Assessing forest resilience after windthrow: linking deadwood, regeneration, and landscape-scale susceptibility - A case study in a Pre-Alpine forest in Italy
iForest - Biogeosciences and Forestry, Volume 19, Issue 2, Pages 102-113 (2026)
doi: https://doi.org/10.3832/ifor4982-019
Published: Apr 07, 2026 - Copyright © 2026 SISEF
Research Articles
Abstract
Severe windstorms account for nearly half of all damage to European forests. South of the Alps, windstorms such as Storm Vaia in 2018 and Storm Alex in October 2020 have left forest managers uncertain on how to assist habitat recovery and how to prevent further damage in the intact portions of the forest. Since storms did not occur regularly in this part of Europe, questions remain about whether to remove windthrown deadwood and which stand, topographic, and microclimatic traits most strongly predispose forests to future windthrow. This study investigates the characteristics affecting the likelihood of windthrow and the effects of windthrow severity on forest regeneration and habitat recovery across three study sites in Alto Verbano, Lombardy, Italy. The objectives of the study were twofold: (i) to examine the relationship between deadwood accumulation and forest regeneration, and (ii) to assess the influence of forest structure, topography, and microclimate on windthrow susceptibility. We combined field sampling with drone-based optical imagery to collect detailed data on deadwood biomass and cover, as well as forest regeneration. We investigated the relationship between deadwood and regeneration indices using Generalized Linear Models and Structural Equation Modeling to test for effects both direct and indirect, i.e., through changes in topographical and environmental factors. To assess windthrow susceptibility, we used Sentinel-2 satellite data, integrated with microtopographic information from satellite products, to assess landscape-level vulnerability to windthrow using non-parametric machine learning methods. The results reveal a negative influence of deadwood on regeneration, irrespective of topographical features and threats to regeneration levels at the sites (i.e., ungulate browsing). On the other hand, forest structure, topography, and microclimate had a critical role in determining forest vulnerability to windthrow. These findings emphasize the importance of carefully considering deadwood management and future forest structural features when carrying out forest restoration actions. Assessing the consequences of deadwood accumulation and understanding forest vulnerability to windthrow are essential for optimizing forest regeneration and resilience.
Keywords
Post-windthrow, Forest Recovery, Ecological Resilience, Deadwood-regeneration Relationship
Introduction
Windstorms represent the leading natural disturbance affecting European forests, being responsible for about 46% of all damage caused by disturbances from 1950 to 2019 ([23]). The consequences of windthrow are substantial from both economic and environmental perspectives, with average annual timber losses reaching 43.8 million cubic meters over the last seven decades ([23]). Such large-scale windthrows profoundly disrupt forest ecosystems, creating canopy gaps that increase light availability in the understory and trigger a cascade of ecological changes ([34]). These effects range from the loss of mature trees to shifts in soil microbiological communities, collectively transforming the original structure and function of the habitat ([10], [15]). Over time, the absence of vegetation in disturbed areas leads to severe soil erosion and a decline in soil organic matter, altering humus quality and significantly impacting soil communities ([36]). These effects make windthrow a critical ecological concern in forest areas, especially those not traditionally hit by this damaging agent until now. Here, understanding which characteristics influence the likelihood of windthrow and the effects on habitat recovery post-windthrow is essential for assessing the vulnerability of forested habitats and assisting their recovery. We hereby operate the assessment of forest resilience based on its ability to recover pre-disturbance structure and composition (also known as engineering resilience).
Remote sensing is a widely used technology for monitoring various stages of disaster management, delivering rapid and relevant information to support planners, scientists, and decision-makers ([4]). This technology enables the collection of detailed information on changes in spectral characteristics, vegetation indices (e.g., NDVI), and red-edge channels, providing critical insights into forest structure and tree health ([11], [20], [18]). Additionally, remote sensing can be used to assess topographic variables such as slope, elevation, and aspect, which influence wind exposure and forest vulnerability. Environmental conditions, particularly soil moisture and precipitation patterns, can also be indirectly monitored using radar and multispectral imagery, enabling the detection of fluctuations in soil water content and vegetation stress following storm events ([18], [12]).
Remote sensing can also support management decisions following windthrow events. Traditional ground surveys, while essential for calibrating ecological models, are too labour-intensive to track the highly patchy and rapidly evolving successional mosaic that follows a major windthrow ([27], [26]). Recent advances in high-resolution satellite, UAV, and LiDAR remote sensing can provide spatially continuous, multi-temporal data on canopy structure, deadwood volume, and early regeneration signals across entire landscapes ([12]). Leveraging these observations allows managers to prioritize intervention zones, optimise deadwood removal, and target assisted regeneration or other active restoration where the greatest ecological and safety benefits can be achieved ([33]).
This study aims to (i) examine the recovery patterns of wind-damaged habitats by assessing the regeneration stages, disturbance legacies, and species composition within the affected areas. By focusing on the structural and compositional dynamics of the forest during regeneration, we seek to understand how these ecosystems respond to severe disturbances over time. This study also aims to (ii) evaluate the environmental, topographical, and structural drivers of forest vulnerability to windthrow, to assess the risk of further wind damage in other sectors of the study area.
Methodology
Site description
The study was conducted in the Alto Verbano Mountain Community, located in the Varese province of Lombardy, Italy, with coordinates ranging from 45° 53′ 24″ N, 08° 36′ 07″ E to 46° 07′ 13″ N, 08° 51′ 48″ E. This area is part of the Varese Prealps, characterized by diverse topographical features, including steep slopes, valleys, and ridges, with elevations ranging from 120 m a.s.l. in the valleys to 1545 m at the highest peaks. The region supports temperate forests with a mix of broadleaf and conifer species, including European beech(Fagus sylvatica), Oak (Quercus spp.), Maples (Acer spp.), Birch (Betula spp.), Ash (Fraxinus spp.). in the past century, extensive conifer plantation was also carried out, using Norway spruce (Picea abies) and European larch (Larix decidua) for pulp production and recreation purposes.
The climate is classified as Dfb (Warm-summer humid continental) and Cfb (Temperate oceanic), according to the Köppen-Geiger system. Based on weather data from the weather station “Aviosuperficie Mascioni” (45° 53′ 56″ N, 08° 43′ 17″ E) recorded between January 2021 and February 2025, the region experiences peak temperatures in July and August, with an average maximum temperature of 17.9 °C and a minimum of 7.4 °C. Precipitation is highest in May, with an average precipitation rate of 8.7%, while July is the driest month. Winds in the region are typically strongest in April, with an average speed of 3.7 knots, and the prevailing wind direction varies throughout the year, predominantly from the northwest.
Regeneration profile before disturbance
Pre-disturbance regeneration conditions were characterized using data from the Italian National Forest Inventory (NFI) sampled in 2015, extracted from sample plots in the western part of the Lombardia region, which corresponds to the study area. Regeneration attributes were analyzed by selecting plots according to the main forest types present within the windthrow gaps, namely “Faggete” (beech forests), “Castagneti” (chestnut forests), “Aceri” (maple- and mixed broadleaf forests), and forests under anthropogenic management (see Tab. S3 in Supplementary material for details).
Overall, the regeneration structure differed markedly among forest types. Faggete exhibited a predominantly top-heavy regeneration profile, with regeneration largely concentrated in the tallest height/diameter class, while smaller saplings were sparse or absent in many plots. This uneven vertical distribution indicates limited recent recruitment relative to past successful establishment events. In contrast, Castagneti showed a more balanced and dynamic regeneration structure, with high densities in both small and intermediate classes and a substantial contribution of younger cohorts to total regeneration biomass and carbon, consistent with continuous multi-cohort regeneration.
Anthropogenically managed forests displayed a discontinuous regeneration layer, strongly dominated by taller saplings and characterized by weak or absent recruitment in the smallest size class. Regeneration biomass and carbon were therefore concentrated in the upper regeneration layer, reflecting episodic regeneration linked to past management interventions rather than continuous establishment. Conversely, Aceri forests were clearly bottom-heavy, with regeneration dominated by very small saplings and only sporadic presence of intermediate individuals, while the tallest regeneration class was largely absent. Biomass and carbon stocks were consequently concentrated in the smallest size class, indicating recent, locally intense recruitment pulses that had not yet translated into a well-developed tall regeneration layer.
Taken together, these patterns indicate that, prior to the storm disturbance, regeneration dynamics across forest types ranged from structurally mature but recruitment-limited systems (Faggete), through actively regenerating and vertically structured stands (Castagneti), to regeneration layers shaped by management legacies (anthropogenic forests), and finally to early-stage, recruitment-dominated systems (Aceri). These contrasting pre-disturbance regeneration profiles provide an essential baseline for interpreting post-windthrow regeneration responses.
Pre-disturbance deadwood profile
Pre-disturbance deadwood conditions were characterized using data from the Italian National Forest Inventory (NFI) carried out in 2015. Data were extracted from sample plots in the western part of the Lombardia region corresponding to the study area and stratified by the dominant forest types present within the windthrow gaps. Deadwood structure differed substantially among forest types, both in quantity and in the relative importance of standing versus downed components (Tab. S4 in Supplementary material).
Faggete (beech stands) exhibited the richest and most structurally complex deadwood profile. Deadwood was clearly dominated by standing dead trees, with generally high snag densities and substantial basal area and volume, indicating the presence of medium- to large-diameter dead stems. Standing deadwood accounted for a large share of total deadwood biomass and carbon, while downed coarse woody debris (CWD) was highly heterogeneous, ranging from nearly absent to locally abundant. This combination reflects mature or partially unmanaged stand conditions with strong spatial variability in mortality and deadwood retention.
