
Extreme climatic events, biotic interactions and species-specific responses drive tree crown defoliation and mortality in Italian forests
iForest - Biogeosciences and Forestry, Volume 17, Issue 5, Pages 300-308 (2024)
doi: https://doi.org/10.3832/ifor4531-017
Published: Sep 30, 2024 - Copyright © 2024 SISEF
Research Articles
Collection/Special Issue: Project LIFE MODERn(NEC)
Workshop "Climate change and forest health monitoring in Italy" - Rome (Italy) May 5, 2023
Guest Editors: Bussotti F, Pollastrini M
Abstract
The frequency of forest disturbances has increased in recent years, provoking widespread defoliation, crown dieback and tree mortality. The ICP Forests monitoring network offers a unique platform for observing the impacts on forests of heatwaves, droughts and other extreme climatic events, as well as the trends of defoliation and mortality. The Italian ICP Forests Level I network consists of 261 permanent plots where tree crown defoliation and damage symptoms are assessed visually each year by well-trained crews of the Corpo Forestale dello Stato (2001-2016) and the Carabinieri Forestale from 2017 onward. This paper aims to assess the main tree species’ responses, in terms of defoliation and mortality, to severe climatic events. The results are discussed in relation to species-specific physiological behaviour and bioclimatic regions. A significant trend toward increasing defoliation and mortality has been observed since 2010 in both conifers and broadleaves. Conifers (especially Picea abies), which are largely diffuse in the Alpine regions, have suffered from bark beetle outbreaks due to severe windstorms (such as Vaia in 2018) and recurrent dry years. In the temperate regions, characterised by deciduous broadleaved trees, the most relevant defoliation events coincided with the driest and hottest years, with low relative humidity (2012, 2017 and 2021-2022), only partially recovering in the subsequent years. Among them, Fagus sylvatica and Quercus cerris, along with increased defoliation, showed symptoms caused by fungi of the genus Biscogniauxia, causal agents of “charcoal canker”, in less favourable site conditions. Quercus pubescens was the most resilient species, able to restore its crown after defoliation. The Mediterranean forests, with evergreen broadleaved species, showed no significant trends but were impacted at the most drought-prone coastal sites. The findings evidenced that the current ICP Forests network in Italy represents a fundamental infrastructure for monitoring impacts and trends connected to climate change and species-specific responses. A local intensification of the grid would help to capture under-represented species or ecological conditions.
Keywords
Climate Change, Crown Defoliation, Emerging Forest Diseases, Forest Health Monitoring, Heatwaves and Droughts, ICP Forests, Tree Ecophysiology, Tree Mortality
Introduction
Forest disturbances (“Damage caused by any factor - biotic or abiotic - that adversely affects the vigor and productivity of the forest, and which is not a direct result of human activities” - [23]) have increased in recent years, as related to global climate changes ([47], [48], [41]). They consist mainly of wind damage, pathogen and insect outbreaks, fires, extreme heatwaves and droughts, invasion of alien pathogenic species, and emerging diseases, and are related to each other. For example, drought increases the occurrence of insect outbreaks in coniferous forests and wound decay fungi in both broadleaf and conifer stands ([2]). The impacts of disturbances on forests are assessed in Europe within the ICP Forests programme (⇒ http://icp-forests.net/). The most widely used parameter to assess the condition of trees is the defoliation of the crown (see Appendix 1 in Supplementary material). Defoliation is defined as needle or leaf loss in the crown as compared to a reference tree ([21]). Defoliation is an unspecific indicator, integrating the intrinsic genetic constitution of trees and site effects ([33]), and can be considered indicative of a tree’s equilibrium in its own environment, representing the result of the accumulated impacts of stressful environmental conditions. External fluctuating pressures, such as abiotic and biotic factors, cause year-to-year variations in defoliation (ΔDef), which are the true indicator of the impacts ([40]). Although the ICP Forests programme was launched to assess the effects of air pollution on forests, the relationship between air pollution and crown condition appears to be weak and limited in time and space, while climatic factors are the most important drivers (De Vries et al. ([20]). In the context of climate change, severe heatwaves and droughts, with decreasing relative humidity, are expected to be more intense and frequent in the years to come ([50], [31]). The challenges imposed by new environmental conditions increase the interest in forest health monitoring among the scientific community and political decision-makers ([24], [8]).
The ICP Forests programme includes two kinds of monitoring networks: Level I (extensive monitoring), to assess the extension at the European level of defoliation and damage, and Level II (intensive monitoring), where cause-effect studies are performed (see Appendix 2 in Supplementary material). The results of extensive and intensive surveys across Europe are reported in annual technical reports (⇒ http://icp-forests.net/page/icp-forests-technical-report) and briefs (⇒ http://icp-forests.net/page/icp-forests-briefs). These documents indicate a general and significant trend of increasing defoliation since 2010 ([43]). The elaboration of the ICP Forests data (Level I) revealed increasing tree mortality rates at the European level ([27], [28]).
Italy can be considered a relevant case study because it includes three bioclimatic regions (alpine, temperate and Mediterranean - [4]). These regions have forests with different tree species compositions. The alpine region is characterised by conifers such as Picea abies (L.) Karst., Larix decidua L. and Pinus sylvestris L.; the temperate region has a prevalent presence of deciduous broadleaved species, both mesophile (Fagus sylvatica L., Castanea sativa Mill.) and thermophile (Quercus pubescens Willd., Quercus cerris L., Ostrya carpinifolia Scop.); the Mediterranean region is characterised by evergreen broadleaved species, with a prevalence of Quercus ilex L. Only Fagus sylvatica, which is widespread on a continental scale, is represented in both the alpine and temperate regions, and it reaches in Italy the southernmost limits of its distribution. For these reasons, Italian forests offer the chance to study the patterns (triggering events, interactions with biotic agents, species-specific and site-specific responses) of climate change impacts and the possible recovery of tree species under different bioclimatic conditions, as well as the possible different behaviour of co-occurring species in the same region ([10]). The progressive shifting of climatic factors, with increasing temperature and air dryness, and recurrent climatic anomalies have impacted Italian forests both in the alpine ([53], [18]) and in the temperate and Mediterranean ([14], [17], [42], [46]) regions. In many cases, climate-stressed trees have been subjected to pathogens, which constituted the ultimate cause of tree death.
