayed the strongest increase in crown damages over the past 20 years ([61]). Concerning conifer species, they showed a smaller vulnerability to crown damages in most recent years ([61]). However, some cases of reduced growth rates and increase of tree mortality have been reported for Pinus spp. in Turkey, Greece and Spain, with peaks in mortality rate in response to 2012 and 2017 droughts in lowland forests ([21]). Pinus pinaster Aiton, P. pinea L. P. sylvestris L. and P. halepensis Mill. have been reported as the most affected species by severe drought events ([52]). Additionally, tree mortality was also documented on Cedrus atlantica (Endl.) Manetti ex Carrière, which was substituted by Quercus ilex and Juniperus oxycedrus L. in North Africa and in the Eastern part of the Mediterranean basin ([62], [21]).
The interaction between the complexity of the landscape, the species assemblage, the kind and intensity of the disturbance, and biotic factors, result in different patterns of forest tree dieback and mortality. Catastrophic events, like extensive forest fires, windstorms and insect outbreaks, can result in large clearings from 10 up to 10,000 km2, i.e., “die-off” events that cause the death of most trees in a forest, regardless of species ([6] - Fig. S1 in Supplementary material). Under severe drought, heat waves and localized stress events, the heterogeneity of the landscape and the genetic structure of the stands may result in a “patchy” dieback, where the patches size can vary from a few square meters to several km2 ([13]). However, a time longer than 10 years of monitoring of patchy decay is needed to further evaluate the temporal worsening of this mortality pattern which could also result in die-off ([68] - Fig. S1 in Supplementary material). Finally, moderate stress events may induce a “salt and pepper” pattern with scattered mortality of individual trees (Fig. S1), i.e., dying trees occur in a matrix of healthy ones ([22], [25], [27], [72]). In this case, dieback and mortality hit the most sensitive species, trees and genotypes and operate a genetic selection of the populations ([2]).
Tree mortality induced by climate change is an issue of worldwide interest, across different biomes and climatic conditions ([46]). This study aims to link a forest dieback pattern to the triggering climatic/events on Mediterranean forests based on available scientific literature. The climatic and weather events able to induce forest mortality (i.e., drought spells, heat waves, compound events or hot drought, cold spells and windstorms) will be described and associated with a particular forest dieback pattern. In particular, we analyzed the ecological and physiological specificity of the responses of Mediterranean forests facing climate stress, referring mostly to the broadleaved evergreen sclerophyllous formations (high forests, maquis, garrigues).
The spatial distribution of dieback and mortality is of special importance since each pattern suggest possible different dynamics of the vegetation after the stressful event and poses specific problems connected to monitoring and management. Although many scientific publications deal with forest mortality and with Mediterranean forest mortality, we argue that a schematic and holistic picture of the issue aimed to disentangle which forest dieback pattern would be expected from some peculiar climatic or weather condition, is still missing.
Extreme climate events driving forest mortality
Drought and heat waves
Drought spells are defined as a lack of precipitation for a variable time (from days to months) in which precipitation deviates from the usual range ([66]). Generally, drought events are recognised based on summary indices, among which the Standardised Precipitation-Evapotranspiration Index and the Standard Precipitation Indices are the most employed ([83]). Such climatic indices have been adopted by scientists to identify and describe moderate and intense drought events, as those occurred in early summer and summer drought episodes in 1985-1986, 1994-1995, 2005, 2011-2012, 2015 and 2017 in southern Europe, that caused severe damages on both tree and shrub species of the Mediterranean forests ([58], [16], [41], [72], [79]).
A heat wave is defined as the highest number of consecutive days in the June-August season, when the daily maximum temperature surpasses the long-term daily 95-th percentile of the daily maximum temperature ([31]). The Mediterranean region is experiencing a 200-500% increase in the frequency of hot extremes and this is particularly hazardous in coastal areas ([48]). One of the most prominent examples of exceptional climatic conditions in the Mediterranean region is the summer of 2003 ([33]). Over the entire 2003, temperatures across the northwestern part of the European continent, southern Adriatic, and southern Europe, were above the 98-th percentile of the 1961-1990 distribution averaged over the entire year ([33]).