In Castagneti (chestnut stands), deadwood was characterized by relatively low snag densities but large individual dead stems, resulting in substantial snag basal area, volume, and carbon stocks despite the limited number of snags. In contrast, downed CWD was scarce across most plots, indicating a deadwood profile dominated by standing elements and limited accumulation of ground material. This pattern is consistent with managed stands where large dead trees persist as structural legacies, while downed wood is either removed or does not accumulate.
Anthropogenically managed forests showed the sparsest deadwood profile overall. Standing dead trees were generally few and of smaller diameter, resulting in low to moderate deadwood biomass and carbon stocks. Downed CWD was almost absent across plots, yielding a simplified deadwood structure with limited vertical and horizontal complexity compared to more natural forest types.
Aceri forests (maple- and mixed broadleaf stands) displayed an intermediate but distinctive deadwood profile, with consistently high snag densities combined with moderate basal area and volume, indicating numerous small- to medium-diameter standing dead trees. Ground CWD was variable but occasionally substantial, contributing to a more balanced distribution between standing and downed deadwood than in managed forest types. This structure suggests active mortality processes and partial retention of deadwood prior to disturbance.
Overall, before the storm disturbance, deadwood conditions ranged from snag-rich, structurally diverse systems (Faggete) through snag-dominated, managed stands (Castagneti) to simplified anthropogenic forests with minimal deadwood, and finally to Aceri stands with abundant snags and variable downed wood. These contrasting deadwood legacies provide an important baseline for interpreting post-windthrow structural changes, habitat availability, and carbon dynamics.
On the night of October 2-3, 2020, Storm Alex swept across southeastern France and Northwest Italy, including the area of Alto Verbano. Severe thunderstorms and heavy rainfall, along with sirocco winds, impacted the region. The weather station at the GV Schiaparelli Astronomical Observatory, located at Punta Paradiso on Mount Campo dei Fiori, recorded wind speeds of 113 km h-1 before the instruments lost power and ceased functioning. This severe weather caused significant damage to our mountains, resulting in the falling of thousands of trees, including beech, chestnut, and ash ([35]). Some of the affected areas are included in the system of regional parks and the EU Natura 2000 network, leaving managers with open questions about how best to assist the recovery of habitat quality and forest value for biodiversity.
In this study, the assessment of the windthrow event is focused on the territories of Sasso del Ferro, Monte Nudo, and Scereda, included in the Site of Community Importance IT2010019 “Monti della Valcuvia”, where the following habitat types occur: Sub-Atlantic and mid-European oak or oak-hornbeam forests of the Carpinion betuli (9160), Asperulo-Fagetum beech forests (9130), and Tilio-Acerion forests of slopes, screes, and ravines (9180*). While the most severe damage in Monte Nudo (75.5%) was to coniferous vegetation, in Scereda and Sasso del Ferro, all the damaged vegetation was broadleaf.
Some salvage logging and soil stabilization measures have already been carried out in Monte Nudo and Sasso del Ferro before this study. According to the salvage logging plan obtained from the owner of the company entitled to the works, 80% of the damaged area in Monte Nudo has already been cleared. In Sasso del Ferro, salvage logging was less extensive, with an estimated 20% of the area cleared. Logging activities in Scereda have been scarce due to a landslide that has further limited access to the site.
Damage mapping
To identify and map windthrow gaps, we used Sentinel-2 satellite images due to their high spatial resolution (10 m) and suitability for large-scale analysis. To ensure optimal visibility of windthrow gaps, images were pre-screened using the following criteria: clear identification of windthrow gaps, consistent color representation, and a maximum cloud cover of 30%. The Sentinel-2 L2A true-color image from June 10, 2021, was selected as the most suitable dataset.
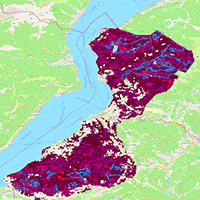
We manually digitized 19 gaps (estimated area: 18.93 ha) and 13 adjacent non-gap patches (estimated area: 24.05 ha) as regions of interest to train a model for supervised classification of gaps across the whole study area. We then trained a Random Forest (RF) algorithm (number of trees = 500) using the Red, Blue, and Green bands of the True Color Sentinel 2 image to classify windthrow gaps within the imagery. The execution of this algorithm was done using the “ranger” package ([38]) for R ([25]). Post-classification filtering was applied to exclude non-windthrow gaps, such as agricultural areas and barren soil, by leveraging supplementary datasets, including barren soil and agricultural land-use layers from the Copernicus Land Cover map for Europe (June 2019). Windthrow gaps were refined by intersecting these polygons with the initial classification, excluding non-windthrow sub-areas. This ensured that the final data set accurately represented true windthrow gaps from the 2020 disturbance event (Fig. 1).
Fig. 1 - Location of windthrow after random forest classification and post-classification filtering.
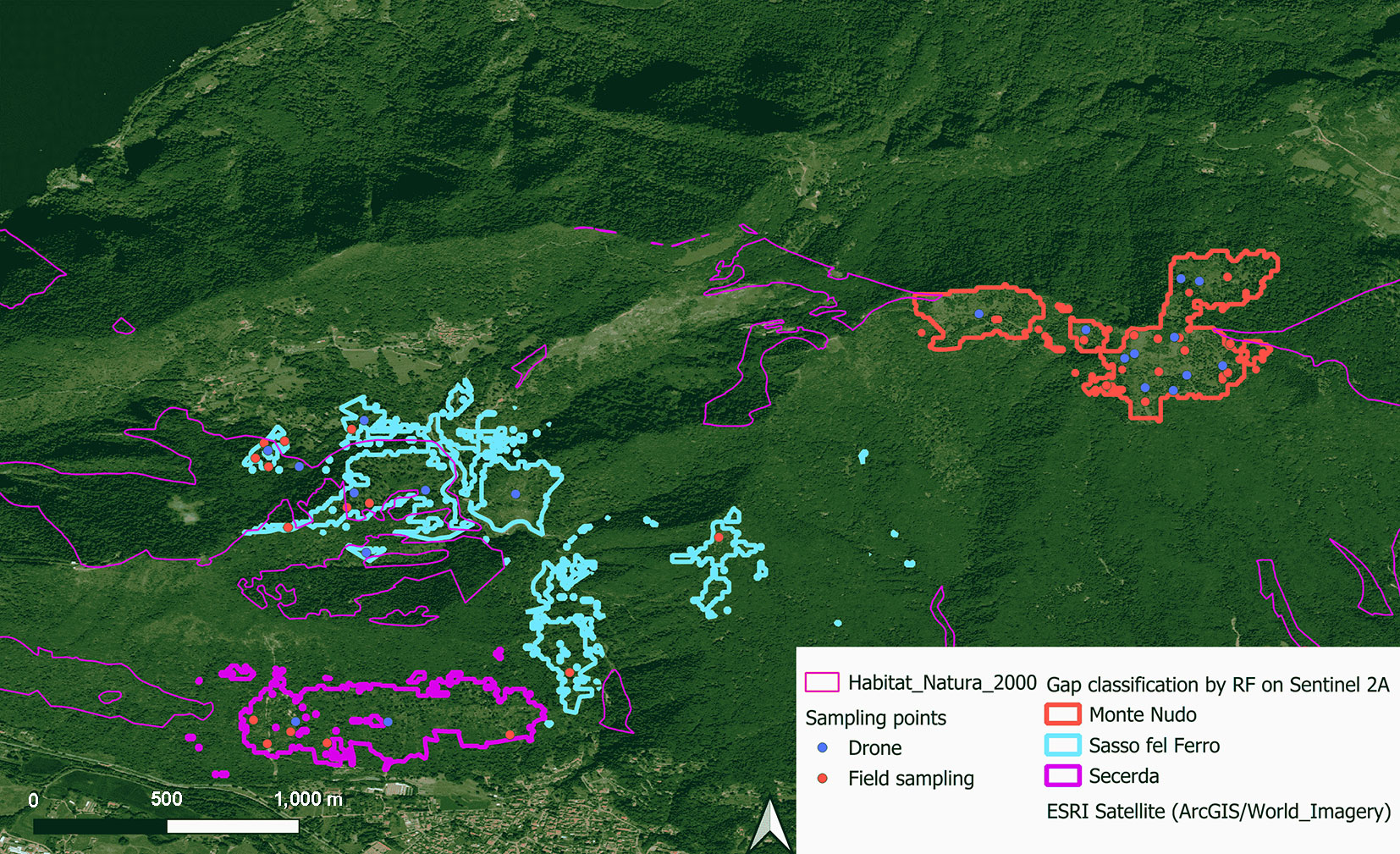
Field survey
Twenty-six 10 × 10 m survey points were established to collect detailed data on forest regeneration in areas affected by windthrow. Areas cleared post-disturbance were avoided. Sampling points were allocated based on the proportion of the windthrow area at each location, with 11 random points assigned to Monte Nudo, 4 random points to Scereda, and 11 random points to Sasso del Ferro, to ensure comprehensive representation of the different conditions in the windthrow areas. To avoid spatial overlap, the sampling points were located at least 100 meters apart. In each plot, the following parameters related to seedling regeneration (individuals with heights from 5 to 330 cm) were recorded: Species identity, Seedling count per species, Percent cover of each species, Total percent seedling cover, Mean seedling height per plot. Moreover, three main deadwood parameters were assessed: Percent soil cover by coarse deadwood (CWD), biomass, and decay classification. Deadwood biomass was converted to carbon content (C) using species-specific conversion factors reported by Birdsey ([3]). Specifically, for broadleaves (eqn. 1):
and for conifers (eqn. 2):
where V is the volume of CWD (m3 ha-1), estimated using the line-intersect sampling method (eqn. 3):
where d is the diameter (cm) at the point of intersection, L is the length of the downed log (m), measured using a Leica laser distance device. Deadwood pieces were further classified into decay stages following the classification scheme described in Waddell ([37]).
