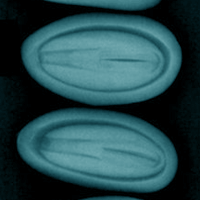
Measuring the damage of Leptoglossus occidentalis in Stone pine seeds in a pine nut production hot spot in Turkey
iForest - Biogeosciences and Forestry, Volume 19, Issue 3, Pages 201-208 (2026)
doi: https://doi.org/10.3832/ifor4854-018
Published: Jun 04, 2026 - Copyright © 2026 SISEF
Research Articles
Abstract
The western conifer seed bug (Leptoglossus occidentalis Heidemann, 1910) is an invasive alien species from North America that causes significant economic losses by damaging cones and reducing seed yield in pines. The pest was first detected in Turkey in 2009, coinciding with increasing seed losses in many conifers, particularly Stone pine, Pinus pinea L. This study aimed to assess the impact of L. occidentalis on seed loss in three-year-old P. pinea cones. Conducted in 2020-2021 in P. pinea stands of the Izmir/Bergama Kozak Plateau, the experiment involved 5103 seeds collected from 30 cones, both caged (excluded) and uncaged (exposed), on 15 trees over two years in a natural setting. The seeds were first categorized by damage level using X-ray imaging, then subjected to a flotation test to determine the floating and sinking ratios. Subsequently, the kernels were manually examined to classify damage, and their weights were measured. The results showed that seed loss in excluded cones was significantly lower than in exposed cones, highlighting the impact of a factor that cannot reach excluded cones, with L. occidentalis as the most likely cause. X-ray, seed flotation, and naked-eye methods yielded similar results in assessing overall seed loss. Seed damage rates in the exclusion treatments were 17.6%, 17.0%, and 17.0%, respectively, whereas in the exposition treatments they were 46.2%, 48.0%, and 53.0%, respectively. As of 2021, these levels of seed loss corresponded to an estimated economic loss of approximately €55 per kg of seed. When damage levels were categorized, X-ray and naked-eye assessments produced inconsistent results. These findings suggest that X-ray, seed flotation, and naked-eye methods can be used interchangeably to determine overall seed damage. However, the naked-eye method appears to be more suitable for categorizing damage levels.
Keywords
Damage Categorization, Pinus pinea L., Western Conifer Seed Bug, Seed Loss, X-ray
Introduction
Seed loss in conifer species can result from various factors, including pollination failure, fertilization issues, seed development defects, climatic conditions, fungal infections, and damage caused by cone- or seed-feeding insects, either individually or in combination ([44], [41], [20]). While it is often difficult to isolate the specific effects of these factors in natural environments, experimental studies have demonstrated that seed insects can cause significant seed losses ([23], [10]).
Due to the high economic value of its seeds, seed loss in Stone pine (Pinus pinea L.) has been a frequently studied topic ([23]). Turkey, along with Italy, Portugal, and Spain, accounts for 94% of the world’s Stone pine stands ([41]). In Turkey, the area of Stone pine forests increased from 89,028 ha in 2012 to 174,596 ha in 2023 ([42]). The pine nut trade is a vital source of income for many rural communities in western Turkey, where pine nut production has declined significantly over the years. Production dropped from an average of 4,385 tons per year between 2005 and 2011 to 1.916 tons per year between 2012 and 2023 ([2]). The key pine nut production regions, including Aydin-Koçarli and Izmir-Bergama’s Kozak Plateau, contribute almost 80% of the country’s total yield ([30]). Notably, in Bergama, Izmir, seed loss reached up to 70% between 2013 and 2017 ([43]).
The western conifer seed bug, Leptoglossus occidentalis Heidemann (1910) (Hemiptera: Coreidae), is a significant insect species native to North America, known for its damage to coniferous tree seeds. Within its natural range, it is one of the primary economic pests in seed orchards, causing considerable seed loss ([31], [5], [11], [23]). The extent of damage caused by L. occidentalis varies depending on several factors, including the host species ([17]), the developmental stage of the bug ([27]) and the cone at which feeding occurs ([50]), the population density of the bug ([24]), and whether the host species is mixed with none-host species ([25]).
Previous studies have reported varying levels of seed loss due to L. occidentalis across different conifer species: 1% in Picea abies ([36]), 2.1% in Pinus albicaulis, 70% in P. contorta ([52]), 75% in P. monticola ([7]), 77.36% in P. nigra, 55% in P. ponderosa ([33], [46]), 53.55% in P. sylvestris ([36]), and between 5% and 50% in Pseudotsuga menziesii ([50], [8]). In Mediterranean regions, studies conducted across different countries and years have estimated seed loss due to L. occidentalis in P. pinea at around 50% ([14], [24]).
Since P. pinea cones take three years to develop, the period of exposure to seed and cone pests is quite long. As a result, damage caused by L. occidentalis, which can produce multiple generations under favorable conditions ([15], [54], [29], [4]), can be severe, making it one of the most significant seed pests of Mediterranean pines ([11]). At the western end of the Mediterranean, Farinha et al. ([24]) (and references therein) have previously demonstrated the relationship between seed loss in P. pinea and L. occidentalis. Moreover, it has been suggested that the bug’s impact is exacerbated by climate change ([20], [14]). At the eastern end of the Mediterranean, research on the effects of climate change on P. pinea has shown that a significant limitation of radial growth has been incurred due to drought over the last 30 years, which may partly explain the recent decline in pine nut yield in the region ([32]). However, the extent of L. occidentalis’s impact, first reported in Turkey in 2009 ([3], [26]), remains unclear. The insect has spread rapidly across the country in a relatively short time ([28], [4]), but how much of the observed seed loss is attributable specifically to L. occidentalis remains to be determined. Understanding this impact is crucial to justify the need for insect control measures.