Both low soil water availability (i.e., delta between precipitation and potential evapotranspiration) and high temperatures are stressful conditions for forest ecosystems, leading to the simultaneous occurrence of extreme droughts and heat waves, also known as “compound events”, with unprecedented impacts on forest ecosystems ([40]). Indeed, compound events, characterized by the simultaneous occurrence of low precipitation and high temperature (thermo-pluviometric stress), leading to high VPD, are linked to local and intense tree mortality events.
Physiological responses of Mediterranean vegetation to drought and heat waves
Although being considered drought-adapted, the evergreen sclerophyllous habitus does not guarantee a greater resistance to drought than their deciduous counterparties that share the same environments. Tognetti et al. ([77]), comparing the physiological responses of Quercus pubescens (deciduous) and Quercus ilex (evergreen) in natural stands to summer water stress, concluded that both the species grow at the limits of hydraulic safety, although highly tolerant to severe water stress and tissue dehydration. Prolonged climate stress might predispose these species to physiological decline and dieback because their limits of hydraulic safety can be surpassed by frequent and repeated stress occurrence.
Deciduous species shed their leaves under extreme water stress conditions, so avoiding severe physiological consequences and protecting xylem tissue from embolism formation. This behavior was observed on xero-thermophilous deciduous tree species in central Italy, as Quercus pubescens and Fraxinus ornus L. under severe heat and drought waves ([11]). On the other hand, in evergreen species retaining the same leaves for multiple years, vulnerability to extreme drought is related to the xylem characteristics and wood anatomy, including wood density, vessel diameter, pith size, number of mechanical elements, like fibers, and non-structural carbohydrates storage ([80], [70]). In addition, the recovery capacity from xylem embolism plays a crucial role in sclerophyllous trees responses to drought stress.
On a daily time scale, the raising values in xylem water potential from midday to night has been suggested to allow a partial embolism recovery in well-watered and droughted plants thank to bubbles dissolution in the xylem sap under low xylem tension, while a complete recovery of xylem functionality after drought was possible only in rewatered plants at low transpiration ([70]).
The great elasticity of the xylem cavitation strain of sclerophyll species depicts tree response to a large drop in stem water potential during the day, followed by night recovery ([77]). This usually occurs in Mediterranean sclerophylls because they grow with high irradiance and air temperature during the day and with high air humidity condensing on the soil surface at night. Moreover, it has been observed that Mediterranean sclerophylls are able to recover xylem hydraulic conductivity from drought induced embolism thank to the production of new functional xylem, though being a slow process ([42]). Therefore, the resistance of the Mediterranean deciduous species is especially connected to their phenology (loss of leaves during the dry summers), whereas the resistance of evergreen sclerophyllous species is connected to their high recovery capacity. Thus, in the Eastern Mediterranean where RH is lower, the presence of summer deciduous species such as Phlomis fruticosa L. increases ([44]).
Alon et al. ([5]) compared five dominant Mediterranean species (i.e., Quercus calliprinos L., Pistacia terebinthus L., Pistacia lentiscus L., Rhamnus lycioides L. and Phillyrea latifolia L.) that experienced raising levels of embolism along a drought gradient. The authors highlighted that these Mediterranean species under drought conditions can harmfully reach the limit of their hydraulic capacity and that even a slight reduction in precipitation would increase the risk of mortality ([5], [71]). Increasing the length and intensity of drought events, with the reduction of RH, can provoke extensive and severe dieback and mortality in Mediterranean evergreen trees, as already observed in Italy and Spain ([58], [41]).
Under moderate drought stress, to avoid mortality due to hydraulic failure, plants close their stomata reducing the photosynthetic carbon uptake, thereby leading to a decrease in NSCs (non-structural carbohydrates) concentration in plant tissue and to the breakdown of stored starch into soluble sugars to supply substrates and energy for primary and secondary metabolism ([10], [69]). A pot experiment on Q. ilex seedlings showed the disappearance of starch grains in wood parenchyma cells in water-stressed plants, accompanied by the upregulation of homologous genes involved in starch catabolism ([42]). Thus, a prolonged negative carbon balance can result in carbon-starved dead trees ([32]).