Drone survey
We collected optical aerial images using a DJI Mavic Pro 2 drone to complement ground-based surveys. The drone captured high-resolution images of windthrow-affected areas, documenting spatial patterns of regenerating vegetation, deadwood distribution, and overall soil conditions in areas difficult to reach from the ground. Drone imagery was analyzed to derive regeneration metrics - including total area of regeneration cover, identification and assessment of occupancy by dominant regenerating species - and deadwood metrics - including total soil cover by deadwood (%), hereby referred to as deadwood occupancy, estimation for deadwood biomass (eqn. 1, eqn. 2), decay stage identified from image characteristics of the plot.
To derive deadwood measurements from UAV imagery, we placed a 2-meter ground-truth ruler within each plot prior to drone flight. The high-resolution UAV images (2 cm spatial resolution) were then geometrically scaled in QGIS v. 4.0 (⇒ https://qgis.org/) using this reference object. Once scaled, the diameter (d) of downed wood at the intersection point and the log length (L) were measured directly from the orthomosaic using the distance measurement tool. These measurements were used to estimate coarse woody debris volume (m3 ha-1) and biomass using the same formula (eqn. 1, eqn. 2) as for field data, enabling consistent comparison between field and image-derived estimates. Each field plot was covered by a 20×20 m UAV orthomosaic; plot-level deadwood estimates were cross-validated between field and UAV data (R2 = 0.89).
This multisource approach, combining field surveys and drone imagery, provided a comprehensive understanding of forest regeneration dynamics and deadwood characteristics within the study area.
Effect of deadwood on regeneration
The structure and composition of regeneration in damaged sites were summarized by identifying the species present and their respective current occupancy in each plot.
We fitted Gamma General Linear Models (GLM) to examine the relationship between standardized deadwood indices (i.e., percent soil cover, biomass, and decay stage) and regeneration indices (i.e., percent seedling cover and mean seedling height) for 26 field survey plots. GLMs included the site as a fixed effect to control spatial clustering. This analysis aimed to reveal in-depth the significant association between the presence of deadwood and regeneration dynamics. We pre-screened GLMs by assessing the explanatory power and parsimony of all possible combinations of predictors using the Bayesian Information Criterion (BIC) from the package “leaps” in R ([25]).
We then used Structural Equation Modelling (SEM) to further explore the influence of deadwood on regeneration through both direct and indirect effects. We developed two models: (i) Direct Influence Model: this model hypothesized a direct relationship where deadwood affects regeneration without intermediate variables. (ii) Indirect Influence Model: this model posits that deadwood indirectly influences regeneration through its effect on microtopography and seedling vulnerability to damage. To account for ungulate browsing, we used field data collected in September 2024 across 80 systematically distributed plots within the same research area extent (methodology in [7]). At each plot (225 m2), regeneration was recorded as the number of seedlings and saplings (5-270 cm), alongside evidence of ungulate damage classified as browsing, fraying, or debarking ([2]). Ungulate browsing pressure was calculated as the proportion of damaged seedlings over the total number of seedlings in each plot. This pressure index was then interpolated into a continuous raster surface using the Inverse Distance Weighting (IDW) method, providing spatially explicit estimates of browsing exposure across the study area.
Windthrow vulnerability of remaining forests
Both in windthrow and non-windthrow patches manually digitized, we extracted the values of the following variables driving forest vulnerability to wind, to fit a Random Forest model of windthrow probability: (i) Slope was sourced from the DTM available at the Geoportale della Lombardia ([14]). (ii) The most recent pre-windthrow data for tree cover density (2018) from the Copernicus Land Monitoring Service (retrieved from: ⇒ https://doi.org/10.2909/59b0620c-7bb4-4c82-b3ce-f16715573137). (iii) Canopy height was acquired from the Global Canopy Height Dataset developed by Lang et al. ([16]). Images used in the model were collected between May and September 2020, making them highly representative of forest structures prior to the windthrow event in October 2020. Forest height heterogeneity was then derived from this data using the Gray-Level Co-Occurrence Matrix index of Dissimilarity, with a 3×3 window and a shift of 1.1 ([22], [39]). (iv) Data on the dominant forest type for 2018 were also obtained from the Copernicus Land Monitoring Service (⇒ https://doi.org/10.2909/7b28d3c1-b363-4579-9141-bdd09d073fd8). (v) Soil properties, including soil packing density and soil bulk density for depths of 0-10 cm and 10-20 cm, were sourced from Panagos et al. ([21]). (vi) Wind regime data, measured at a height of 50 meters above ground level, were retrieved from the Global Wind Atlas (⇒ https://globalwindatlas.info).
Values were extracted from each source raster layer at its original spatial resolution, then resampled to 10 meters using bilinear interpolation.
We ran a t-test to estimate each predictor’s contribution to the gap vs. no-gap classification. We then divided the dataset into training (70%) and testing (30%) subsets and used them to fit a Random Forest model (number of trees = 500). We computed permutation importance for each variable to identify key factors influencing gap formation. Using the trained Random Forest model, we then extrapolated predictions across the raster stack that encompassed all Natura 2000 sites in the study area. This provided spatial predictions of the probability of windthrow damage for every forested pixel across the landscape, under the implicit assumption that such forests would be exposed to events similar in magnitude and characteristics to the Alex storm (i.e., peak wind speed during the event was not explicitly accounted for in this analysis).
Results
Effect of deadwood on regeneration
Faggete (beech stands) represents late-successional native beech forests that regenerate primarily through natural seed dispersal in canopy gaps, forming shade-tolerant cohorts within uneven-aged structures typical of old-growth dynamics. Castagneti (chestnut stands) are cultural forests, often of planted or selected origin, that regenerate via prolific stump sprouting and seed production, supporting continuous recruitment under managed coppice systems. Anthropogenic forests arise from human interventions such as reforestation or afforestation, relying on planted stock with limited natural regeneration pathways, shaped by site preparation and species choice. Aceri (Aceri-frassineti and aceri-tiglieti) are mixed late-successional broadleaf stands dominated by shade-tolerant native maples, ashes, and limes that regenerate naturally from advance growth and small-scale disturbances. These pre-disturbance characteristics explain the observed profiles: top-heavy in Faggete and anthropogenic forests from episodic native pulses, bottom-heavy in Aceri from recent shade-tolerant recruitment, and balanced in Castagneti through vegetative propagation. Deadwood biomass varied across forest types and sites. In Sasso del Ferro broadleaf stands, values were moderate, with most plots below 10 m3 ha-1 and a median around 6-7 m3 ha-1. Monte Nudo conifer stands showed the widest variability, with a median near 8 m3 ha-1, but some plots exceeded 20 m3 ha-1, indicating localized accumulation. Monte Nudo broadleaf and Scereda broadleaf forests had comparable levels, centered around 6-7 m3 ha-1, though Scereda displayed slightly greater spread. Monte Nudo mixed stands were more uniform, consistently around 10 m3 ha-1 with little variation, suggesting more homogeneous conditions.
The identification of dominant species in the regeneration layer was consistent across field surveys and drone imagery (Tab. 1, Tab. 2). In Monte Nudo (MN), pioneer species like Betula spp. and Larix decidua dominate the regeneration scenario (totally 70.95% area of regenerating species from the field survey - Tab. 1, 58.85% area of regenerating species from the drone survey - Tab. 2), but the height variability in Betula spp. (75.8 ± 126 cm - Tab. 1) and the low cover of European larch (1.68% and 10.36% for the regenerating proportion from field and drone surveys, respectively) suggests uneven regeneration success or establishment at different times. Corylus avellana (common hazel), an early-successional species, ranked second in coverage, showing robust growth where it occurs (22.35% area of regenerating species), and potentially creating favorable microenvironments for late-successional species. Late-successional species (e.g., European beech and Norway spruce) are present but not yet significantly contributing to the site regeneration (3.35% of regenerating species). The height of these late-successional species indicates that they are likely in the initial establishment phase (30 ± 22.3 cm for beech, 6.5 ± 2.12 cm for spruce). This suggests an early phase of forest recovery in Monte Nudo, where pioneer species are crucial for stabilizing the site.