Direct observation of bug damage is challenging. Its feeding activity does not leave visible traces on cones (see [34], [35] for detectable traces of L. occidentalis), no effective trapping method currently exists, and observing bug activity in the canopy is difficult due to tree height ([24]). Consequently, further studies are needed to confirm the extent of bug damage. Radiographic analysis of harvested seeds has been successfully used to classify seed damage in Douglas fir ([5]) and P. pinea ([22]). However, this method alone cannot differentiate between seeds emptied by seed bug feeding and naturally unfilled (aborted) seeds ([6]). When combined with controlled bagging experiments, radiographic methods can facilitate the rapid detection of seed damage ([22]).
The aim of this study was to quantify seed loss caused by L. occidentalis in the Bergama Kozak Plateau, one of Turkey’s two major pine nut production hotspots. Specifically, we aimed to: (1) determine the seed loss in P. pinea in Bergama, (2) compare the effects of excluding and exposing P. pinea seeds to L. occidentalis damage, and (3) evaluate the reliability of radiographic analysis in assessing seed damage levels.
Material and methods
Study site
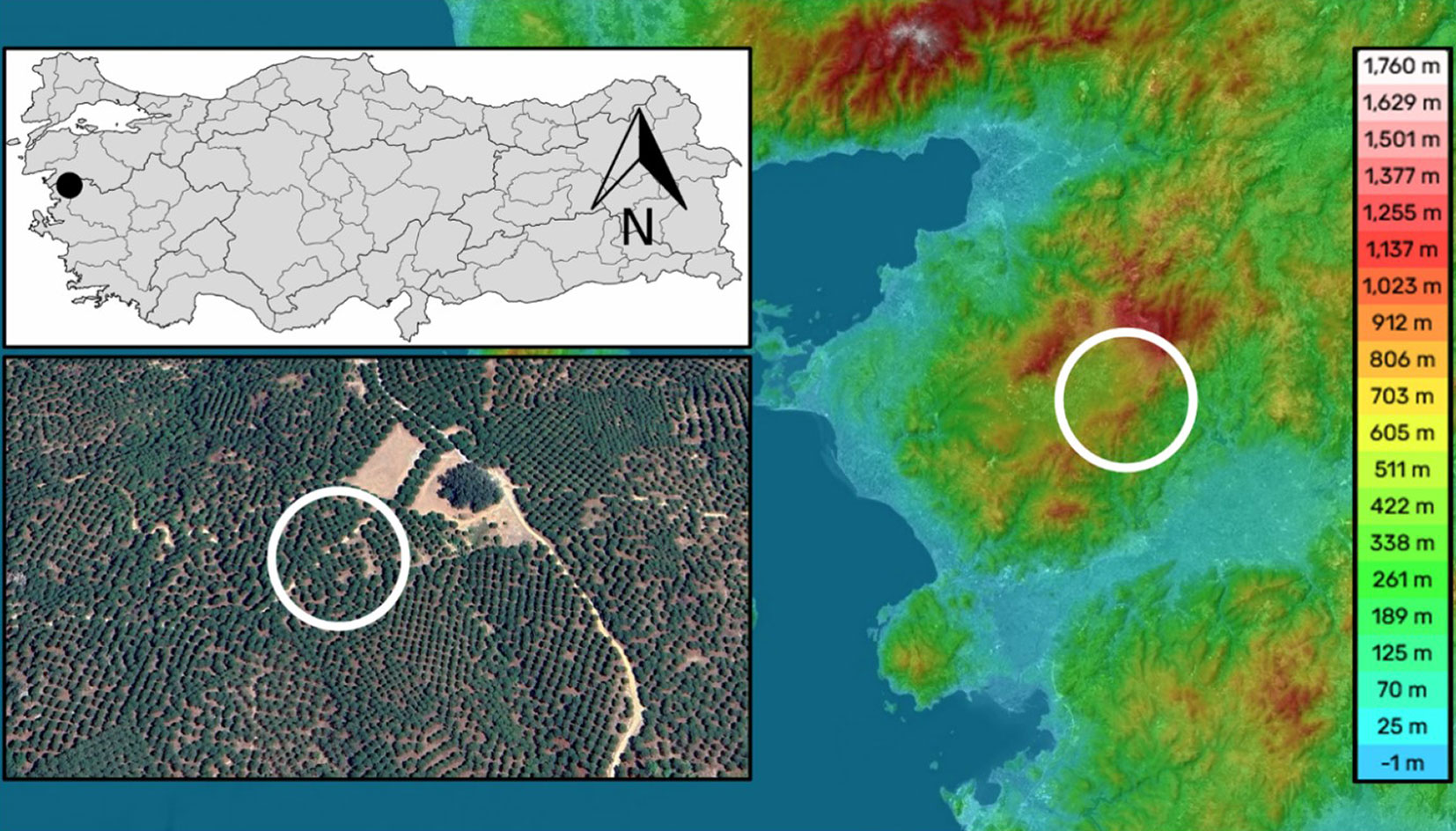
The study area is located within the borders of Karaveliler Village in the Bergama Kozak Plateau of Izmir Province, Turkey (Fig. 1). This location was selected because it is the largest habitat of P. pinea and has the highest pine nut production in the country. The site is situated at an elevation of 787 m a.s.l., with central coordinates of 39° 15′ 23″ N, 27° 08′ 42″ E (Fig. 1). The climate is typically Mediterranean, characterized by mild, wet winters and hot, dry summers. The total annual rainfall is 720 mm, and the average annual temperature is 16.0 °C. The site was established through planting in 1997 and is owned by the Karaveliler village community. The Stone pine trees within the study area have diameters ranging from 36 to 52 cm and heights of 7 to 8 m. The forest has a general coverage of 80-90% and is surrounded by P. brutia forests.