The heat wave of 2003 caused visible declining symptoms (i.e., foliage yellowing and browning, branch dieback and premature leaf loss) in many forests stands both at local and regional scale in Mediterranean and Middle European region, leading to both patchy dieback and die-off ([56], [34], [76]). Some authors proposed that during heat stress, the enhancement of respiration processes may lead tree die due to a temperature-driven carbon starvation ([1]). Alternatively, Cochard ([24]) proposed that high temperatures may lead trees to death due to the increase in residual plant transpiration (gmin) imposed by high VPD that may significantly contribute to the formation of runway embolism resulting in hydraulic failure. Isolating the impact of heat stress on forests is challenging as a certain degree of thermal stress (heat stress) often accomplices drought episodes, as in the case of heat waves of 2003 and 2018 ([56], [76], [73]). To underline the predominant components of temperatures on those climatic extremes, other authors have used the terms “heat driven drought” to refers to a dry period in which the most detrimental effect to forest is imposed by high temperatures, rather than low precipitations ([63]). Although it is not clear how plants die under heat stress, what is established is that the worst impacts on vegetation occur when heat and drought stress co-occur ([24]).
Recent studies ([40]) evidenced that in the last decades drought and heat waves are accompanied by low relative humidity, with increasing vapour pressure deficit (VPD). These are the so-called “compound events”, that are responsible of the increased mortality in European forests ([40]. The authors stated that the increase in background mortality rates was associated with an increase in summer VPD, imposed by increasing temperatures. Thus, given the well-known temperature increasing trend, moderate droughts can have a significant impact on forest health ([81]). The increasing atmospheric water demand has been established as a key element driving plant mortality at least equally important as the reduced soil moisture. Furthermore, heat stress and high VPD may further worsen tree vitality in individuals that are already experiencing hydraulic dysfunction and carbon starvation.
Increasing precipitation deficit, higher temperature and reduced relative humidity act simultaneously in the Mediterranean forests, therefore it is hard to disentangle the relative role of each factor in inducing tree dieback and mortality. The combination of these factors affects gas-exchanges processes, with increasing risks of xylemic embolism and carbon starvation. Additionally, a temporary reduction of carbon reserves may turn trees more vulnerable to other unexpected biotic and abiotic disturbances, as the availability of NSCs could not reach the demand to face the stress ([47]).
Declining trees are often attacked by opportunistic parasites. Mortality in oak species is often associated to attacks of the endophyte fungal parasite Biscougnautia mediterranea (De Not.) Kuntze ([20]), or to the oomycete Phytophthora sp.pl. ([26]).
Cold spells
Although heatwaves and droughts are the most common issues in the Mediterranean region, a few cold spells have been reported in the eastern section (Balkans - [55], [48]). A cold spell is defined as the number of consecutive days with minimum temperature below the 10-th percentile of the temperatute distribution ([85]). Climatic studies have shown a significant decrease in the number and duration of cold spells in the eastern Mediterranean region ([84]). Despite the limited number of studies in Mediterranean Europe, cold spells have been reported to induce a patchy dieback characterized by decline in the radial growth of Quercus robur at wet sites in northern Spain ([17]). Spring frost may induce strong defoliation, increasing the risk of mortality due to carbon starvation, as reported in the Fagus sylvatica L. forests of Central Italy ([29]). In another case of study, the passage of storm Filomena across the Iberian Peninsula at the beginning of 2021 (when temperatures reached the minimum of -21 oC) caused intense crown defoliation on Pinus halepensis and Quercus ilex forest stands in central and eastern Spain, accompanied by reduced radial growth rate and diminished sapwood NSC ([18]). Additionally, although not directly measured in the study, authors did not exclude that freeze-thaw cycles may have also caused hydraulic dysfunctions ([18]). Thus, at least in Mediterranean and temperate forests, both climate extremes, such as dry and cold spells, may potentially induce a patchy dieback mortality pattern characterized by radial growth decline.