Tab. 1 - Summary of the structure and composition of regeneration from field observation.
| Site | Regenerating species |
Successional group |
Cover (m2) |
Regeneration proportion (%) |
Mean height (cm) |
|---|---|---|---|---|---|
| MN | Betula spp. | Pioneer species | 62 | 69.27 | 75.8 ± 126 |
| Fagus sylvatica | Late successional | 3 | 3.35 | 30 ± 22.3 | |
| Larix decidua | Pioneer to mid-successional | 1.5 | 1.68 | 11 ± 5.66 | |
| Corylus avellana | Early successional | 20 | 22.35 | 350 ± 84.2 | |
| Picea abies | Mid- to late-successional | 3 | 3.35 | 6.5 ± 2.12 | |
| SdF | Betula spp. | Pioneer species | 120 | 15.13 | 153.75 ± 69.9 |
| Fagus sylvatica | Late successional | 260 | 43.70 | 87.71 ± 53 | |
| Corylus avellana | Early successional | 215 | 36.13 | 390 ± 36.1 | |
| Scereda | Ailanthus altissima | Invasive pioneer species | 10 | 90.91 | 300 ± 62.1 |
| Castanea sativa | Mid-successional | 1 | 9.09 | 50 ± 13.5 |
Tab. 2 - Summary of the structure and composition of regeneration from drone observation.
| Site | Regenerating species |
Successional group |
Cover (m2) |
Regeneration cover (%) |
|---|---|---|---|---|
| MN | Acer spp. | Mid to late succession | 14 | 1.50 |
| Betula spp. | Pioneer | 454 | 48.50 | |
| Fagus sylvatica | Late succession | 371 | 39.64 | |
| Larix decidua | Pioneer | 97 | 10.36 | |
| SdF | Fagus sylvatica | Late succession | 672 | 97.11 |
| Robinia pseudoacacia | Invasive early successional | 20 | 2.89 | |
| Scereda | Ailanthus altissima | Invasive pioneer | 150 | 100.00 |
In Sasso del Ferro (SdF), field survey showed that pioneer species like birch are well-established in terms of height (153.75 ± 69.9 cm). Common hazel showed robust growth and was a significant contributor to regeneration, indicating the transition of dominance from pioneer to early-successional species. The coverage proportion at this site was skewed towards late successional species, such as beech. This group was widespread in the understory, covering 260 and 672 m² according to field and drone surveys, respectively. This represents 43.70% and 97.11% of the area occupied by regenerating species. However, the shorter height of late successional species suggests that they are still competing with faster-growing early successional species (average height: 87.71 ± 53 cm vs. 390 ± 36.1 cm for beech and hazel, respectively). This competition is concerning, especially in the presence of the invasive Robinia pseudoacacia (Tab. 2). This site represents a transitional stage of regeneration. Pioneer species are well-established, early successional species are thriving, and late-successional species are becoming increasingly abundant.
At the Scereda site, the pioneer species is Ailanthus altissima, an invasive species with rapid growth (90.91% and 100% area of regenerating species, respectively, according to field and drone surveys) and dominant (height: 300 ± 62.1 cm) among the regenerating species. The lack of native early-successional species at this site may be due to high competition from the rapidly growing invasive pioneer species. No other pioneer or early successional species were registered.
In Monte Nudo, regeneration cover and regeneration height were significantly and negatively correlated with deadwood occupancy and deadwood biomass (Tab. 3), meaning that this site has the more deadwood occupancy, the less cover (β = -0.56) and the shorter regeneration (β = -0.71). In Sasso del Ferro, the same negative correlation between regeneration coverage and deadwood occupancy was observed, although less pronounced than in Monte Nudo (β = -0.41), meaning that for the same increase in deadwood occupancy, the decrease in regeneration is less intense than in Monte Nudo. In Scereda, the lower deadwood biomass is associated with higher regeneration cover. We found a significant positive correlation between regeneration height and deadwood decay class (β = 0.71).
Tab. 3 - Site-specific correlation between regeneration indices and deadwood indices. (N) Number of plots; (*): p<0.05; (**): p<0.01; (***): p<0.001.
| Site | N | Correlation pair | |||||
|---|---|---|---|---|---|---|---|
| Regeneration coverage vs. | Regeneration height vs. | ||||||
| DW occupancy |
DW biomass |
DW decay |
DW occupancy |
DW biomass |
DW decay |
||
| MN | 12 | -0.56 *** | -0.31 *** | -0.11 | -0.71 *** | -0.34 | -0.27 |
| Scereda | 10 | -0.32 | -0.31** | -0.51 | -0.38 | 0.04 | 0.71* |
| SdF | 4 | -0.41*** | 0.04 | 0.10 | -0.47 | -0.03 | 0.07 |
| All site | 26 | -0.51*** | -0.26 | -0.15 | -0.62*** | -0.36 | -0.002 |
The GLMs for deadwood soil cover and deadwood biomass, as a function of regeneration height and cover, revealed significant negative effects on both response variables (Tab. 4).
Tab. 4 - Detailed results of Gamma GLM.
| Independent variable |
Variables | General model | Beech-specific model |
Birch-specific model |
|---|---|---|---|---|
| Regeneration height |
Intercept | 4.29*** | 4.17*** | 4.63*** |
| Deadwood occupancy | -1.14*** | -0.67* | -0.88** | |
| Regeneration height |
Intercept | 4.58*** | 4.49*** | 4.68*** |
| Deadwood biomass | -0.77*** | 0.31 | -0.39 | |
| Regeneration coverage |
Intercept | 2.57*** | 3.00*** | 2.87*** |
| Deadwood occupancy | -1.24*** | -1.29* | -0.92* | |
| Regeneration coverage |
Intercept | 2.86*** | 3.59*** | 2.90*** |
| Deadwood biomass | -0.58* | 0.28 | -0.02 |
After standardizing the predictors to facilitate direct comparison of effect sizes, we found that deadwood occupancy consistently exhibited a stronger influence on regeneration metrics than deadwood biomass across all models. In the general model, a one-standard-deviation increase in deadwood occupancy was associated with significant decreases in both regeneration height (β = -1.14, p < 0.001) and coverage (β = -1.24, p < 0.001), suggesting a strong negative impact of deadwood occupancy on overall regeneration.
For the Beech-specific model, the standardized effect of deadwood occupancy on regeneration height was relatively weaker (β = -0.67, p = 0.05), while its effect on coverage was remarkable (β = -1.29, p < 0.05). This indicates that, while deadwood occupancy modestly suppresses beech regeneration height, it exerts a stronger negative influence on its coverage. In contrast, deadwood biomass had no significant effect on either height or coverage in beech, confirming its limited role in shaping regeneration for this species.
The Birch-specific model showed moderate sensitivity to deadwood occupancy, with significant negative effects on both height (β = -0.88, p < 0.01) and coverage (β = -0.92, p < 0.05), though weaker than those observed in the pooled model. Deadwood biomass showed no significant effect, confirming that deadwood occupancy is the more influential predictor.
Overall, standardization clarified that deadwood occupancy is a consistently stronger predictor of seedling regeneration than deadwood biomass, particularly for explaining regeneration coverage. This highlights that structural occupancy by deadwood may create microsite conditions or competition effects that constrain regeneration more than the quantity of deadwood itself.
The direct SEM model revealed a significant negative correlation between deadwood occupancy and regeneration cover (β = -0.532, p < 0.001), indicating that higher deadwood occupancy is associated with lower regeneration cover. However, other deadwood indices, such as deadwood biomass and decomposition stage, did not significantly affect regeneration. Deadwood biomass had a marginally significant effect on dominant regeneration species (β = -0.059, p = 0.059), suggesting that deadwood biomass may influence which species regenerate, favoring species such as Chestnut, Tree-of-heaven, Birch, Robinia, and European beech, while suppressing regeneration of species such as Norway spruce, European larch, and Maples. Other deadwood indices did not show a significant direct influence on regenerating species (Fig. 2).
Fig. 2 - SEM models of A - Direct impacts of deadwood indices on regeneration indices and B- Indirect impacts of deadwood indices via slope, site, and ungulate pressure characteristics on the regeneration indices. (DW_c): Deadwood soil cover; (DW_m): Deadwood biomass; (dcm): Deadwood decomposition stage; (rg_p): Regeneration cover; (rg_s): Regenerating species (order: Castanea sativa, Ailanthus altissima, Betula spp., Robinia, European beech, Norway spruce, European larch, Acer); (slp): Slope; (sit): Site (order: Scereda, Monte Nudo, Sasso del Ferro); (u_P): Ungulate pressure.