Fig. 1 - Location of the study site, Bergama Kozak Plateau, Izmir, Turkey.
Experimental design
The experiment was conducted over two consecutive years, 2020 and 2021. In May 2020, we selected and marked 15 trees with red spray paint, ensuring each tree bore at least six branches with third-year cones. A hydraulic lift mounted on a truck was used to provide access (Fig. 2a). On each tree, we selected two third-year cones and marked them with red and white hazard tape. We selected the cones at the same height and aspect to create a homogeneous trial as much as possible. One of the selected cones per tree was enclosed in a protective bag (hereafter: “Exc”, short for “exclusion treatment”) to prevent insect damage (Fig. 2b). The bag, made of 0.5 mm mesh and measuring 35-60 cm in length depending on the branch size, covered the cone-bearing branch from the fork base to the tip, allowing room for growth. The wire supports inside the bags kept the cones from touching the mesh, preventing insect feeding from the outside. The other selected cone per tree was left exposed (hereafter: “Exp”, short for “exposition treatment”) to allow natural insect interaction (Fig. 2c). The experiment continued until December 2020, the pine nut harvesting period in Turkey, when all marked cones were collected and transferred to the laboratory for further analysis. The same experiment was repeated from June to November 2021.
Fig. 2 - Experimental design.

All sampled third-year cones were dried in an incubator at 40-55 °C in the laboratory until they opened (Fig. 2d). We then counted the number of seeds per cone. To assess seed health status, we employed the following methods:
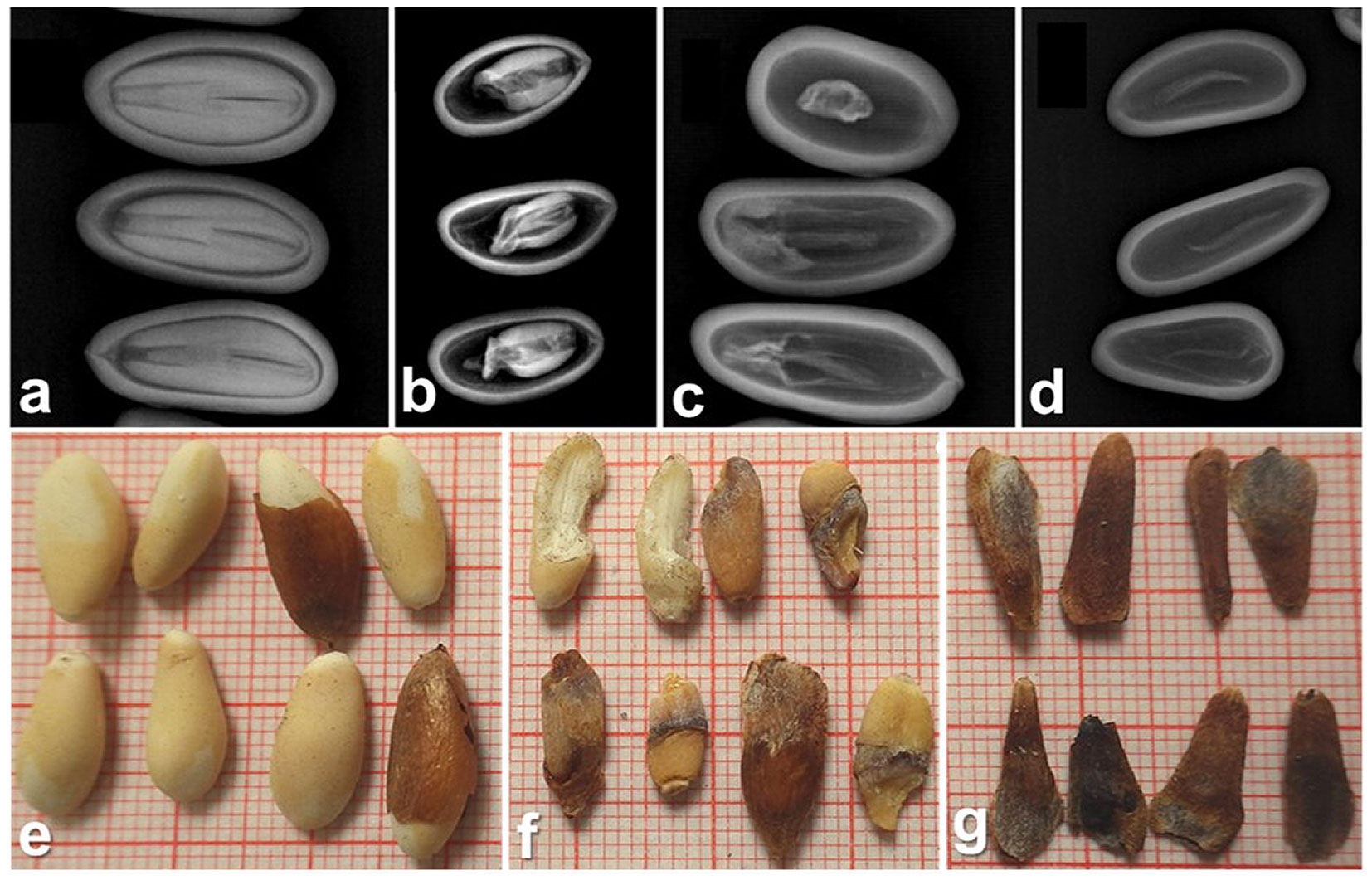
- X-ray evaluation of seeds - we randomly selected 25 seeds per cone from 15 trees over two years, for a total of 1,475 seeds. X-ray images were taken using a Siemens Mammomat Inspiration imager (32 kV, 50 mAs, print range: 1 cm - Fig. 2e, Fig. 2f). Each seed was visually scored from x-ray images for damage based on Farinha et al. ([20]) (Farinha’s scale hereafter), categorizing them as: (i) “Viable”: intact kernel (Fig. 3a, Fig. 3e); (ii) “Type-I”: partially damaged kernel (Fig. 3b, Fig. 3f); (iii) “Type-II”: dry embryo with no endosperm (Fig. 3c, Fig. 3g); (iv) “Type-III”: empty shell without an embryo (Fig. 3d).