Physiological responses to cold spells
Droughts and heat waves are common in seasonally dry regions, such as the Mediterranean, though unforeseen cold spells are also possible to occur. Under global warming, enhanced variability in temperature results in more frequent, intense, and lasting heat waves as well as cold spells. It has been reported that Mediterranean tree species of subtropical origin adapted to withstand drought are highly vulnerable to cold spell disturbances ([18]). In this regard, an extreme cold event in 2001 induced severe Mediterranean Scots pine dieback over 14.000 ha of forests in eastern Spain ([18]). Hydraulic failure is the most likely mortality mechanism in plants under extremely cold conditions. Indeed, when temperatures cross the zero threshold many times, xylem tissues are exposed to freeze-thaw cycles with a high likelihood of experiencing freeze-induced embolism ([59]). Bubbles produced during the freezing of xylem water expand during thawing, inducing tension in the xylem. It is worth noting that for freeze-induced embolism (as in the case of drought-induced embolism), xylem anatomical traits affect vulnerability to hydraulic damages, with wider vessels being more prone to experience reduction in xylem conductivity ([30]). Additionally, forest site conditions can modulate the effects of cold spells on forests. It has been reported that south-facing slopes are the most affected by cold-induced dieback due to the highest diurnal temperature ranges experienced by trees, thus increasing the likelihood of xylem embolism through multiple freeze-thaw events ([15]). Additionally, if starch breakdown occurs to provide sugars to be employed as osmolytes for freezing tolerance, thus cold spells can affect tree carbon stocks diminishing the capacity of trees to withstand abiotic and abiotic stresses ([18]). A reduction in the pool of NSCs imposed by cold spells may also compromise tree resprouting capacity, which is one of the main resilience mechanisms of Mediterranean species ([3]).
Windstorm
Among the occasional forest disturbances that can induce tree mortality, windstorms must also be considered ([7]), though such events are more frequent in Atlantic and Central Europe than in the Mediterranean Region. In Mediterranean areas, environmental perturbations caused by storms, that combine the actions of strong winds, heavy rain, and/or snowfall, are relatively frequent (every 1-7 years - [65]). Nevertheless, windstorm frequency and impact in Mediterranean forests have been increasing since the mid-twentieth century, putatively due to climate change and forest management activities ([45], [48]), which should take into account that pure and even-aged stands with long rotation cycle are more vulnerable to storm damage compared to mixed and uneven-aged stands ([78]).
One of the most recent and striking examples is the Vaia storm which hit the North-East of Italy destroying approximately 42.500 ha of forests, with damage still visible today ([82]). Visible damage induced by windstorms can range from uprooting or the breakage of a single tree to massive catastrophic losses of thousands of hectares of forest ([35]). It has been reported that a microburst characterized by strong wind speeds (more than 100 km h-1) and violent shower (more than 50 mm h-1) uprooted most of the holm oak trees present in forests close to urbanized metropolitan areas of Naples (Southern Italy - [9]) .
In general, tree susceptibility to storm damage depends on tree height and rooting habit ([55], [48]). During such windstorms, tree damage is mediated by the physical resistance of trees to the lever arm exerted by the wind on the tree crowns ([35], [37]). Thus, the social status of trees within a stand indirectly influences their susceptibility to wind damage. Further, the presence of high stem density, abundant standing dead elements and weakened plants (i.e., due to biotic or abiotic factors), increase stand risk to suffer a “domino effect” ([19]). The domino effect occurs when falling trunks damage adjacent trees, resulting in higher proportions of uprooted, broken, bent, and inclined trees. Although windstorms can cause a reduction in NDVI due to fallen trees within a forest, an increase in radial growth of the surviving trees associated to a relief of competition for light and soil water can be expected, as shown by Camarero et al. ([17]). Interestingly, the authors showed that windstorm, or more appropriately the canopy gap generated by a windstorm, can induce also important changes in soil conditions towards a nutrient poor soil with fewer fungi-dominated communities, especially in soils with a low clay content ([17]). Additionally, previous studies have shown that storm damages can favour the spread of invasive plant species and modify soil microbial community composition, resulting in an increase in plant pathogens ([49], [74]).