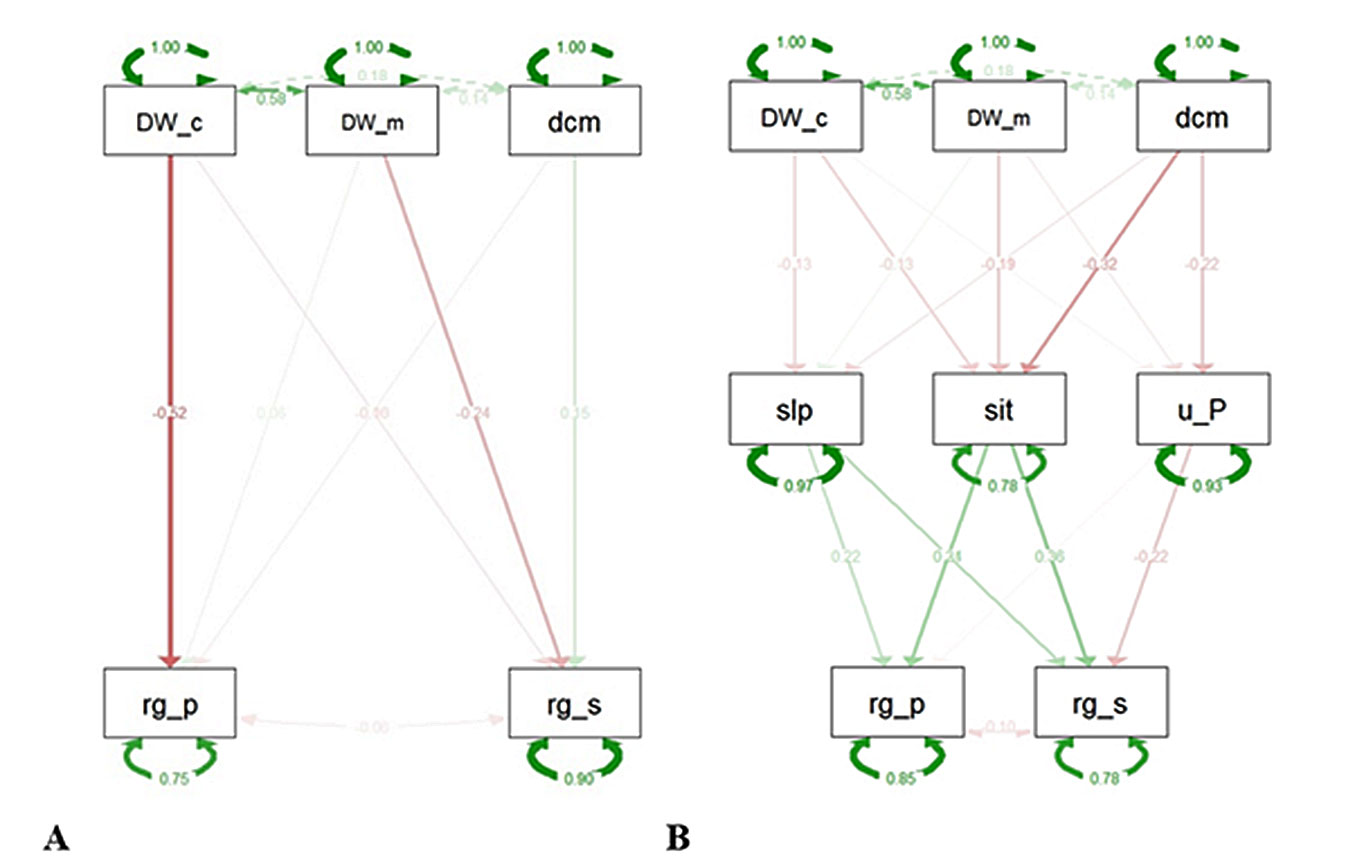
In the indirect model, none of the deadwood indices had a significant intermediating effect on microsite slope. However, the decay stage varied significantly across sites (p < 0.001), with more advanced decomposition at Sasso del Ferro and less advanced at Monte Nudo and Scereda. Decay stage significantly affected ungulate pressure (β = -3.048, p = 0.047), indicating that higher decomposition was associated with reduced ungulate pressure. Slope had a small but significant positive effect on regeneration cover (β = 0.016, p = 0.032). The site also showed a positive (β = 0.325) and significant (p-value = 0.003) effect, suggesting that regeneration cover was higher in Sasso del Ferro and Monte Nudo than in Scereda.
For the regeneration composition, all intermediate variables had a significant influence. Slope had a positive influence on regeneration genus, indicating seedlings of certain species prefer different slope conditions. Ungulate pressure had a negative influence on regeneration, indicating that ungulates suppressed certain regenerating species, such as Castanea sativa, Ailanthus altissima, and Betula spp.
Vulnerability to future windthrow
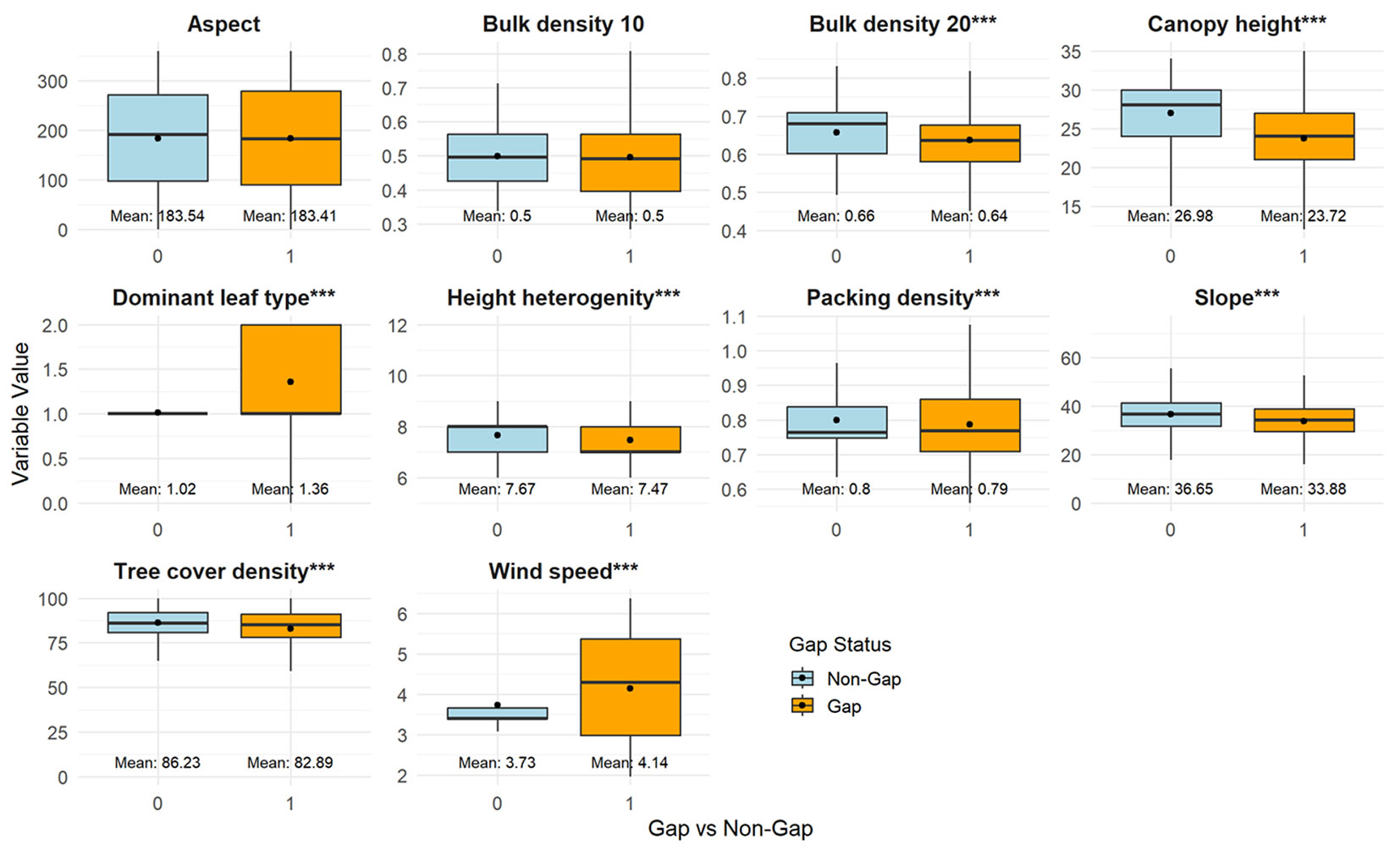
Areas with windthrow gaps differed from undamaged areas in several ways. These included soil bulk density at a depth of 10-20 cm, dominant leaf type, canopy height, height heterogeneity, soil packing density, slope, tree cover density, and average wind speed (Fig. 3). The boxplot comparison shows that gap probability is strongly linked to structural and environmental differences between forest stands. Gaps are more frequent in lower-canopy stands, suggesting that shorter stands are more vulnerable to windthrow, whereas taller canopies contribute to stability. Similarly, tree cover density, height heterogeneity, and packing density are slightly reduced in gap areas, indicating that denser, more heterogeneous stands provide greater mechanical resistance to wind disturbance. In contrast, bulk density at 20 cm is lower in gap areas, indicating looser soils that may weaken anchorage and increase the risk of uprooting. Slope is also shallower in gaps, implying that steeper terrain may buffer against windthrow. Among vegetation traits, the dominant leaf type differs significantly, with certain broadleaf morphologies more associated with gap formation, possibly due to aerodynamic effects. As expected, wind speed is consistently higher in gap areas, directly increasing the likelihood of disturbance. Meanwhile, aspect and bulk density at 10 cm differ little, suggesting a negligible role in gap probability. Overall, these results highlight that windthrow risk is amplified by weaker soil properties, lower stand structural complexity, and higher wind exposure.
Fig. 3 - Boxplots of values of potential vulnerability predictors in Gap (1) vs. Non-Gap (0) areas. (***): p < 0.001.