- Seed floating test - we tested the buoyancy of 2.516 Exc and 2.587 Exp seeds by floating them in water and recording how many seeds floated or sank.
- Naked-eye evaluation of kernels - we cracked open all 5.103 seeds, extracted the kernels, and classified them visually into damage categories using Farinha’s scale (Fig. 3e-Fig. 3g).
- Kernel weighing - after visual examination, we weighed the kernels with 0.1 g precision (Fig. 2g).
Fig. 3 - Seed damage categories according to Farinha’s scale obtained from X-ray image evaluation: (a) Viable; (b) Type-I; (c) Type-II; (d) Type III; and from naked-eye evaluation: (e) Viable; (f) Type-I; (g) Type-II (Type-III is not included as this category has no kernel).
Statistical analysis
To test differences in seed health status between the two treatments, Exc and Exp, we used the Student’s t-test or the Mann-Whitney test based on the data’s normality, with a statistical significance level of 0.001 (p ≤ 0.001). Tested variables were the scores for damage visualization by X-ray and naked eye, the number of seeds that floated and sank, and seed weight. The statistical analysis was performed in R ([48]).
Results
Stone pine seed damage in Bergama
Seed yield was 2.117 seeds in 2020 (approximately 71 seeds per cone) and 2.986 seeds in 2021 (approximately 100 seeds per cone), based on 30 cones sampled from the same 15 trees each year. Despite differences in total seed counts, no significant difference was found between the two years in the percentage of viable seeds (t = 0.013, df = 28, p > 0.05). However, a statistically significant difference was observed between the percentage of viable seeds of bug-protected Exc and unprotected Exp (t = -6.485, df = 40, p < 0.05). In the Exc treatment, the percentage of viable seeds ranged from 82.4% to 83.0%, whereas in the Exp treatment, it ranged from 47.0% to 53.8% (Tab. S1 in Supplementary material). Accordingly, seed damage ranged from 17.0% to 17.6% in Exc and from 46.2% to 53.0% in Exp. Thus, excluding P. pinea seeds from insect damage significantly increased the number of healthy seeds.
Comparative evaluation of methods for assessing seed damage
Since no statistically significant differences were found between 2020 and 2021 across X-ray, naked-eye, seed weight, or flotation trials, the data from both years were combined for further analysis.
X-ray image evaluation of seeds
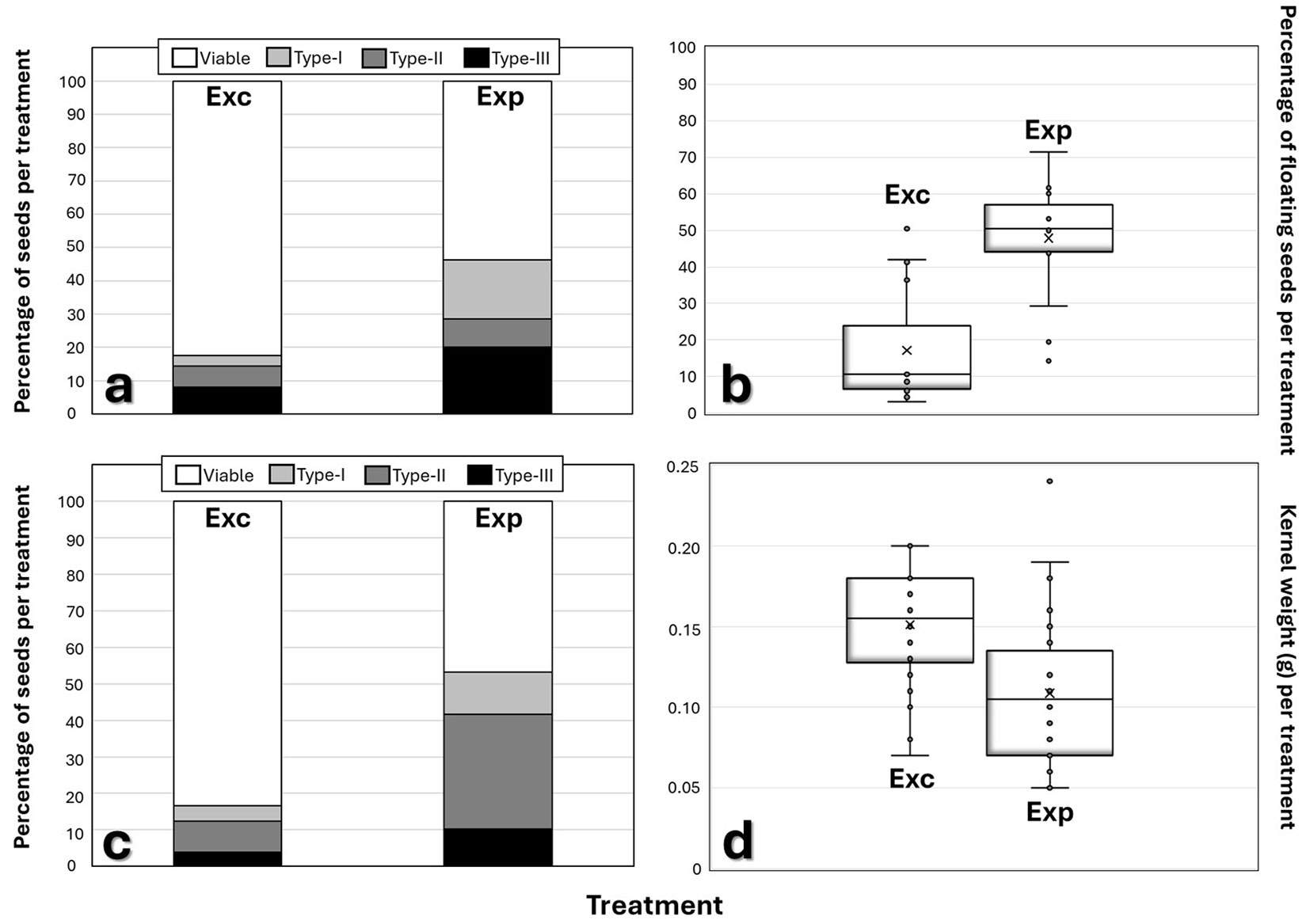
The percentage of seeds per treatment in the Viable category in Exc (82.4%) was significantly higher (p < 0.05) than in Exp (53.8% - Fig. 4a, Tab. S1). The percentage of seeds per treatment categorized as Type-I in Exc (3.1%) was significantly lower (p < 0.05) than in Exp (17.6% - Fig. 4a, Tab. S1). Although there was no statistically significant difference (p = 0.10) in the number of Type-II seeds between Exc and Exp, the percentage of Type-II seeds was lower in Exc (6.4%) compared to Exp (8.6% - Fig. 4a, Tab. S1). Similarly, the percentage of Type-III seeds in Exc (8.0%) was significantly lower (p < 0.05) than in Exp (20.0% - Fig. 4a, Tab. S1). Overall, seed damage in Exc was lower than in Exp, with the most notable difference observed in the Type-I damage class ( Fig. 4a, Tab. S1).
Fig. 4 - Results of (a) X-ray image evaluation of seeds, (b) seed floating test, (c) naked-eye evaluation of kernels, and (d) kernel weighing in each treatment (Exc and Exp).
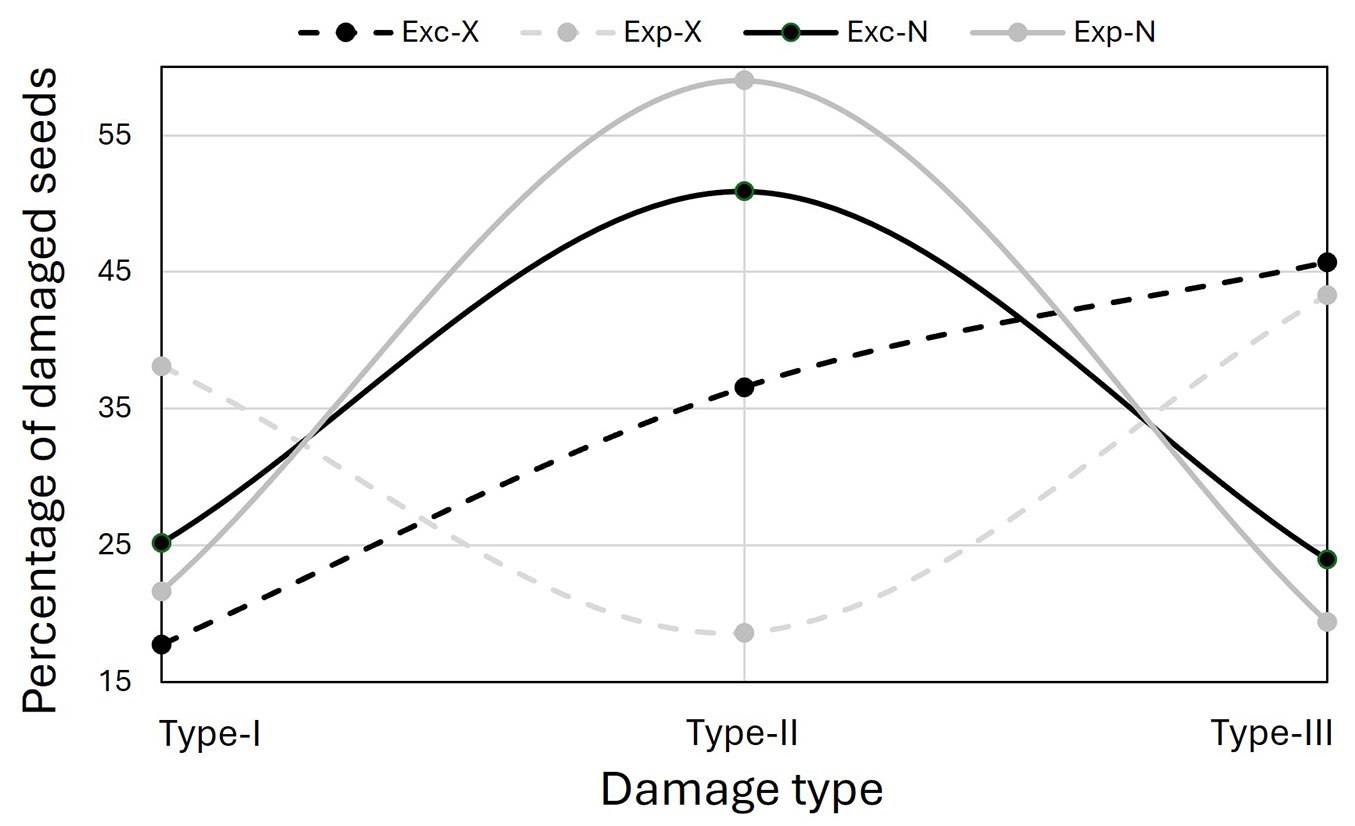
When only the damaged seeds were considered, 45.7% of all damaged seeds in Exc fell into Type-III, 36.6% into Type-II, and 17.7% into Type-I. In Exp, 43.3% of all damaged seeds fell into Type-III, 38.1% into Type-I, and 18.6% into Type-II (Fig. 5).