Physiological responses to windstorm
The most destructive impact of windstorms on forests occurs when wind loads exceed the resistance of stem or root/soil systems, resulting in tree breakage and uprooting ([37]). However, Mediterranean species, characterised by a high resprout capacity, should be more resilient than those of other biomes to windstorm damage.
Wind exposure can induce peculiar morphological responses (i.e., thigmomorphogenetic responses), including the formation of flexure wood, increased secondary thickening of stems and structural roots, and reduced shoot extension ([36]). These growth responses produce trees with an higher resistance to deflection and firm root anchorage. In addition, airflow across the leaf surface removes the boundary layer of the leaves and induces stomatal closure; therefore, wind can influence the photosynthetic performance of plants. However, only a few studies - not dealing with Mediterranean plants - have addressed the impact of wind on leaf gas exchange and water relations ([23]). Previous research showed that wind can greatly affect leaf transportation compared to leaf photosynthesis because of a wind-induced stomatal limitation to photosynthesis without altering CO2 diffusion through the mesophyll ([43]). Additionally, depending on temperature and speed, wind may induce a reduction in leaf light interception, thus reducing plant photosynthetic carbon uptake or inducing leaf cuticular damage ([23]). Concerning the latter aspect, it is highly likely that minimum leaf conductance (gmin), i.e., the lowest leaf conductance reached when stomata are maximally closed, as well as the biochemical and morphoanatomical traits affecting gmin, play an important role in leaf water status under wind flow. Overall, the effect of wind on plant physiology remains under-investigated, probably because of the difficulties in artificially manipulating this stress factor; even if this were to be achieved, it would be difficult to artificially obtain windstorms. Wind tunnels provide a viable way to deepen the investigate of the impact of wind loads on trees under controlled conditions, though such kind of facilities are not common ([53]). However, these aspects should be further investigated, given the forecasts of cyclones and storms in Mediterranean areas.
Patterns of mortality observed after climatic extremes and forest monitoring
Patterns of mortality
Local factors such as stand structure, soil water holding capacity, nutrient availability can modulate tree sensitivity to climate significantly affecting tree mortality patterns ([39]). Mortality rates and patterns depend also on the composition and genetic structure of the forest stands, being homogeneous, monospecific forests more sensitive than the mixed ones.
Extensive cases of dieback and mortality (die-off) occur in consequence of catastrophic events, e.g., the aforementioned Vaia windstorm in the central-eastern Alps ([82]), large scale insect outbreaks or extreme heat and drought waves at regional scale. In north-eastern Spain the severe winter-spring drought in 2011-2012 induced Pinus sylvestris L. and Abies alba Mill die-off ([38]). Die-off has been reported also in a Mediterranean shrublands in southern Spain due to low precipitation and a severe cold winter in 2005 ([57]). Gazol et al. ([39]) stated that a high mortality rates at regional scales can be expected in Spanish gymnosperms forests due to their higher tree growth reduction under drought and their higher distribution in dry region compared with angiosperms. Extensive mortality is favoured in homogeneous environment, both in relation to the physical properties of the stands (soil, exposure etc.) and in the composition and genetic structure of the stand.
Patchy dieback and mortality can occur in heterogeneous environments, with local differences in site conditions (bedrock, soil thickness, slope, exposure) and forest structure and composition with patches of drought sensitive species or genotypes alternate with resistant ones. Drought stress have been reported to induce also other mortality pattern, i.e., patchy dieback. Galiano et al. ([32]) reported a patchy dieback pattern in a Quercus ilex stand in Northeastern Spain, revealing signs of carbon starvation in declining trees even seven years after a single drought event.
Moderate heat and drought stress can provoke dieback and mortality at individual level, so inducing the so-called “salt and pepper” pattern. Salt and pepper dieback pattern have been associated to drought episodes (e.g., 2012 and 2017), as in the case of Pinus nigra J.F. Arnold reported in Northeastern Italy from 2013 ([64]). The authors hypothesized that intraspecific differences in the hydraulic strategy may underlie the observed salt and pepper dieback pattern. Non-declining trees resulted to adopt a more conservative water-use strategy under drought stress compared non-declining trees ([64]). Similar results have been reported also for Quercus species. In Southern Italy, declining and non-declining Quercus frainetto Ten. trees within the same forest patch differed in terms of intrinsic water-use efficiency (WUEi), with declining trees showing lower WUEi compared with non-declining ([25]). Additionally, a patchy dieback has been documented in a Q. ilex forest in Central Italy, where close trees showed a strong divergence in intrinsic water use efficiency, as inferred from tree-ring δ13C associated to different genetic background based on simple-sequence repeat (SSR) markers ([2]).
The patterns of dieback and mortality may have different ecological consequences. Die-off and patchy patterns can trigger changes in the composition and structure of the forests, with the decline of the sensitive species. Conifers can be replaced by broadleaves ([46]), whereas the Mediterranean high forest of Quercus ilex may go towards a regressive dynamic from forests to maquis and garrigue ([11]). The “salt and pepper” pattern, although less showy, may have relevant consequences on the composition (biodiversity) and the genetic structure of the forests. Likely, mortality affects the more sensitive species and genotypes with a loss of biodiversity and simplification of the genetic structure of the stand.
Monitoring tree mortality in Mediterranean forests
The knowledge of the patterns, in addition to the intensity of mortality, is relevant for forest planning and management. Tree mortality is currently assessed by periodic terrestrial surveys on systematic networks within the national forest inventories (NFI) and the ICP Forests programme (⇒ https://www.icp-forests.net/). The latter network is aimed to monitor the general conditions of trees in terms of defoliation and damage symptoms ([12]). These surveys produce data concerning the percentage of dead and declining trees within a population but are not aimed at describing the spatial mortality patterns.
Remote sensing (RS) is a powerful and widespread approach to detect the extension and intensity of the climatic impacts, but its efficacy can be limited, and the results biased when applied in heterogeneous environment and complex forests in terms of structure and composition. The die-off is ideally the more easily detectable pattern using the Normalized Difference Vegetation Index (NDVI) through remote sensing techniques ([38]). In the years after the impact, however, the possible recovery of NDVI may be linked to the vegetational dynamics triggered by the stressful event (with the development of secondary successions) rather than the recovery of the canopies. Patchy and “salt and pepper” patterns can escape from RS detection and require high-resolution data supported by proximal sensing (drone) and field validation ([54]).
The combination of terrestrial and RS monitoring methods is therefore necessary for a more comprehensive picture, to follow the ongoing ecological transformations in terms of mortality in the understorey, substitution of species, etc.
Conclusions
Although being considered resistant to drought, Mediterranean ecosystems are fragile to current global climate changes. Mediterranean forests evolved under different climatic conditions, and show a very fragile ecological equilibrium. Indeed, they are characterized by patches of microhabitats and refugia rich of endemic species. Climate change is expected to exert a selective pressure leading to the loss of the less adapted genotypes and species. Changes can occur on large (die-off mortality pattern), medium scale (patchy mortality pattern) or on singular individual tree (“salt and pepper” pattern), depending on the intensity and frequency of the climatic impacts, the physical characteristics of the sites, and the structure and composition of forests. Among the abovemetioned forest dieback patterns, salt and pepper dieback is not easily to detect by satellite images with low spatial resolution (10 m2), which could not be enough to detect the presence of healthy and declining neighbor trees. Annual field monitoring campaigns could be recommended to further evaluate the temporal worsening of this mortality pattern into patchy decay.
Mediterranean forests are likely going toward more simplified structures with prevalence to shrub formations and garigue. This process, together with the increase of wildfire, may led to erosion and desertification processes. All these aspects will have impact, not only at biological, ecosystem and environmental levels, but also on the landscape level, hindering the touristic fruition of the Mediterranean regions. It is therefore necessary to think about new ecological equilibriums and concepts for a bio-economy sustainable in the new ecological conditions.
Acknowledgements
This paper is part of the project “LIFE MODERn(NEC) - new monitoring system to detect the effects of reduced emissions of pollutants resulting from the adoption of the NEC Directive” - LIFE20 GIE/IT/000091.
The authors are grateful to Maremma Regional Park to allow the access to the park in the frame of the project “Impatto dei cambiamenti climatici sulla vegetazione di ecosistemi costieri mediterranei”.
Funds from the National Biodiversity Future Center (NBFC) to Martina Pollastrini and Francesco Ferrini are acknowledged.
References
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Gscholar
CrossRef | Gscholar
Supplementary Material
Authors’ Info
Authors’ Affiliation
Cecilia Brunetti 0000-0002-8531-6076
Antonella Gori 0000-0002-7304-7526
National Research Council of Italy - CNR, Institute for Sustainable Plant Protection, Sesto Fiorentino, I-50019 Florence (Italy)
Filippo Bussotti 0000-0002-8353-4459
Cecilia Brunetti 0000-0002-8531-6076
Francesco Ferrini 0000-0003-2222-0437
Antonella Gori 0000-0002-7304-7526
Martina Pollastrini 0000-0003-0959-9489
University of Florence, Department of Agriculture, Food, Environment and Forestry, I-50044 Florence (Italy)
NBFC, National Biodiversity Future Center, I-90133 Palermo (Italy)
Corresponding author
Paper Info
Citation
Alderotti F, Bussotti F, Brunetti C, Ferrini F, Gori A, Pollastrini M (2024). Linking patterns of forest dieback to triggering climatic and weather events: an overview on Mediterranean forests. iForest 17: 309-316. - doi: 10.3832/ifor4545-017
Academic Editor
Marco Borghetti
Paper history
Received: Dec 21, 2023
Accepted: Sep 17, 2024
First online: Sep 30, 2024
Publication Date: Oct 31, 2024
Publication Time: 0.43 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2024
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 15413
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 8956
Abstract Page Views: 3944
PDF Downloads: 2089
Citation/Reference Downloads: 8
XML Downloads: 416
Web Metrics
Days since publication: 669
Overall contacts: 15413
Avg. contacts per week: 161.27
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Jul 2026)
Total number of cites (since 2024): 11
Average cites per year: 3.67
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Review Papers
Impacts of climate change on the establishment, distribution, growth and mortality of Swiss stone pine (Pinus cembra L.)
vol. 3, pp. 82-85 (online: 15 July 2010)
Review Papers
Drought-induced mortality of Scots pines at the southern limits of its distribution in Europe: causes and consequences
vol. 3, pp. 95-97 (online: 15 July 2010)
Research Articles
Is Pinus pinea growth affected by climate change in western Anatolia?
vol. 18, pp. 93-101 (online: 28 April 2025)
Review Papers
SilvaCuore: a web-application to monitor the health status of Italian forests
vol. 17, pp. 317-322 (online: 30 September 2024)
Research Articles
Effects of warmer and drier climate conditions on plant composition and biomass production in a Mediterranean shrubland community
vol. 1, pp. 39-48 (online: 28 February 2008)
Research Articles
Predicting the effect of climate change on tree species abundance and distribution at a regional scale
vol. 1, pp. 132-139 (online: 27 August 2008)
Research Articles
Perceptions of forest experts on climate change and fire management in European Mediterranean forests
vol. 7, pp. 33-41 (online: 14 October 2013)
Research Articles
Seeing, believing, acting: climate change attitudes and adaptation of Hungarian forest managers
vol. 15, pp. 509-518 (online: 14 December 2022)
Research Articles
Potential impacts of regional climate change on site productivity of Larix olgensis plantations in northeast China
vol. 8, pp. 642-651 (online: 02 March 2015)
Review Papers
Climate change impacts on spatial distribution, tree-ring growth, and water use of stone pine (Pinus pinea L.) forests in the Mediterranean region and silvicultural practices to limit those impacts
vol. 14, pp. 104-112 (online: 01 March 2021)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword