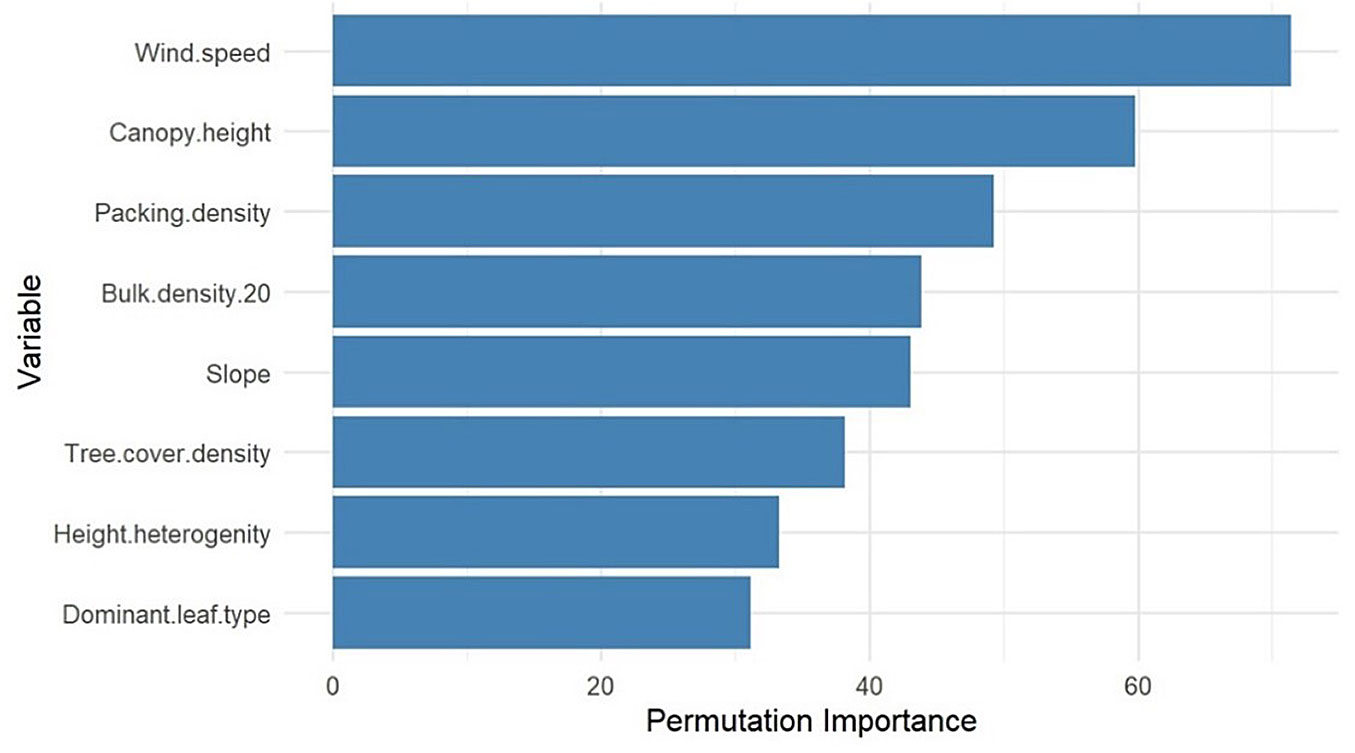
After we entered all variables with significant differences between gap and non-gap areas into our Random Forest model, wind speed emerged as the most critical determinant of windthrow likelihood. The second- and third-most important predictors of windthrow were canopy height, underscoring the significance of forest structural characteristics, and soil packing density (Fig. 4).
Fig. 4 - Variable importance for the probability of gap occurrence calculated from the Random Forest model.
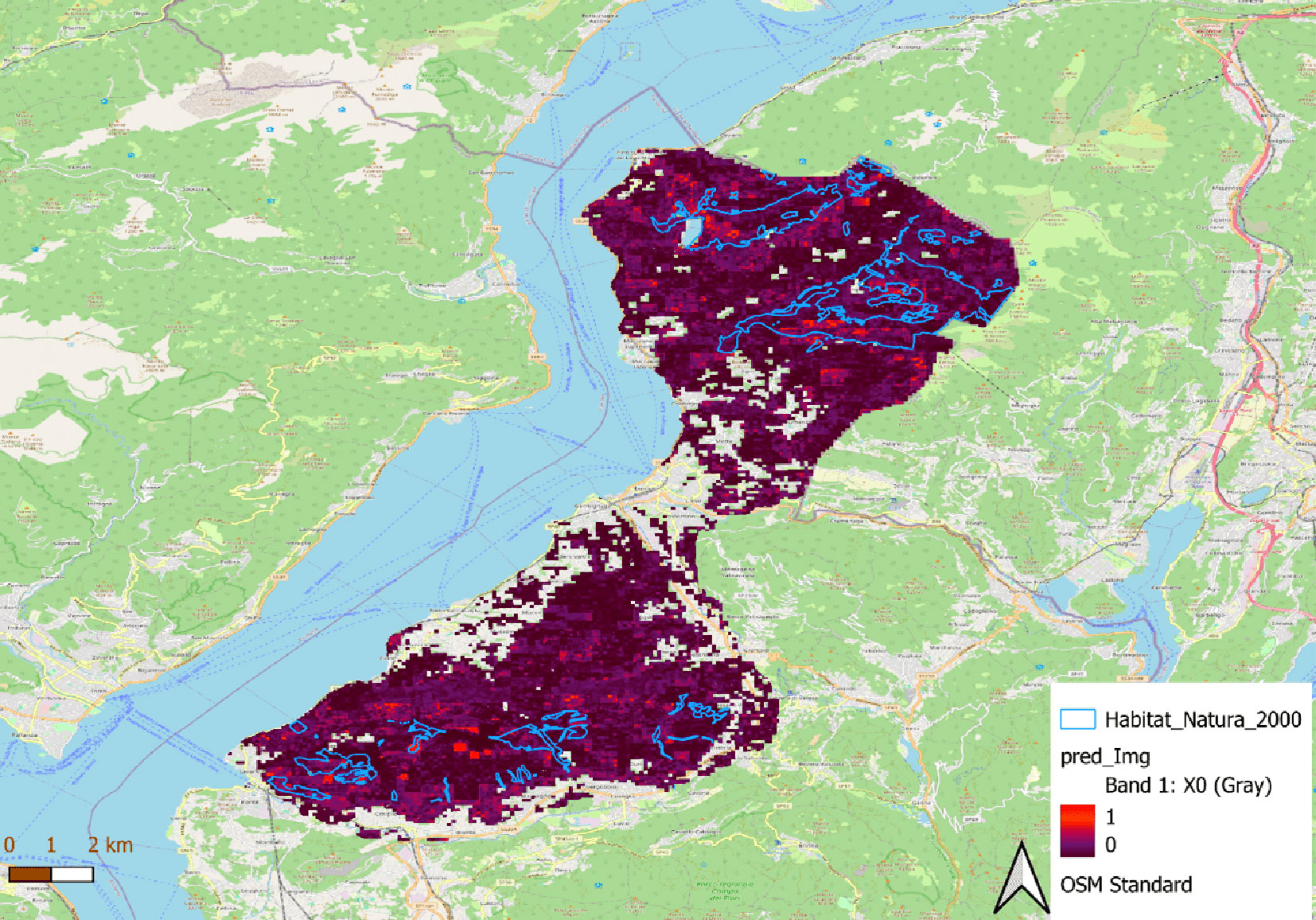
The extrapolation of the Random Forest model highlighted areas with high windthrow vulnerability, such as those in Monte Gradisea, Alpe Chedo, Monte Colmegnino, Alpe Ca’ del Sasso, Monte Sirti, North of Lago Delio in the North of the study area; and Sasso del Ferro, Monte Nudo, Alpe San Michele, San Martino, Pian Cuvinicc in the South of study area. These areas require prioritized mitigation strategies to reduce their vulnerability to windthrow (Fig. 5).
Fig. 5 - Prediction of the probability of windthrow from the Random Forest model.
Discussion
Regeneration pre- vs. post-disturbance
Pre-disturbance regeneration in beech forests (Faggete) showed top-heavy profiles dominated by taller class 3 saplings (795-1989 ind ha-1), reflecting episodic natural seed-based recruitment of shade-tolerant late-successional beech under canopy gaps. After disturbance at sites like Monte Nudo and Sasso del Ferro, pioneer Betula spp. (69% field cover, heights 76-154 cm) and early-successional Corylus avellana (22-36% cover, 350-390 cm) surged ahead of late-successional species like Fagus sylvatica (3-44% cover, shorter 30-88 cm), signaling a shift to faster-establishing species that stabilize soil and create microsites for future beech ingrowth. Chestnut stands (Castagneti) maintained balanced densities across classes via vegetative sprouting, but post-disturbance scenarios favor hazel dominance alongside invasives such as Robinia pseudoacacia (3% drone cover in SdF), potentially delaying chestnut recovery without intervention.
Antropogene stands pre-disturbance exhibited discontinuous, top-heavy regeneration from planted pulses, contrasting with post-disturbance reliance on birch and larch pioneers (up to 59% drone cover in MN) that fill gaps left by anthropogenic legacies. Aceri-frassineti/tiglieti displayed bottom-heavy small-sapling dominance pre-disturbance, aligning with shade-tolerant advance growth; after disturbance, however, invasive pioneers like Ailanthus altissima monopolize Scereda (91-100% cover, 300 cm height), suppressing natives and inverting expected late-successional pathways. Overall, disturbance accelerates pioneer shifts across types, with negative correlations between regeneration cover/height and deadwood occupancy (β = -0.56 to -0.71 in MN/SdF), indicating legacy constraints on native recovery (Tab. 3).
Deadwood pre- vs. post-disturbance
Pre-disturbance beech forests hosted rich, snag-dominated deadwood (18-2100 trees ha-1, 6-17 Mg ha-1 C), supporting high biodiversity, while chestnut stands emphasized large veteran snags (9-18 Mg ha-1) with scant downed CWD due to management. Post-disturbance, Monte Nudo conifer/mixed stands exhibit elevated variability (median 8 m3 ha-1, peaks >20 m3 ha-1) and occupancy that suppresses regeneration (GLM β = -1.14 for height), as dense legacies hinder pioneer establishment despite facilitating microsites in advanced decay (Tab. 4). Antropogene pre-disturbance sparsity (low snags/CWD) persists post-event, mirroring uniform MN mixed stands (~10 m3 ha-1), but with amplified patchiness that limits structural diversity.
“Aceri” deadwood balanced moderate snags (94-180 trees ha-1, 1-4 Mg C ha-1) and variable CWD pre-disturbance, yet SdF/Scereda broadleaves maintain 6-7 m3 ha-1 medians post-disturbance, where occupancy strongly curbs cover (β = -1.24) more than biomass, favoring invasives over natives. SEM confirms direct negative deadwood occupancy-regeneration links (β = -0.532), with biomass marginally shaping composition, suppressing spruce and larch, boosting birch, beech, and chestnut; (β=-0.059 - Fig. 2). Disturbance thus boosts deadwood volume 2-3× in variable sites (e.g., MN), inverting pre-disturbance patterns and constraining late-successional recovery unless decay advances to reduce occupancy barriers.
Deadwood occupancy limits regeneration cover and height
Our results illustrate early stages of secondary succession following a stand-replacing disturbance. The observed regeneration patterns fit the disturbance-response framework, in which pioneer species dominate early phases, but site legacies (e.g., deadwood, invasive presence) mediate the transition toward mature composition. The regeneration profiles across the three study sites, Monte Nudo, Sasso del Ferro, and Scereda, reveal distinct recovery trajectories and therefore require site-specific management responses. While SEM aggregated the dataset to test general pathways, site-specific GLMs and correlations (Tab. 3) reveal distinct local processes (e.g., strong physical obstruction at Monte Nudo vs. facilitative decay effects at Scereda). This multiscale approach reconciles local heterogeneity with overarching drivers. At Monte Nudo, the regeneration is dominated by pioneer birch and larch and robust patches of hazel; this pattern, combined with high variability in birch height and low larch cover, indicates an early-stage, spatially uneven recovery where pioneer species stabilize the substrate but do not yet form a uniform successional advance to late-successional taxa. The strong, negative association between deadwood occupancy and both regeneration cover and height at Monte Nudo implies that when fallen wood occupies soil surface area, it can substantially reduce available seedbed and growing space for seedlings (general model β for deadwood occupancy on cover and height - Tab. 4). This supports targeted interventions such as creating localized openings in dense deadwood patches or redistributing logs to preserve nurse-log microsites, while freeing ground space for seedling establishment ([5], [31]).
At Sasso del Ferro, the system appears transitional: pioneer birch and vigorous hazel co-occur with an increasingly widespread beech regeneration that is shorter than the faster-growing hazel. Although deadwood occupancy again shows a negative relationship with regeneration indices (albeit weaker than at Monte Nudo), our models suggest that deadwood effects can be species-specific: for beech, the negative effect was stronger on horizontal cover than on height (Beech-specific model). One plausible explanation is that beech establishes by lateral spread and clumped recruitment under partial canopy openings before committing to rapid height growth; therefore, deadwood that limits horizontal space will disproportionately reduce cover but not immediately reduce vertical stature, a mechanism proposed based on ecological strategy and our standardized-effect results ([8]).
Scereda is dominated by the invasive tree-of-heaven, which occupies all regenerating cover essentially and attains large sapling height. The near-absence of native early-successional species suggests competitive exclusion by the invasive and a strong seed rain/sprouting advantage for tree-of-heaven at this site. Management should prioritize invasive control (cut targeted herbicide or integrated methods reviewed by [28]) and combine this with selective deadwood retention/removal: retain or arrange some deadwood to maintain fungal and soil legacies while removing or redistributing obstructive log accumulations that are directly associated with reduced seedling ground cover in our models ([6], [17]).
Our analysis of windthrow vulnerability aligns closely with broader empirical and modelling studies; wind speed repeatedly emerges as the most critical driver of windthrow likelihood, followed by structural forest attributes (such as canopy height, height heterogeneity, and stand density), soil physical properties (including compaction and packing density), slope and topographic exposure, and species-specific rooting traits. These factors combine to determine mechanical stability and wind resistance at both tree- and stand-levels ([9], [24], [30]).
Contrasting our results with the broader literature highlights why deadwood may show both strongly positive and strongly negative effects on regeneration, depending on context. Numerous studies have documented beneficial microsite effects of coarse and downed woody debris, e.g., nurse-log effects that provide moisture, nutrient hotspots, and reduced competition, and experiments showing improved seedling survival and growth near logs under heat or drought stress ([5], [31]). Likewise, retention of deadwood buffers soil fungal communities and belowground functioning after disturbance ([6], [17]). These positive mechanisms are particularly important in water- or temperature-limited environments, where logs moderate extremes and conserve moisture, thereby promoting seedling establishment ([31]).
By contrast, our sites show net negative effects of deadwood occupancy on regeneration cover and height. Several hypotheses reconcile this apparent contradiction. First, the dominance of negative deadwood effects in our GLMs and SEM may reflect the occupancy measure (proportion of soil surface covered by logs), which directly reduces available seedbed area and physically hampers seedling establishment; this structural obstruction can outweigh facilitative microclimate benefits in systems where moisture and temperature are not strongly limiting. Second, pre-disturbance stand structure may co-vary with post-disturbance deadwood (i.e., dense pre-windthrow stands lead to more high log cover, meaning fewer pre-existing advance regeneration), therefore the observed correlations may partly capture legacy effects of stand density or composition rather than purely contemporary log-mediated microsite processes, and an alternative explanation consistent with literature on pre-disturbance legacies ([29]). Third, species-specific regeneration strategies will modulate responses: pioneer species such as birch can exploit disturbed, exposed mineral soil and may be less hindered by log cover than species that require fine seedbeds; conversely, species that recruit on nurse logs will benefit from retaining deadwood ([5], [6]). Our results show deadwood occupancy having a stronger standardized negative effect than deadwood biomass, which is consistent with the idea that spatial obstruction (occupancy) is more limiting to ground-covered sapling recruitment than the mere amount of woody material in the stand.
Management implications, therefore, need to be nuanced. Blanket salvage logging to remove all deadwood would likely harm soil fungal communities and long-term soil functioning ([6], [17]). Instead, a triage or heterogeneity-oriented approach is preferable: (i) remove or redistribute dense patches of surface-occupying logs that effectively eliminate seedbed availability, especially where pioneer recruitment is desired; (ii) retain or create nurse-log arrangements (or leave standing snags) where logs provide microclimate buffering that increases seedling survival in dry or heat-prone microsites ([31], experimental evidence); and (iii) in stands with problematic invasive recruitment (e.g., tree-of-heaven or black locust), pair invasive control with targeted deadwood manipulation to allow native seedlings to re-establish. The positive role of deadwood for fungal communities and nutrient cycling argues for selective retention rather than wholesale removal ([6], [17]).
Windthrow vulnerability assessment
Our windthrow-vulnerability analysis further informs prioritization of interventions. Wind speed, canopy height, and soil packing density emerged as primary predictors of gap occurrence in the random-forest model; this aligns with other modelling and empirical studies identifying wind exposure and stand structure as principal controls on windthrow risk ([30]) and agreeing with the critical environmental factors include wind speed and storm intensity; sustained winds of 95 km h-1 or higher, or gusts reaching 130 km h-1, substantially elevating the likelihood of windthrow by over 50% ([32]). However, some of the directions in our gap vs. non-gap comparisons require cautious interpretation. For instance, gaps in our data were associated with lower canopy height and lower height heterogeneity (Fig. 3), a pattern opposite to some expectations, as in the studies of Taylor et al. ([32]) and Forzieri et al. ([13]), shows that taller stands face greater bending stress, and dense, uniform plantations are more prone to windthrow than mixed, natural forests. Several factors could explain this: (i) selective removal of taller trees by past disturbances or logging leaving shorter remnant canopies where subsequent windthrow occurs; (ii) local site exposure interacting with stand architecture (e.g., short uniform stands on exposed ridges being more vulnerable than taller heterogeneous stands on sheltered slopes); or (iii) sampling scale effects where within-gap measures reflect remnant stumps and resprouts rather than pre-failure canopy structure. Therefore, we cannot verify which mechanism predominates without pre-disturbance tree-level data, especially the slenderness ratio between diameter at breast height and tree height as a principal parameter of tree vulnerability to windthrow.
Soil packing (bulk density) also showed an association with gap occurrence. Mechanistically, compacted soils restrict rooting depth and root development, reducing anchorage and increasing susceptibility to uprooting during storms ([9]). In-depth studies on the structural vulnerability of tree species such as Norway spruce (Picea abies) in Europe and balsam fir (Abies balsamea) in North America show that it is primarily due to shallow root systems and rigid stems ([32], [19]). This also suggests that soil physical status, including compaction from past human use or machinery, should be considered in risk-reduction planning. Finally, a landscape-level prioritization for windthrow mitigation should combine predicted vulnerability (random forest extrapolation) with regeneration and invasion status; zones predicted to be both vulnerable and lacking desirable native regeneration (or dominated by invasive recruits) demand special attention for active restoration.
Our analysis confirmed the central role of forest structure, but proxies of naturalness (height heterogeneity, mixed-species cover) also affected susceptibility, consistent with the idea that structural complexity confers mechanical buffering and adaptive capacity ([24], [1]). The limitation of the available data’s spatial resolution (10 m) also limits the model’s ability to detect fine-grained heterogeneity.
Limitations
The ability of Sentinel-2 imagery to capture windthrow-related features in structurally complex forests is inherently constrained by its spatial resolution (10-20 m), which limits the reliable detection of small or narrow canopy openings, fragmented deadwood patches, and early-stage regeneration beneath partially intact canopies. In heterogeneous stands, windthrow often results in fine-scale gap mosaics and irregular accumulations of coarse woody debris that may fall below the sensor’s effective detection threshold or be spectrally mixed with the surrounding intact canopy. As a consequence, Sentinel-2-based classifications are more sensitive to extensive, high-severity disturbances than to subtle structural damage or incipient regeneration processes. These limitations are further amplified by canopy complexity, where residual overstory trees and uneven vertical structures can obscure ground-level signals associated with deadwood and seedling establishment.
In this context, the use of supervised random forest classification introduces additional sources of uncertainty. While random forest models are robust to multicollinearity and non-linear relationships, their performance strongly depends on the quality and representativeness of the training data. Manual digitization of windthrow gaps, although necessary, introduces a degree of subjectivity in defining gap boundaries, particularly in transitional zones where partial canopy cover persists. This subjectivity, combined with spectrally heterogeneous forest cover, may lead to misclassification between lightly disturbed stands, shadowed areas, and intact forest. We therefore interpret the classification outputs as indicative of relative disturbance patterns rather than precise spatial delineations of windthrow gaps or deadwood distribution, and emphasize that higher-resolution imagery or LiDAR data would be required to fully resolve fine-scale disturbance and early regeneration dynamics.
Our observational design and the post-hoc nature of distinguishing pre- vs. post-windthrow regeneration mean that some effects attributed to deadwood could reflect pre-existing stand structure and seed-source limitations rather than solely log-mediated microsite processes. We did not separate advance regeneration (pre-disturbance) from post-disturbance recruits at all sites; future work should measure age cohorts (e.g., bud-scale, whorl counts, or dendro methods) to disentangle pre-existing regeneration from post-windthrow establishment. Our data represent early post-disturbance trajectories (4-5 years after Storm Alex), when regeneration signals primarily reflect immediate microsite conditions rather than full successional reorganization. Longer-term monitoring is needed to assess whether the observed negative effects of deadwood occupancy persist or diminish as decomposition advances. Also, where we recommend partial deadwood removal/redistribution, experimental manipulations (salvage logging treatments, controlled log redistribution, paired invasive-control interventions) would provide stronger causal evidence for optimal management prescriptions.
Management implications
Overall, our results indicate that deadwood is neither unambiguously harmful nor purely beneficial. Its net effect depends on spatial occupancy, species-specific regeneration strategies, the presence of invasions, and local site constraints (moisture, temperature, soil compaction). A management approach that promotes spatial heterogeneity (retaining some deadwood and removing obstructive patches) is most consistent with both fostering regeneration and preserving belowground ecosystem functioning ([6], [17], [31]).
The findings from this research are critical for informing habitat conservation strategies before and after extreme climate events, using cost-effective remotely sensed input. By evaluating the current state of forest recovery, the study will help identify the role of windthrow-induced deadwood in regeneration processes. Additionally, the evaluation of environmental, topographical, and structural drivers of windthrow vulnerability will provide a workflow for wind risk assessment across forests, highlighting priority areas for targeted preventive actions. These insights will support evidence-based conservation planning, ensuring long-term ecological stability and the sustained provision of critical ecosystem services that forests offer amid increasing climatic uncertainties.
Our results suggest that post-windthrow management should move beyond uniform salvage or non-intervention strategies and instead adopt context-dependent approaches that account for deadwood spatial configuration, forest structure, and regeneration trajectories. In particular, the strong negative relationship between deadwood occupancy and early regeneration indicates that excessive surface coverage by coarse woody debris can hinder seedling establishment, especially in sites dominated by even-aged stands and limited microsite heterogeneity. This supports management strategies that retain deadwood while avoiding continuous, high-coverage accumulations, for example, through partial redistribution rather than complete removal.
At the same time, the observed variability among sites highlights that deadwood cannot be treated as a universally negative or positive legacy. In more structurally complex stands, deadwood may contribute to long-term resilience by enhancing habitat diversity and future microhabitat availability, even if short-term regeneration is locally constrained. This implies that selective retention of deadwood, coupled with targeted interventions to create regeneration niches, may reconcile biodiversity conservation goals with forest recovery objectives.
The windthrow susceptibility analysis further indicates that forests characterized by high canopy height and structural homogeneity are disproportionately vulnerable to extreme wind events. From a preventative perspective, this underscores the importance of promoting structural diversity and mixed-species stands as part of long-term forest planning, particularly under increasing disturbance regimes linked to climate change. Whether such structural diversification consistently reduces wind damage across broader spatial and temporal scales remains to be tested with long-term datasets.
Overall, our findings advocate for adaptive, site-specific management that integrates disturbance legacies into restoration planning, rather than relying on fixed post-disturbance prescriptions. Continued monitoring is essential to determine whether early negative effects of deadwood on regeneration persist or transition into facilitative roles as decomposition progresses.
Conclusions
This study provides valuable insights into the regeneration dynamics of wind-damaged forests and the key environmental, structural, and topographical factors influencing windthrow vulnerability across three sites in the Pre-Alps of Varese (Italy): Monte Nudo, Sasso del Ferro, and Scereda. Our findings highlight that regeneration success varies across sites, with distinct species compositions and responses to disturbance legacies. While Monte Nudo exhibits early-successional patterns dominated by birch and European larch, Sasso del Ferro demonstrates a transition towards a more stable forest structure, and Scereda faces challenges due to the dominance of the invasive Ailanthus altissima. These variations emphasize the need for site-specific management strategies to promote successful forest recovery.
A critical factor affecting regeneration across all sites is deadwood accumulation, which negatively influences regeneration cover and height by limiting available space and altering microclimatic conditions. The results from both the Gamma GLM and SEM models confirm that excessive deadwood occupancy suppresses regeneration, though its effects vary among species. European beech exhibits sensitivity in lateral expansion due to deadwood occupancy, whereas Betula spp. shows a more balanced response in both height and coverage. These findings highlight the complexity of deadwood’s role in post-windthrow dynamics and reinforce the need for adaptive deadwood management to optimize species-specific regeneration processes.
Beyond regeneration, our study underscores the drivers of forest vulnerability to windthrow identified by the Random Forest model. Wind speed remains the most influential factor, followed by canopy height and topographical features such as slope inclination and aspect. Soil characteristics, particularly packing density, also play a crucial role in determining forest stability. Additionally, species composition and structural heterogeneity influence windthrow susceptibility, with pure stands of Norway spruce and forests with high broadleaf exposure during leaf-on periods being more vulnerable. These insights stress the importance of integrating structural and compositional factors into windthrow risk assessments.
From a conservation perspective, these results provide a decision-making framework for enhancing forest resilience before and after disturbance events. Targeted actions, such as regulating deadwood levels, controlling invasive species, and promoting wind-resistant species mixtures, can strengthen recovery capacity. Moreover, spatially explicit vulnerability maps can help prioritize interventions both within Natura 2000 sites and across the broader landscape. Embedding these measures into adaptive management will support ecosystem stability, safeguard biodiversity, and sustain forest services under intensifying climate-related disturbances.
Acknowledgements
We thank Susanna Galimberti for assistance with fieldwork, Sebastian Brocco for technical support in drone survey planning, and Federico Romanato for his valuable comments on the manuscript. This work was funded by Comunità Montana Valli del Verbano within the BioClima project.
Authors’ contribution
Conceptualization: BTU, GV; Methodology: BTU, GV; Formal analysis: BTU, Visualization: BTU, GV; Writing - original draft: BTU; Writing - review & editing: BTU, GV; Supervision: GV; Funding acquisition: GV.
References
Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Authors’ Info
Authors’ Affiliation
Giorgio Vacchiano 0000-0001-8100-0659
Forest Lab, Department of Agricultural and Environmental Sciences, University of Milan (Italy)
Corresponding author
Paper Info
Citation
Bui TU, Vacchiano G (2026). Assessing forest resilience after windthrow: linking deadwood, regeneration, and landscape-scale susceptibility - A case study in a Pre-Alpine forest in Italy. iForest 19: 102-113. - doi: 10.3832/ifor4982-019
Academic Editor
Marco Borghetti
Paper history
Received: Sep 09, 2025
Accepted: Feb 01, 2026
First online: Apr 07, 2026
Publication Date: Apr 30, 2026
Publication Time: 2.17 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2026
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 1887
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 362
Abstract Page Views: 734
PDF Downloads: 731
Citation/Reference Downloads: 0
XML Downloads: 60
Web Metrics
Days since publication: 96
Overall contacts: 1887
Avg. contacts per week: 137.59
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
(No citations were found up to date. Please come back later)
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Review Papers
Remote sensing-supported vegetation parameters for regional climate models: a brief review
vol. 3, pp. 98-101 (online: 15 July 2010)
Review Papers
Remote sensing support for post fire forest management
vol. 1, pp. 6-12 (online: 28 February 2008)
Review Papers
Accuracy of determining specific parameters of the urban forest using remote sensing
vol. 12, pp. 498-510 (online: 02 December 2019)
Review Papers
Remote sensing of selective logging in tropical forests: current state and future directions
vol. 13, pp. 286-300 (online: 10 July 2020)
Technical Reports
Detecting tree water deficit by very low altitude remote sensing
vol. 10, pp. 215-219 (online: 11 February 2017)
Research Articles
Assessing water quality by remote sensing in small lakes: the case study of Monticchio lakes in southern Italy
vol. 2, pp. 154-161 (online: 30 July 2009)
Research Articles
Afforestation monitoring through automatic analysis of 36-years Landsat Best Available Composites
vol. 15, pp. 220-228 (online: 12 July 2022)
Technical Reports
Remote sensing of american maple in alluvial forests: a case study in an island complex of the Loire valley (France)
vol. 13, pp. 409-416 (online: 16 September 2020)
Research Articles
Geostatistical techniques for estimating aboveground biomass in eastern Amazonia
vol. 19, pp. 85-93 (online: 13 March 2026)
Research Articles
Evaluation of hydrological and erosive effects at the basin scale in relation to the severity of forest fires
vol. 12, pp. 427-434 (online: 01 September 2019)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