Fig. 5 - Partitioning of damaged seeds in each treatment (Exc and Exp) into damage categories of Farinha’s scale according to each evaluation method (X: X-ray; N: naked-eye).
Seed floating test
The percentage of floating seeds per treatment in Exc (17%) was lower than in Exp (48%), and this difference was statistically significant (p < 0.05; Fig. 4b).
Naked-eye evaluation of kernels
The percentage of seeds per treatment in the Viable category in Exc (83.0%) was significantly higher (p < 0.05) than in Exp (47.0% - Fig. 4c, Tab. S1). The percentages of seeds categorized as Type-I, Type-II, and Type-III in Exc (4.2%, 8.5%, and 4.0%, respectively) were significantly lower (p < 0.05) than in Exp (11.5%, 31.4%, and 10.3%, respectively - Fig. 4c, Tab. S1). Overall, seed damage in Exc was lower than in Exp, with the most notable difference observed in the Type-II damage class (Fig. 4c, Tab. S1).
When only the damaged seeds were considered, 50.9% of all damaged seeds in Exc fell into Type-II, 25.1% into Type-I, and 24.0% into Type-III. In Exp, 59.0% of all damaged seeds fell into Type-II, 21.6% into Type-I, and 19.4% into Type-III (Fig. 5).
Kernel weighing
Mean kernel weight per treatment in Exc (0.15 g) was higher than that in Exp (0.11 g), and this difference was statistically significant (p < 0.05 - Fig. 4d).
Discussion
Seed damage in P. pinea
The seed loss in our study plot in Bergama, Izmir, in 2020 and 2021 was between 46-53% in cones that were not bagged. Calama et al. ([14]) found that seed loss in P. pinea in Spain was around 50%. Farinha et al. ([24]) reported that seed loss in Portugal varied significantly from year to year, ranging from 2% to 50%. Ponce-Herrero et al. ([47]) reported a seed loss rate of 35.6% in Spain. Differences in seed loss rates across studies may be due to fluctuations in L. occidentalis population size ([24]) and various uncontrolled factors affecting seed yield, such as temperature, drought, stand composition (pure vs. mixed, in line with the theory of biotic resistance - [25]), as well as tree genotype and age. Without accounting for these factors, comparing seed loss rates across years and locations may be of limited significance.
Impact of L. occidentalis on seed damage in P. pinea
Exclusion experiments are a crucial method for demonstrating the impact of insects on seed yield ([53], [51]). In this study, seed loss in exposed cones ranged from 46% to 53%, whereas in excluded cones, it ranged from 16.6% to 17%. Likewise, the percentage of viable seeds was significantly higher in excluded cones, while the proportion of damage across all damage classes (Type-I, Type-II, and Type-III) was lower. Indirect methods such as X-ray and seed flotation provided results consistent with the more direct naked-eye assessments, suggesting they can serve as proxies when direct evaluation is not feasible. Collectively, these results strongly suggest that the primary cause of low cone yield in Bergama-Izmir is an agent that is prevented from accessing the excluded cones, most likely L. occidentalis.
Besides L. occidentalis, Camptomyia pinicola Mamaev, 1971 (Diptera: Cecidomyiidae), Dioryctria pinea (Staudinger, 1859) (Lepidoptera: Pyralidae), and Ernobius pini (Sturm, 1837) (Coleoptera: Anobiidae) are also known to feed on P. pinea cones in the study region ([45]). However, the damage patterns of these species differ significantly from those of L. occidentalis, and they can hardly be confused. No damage by these species was found on cones left in the exposition treatments. Moreover, Strong et al. ([52]) and Strong ([51]) have shown that the hypothesis that cone closure can change microclimatic conditions in favor of the seed and positively affect seed health is incorrect. All this strongly suggests that the most important reason for the high seed yield in the exclusion treatment is the exclusion of L. occidentalis.
Based on the seed loss (ca. 40-60%) observed in the present study, the potential economic impact in Turkey was estimated at approximately €55 per kg (April 2021 values), highlighting the potential economic significance of the damage. On the other hand, the actual impact is likely to be considerably greater than this estimate. Although this study demonstrated the damage caused by L. occidentalis on 3-year-old cones, it likely underestimates the impact of abortion in 1- and 2-year-old cones induced by L. occidentalis ([40]), which was not accounted for in the present study. Moreover, this study does not capture fluctuations in L. occidentalis populations or their impact on kernel yield, as shown by Farinha et al. ([24]). While these questions have been explored across the Mediterranean, from Lebanon to Portugal ([19], [14], [18], [24]), further research is needed in Turkey’s primary pine nut production region to assess these factors under its specific climatic and vegetation conditions.