This paper presents the results of a multiyear (2001-2022) assessment of the Italian plots belonging to the ICP Forests Level I network. The trends over time are analysed for all the sampled tree species, functional groups (conifers and broadleaves) and the main singular tree species. The objectives are:
- the European ICP Forests Level I network is designed with a low-intensity grid, with the purpose of producing data on a continental scale. Its representativeness at the national level may be questioned; the first goal is to verify if the current Italian monitoring system can capture the general impacts of extreme climatic events and trends of defoliation and mortality at the national level;
- the levels of defoliation fluctuate with year-to-year changes (ΔDef) in relation to occasional disturbances, although the long-term trend can be expected to be constant and represented by a linear regression; permanent changes in environmental conditions, on the other hand, can modify this trend; a severe event (climatic or biotic) can represent a key year; after that, the linear regression of the trend may assume a different slope or intercept ([30]). Our goal is to determine the occurrence of possible key years (and impact events) that significantly affected the patterns of defoliation for groups or singular species.
- The bioclimatic regions are subjected to different kinds of climatic and biotic impacts. In the last few decades, the temperate and Mediterranean regions have been subjected to extreme heat and drought events ([42], [46]), whereas alpine coniferous forests have been impacted by severe windstorms ([18]); we expect to find in each region different key years and triggering events, as well as specific patterns of defoliation and mortality;
- although defoliation is considered a common and nonspecific symptom, the patterns of defoliation, mortality, and recovery can be expected to differ among species co-occurring in the same bioclimatic region; such differences are likely related to different ecophysiological strategies to cope with extreme climatic events and pathogens; from this perspective, the observed trends will be discussed considering the available literature.
Materials and methods
Size and distribution of the sample
The dataset consists of the defoliation data from the Italian ICP Forests Level I network and includes information from 261 permanent monitoring plots (distributed across the country with a 15 x 18 km grid - Fig. S1 in Supplementary material), assessed annually in the period 2001-2022. The plots are circular areas (13 m radius) in which all dominant trees with a diameter at breast height >9.5 cm were assessed. The mean number of trees, in total and for the main species, is shown in Tab. 1. More than 60 species were identified and assessed; however, 85% of the sample trees are represented by the 10 main species listed in Tab. 1.
Tab. 1 - Mean number of trees assessed (2001-2022) in total, grouped in broadleaves and conifers, and the main species.
| Species | N |
|---|---|
| All Trees | 5966 |
| Broadleaves | 4330 |
| Conifers | 1639 |
| Fagus sylvatica | 1104 |
| Quercus pubescens | 694 |
| Quercus cerris | 563 |
| Picea abies | 550 |
| Castanea sativa | 441 |
| Ostrya carpinifolia | 348 |
| Larix decidua | 339 |
| Quercus ilex | 223 |
| Pinus sylvestris | 206 |
| Pinus nigra | 189 |
Assessment of defoliation
The assessment of defoliation is carried out each year by the personnel of the Corpo Forestale dello Stato (until 2016) and the Carabinieri Forestale (from 2017 onward) according to the standardised methods formalised in the European ICP Forests common protocols ([21]) and translated into the national manual ([11]). The defoliation of the visible upper part of the crown is evaluated visually by field crews according to a proportion scale in 5% intervals (0 = no defoliation; 5%; 10%; 15%; …; 100% = dead tree - 99% indicates a tree completely defoliated but not dead) by comparing the crowns of the sampled trees with photographic standards of reference trees for the main forest species, i.e., the photo guide method proposed by Müller & Stierlin ([36]) for alpine conifers and mesophile broadleaves and for Mediterranean evergreen species and thermophile broadleaves. The evaluation includes an assessment of damage symptoms and causal agents. Dead trees (standing, fallen and uprooted) were assessed only in the year in which their death occurred. Yearly surveys are carried out during the summer months, when leaves are fully expanded and before autumnal senescence (the recommended period ranges from mid-July to mid-September). Field crews are trained before each yearly assessment to guarantee the scores’ comparability and repeatability ([9]).
Statistics
The significance of the trends of defoliation and mortality (all species, conifers and broadleaves, and the main singular species) over the 2001-2022 period was calculated according to the non-parametric Mann-Kendall trend test (R Package “Trend”). Defoliation data of living trees are presented as the average percentage of trees over specific thresholds: >25% (all defoliated trees); >60% (severe defoliation), and >85% (extreme defoliation). Data on dead trees are reported separately. Structural changes in chronologies were tested with the Chow test ([13], [55]), a statistical test that allows one to confront two regression lines with different slopes or intercepts and identify the presence of two different processes underlying time series’ generation ([30]). The Chow test was applied to determine the key year for increasing defoliation. Significance is reported as p<0.05 (significant), p<0.01 (very significant) and p<0.001 (highly significant).
Results and discussion
Climatic trends and extreme events
Temperatures are increasing in Italy, with a very marked trend since the 1980s, and this increase is driven mainly by the summer temperature ([6]). From 1981 to 2019, the increasing rates recorded for the summer temperatures were 1.65, 1.77 and 2.00 °C, respectively, for Northern, Central and Southern Italy. The highest increase in the mean temperature on a monthly basis was registered in the Alps for June (about 3 °C in a 38-year period - [6]). In the last few decades, heatwaves originating in North African regions have been reaching the peninsular areas of Italy more and more often, bringing torrid temperatures already in early June ([6]).
Climatic anomalies in Italy from 1990 to 2022 are presented in Fig. S2 (Supplementary material - [49]). The anomalies are expressed as the percentage deviation compared to the 30-year average. The temperature anomalies show a significantly positive trend (p<0.001 - Fig. S2A in Supplementary material), whereas relative humidity shows a significantly negative trend (p<0.001 - Fig. S2B in Supplementary material). Heatwaves and droughts were recorded in 2003 ([45]), 2012 ([34]) and 2017 ([46]). The year 2022 was the hottest of the 30-year period, with an anomaly of +1.23 °C on a yearly level and +2.18 °C in the summer. Precipitation shows no significant trends (Fig. S2C in Supplementary material). We can recognise, however, a series of dry years in the 1990s (with negative anomalies for temperature) and some sparse severe drought events. Other extreme climatic events were the late frost in the central Apennine in spring 2016 ([17]) and the windstorm Vaia ([18]) that hit the forests of the North-Eastern Alpine Belt in October 2018.
General trends of defoliation and mortality
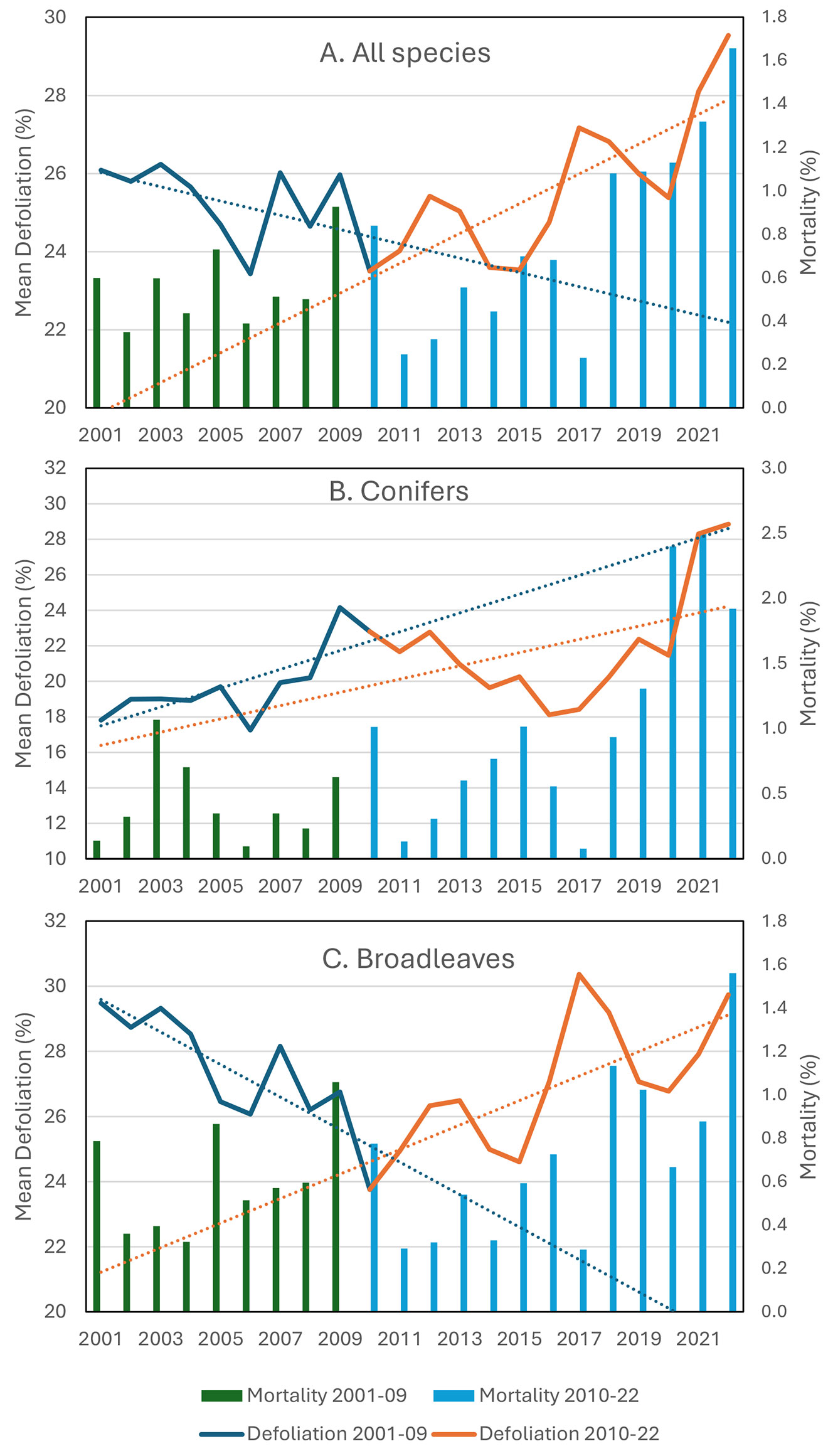
The general results (all species, 2001-2022 period - Fig. 1A, Tab. 2) show no significant trends in the mean defoliation for the whole 2001-2022 period, but there is a significant increase in the percentage of trees in the highest defoliation classes (ΔDef >60%) and increasing tree mortality. The increase in mean defoliation became significant in the second sub-period (2010-2022 - Tab. 2).
Fig. 1 - Mean defoliation (on living trees) and mortality (A: All trees; B: Conifers; C: Broadleaves) during the 2001-2022 period. The subperiods 2001-2009 and 2010-2022 are indicated with different colors. Significance of trends are reported in Tab. 2.
Tab. 2 - Significance of the trends of defoliation (living trees) and mortality in the whole 2001-2022 period, and in the 2 sub-periods (2001-2009 and 2010-2022), according to the Mann-Kendall test. Data in brackets indicate negative trends (decreasing defoliation). (ns): not significant.
| Species | Period | Mean Def. | >25% | >60% | >85% | Mortality |
|---|---|---|---|---|---|---|
| All species | 2001-2022 | ns | ns | p<0.05 | p<0.01 | p<0.05 |
| 2001-2009 | ns | (p<0.10) | p<0.01 | p<0.01 | ns | |
| 2010-2022 | p<0.001 | p<0.01 | p<0.05 | p<0.01 | p<0.01 | |
| Conifers | 2001-2022 | p<0.01 | p<0.01 | p<0.05 | p<0.05 | p<0.05 |
| 2001-2009 | p<0.05 | p<0.05 | p<0.1 | ns | ns | |
| 2010-2022 | ns | ns | p<0.05 | p<0.01 | p<0.01 | |
| Broadleaves | 2001-2022 | ns | ns | ns | p<0.05 | ns |
| 2001-2009 | p<0.05 | p<0.05 | p<0.05 | p<0.05 | ns | |
| 2010-2022 | p<0.05 | p<0.05 | n | p<0.05 | p<0.01 | |
| Picea abies | 2001-2022 | p<0.05 | p<0.01 | p<0.001 | ns | p<0.001 |
| 2001-2009 | p<0.01 | p<0.05 | p<0.05 | p<0.05 | ns | |
| 2010-2022 | ns | ns | ns | ns | p<0.05 | |
| Larix decidua | 2001-2022 | ns | ns | ns | ns | ns |
| 2001-2009 | ns | p<0.05 | ns | ns | p<0.05 | |
| 2010-2022 | p<0.05 | p<0.05 | ns | ns | p<0.01 | |
| Pinus sylvestris | 2001-2022 | p<0.01 | p<0.05 | ns | ns | ns |
| 2001-2009 | ns | ns | ns | ns | ns | |
| 2010-2022 | ns | ns | ns | ns | p<0.05 | |
| Pinus nigra | 2001-2022 | p<0.05 | ns | (p<0.001) | ns | ns |
| 2001-2009 | ns | ns | ns | ns | ns | |
| 2010-2022 | p<0.001 | p<0.001 | p<0.01 | p<0.05 | ns | |
| Fagus sylvatica | 2001-2022 | ns | ns | ns | ns | ns |
| 2001-2009 | p<0.05 | p<0.05 | ns | p<0.01 | ns | |
| 2010-2022 | p<0.05 | p<0.05 | ns | ns | ns | |
| Quercus pubescens | 2001-2022 | (p<0.001) | (p<0.001) | (p<0.05) | ns | ns |
| 2001-2009 | p<0.05 | p<0.05 | ns | ns | ns | |
| 2010-2022 | ns | ns | ns | ns | ns | |
| Quercus cerris | 2001-2022 | ns | ns | ns | ns | p<0.05 |
| 2001-2009 | (p<0.01) | (p<0.01) | ns | p<0.05 | ns | |
| 2010-2022 | p<0.01 | p<0.05 | p<0.1 | ns | p<0.01 | |
| Quercus ilex | 2001-2022 | ns | ns | ns | ns | ns |
| 2001-2009 | ns | ns | ns | ns | ns | |
| 2010-2022 | ns | ns | ns | ns | ns | |
| Castanea sativa | 2001-2022 | (p<0.001) | p<0.05 | p<0.01 | p<0.01 | p<0.01 |
| 2001-2009 | p<0.05 | ns | p<0.01 | p<0.05 | ns | |
| 2010-2022 | ns | ns | ns | ns | p<0.01 | |
| Ostrya carpinifolia | 2001-2022 | ns | ns | ns | ns | ns |
| 2001-2009 | ns | ns | ns | ns | ns | |
| 2010-2022 | p<0.01 | p<0.001 | p<0.1 | ns | p<0.05 |
Mean defoliation increased significantly in conifers (Fig. 1B, Tab. 2) during the whole study period (2001-2022), but not in broadleaves (Fig. 1C, Tab. 2). Mean defoliation and mortality in broadleaves increased significantly in the second sub-period (2010-2022), especially in the highest defoliation classes (>85%-99%).
Conifers suffer from recurrent biotic and abiotic disturbances common in the Alpine region ([29]); however, the dramatic recent increase in defoliation and mortality was attributed to bark beetle outbreaks, which were widespread after the catastrophic storm Vaia (2018), increasing temperatures, and recurrent dry years, thus mirroring what was registered at the European level ([32]). Broadleaves show defoliation peaks corresponding to the severe drought events of 2012, 2017 and 2021-2022 (Fig. 1C) that affected the temperate and Mediterranean regions, especially Central and Southern Italy, as well as the late frost of 2016 in central Apennine beechwoods ([17]).
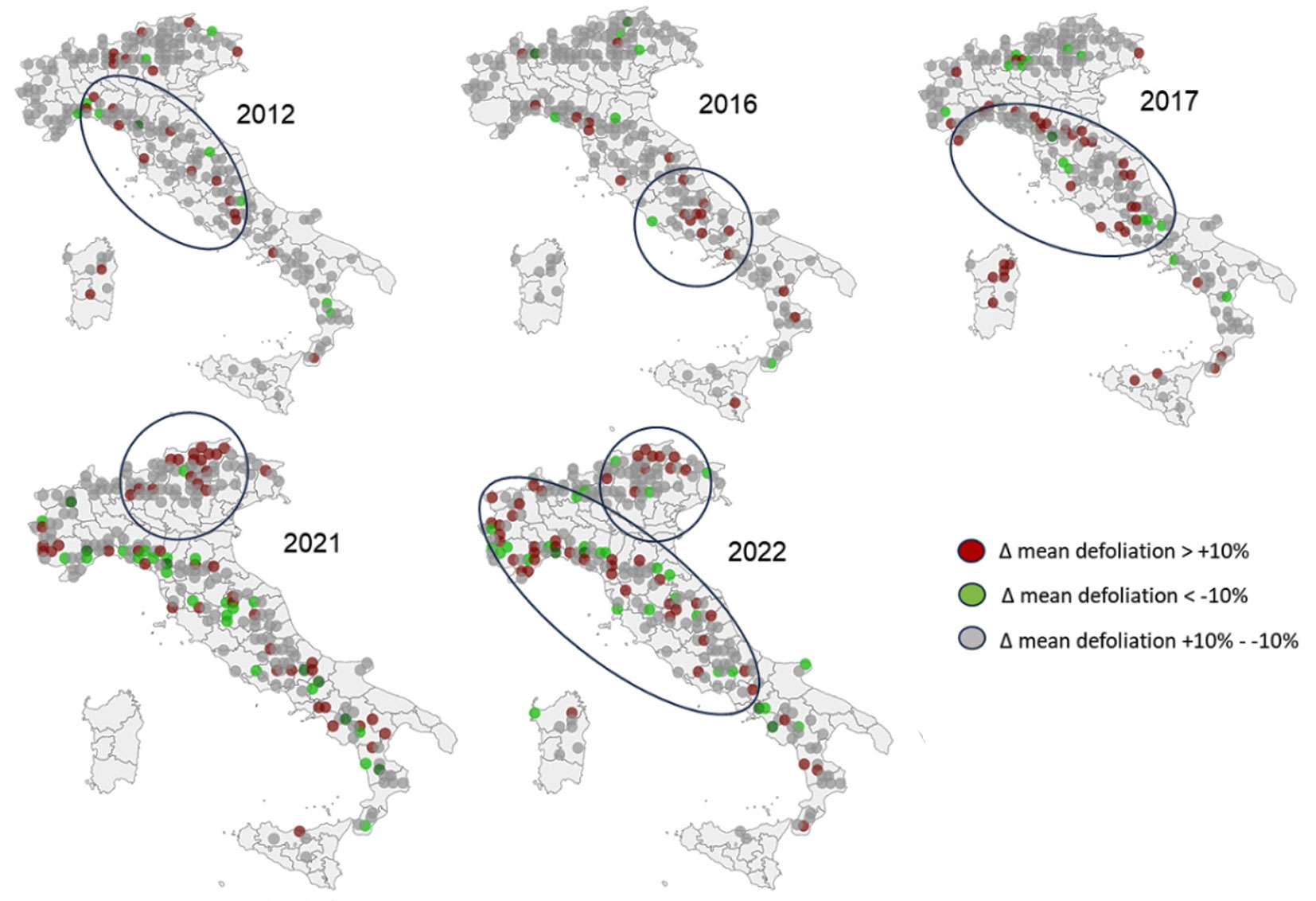
The plots with the highest year-to-year changes in mean defoliation are shown in Fig. 2. The central Apennine and North-Western regions were the most affected by the 2012, 2016 (late frost) and 2017 events. The higher defoliation in 2021 was driven by the coniferous forests in the North-Eastern Alpine Belt. The increasing defoliation of trees in 2022 shows two different components, in the Alpine Belt and the Central and North-Western regions.
Fig. 2 - Maps of the changes in defoliation at the Italian Level I plots, with respect to the previous year. Brown colored plots indicate an increase of the mean defoliation (> +10%); green colored plots indicate a reduction of defoliation (< -10%). Grey plots indicate a change of defoliation between +10% and -10%.
Drought events in the first sub-period (2001-2009) were not accompanied by high mortality. There is a substantial difference between the past and current drought events: according to Gazol & Camarero ([27]), recent events can be classified as “composite droughts”, characterised by higher temperatures and vapour pressure deficit (VPD).
Trends of defoliation and mortality on coniferous species
Defoliation and mortality in Picea abies showed a positive trend during the whole assessment period (2001-2022) and increased dramatically starting in 2018 (Fig. 3A, Tab. 2). This species suffered from one of the most aggressive bark beetles, Ips typographus (L.). Attacks were stronger in the years after the storm Vaia (2018), since the large amount of deadwood produced represented an ideal habitat and substrate for this insect’s growth and reproduction. The increasing frequency and severity of dry conditions were also contributing causes of bark beetle damage ([32]).
Fig. 3 - Mean defoliation (on living trees) and mortality on conifer species. (A: Picea abies; B: Larix decidua; C: Pinus sylvestris; D: Pinus nigra). The subperiods 2001-2009 and 2010-2022 are indicated with different colors. Significance of trends are reported in Tab. 2.
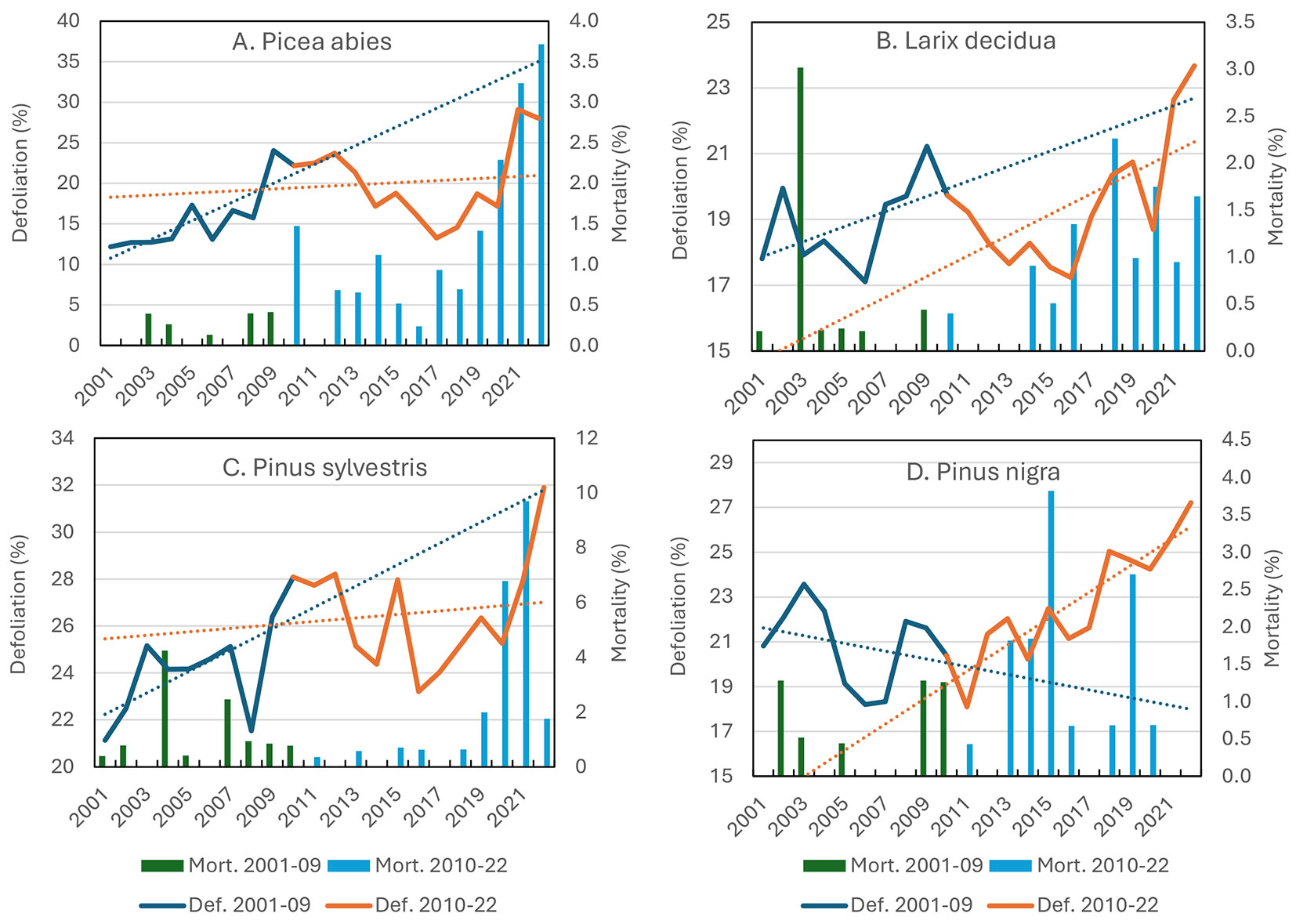
Larix decidua shows significant trends of increasing defoliation and mortality rates in the second sub-period (2001-2022 - Fig. 3B, Tab. 2).
Pinus sylvestris showed the highest mortality rates (Fig. 3C, Tab. 2). This species’ decline began in the first years of the 21st century on the southern slopes of the Alps because of climate warming and subsequent bark beetle attacks ([15], [53]).
Pinus nigra Arn., largely used for afforestation programmes, may be subject to early senescence caused by its ecological incompatibility with the conditions in which it was planted, aggravated by increasing drought that favoured the emergence of the endophyte fungal pathogen Sphaeropsis sapinea (Fr.) Dyko & Sutton [Syn.: Diplodia pinea (Desm.) Kickx] ([7] - Fig. 3D, Tab. 2).
Trends of defoliation and mortality in deciduous broadleaves
Although the trends of defoliation were not significant for the entire monitoring period (Tab. 2), Fagus sylvatica suffered leaf loss under climatic disturbances like the late frost in the central Apennine (2016) and the heatwave and drought in 2017. Subsequent pulses of heat and drought significantly induced positive defoliation trends in the second sub-period (Fig. 4A, Tab. 2). Fagus sylvatica demonstrated a sensitivity to cavitation ([54]), which is one of the most likely causes of leaf shedding, branch desiccation and tree death under extreme drought and may have consequences for tree water transport in the years after drought ([3]). No significant trends of mortality were observed in Fagus sylvatica, although extensive charcoal canker symptoms caused by Biscogniauxia nummularia (Bull.) Kuntze, a common endophyte that changes into a pathogen in stressed trees ([35], [37]), were observed in drought-prone sites and at lower elevations ([10]).
Fig. 4 - Mean defoliation (on living trees) and mortality on the main broadleaved species (A: Fagus sylvatica; B: Quercus pubescens; C: Quercus cerris; D: Quercus ilex; E: Castanea sativa; F: Ostrya carpinifolia). The subperiods 2001-2009 and 2010-2022 are indicated with different colors. Significance of trends are reported in Tab. 2.
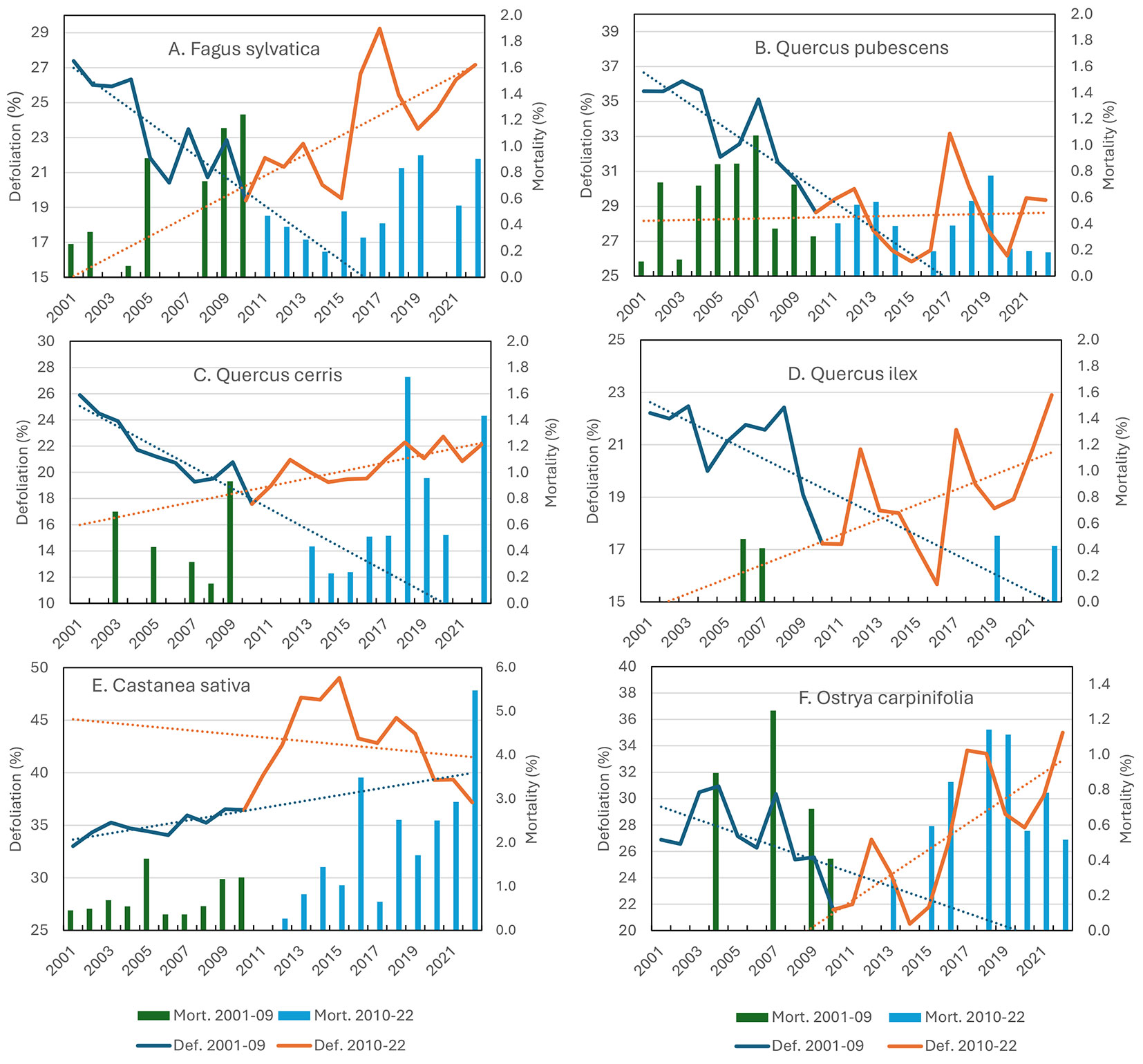
Quercus pubescens showed high levels of defoliation over time, although with different behaviours between the first (significant decrease in defoliation) and the second sub-period (no trend - Tab. 2, Fig. 4B). The levels of defoliation in this species present very strong year-to-year differences, but the mortality rates are constant throughout the whole 2001-2022 period. Quercus pubescens lost its leaves in the hottest and dryest summers but showed a substantial recovery in the following years, demonstrating high resilience. According to Nardini & Pitt ([39]), Quercus pubescens is very drought tolerant and can maintain high foliar relative water content under water stress conditions. This species compensates for the foliar loss of water with an equal amount of water uptake. Such a drought avoidance strategy is made possible by the high hydraulic efficiency of its stems and roots under water stress ([39]). Quercus pubescens has high water potential despite high stomatal conductance, indicating that its leaf water status is independent of stomatal activity ([25]). According to Gallé et al. ([26]) and Damesin & Rambal ([19]), a network of photoprotective mechanisms can preserve the potential functionality of the photosynthetic apparatus during severe drought, leading to a rapid recovery of photosynthetic activity after rewatering. Thus, Quercus pubescens seems to be capable of withstanding and surviving extreme drought events. Summer foliar abscission can be considered an active mechanism to avoid damage to the water conduction system.
Quercus cerris shows different behaviour than Quercus pubescens. After a significant negative trend (reduction of defoliation) in the first sub-period, defoliation increased significantly in the second sub-period (Tab. 2, Fig. 4C). Quercus cerris shows high mortality rates, with a significant positive trend. Defoliation and mortality are often delayed with respect to climatic impacts. Although it is described as a drought-tolerant species, the understanding of this species’ behaviour under extreme drought conditions is still limited. According to Nardini et al. ([38]), Quercus cerris has restricted water absorption compared to other Mediterranean oak species. Compared with Quercus pubescens, Quercus cerris has a less conservative water-use strategy, and it has higher photosynthetic rates than Quercus pubescens ([52]) but exhibits a steep decline in photosynthetic efficiency and leaf hydration under drought ([16]). Quercus cerris stems are strongly hydrated, and water storage reservoirs could play an important role in tree survival during extended periods of low soil water availability ([51]). Field evidence ([10]) revealed that mortality is often associated with attacks of the fungal endophyte and opportunistic pathogen Biscogniauxia mediterranea (De Not.) Kuntze ([12], [37]).
In Castanea sativa Mill., the trends of defoliation and mortality are significantly positive during the monitoring period (Fig. 4E, Tab. 2) and seem to be related to the attacks from the Asian chestnut gall wasp Dryocosmus kuriphilus Yasumatsu, which lead to the production of galls on growing shoots and leaves ([5]). The recent decrease in defoliation was attributed to the successful introduction of its natural antagonist, Torymus synensis Kamijo ([44]). Castanea sativa growing at lower elevations and in marginal areas was also affected by leaf shedding ([10]).
Ostrya carpinifolia showed high sensitivity to drought events, showing a positive trend of defoliation in the second sub-period (Tab. 2). Peaks of defoliation were observed in the driest years 2012, 2017 and 2021-22 (Fig. 4F). Peaks of defoliation were followed by substantial but incomplete recovery in the subsequent years. Highly defoliated trees often have abundant fructification. The sensitivity of this species to drought can be attributed to its shallow root system.
Trends of defoliation and mortality in evergreen broadleaves
Quercus ilex shows no significant trends of defoliation (Tab. 2, Fig. 4D). Field evidence ([42], [1], [10]) reveals the presence of strong impacts of extreme heat and drought events in this species at several coastal sites in Central Italy, with frequent tree mortality and crown dieback. Sclerophyll species skilfully manage water stress recovery during summer droughts. The large drop in stem water potential and cavitation strain during the day is followed by nighttime recovery thanks to the high air humidity condensing on the soil surface at night ([51]). Prolonged climate stress and high VPD decrease the possibility of recovery after a drought and might predispose these species to physiological decline and dieback because their hydraulic safety limits can be surpassed by frequent and repeated stress.
Comparison of trends
Different defoliation patterns for Alpine conifers and broadleaves (both deciduous and evergreen species) were revealed by the application of the Chow test (Tab. 3). The year 2010 was a key year for broadleaves, since significant changes in the trend directions (from negative to positive trends of defoliation) were observed after this year. The change in the direction of the trends is attributed to subsequent heat and drought events in the temperate and Mediterranean regions. In Castanea sativa, the key year corresponds to the outbreak of Dryocosmus kuriphilus. Fagus sylvatica shows different patterns of defoliation in the bioclimatic regions where this species grows (alpine and temperate), with non-significant trends in the Alpine regions and a significant key year (p<0.001) of 2010 in the temperate regions (Fig. S3 in Supplementary material). Conifers in the Alpine regions were not subjected to the same disturbances as broadleaves, and a possible key year can be individuated in 2018, with the windstorm Vaia and subsequent bark beetle attacks (the time series, however, is still too short to identify significant trends). Pinus nigra, which is often planted in Central Italy, was subjected to the same disturbances as the temperate region and shows the key year of 2010.
Tab. 3 - Results of the Chow test. The year 2010 was tested to check the significance of the change of direction in the regression of the series of data.
| Species | p-value |
|---|---|
| All | ns |
| Broadleaves | p<0.001 |
| Conifers | ns |
| Picea abies | ns |
| Larix decidua | ns |
| Pinus sylvestris | ns |
| Pinus nigra | p<0.01 |
| Fagus sylvatica | p<0.01 |
| Quercus pubescens | p<0.05 |
| Quercus cerris | p<0.001 |
| Quercus ilex | p<0.01 |
| Castanea sativa | p<0.05 |
| Ostrya carpinifolia | p<0.01 |
Conclusions
The behaviour of defoliation and mortality at the national level is the product of different species-specific physiological and pathological patterns and the distinct species composition in the bioclimatic regions. These results do not therefore have a unique ecological or physiological meaning but permit the comparison of different national trends at the European level in the ICP Forests reports. The results of a 22-year monitoring period in Italy suggest the role of severe heatwaves and droughts and other extreme events (windstorms, late frost) in increasing tree defoliation and mortality. These events constitute a point of rupture and discontinuity in the temporal trends of defoliation and mortality. Such events, however, have different natures, intensities and frequencies in the different bioclimatic regions. Alpine coniferous forests are the most fragile formations, with a quick decline in recent years connected to extreme windstorms followed by insect outbreaks. Such events, in our opinion, are not sufficient to explain these species’ decline (the decline of Pinus sylvestris, for example, began many years before the impacts on Picea abies), and the sharp increase in the summer temperature in the Alps may have played an important role. Temperate deciduous broadleaved species suffered from the harsh heatwaves and droughts that have occurred since 2010. These species showed a certain species-specific degree of resilience (the capacity to restore the condition prior to the impact) depending on their ability to recover from cavitation, but they may be prone to attacks by opportunistic fungal parasites.
The ICP Forests Level I network was established to produce reliable data at the European level (and in some cases at the national level), but it can miss localised forest types and events (e.g., in our research, coastal Mediterranean forests and shrublands). It may be of interest for the forest policies to deploy a locally intensified network to improve the informative potential.
Acknowledgements
The research has been carried out within the project “LIFE MODERn(NEC) - new MOnitoring system to Detect the Effects of Reduced pollutants emissions resulting from NEC Directive adoption” - LIFE20 GIE/IT/000091.
The authors acknowledge the support from the National Biodiversity Future Center to University of Florence, funded by the Italian Ministry of University and Research, PNRR, Missione 4 Componente 2, “Dalla ricerca all’impresa”, Investimento 1.4, Project CN00000033.
References
CrossRef | Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Online | Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Enrico Cenni
Davide Bettini
Giovanni Iacopetti 0000-0002-1472-4435
Luisa Ghelardini 0000-0002-3180-4226
Salvatore Moricca 0000-0003-3097-559X
Tiziana Panzavolta 0000-0002-8985-3071
Matteo Bracalini 0000-0002-0402-8604
Martina Pollastrini 0000-0003-0959-9489
Department of Agriculture, Food, Environment and Forestry (DAGRI), University of Firenze, Firenze (Italy)
Domenico Di Martino
Cristiana Cocciufa
Claudia Cindolo
Arma dei Carabinieri, Comando per la Tutela della Biodiversità e dei Parchi, Roma (Italy)
NBFC, National Biodiversity Future Centre, Palermo 90133 (Italy)
Corresponding author
Paper Info
Citation
Bussotti F, Papitto G, Di Martino D, Cocciufa C, Cindolo C, Cenni E, Bettini D, Iacopetti G, Ghelardini L, Moricca S, Panzavolta T, Bracalini M, Pollastrini M (2024). Extreme climatic events, biotic interactions and species-specific responses drive tree crown defoliation and mortality in Italian forests. iForest 17: 300-308. - doi: 10.3832/ifor4531-017
Academic Editor
Marco Borghetti
Paper history
Received: Nov 29, 2023
Accepted: Jul 21, 2024
First online: Sep 30, 2024
Publication Date: Oct 31, 2024
Publication Time: 2.37 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2024
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 10018
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 5504
Abstract Page Views: 2404
PDF Downloads: 1694
Citation/Reference Downloads: 1
XML Downloads: 415
Web Metrics
Days since publication: 643
Overall contacts: 10018
Avg. contacts per week: 109.06
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
(No citations were found up to date. Please come back later)
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Review Papers
Monitoring the effects of air pollution on forest condition in Europe: is crown defoliation an adequate indicator?
vol. 3, pp. 86-88 (online: 15 July 2010)
Research Articles
Influence of climate on tree health evaluated by defoliation in the ICP level I network (Romania)
vol. 10, pp. 554-560 (online: 05 May 2017)
Research Articles
First results of a nation-wide systematic forest condition survey in Turkey
vol. 4, pp. 145-149 (online: 01 June 2011)
Research Articles
Predicting tree crown defoliation using color-infrared orthophoto maps
vol. 6, pp. 23-29 (online: 14 January 2013)
Research Articles
Estimating crown defoliation of Scots pine (Pinus sylvestris L.) trees using small format digital aerial images
vol. 6, pp. 15-22 (online: 14 January 2013)
Editorials
COST Action FP0903: “Research, monitoring and modelling in the study of climate change and air pollution impacts on forest ecosystems”
vol. 4, pp. 160-161 (online: 11 August 2011)
Research Articles
Long-term monitoring of air pollution effects on selected forest ecosystems in the Bucegi-Piatra Craiului and Retezat Mountains, southern Carpathians (Romania)
vol. 4, pp. 49-60 (online: 05 April 2011)
Commentaries & Perspectives
Availability, accessibility, quality and comparability of monitoring data for European forests for use in air pollution and climate change science
vol. 4, pp. 162-166 (online: 11 August 2011)
Editorials
Adaptation of forest ecosystems to air pollution and climate change: a global assessment on research priorities
vol. 4, pp. 44-48 (online: 05 April 2011)
Research Articles
A fast screening approach for genetic tolerance to air pollution in Scots pine field tests
vol. 6, pp. 262-267 (online: 01 July 2013)
iForest Database Search
Search By Author
- F Bussotti
- G Papitto
- D Di Martino
- C Cocciufa
- C Cindolo
- E Cenni
- D Bettini
- G Iacopetti
- L Ghelardini
- S Moricca
- T Panzavolta
- M Bracalini
- M Pollastrini
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
- F Bussotti
- G Papitto
- D Di Martino
- C Cocciufa
- C Cindolo
- E Cenni
- D Bettini
- G Iacopetti
- L Ghelardini
- S Moricca
- T Panzavolta
- M Bracalini
- M Pollastrini
Search By Keywords
PubMed Search
Search By Author
- F Bussotti
- G Papitto
- D Di Martino
- C Cocciufa
- C Cindolo
- E Cenni
- D Bettini
- G Iacopetti
- L Ghelardini
- S Moricca
- T Panzavolta
- M Bracalini
- M Pollastrini
Search By Keyword