Impact of other factors on seed loss in P. pinea
In their exclusion experiment, Farinha et al. ([24]) found that the percentage of viable seeds in exposed cones ranged from 70% to 86%, while in excluded cones, it averaged 92%. Other studies reported similar rates, with 94% viable seeds in excluded cones ([19], [21]). We found that the percentage of viable seeds in excluded cones (ca. 83%) was lower than in exposed cones (ca. 50%). These results align with those of Ponce-Herrero et al. ([47]), who reported 73.6% viable seeds in excluded cones and 35.6% in exposed cones in their exclusion experiment on P. pinea in Valladolid, Spain. The lower percentage of intact seeds in excluded cones in our study, compared with other studies, may be due to climatic factors, such as drought, which can reduce overall seed yield. Spring and summer droughts in the Mediterranean region, exacerbated by climate change, have been suggested as key contributors to the recent decline in pine nut yield ([41], [21]). Indeed, Stone pine is known for its drought resistance. However, trees in arid sites tend to have a lower leaf area index and reduced cone production compared to those grown under irrigation ([9], [12], [13], [16], [37], [38]). Köse et al. ([32]) reported a significant increase in winter temperatures and drought in the Kozak Plateau-Bergama, particularly after the late 1990s. Thus, climate-related factors, mainly high air temperatures and drought, appear to contribute significantly to seed loss in P. pinea.
Categorization of seed damage in P. pinea and reliability of X-ray images
Among the four evaluation methods used to assess and categorize damage levels in P. pinea kernels, naked-eye evaluation and kernel weighing were direct methods focused on kernels, whereas X-ray evaluation and seed floating were indirect methods focused on seeds. Kernel weight is a variable characteristic influenced by locality and growing year ([1]). Although kernel weighing effectively distinguished between exclusion and exposition treatments, it may not reliably categorize damage without these treatments unless the average kernel weight for the specific locality and year is known. The other direct method, naked-eye evaluation, can be considered a reference method due to its low error rate, as it involves the direct and individual assessment of each kernel. Using this method, we found that approximately 17% and 53% of seeds were damaged in exclusion and exposition treatments, respectively. The indirect methods, X-ray evaluation and seed floating, yielded damage ratios similar to naked-eye evaluation: around 18% and 17% in the exclusion treatment, and approximately 46% and 48% in the exposition treatment, respectively. These findings suggest that naked-eye evaluation, X-ray evaluation, and seed floating can be used interchangeably to determine overall kernel damage. However, while the seed floating test can indicate total damage, it cannot categorize damage types, unlike the naked-eye and X-ray methods. Notably, there was a discrepancy between the latter two methods when categorizing damage according to Farinha’s scale, as detailed below.
Type-I: Farinha et al. ([20]), using a combination of X-ray and naked-eye methods (though without directly comparing them), reported that Type-I damage was nearly absent in excluded cones and approximately 7% in those exposed. Similarly, Elvira-Recuenco et al. ([19]), using naked-eye evaluation, found no Type-I damage in excluded cones. However, our findings differed, as we detected Type-I damage in both treatments, with higher levels in exposed cones than in those excluded. Among the damaged seeds, X-ray analysis revealed that approximately 18% of seeds in protected cones and 38% in exposed cones exhibited Type-I damage. In contrast, the naked-eye evaluation identified around 25% and 22%, respectively, significantly different from the percentages observed in the X-ray evaluation.
Type-II: Ponce-Herrero et al. ([47]), using naked-eye evaluation, reported Type-II damage rates of 4.6% in excluded cones and 20.6% in exposed. Our study found similar results with the naked-eye evaluation, identifying Type-II damage in 8.5% of excluded cones and 31.4% of exposed cones. However, in the X-ray evaluation, these rates were 6.4% in excluded cones and 8.6% in exposed cones, with no statistically significant difference between them. Among the damaged seeds, approximately 37% in excluded cones and 19% in exposed cones were categorized as Type-II damage. This suggests that Type-II damage was the least common type among damaged seeds in exposed cones but the second most common in excluded cones. In contrast, the naked-eye evaluation indicated that Type-II damage was the most frequent damage type in both excluded and exposed cones, with rates of 51% and 59%, respectively. A similar result was reported by Farinha et al. ([20]), who suggested that the high prevalence of this damage type in protected cones could be attributed to climatic, physiological, or other biotic factors affecting cone development in its early stages.
Type-III: Farinha et al. ([20]) and Elvira-Recuenco et al. ([19]) reported no significant difference in Type-III damage between excluded and exposed cones, concluding that this damage type was unrelated to L. occidentalis and likely linked to fertilization issues. However, our study found a significant reduction in Type-III damage in excluded cones, decreasing from 20% to 8% in the X-ray evaluation and from 10.3% to 4% in the naked-eye evaluation. These findings align with those of Ponce-Herrero et al. ([47]), who reported Type-III damage rates of 14% in excluded cones and 19% in exposed cones. Similar patterns have also been observed in P. ponderosa and P. menziesii ([46], [49]). According to the X-ray evaluation, Type-III damage was the most common damage type in both excluded and exposed cones. In contrast, the naked-eye evaluation identified it as the least common type of damage in both cases. This discrepancy further highlights the differences between the two evaluation methods in how they categorize damage types.
Our findings revealed different distributions of damage types between X-ray and naked-eye evaluations, despite no significant difference in the percentage of viable seeds identified by either method. This discrepancy may be due to sampling errors since the naked-eye assessment was performed on the entire seed sample (2.986 seeds), while the X-ray evaluation was conducted on a randomly selected subset (1.475 seeds). Alternatively, it may indicate that radiographic methods are not entirely reliable for detailed damage classification. For example, the method may overestimate the proportion of Type-III seeds, as Type-II seeds are prone to misclassification due to limitations in detecting subtle internal opacity differences that can only be discerned through naked-eye inspection. X-ray damage assessment has been successfully applied in various conifer species ([5], [36], [20], [21]). This method allows for rapid damage diagnosis; however, previous studies have reported that it may underestimate damage ([21]). Our study further suggests that while X-ray evaluation is consistent with the naked-eye method in identifying healthy seeds, it may yield different results when categorizing specific types of damage.
Conclusion
In pine nut-producing regions of Turkey, the pine nut trade has become the backbone of the local economy, and the Stone pine plays a significant role in cultural traditions. Pine nuts have been consumed as food since the Paleolithic era ([38], [39]). In the Bergama region, they hold such significance in local culture that a groom’s skill in climbing pine trees to collect cones plays an important role in marriage selection. The decline in pine nut yields has far-reaching socioeconomic consequences, pushing producers to seek alternative sources of income. This shift accelerates the decline of pine nut ecosystems, further threatening both the economy and cultural heritage tied to these forests.
Our results suggest that L. occidentalis is a major contributor to the ongoing yield loss in Stone pine in western Turkey. Therefore, the severity of the problems caused by this pest should be carefully evaluated, and decision-makers and producers should be adequately informed to implement concrete management plans to mitigate the impact of L. occidentalis. Furthermore, continuous monitoring of its populations and the associated seed losses should be established in western Turkey. Seed damage can be quantified using various evaluation methods, though their sensitivities differ. Depending on the objective, they can be used individually or in combination to monitor annual seed loss in Stone pine, ultimately helping to develop improved management plans and ensuring the sustainability of pine nut production in a changing environment.
Acknowledgements
This study was conducted in 2020-2021 as part of the project “Determination of Leptoglossus occidentalis Damage in Stone Pine Cones in Izmir-Bergama Kozak Region” under the broader initiative “Causes of Decreases in Cone and Seed Yield in Stone Pine and Other Conifers and Control Possibilities Integrated Project” funded by the Turkish General Directorate of Forestry. We also thank Ferudun Gürkaya, the chief of Karaveliler Village, and the personnel of the Bergama Forest Department for their support during the field studies, as well as the anonymous reviewer who contributed to improving the manuscript.
Author contributions
MA, SO, and KI developed the study idea; MA, SO, TC, FÖ, MÖ, and KI carried out the field studies; MA, SO, and KI carried out the statistical analyses; and KI wrote the manuscript.
References
Online | Gscholar
Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Sükran Oguzoglu 0000-0002-1816-8691
Isparta University of Applied Sciences Faculty of Forestry Department of Forest Engineering, 32260, Isparta (Turkey)
Bursa Regional Directorate of Forestry, Altinova Mahallesi, Istanbul Caddesi 4. Km, no. 420/1, 16250, Osmangazi, Bursa (Turkey)
I Meltem Özçankaya 0000-0002-5144-8345
Izmir Regional Directorate of Forestry Department of Forest Pest Management, Karsiyaka, Izmir (Turkey)
Hacettepe University Department of Biology, 06800, Beytepe, Ankara (Turkey)
Hacettepe University Biological Diversity Research and Application Center, HUBIOM, 06800, Beytepe, Ankara (Turkey)
Corresponding author
Paper Info
Citation
Avci M, Oguzoglu S, Can T, Öçal F, Özçankaya IM, Ipekdal K (2026). Measuring the damage of Leptoglossus occidentalis in Stone pine seeds in a pine nut production hot spot in Turkey. iForest 19: 201-208. - doi: 10.3832/ifor4854-018
Academic Editor
Matteo Marchioro
Paper history
Received: Mar 20, 2025
Accepted: Nov 04, 2025
First online: Jun 04, 2026
Publication Date: Jun 30, 2026
Publication Time: 7.07 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2026
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 1114
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 216
Abstract Page Views: 406
PDF Downloads: 457
Citation/Reference Downloads: 0
XML Downloads: 35
Web Metrics
Days since publication: 57
Overall contacts: 1114
Avg. contacts per week: 136.81
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Jul 2026)
(No citations were found up to date. Please come back later)
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
The effect of seed size on seed fate in a subtropical forest, southwest of China
vol. 9, pp. 652-657 (online: 04 April 2016)
Research Articles
Seed trait and rodent species determine seed dispersal and predation: evidences from semi-natural enclosures
vol. 8, pp. 207-213 (online: 28 August 2014)
Research Articles
Seed germination traits of Pinus heldreichii in two Greek populations and implications for conservation
vol. 15, pp. 331-338 (online: 24 August 2022)
Research Articles
The effectiveness of short-term microwave irradiation on the process of seed extraction from Scots pine cones (Pinus sylvestris L.)
vol. 13, pp. 73-79 (online: 13 February 2020)
Research Articles
The maternal environment of European beech (Fagus sylvatica L.) affects intrapopulation variability in seed traits and germination
vol. 19, pp. 114-121 (online: 10 April 2026)
Short Communications
Evidence of Alectoris chukar (Aves, Galliformes) as seed dispersal and germinating agent for Pistacia khinjuk in Balochistan, Pakistan
vol. 14, pp. 378-382 (online: 22 August 2021)
Research Articles
Maximizing growth of Acacia confusa through native plant growth-promoting bacterial inoculation and seed pelleting for revegetation in landslide areas
vol. 19, pp. 28-37 (online: 11 January 2026)
Research Articles
Influence of mother plant and scarification agents on seed germination rate and vigor in Retama sphaerocarpa L. (Boissier)
vol. 7, pp. 306-312 (online: 08 April 2014)
Research Articles
Decline in commercial pine nut and kernel yield in Mediterranean stone pine (Pinus pinea L.) in Spain
vol. 13, pp. 251-260 (online: 03 July 2020)
Review Papers
Soil seed banks of pioneer tree species in European temperate forests: a review
vol. 11, pp. 48-57 (online: 25 January 2018)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword






