Spatial distribution of aboveground biomass stock in tropical dry forest in Brazil
iForest - Biogeosciences and Forestry, Volume 16, Issue 2, Pages 116-126 (2023)
doi: https://doi.org/10.3832/ifor4104-016
Published: Apr 17, 2023 - Copyright © 2023 SISEF
Research Articles
Abstract
Climate change is being intensified by anthropogenic emission of greenhouse gasses, highlighting the value of forests for carbon dioxide storing carbon in their biomass. Seasonally dry tropical forests are a neglected, threatened, but potentially critical biome for helping mitigate climate change. In South America, knowing the amount and distribution of carbon in Caatinga seasonally dry vegetation is essential to understand its contribution to the global carbon cycle and subsequently design a strategic plan for its conservation. The present study aimed to model and map the spatial distribution of the potential forest biomass stock across 32 forest fragments of Caatinga, in the state of Bahia, Brazil, using regression kriging and Inverse Square of Distance techniques, building from point measurements of vegetation biomass made on-the-ground in ecological plots. First, a model for estimating biomass was fitted as a function of environmental variables to apply regression kriging, and then applied to the maps of the selected components. Elevation, temperature, and precipitation explained 46% of the biomass variations in the Caatinga. The model residuals showed strong spatial dependence and were mapped based on geostatistical criteria, selecting the spherical semivariogram model for interpolation by ordinary kriging. Biomass was also mapped by the Inverse Square of Distance approach. The quality of the regression model suggests that there is good potential for estimating biomass here from environmental variables. The regression kriging showed greater detail in the spatial distribution and revealed a spatial trend of increasing biomass from the north to south of the domain. Additional studies with greater sampling intensity and the use of other explanatory variables are suggested to improve the model, as well as to maximize the technique’s ability to capture the actual biomass behavior in this newly studied seasonally dry ecosystem.
Keywords
Geostatistics, Regression Kriging, Spatial Analysis, Forest Inventory
Introduction
Seasonally dry tropical forests (SDTFs) act as important CO2 sinks as they promote carbon fixation from the atmosphere in their plant biomass and contribute to the quantification and cycling of nutrients ([1]). Knowing the biomass retained in plants allows us to understand carbon dynamics, thus providing a better understanding of the impacts of deforestation on global warming. In addition, they provide helpful information to guide forest management practices and inform decision-making in public and non-governmental sectors. For example, many efforts including floristic inventory and vegetation monitoring initiatives in tropical regions, such as the ForestPlots Plot Network ([19]) and SDTF research protocols ([13], [35]), have motivated civil society and government agencies to promote sustainable environmental land management through tree planting and ecological restoration ([8]). Setting targets and assessing the proportionate contribution of such projects requires a solid initial understanding of the potential stock of aboveground biomass for each ecosystem at both regional and global scales ([9]).
While the importance of this information for the conservation, management, and sustainable use of SDTFs is recognized ([10]), regional spatial models of aboveground biomass (i.e., green or dry weight at tree level or species at a local scale) in exact spatial resolution are still absent in many SDTF areas, especially in the Caatinga region of northeastern Brazil. These biomass maps are needed to, in turn, create spatial understanding of variation in carbon stocks and carbon storage potential. Some maps have been developed continental and globally ([43]), however, their relatively coarse spatial resolutions (≥ 1 km) limit their applications for local forest management, as does the fact that they contain relatively little information about the distribution potential of biomass in space ([51]) and its relationships with the region’s intrinsic environmental factors or gradients.
Measuring environmental variables is an essential step towards understanding on how to better quantify ecological patterns and the distribution of biomass and carbon ([11]). For example, positive relationships between biomass stock and environmental factors are shared across various vegetation types and ecological gradients ([48]). Despite encompassing the largest expanse of the South American SDTF biome and with the highest species richness and endemism ([12], [17]), in situ observations on biomass stock and environmental factors for the Caatinga seasonally dry forests are, however, still incipient and little known ([38]).
The Caatinga vegetation occurs under extreme edaphoclimatic characteristics and is perhaps the most vulnerable Brazilian region to climate change ([20]). In addition, the Caatinga is highly threatened by inappropriate land use practices. Due to the high population density of the northeastern semi-arid region, its natural resources are widely exploited, which results in reduced production and biomass stocks, in addition to fragmenting the landscape into different physiognomies, stages of regeneration, and land uses ([1]). This culminated in an intense degradation process that has advanced recently ([50]).
Thus, there is a need to develop techniques to estimate and map biomass and, in turn, produce reliable information on the potential aboveground stock of forest formations and support estimates of carbon dynamics. This work examines the potential use of global climate datasets developed from satellite observations to map patterns and potential distribution of aboveground biomass in Caatinga vegetation. Unlike wetlands with high biomass driven primarily by climatic factors (e.g., temperature and precipitation) among other spatially explicit environmental factors or gradients, the Caatinga vegetation has lower biomass due to its adaptive metabolic and functional characteristics for the region, but a comparatively high potential for carbon storage ([45]). However, there is little local evidence to predict biomass patterns and potential distribution from environmental factors.
These regional characteristics favor the use of environmental variables to develop ecological and distribution models. The available remote sensing data provide spatially refined information on landscape and vegetation heterogeneity over the Caatinga SDTFs that can be readily incorporated into models to predict the potential distribution of biomass. These models are strictly mathematical or based on specific ecological theories ([16]). A detailed discussion or review of these ecological models and theories is beyond the scope of this paper, and has already been fully addressed elsewhere ([14], [24]). Here, we are interested in modeling and mapping the Caatinga biomass distribution to understand how available resources and environmental factors condition these biomass patterns at the biome scale. We tested geostatistical methods that integrate remote sensing data and the location of geographic points from ground-based plot inventories in different Caatinga forest fragments. With this approach, we aim to model and map the potential distribution of biomass and evaluate the contribution of each environmental variable in predictive maps for the Caatinga SDTF in Bahia. Therefore, we started from the hypothesis that climatic and relief factors have a strong influence on aboveground biomass patterns in Caatinga vegetation. The paper is organized into four sections: (i) description of biomass and environmental data; (ii) description of spatial modeling and main environmental factors; (iii) assessment of potential range distributions of biomass; (iv) discussion on the potential distribution of total biomass for the characterization of suitable areas for conservation and forest management.
Material and methods
Characterization of the study area
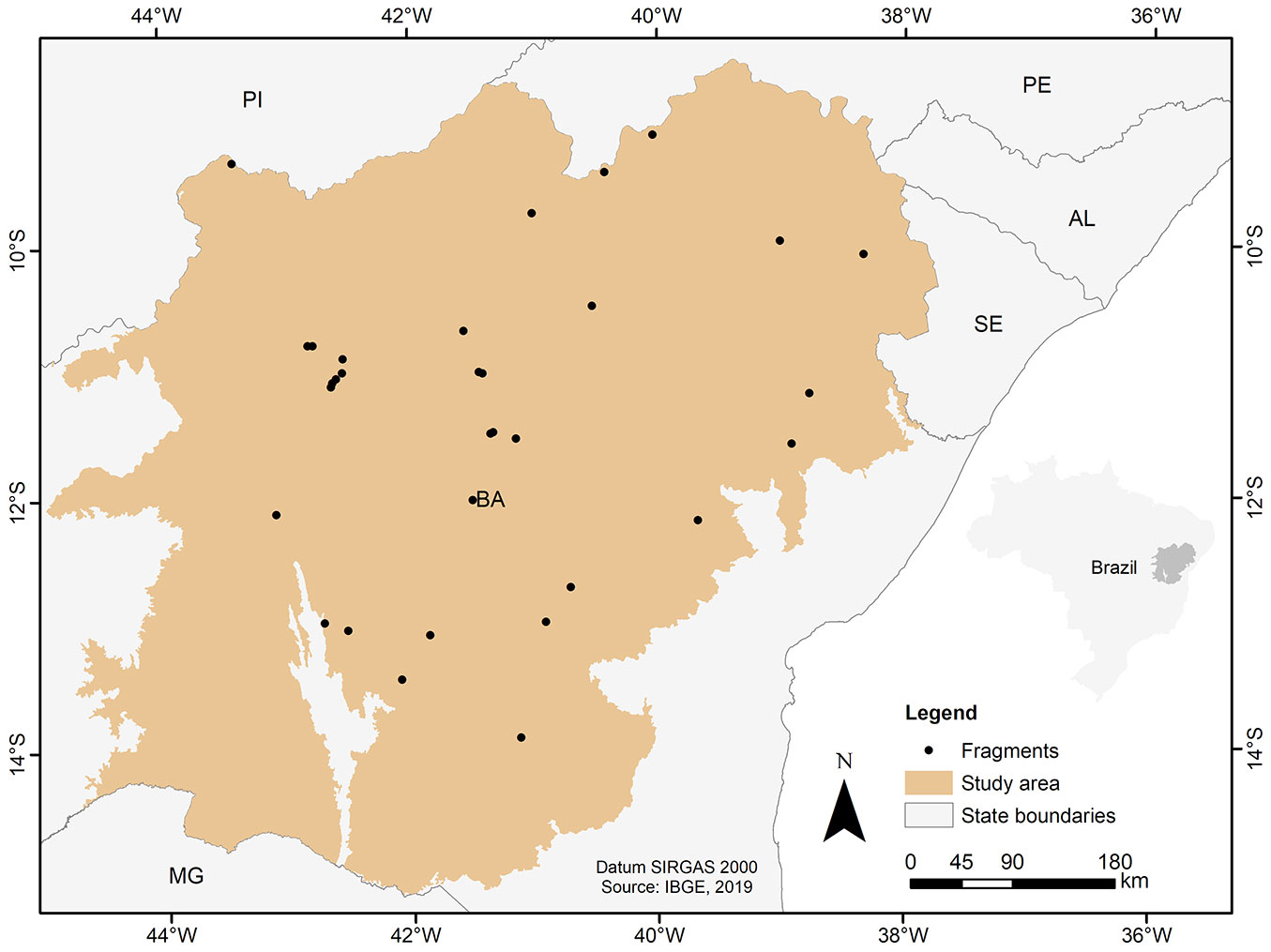
The area covered by this study is located in the state of Bahia (Fig. 1) and comprises a seasonally dry vegetation widely known as Caatinga, the most species-rich and largest expanse of the South American SDTF biome ([12], [17]). The Caatinga domain occupies an area of 351,402 km² in the state, covering about 62% of the Bahian territory ([26]).
Fig. 1 - Location of the sampled fragments in the Caatinga biome, in the state of Bahia, Brazil.
BSh climate according to the Köppen climate classification ([3]) is predominant in this area, characterized as a dry and hot semi-arid climate, which associates a scarcity of rainfall with strong insolation and high temperatures. The tropical As climate is also present, with a dry summer season, and the Aw type on the west side, with a dry winter. There are also subtropical humid zones with mild summers (Cwb climate with dry winters, and Cfb without a dry season) in a small region in the center-south, where altitudes are above 1000 m a.s.l.
There is irregularity in the rainfall distribution in time and space. This region receives from 300 to 1180 mm of annual rainfall; however, an expressive part of the area receives average rainfall of less than 750 mm year-1. Moreover, the precipitation is less than 400 mm per year in a small part. About 50% of these annual totals are generally concentrated in just three months ([18]).
Temperatures also vary in space, with annual averages between 16 and 29 °C, however, they are predominantly above 24 °C ([18]). There are plateaus and mountains which can exceed 1000 m in altitude. Among these geomorphologies are depressions which occur more expressively throughout the area. The relief has an influence on the spatial patterns of temperatures ([7]).
The soil orders that occur are: Neosols (typical in the semi-arid environment of northeastern Brazil, characterized by low natural fertility, low water retention capacity and low levels of organic matter), Latosols, Planosols, and even small fragments of Cambisols, Argisols and Luvissols ([31], [26]).
The most representative vegetation is steppe savanna, which has a deciduous, gray and thorny physiognomy in the dry season, interspersed with cactus and bromeliads. This landscape is interrupted by seasonal semi-deciduous and deciduous forest fragments which occupy regions of soils with rocky outcrops and karst relief associated with more clayey soils, and have the presence of limestone in the valleys and slopes. The latter is more expressive on the west side of the study area. There is the occurrence of savanna and small dense ombrophilous forest fragments in mountainous regions, swamps and milder climatic pockets, common in the center-south of the state of Bahia ([26]).
Biomass data estimates
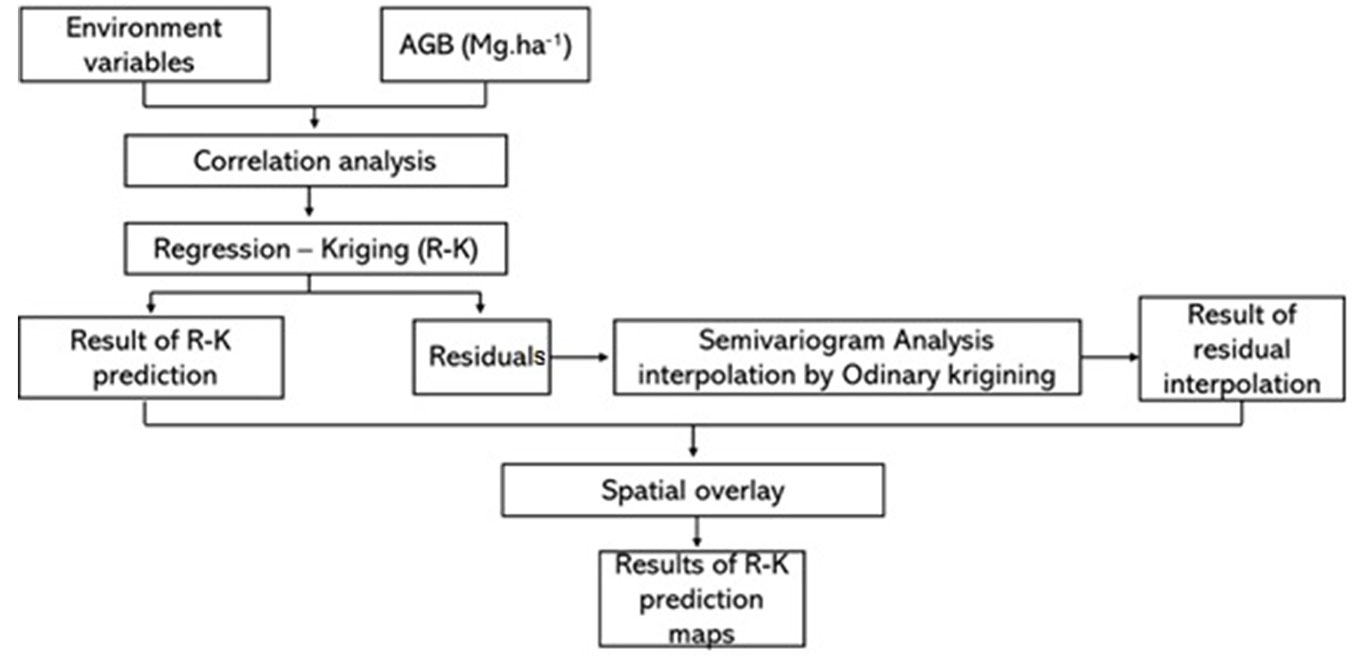
The database for biomass estimation was obtained from forest surveys carried out within the study area, which include a total of 836 plots distributed in 32 forest fragments (Fig. 1). A total of 21 forest inventories carried out between 2016 and 2020 were provided by the Institute for the Environment and Water Resources of the State of Bahia (INEMA) to compose this database. The number and size of plots varied between inventories at 0.02 to 0.1 ha. Tree individuals were measured in terms of diameter at breast height (DBH), measured at 1.30 m above ground level, adopting DBH greater than or equal to 10 cm as an inclusion criterion (Fig. 2).
Fig. 2 - Flowchart of Regression Kriging (GWR-K) and AGB mapping predictions in this study.
Data from 10 permanent plots of up to 1 ha were added to this database, registered in ForestPlots.net ([30], [19]), and which contribute to the Nordeste and Dryflor projects. These plots were inventoried between 2017 and 2018. Each plot for this data was assumed to be a forest fragment and only trees with DBH ≥ 10 cm were considered.
The Contendas do Sincorá National Forest was still considered as a single forest fragment in a survey carried out in 2015, inventoried by a partnership between the Caatinga Forest Management Network and the Forestry Soil and Forest Ecology and Protection Laboratories of the University State of Southwest Bahia (UESB). The measurement of arboreal individuals with DBH ≥ 15 cm was performed for this survey.
The equation developed by Sampaio & Silva ([44]) was applied to the individual data of the trees in the plots to predict the above ground woody biomass. The equation is specific to the Caatinga vegetation and is the most used by the scientific community (eqn. 1):
(R2 = 92%, RMSE = 3.8 kg, bias = 7.6%) in which AGB is the aboveground dry biomass for each tree individual (kg), DBH is the diameter measured at 1.30 m above the ground (cm) and R² is the coefficient of determination. RMSE (root mean squared error) is the measure that calculates the “root mean squared error” of the errors between observed values (actual) and predictions (hypotheses).
The biomass stocks of trees per plot were summed and were extrapolated in ton per hectare (Mg ha-1) according to the size of the plots. Due to the heterogeneity in the biomass stock of the plots within the same fragment, the dataset used in the analyzes was the average of the biomass stock of the plots in each of the 32 forest fragments ([46]).
Spatial modeling of biomass stock
The biomass stock mapping was performed using regression kriging (RK) and Inverse Squared Distance (IQD) techniques. Kriging with regression combines two processes: elaboration of a global map which illustrates the spatial behavior of the biomass, generated from a multivariate regression model; and subsequent application of ordinary kriging on the residuals generated by the regression model.
In order to introduce specific characteristics of each micro-region of the study area and provide a continuous data base, the biomass was modeled as a function of spatial variables obtained in raster layers (Tab. 1). These variables demonstrated a potentially significant relationship with the spatial distribution of forest attributes in previous studies ([47]).
Tab. 1 - Bioclimatic variables and elevation above sea used to adjust the biomass prediction model in the caatinga, Bahia.
| Variables | Descriptions |
|---|---|
| BIO 1 | Average annual temperature (°C) |
| BIO 2 | Average daytime range (monthly average) (°C) |
| BIO 3 | Isothermal (Bio 2/ Bio 7) (·100) (°C) |
| BIO 4 | Temperature seasonality (standard deviation·100) (°C) |
| BIO 5 | Maximum temperature of the warmest month (°C) |
| BIO 6 | Minimum temperature of the coldest month (°C) |
| BIO 7 | Override temperature range (Bio 5 - Bio 6) (°C) |
| BIO 8 | Average temperature of the wettest quarter (°C) |
| BIO 9 | Average temperature of the driest quarter (°C) |
| BIO 10 | Average temperature of the warmest quarter (°C) |
| BIO 11 | Average temperature of the coldest quarter (°C) |
| BIO 12 | Average annual rainfall (mm) |
| BIO 13 | Rainfall in the wettest month (mm) |
| BIO 14 | Rainfall of the driest month (mm) |
| BIO 15 | Seasonality of precipitation (Coefficient of variation) (mm) |
| BIO 16 | Precipitation of the wettest quarter (mm) |
| BIO 17 | Rainfall in the driest quarter (mm) |
| BIO 18 | Precipitation of the warmest quarter (mm) |
| BIO 19 | Rainfall of the coldest quarter (mm) |
| Elev | Elevation (m) |
After estimating the biomass stock, the data were associated with a set of predictor variables at each location. Twenty-eight geospatial covariates were selected and grouped into climatic and topographical subsets. The covariates were obtained using satellite remote sensing and globally distributed terrestrial weather stations in raster format. Each raster layer is a spatially explicit grid image, where each pixel represents the value of the described covariate.
The covariates were grouped into two distinct categories: topographical and climatic. Topographic covariates will include elevation, terrain slope, aspect (such as north and east), latitude (such as absolute latitude value), and a terrain roughness index (IRT). The climatic covariates will be composed of potential evapotranspiration, solar radiation, wind speed, cloud cover, and the set of 19 bioclimatic variables, 11 of which are derived from temperature: Average annual temperature, Average daytime interval (Monthly Average: max temp - min temp), Isothermal, Temperature Seasonality (standard deviation · 100), Maximum temperature of the hottest month, Minimum temperature of the coldest month, Annual temperature range, Average of the wettest quarter, Average of the driest quarter, Average of the warmest quarter of the year and Average of the coldest quarter of the year; and eight variables derived from Rainfall: Annual Rainfall, Rainfall in the coldest month, Rainfall in the driest month, Seasonality of Rainfall (Coefficient of variation), Rainfall in the wettest quarter, Rainfall in the driest quarter, Rainfall in the hottest quarter and Rainfall of the coldest quarter). The complete description of the topographic covariates can be consulted at ⇒ http://earthenv.org/, and for the climatic covariates, more information can be obtained at ⇒ https://www.worldclim.org/.
All variables acquired to fit the model, except latitude and longitude, were processed in the R software ([40]). To obtain the values of the geospatial covariates corresponding to each study site (at the plot and fragment level), they were initially obtained for the state of Bahia, and later, using the coordinates of each site, the extraction of environmental values was performed. For each point sampled using the “raster::extract” function of the R “raster” package ([25]) in each raster file of the geospatial covariates, this information was then stored and saved in a final matrix and used as predictor variables in the model.
The biomass values used in the modeling were submitted to exploratory analysis to identify the presence of outliers and their influence on the regression assumptions. Multiple linear regression analysis was used to adjust the model, and the parameters were estimated using the least squares method. The stepwise technique was used based on the Akaike Information Criterion (AIC) with both directions in order to define which variables best explain the biomass to construct a multivariate model.
The Shapiro-Wilk and Breusch-Pagan tests were applied to verify if the model residuals presented normality and homoscedasticity, respectively. In addition, the model was submitted to the Variance Inflation Factor (VIF) test to analyze possible correlations between the explanatory variables and ensure that the model is free of multicollinearity.
The coefficient of determination (R2), Mean Absolute Error (MAE) and graphical analysis of residuals were evaluated to test the accuracy of the fitted model. We also used the scatter plot of the predicted values around the 1:1 line to observe the behavior of the predictions made by the model. All analyzes were performed using the R software program ([40]).
Spatialization of the biomass stock
The maps of the variables selected in the multivariate model were designed for the study area in raster format, with a spatial resolution of 1 hectare (100 × 100 m). The regression model was applied on these cells, making it possible to estimate the biomass stock in each pixel and consequently generate a global predictive map of aboveground biomass. However, this map needs to be detailed in the regression kriging procedure by adding a spatial distribution map of the regression model residuals.
Next, an exploratory analysis of the data was used to apply geostatistics on the residuals in order to know their distribution, trends and identify atypical observations (outliers). This preliminary analysis is fundamental for decision making in geostatistical procedures.
The classic estimator of Matheron ([32]) was used to calculate the experimental semivariogram of the residuals, and the range was limited to half the maximum distance between two points. The behavior of semivariograms constructed in four horizontal directions was then verified to investigate the existence of anisotropy in the residuals: 0°, 45°, 90° and 135° ([22]).
The spherical, exponential, and Gaussian theoretical models were directly fitted to the data using the Maximum Likelihood method, considering the stationarity assumption of the intrinsic hypothesis ([28]). This fitting method estimates the parameters as a function of the original data set and does not show dependence on the experimental semivariogram points; however, it requires data normality ([46]). After the fittings, the semivariogram parameters were determined and the ratio between the nugget effect and the threshold showed the degree of spatial dependence, in accordance with Cambardella et al. ([6]).
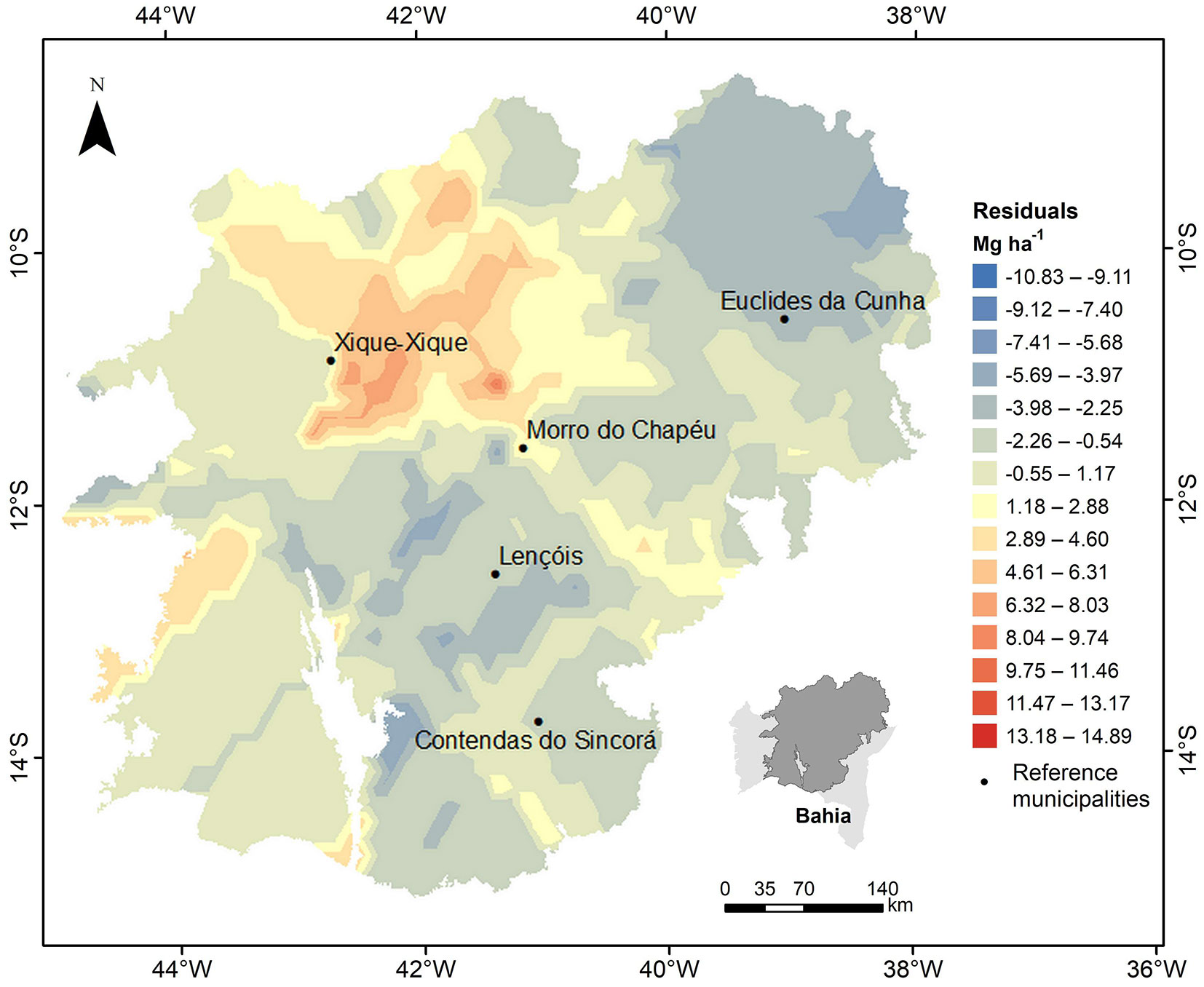
The theoretical model for the interpolation was selected by evaluating the Akaike Information Criterion (AIC), the degree of spatial dependence, the reduced mean error (RME) and the standard deviation of the reduced mean error (SDRME) provided by Jackknife cross-validation ([36]). The closer to zero the RME and the closer to 1 the SDRME, the better the performance of the model. Then the regression residuals were interpolated from the parameters of the selected spatial model by ordinary kriging, obtaining the residual map to add a stochastic aspect to the global map of the biomass stock. This approach aims to improve the initial quality of the global map estimates, which may present trends in the estimates and not generate a sufficiently detailed spatial distribution of the biomass (Fig. 3).
Fig. 3 - Mapping by ordinary kriging of the residuals of the regression model.
The predictive kriging quality was assessed by the mean error (ME), mean standard error (MSE), and root mean squared error (RMSE), which measure the accuracy of estimates and are provided by cross-validation by the k-fold method in the package “MASS” in R. Therefore, the final unbiased map of the spatial distribution of biomass was obtained for the entire Caatinga domain area in the state of Bahia by adding the residual map to the global map using map algebra tools. The semivariogram fitting was performed using the R software program ([40]), with the help of the “geoR” package ([42]) and the maps and interpolation were prepared in the ArcGIS® software program ([15]).
The prediction of biomass using the deterministic interpolator was also performed using the Inverse Weighted Distance (IWD) with exponent 2, a method known as Inverse Squared Distance (ISD). Neighborhood parameters were defined for a minimum of 10 points and a maximum of 32 points, which corresponds to the total number of sampling points in the study area. Interpolation by ISD was performed in the ArcGIS® software program ([15]). Mapping performance was evaluated on visual and statistical criteria. The mean error (ME) and the Root Mean Square Error (RMSE) were used for the statistical evaluation. The ME measured the prediction bias and its values must be close to zero for unbiased predictions ([41]). Negative ME values suggest that the prediction technique is overestimating, while positive values point to an underestimation.
Results and discussion
Exploratory analysis
The estimated biomass stock values in the studied fragments ranged widely, from 2.85 to 80.88 Mg ha-1. There is similarity between the mean (25.17 Mg ha-1) and median values (20.33 Mg ha-1), which points to an approximately symmetrical data distribution, but the superiority of the mean indicates that the highest amounts of biomass are further from the center compared to the lowest stocks. The marked difference between the minimum and maximum values resulted in a high coefficient of variation (73.7%), reflecting the heterogeneity of the study area. This variation portrays how the conditions of the sampled fragments are different. Different successional stages and anthropization degrees occur in these fragments, leading to variation in the number of individuals per hectare, in the richness and diversity of species, and in the diameters and heights of the trees.
The average biomass found was similar to that estimated by Souza et al. ([49]) in a protected area in Pernambuco (28.48 Mg ha-1), and lower than a fragment in Sergipe, which exhibited a contribution of 54.93 Mg ha-1 ([37]). Lima Júnior et al. ([29]) estimated values between 5.93 and 60.74 Mg ha-1 in Petrolina (PE), while Santos et al. ([45]) observed a smaller range in Caicó (RN) of between 10 and 17.8 Mg ha-1 of biomass. The discrepancy between the biomass stocks observed in the referenced works evidences the physiognomic variation that occurs along fragments of the Caatinga.
These differences in stocks between Caatinga areas are associated with precipitation, irregular rainfall distribution and the successional stage of the forest, which promote variation in the species biomass accumulation and distribution ([29]). Souza et al. ([49]) confirmed that the formation biomass is highly variable because it results from a combination of the age of the forest fragment, rainfall and species richness. The authors found that older forests exposed to greater precipitation support a greater number of species and have twice the biomass when compared to successional and species-poor areas.
Modeling the biomass stock
We identified fragments with outliers for biomass removed from the database used for subsequent analyzes as they influenced the regression assumptions. Thus, 27 fragments were used for modeling and geostatistical analysis.
The parameters used in the regression model showed significant coefficients (Tab. 2), which implies the importance of the selected variables. The VIF test confirms the absence of multicollinearity. The generated model is parsimonious and retained three variables, which is desirable as it reduces the complexity and time spent collecting data and using the model.
Tab. 2 - Coefficients estimated by the regression model and Variance Inflation Factor (VIF).
| Parameter | Variable | Coefficient | p-value (<0.05) | VIF |
|---|---|---|---|---|
| b0 | intercept | 519.3 | 0.000346 | - |
| b1 | Elev | -0.0000594 | 0.000293 | 1.70 |
| b2 | Temp | 1.831 | 0.045831 | 1.18 |
| b3 | Prec | -0.03326 | 0.016238 | 1.59 |
The model presented a coefficient of determination (R2) equal to 46% and Mean Absolute Error (MAE) of 5.9%, which indicated that the model performed well. The MAE demonstrates the model’s ability to make estimates closer to the real ones when its value tends to zero ([41]). The R2 demonstrates how much the model is able to explain the observed data. The value found herein is considered acceptable given the magnitude of the study area and the wide variation that occurs between the biomass stocks of the studied fragments. Similar conditions are also reported by Scolforo et al. ([46]), who found R2 equal to 53% for a geographic model that estimates the carbon stock in the state of Minas Gerais.
The regression residuals follow a normal distribution (Shapiro-Wilk, p-value = 0.32) and are homoscedastic (Breusch-Pagan, p-value = 0.36). These characteristics are essential to define the adequacy of the model and allow adopting the Maximum Likelihood method in fitting the semivariogram, which requires a priori knowledge of the data distribution ([2]). The quality of the model can also be verified in Fig. S1 (Supplementary material). The model presented randomly distributed residuals and did not show significant deviations from the mean or variance heterogeneity, suggesting that there were no underestimation or overestimation trends (Fig. S1a). These characteristics configure an estimation bias-free model. Fig. S1b illustrates the behavior of the estimated vs. the observed biomass. The closer the predicted values are to the 1:1 line, the better the fit. This means there is greater precision, less chance of estimation bias, and consequently the sum of residuals tends to zero.
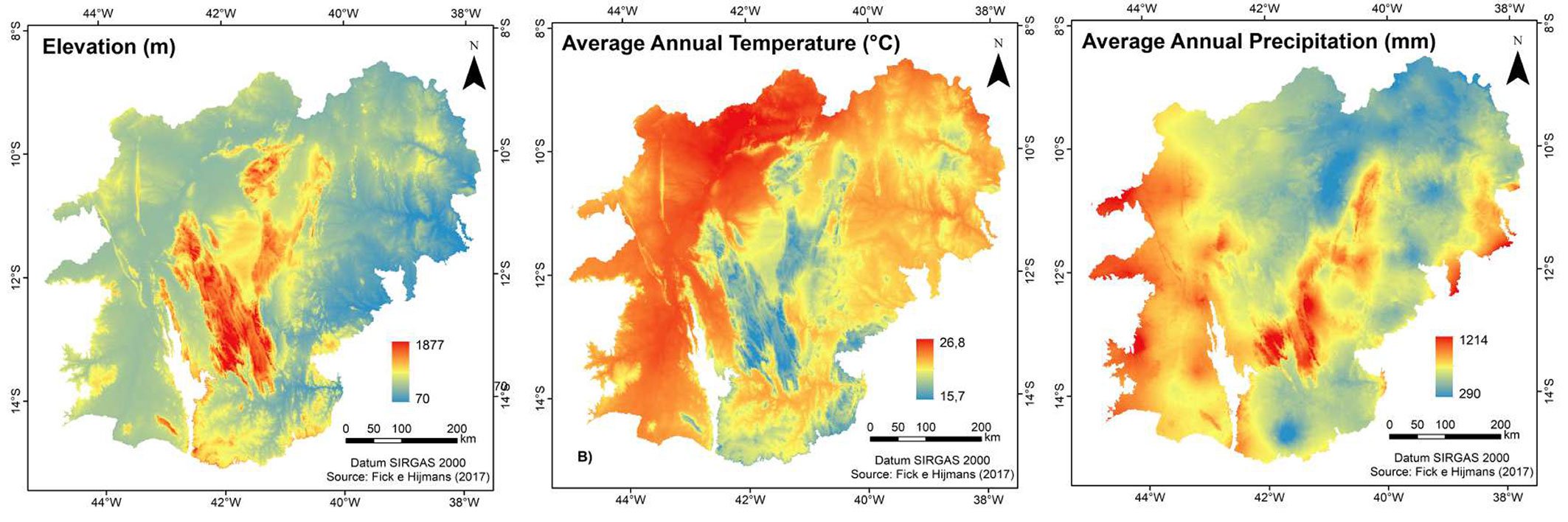
Elevation, average annual temperature and average annual precipitation are the main variables responsible for the variation in the Caatinga biomass stock in the state of Bahia (Fig. 4). The contribution of precipitation is already consolidated in the literature ([4], [39], [47], [49]). The availability of water in the soil increases the photosynthetic activity of plants, influencing the seasonality and intensity of leaf production, and consequent biomass production ([49]).
Fig. 4 - Variables selected by the multivariate regression model to estimate biomass in the Caatinga, Bahia.
The prediction error associated with the developed model may be due to the precision degree of applying the allometric equation used in this study, which presents R2 equal to 92% ([44]). The error can also be attributed to the measuring process of the trees which was performed on different occasions and by different teams. The high variability that occurs in the biomass values, which promoted the high coefficient of variation, also represents an additional difficulty for fitting the multivariate model. This was also observed by Mello et al. ([33]), who reported losses in precision statistics when modeling rainfall erosivity for the Northeast region attributed to the high variability of precipitation in this region.
Scolforo et al. ([46]) and Silveira et al. ([47]) proved that environmental components can be highly explanatory variables capable of estimating forest attributes. The present study corroborates these authors by identifying that there is a potential for developing models capable of estimating the biomass of the Caatinga vegetation using environmental variables. However, it is necessary to improve these models, aiming at increasing the accuracy of the estimate, given that regression equations are always associated with uncertainties ([46]). Studies using a larger sample population are suggested, which will allow construction of more robust equations with fewer errors associated with the predictive capacity.
It should be noted that the accumulation of biomass in forests does not only respond to the variables considered, but also to other environmental conditions, such as soil fertility, water deficit, seasonality of rainfall and successional stages ([39], [49]). The use of vegetation indices and remote sensing data is certainly a convincing option for biomass characterization and for developing regression models with lower degrees of uncertainty. Work in this direction has already been developed by Lima Júnior et al. ([29]), Xue et al. ([52]) who found good performance in the fitted functions.
Spatialization of the biomass stock
The semi-variance calculated in the four directions did not show significant differences and the semivariogram was considered isotropic, indicating that the spatial dependence of the regression residuals only depends on the distance between the points and is the same in all directions ([34]).
The theoretical semivariograms fitted to the data by the Maximum Likelihood method are represented in Fig. S2 (Supplementary material). It is noteworthy that the residuals had a normal distribution, which legitimizes the use of this method ([41]). It is possible to detect that the residuals of the regression model are spatially structured independent from the theoretical model.
Tab. 3shows the parameters of the fitted theoretical models and their respective degrees of spatial dependence of the residuals, in addition to the statistical criteria used to define the best model. The spherical model presented the best fit to the regression model residuals due to the lower degree of spatial dependence and nugget effect, lower AIC and reduced mean error (RME) closer to zero.
Tab. 3 - Statistical parameters and criteria of the theoretical models adjusted for the residuals of the regression model. (C0): nugget effect; (C1): threshold; (A): range; (GD): degree of spatial dependence; (EMR): mean reduced error (SER): standard deviation of reduced errors (AIC): Akaike Information Criterion.
| Model | C0 | C1 | A (m) | GD (%) | EMR | SER | AIC |
|---|---|---|---|---|---|---|---|
| Exponential | 11.75 | 49.00 | 21000 | 23.90 | 0.018 | 1.001 | 187.3 |
| Gaussian | 13.69 | 48.12 | 21000 | 28.45 | 0.020 | 1.002 | 186.3 |
| Spherical | 5.06 | 45.45 | 21000 | 11.14 | 0.016 | 0.992 | 185.4 |
We can see the smallest nugget effect associated with the spherical model necessary to provide more accurate estimates in the kriging interpolation in Fig. S2 ([2]). The nugget effect different from zero indicates that there is a random variation of the residual data that cannot be explained by the spatial component due to the lack of sampling in the distances smaller than the smallest distance between the plots ([46]). The lowest average distance recorded for the sampled fragments was 2447 meters.
The range found was 21.000 meters and suggests that the residuals are spatially correlated up to this distance. The high range value is due to the distances between the sampled fragments and the magnitude of the study area. The relationship between the nugget effect and the threshold expresses the degree of spatial dependence of the variable, which is classified as strong (< 25%), moderate (25 to 75%) and weak (> 75%), according to Cambardella et al. ([6]). All models showed a residual spatial dependence structure from strong to moderate, constituting desirable indices in geostatistical modeling ([34]).
The ordinary kriging mapping of residuals by the spherical model is shown in Fig. 3. It is observed that closer points in space tend to have similar values to each other, which is the main feature revealed by geostatistics ([32]). Mello et al. ([33]) highlighted that the positive values of the residuals represent underestimation of the model, while the negative values correspond to overestimation. Thus, the authors assure that a balanced distribution of these estimates is desirable, as it suggests that there are no trends produced by the regression model. A balance between under- and overestimates is noticeable on the map, showing that the model is suitable for the study area.
The error metrics associated with the interpolated surface in the ordinary kriging of residuals are presented in Tab. 4and reinforce that the spherical theoretical model was adequate in interpolating residuals. The difference found between the RMSE and ASE indicates that the forecast was underestimated by 0.43 Mg ha-1 ([34]).
Tab. 4 - Statistics of the interpolation by ordinary kriging of the residuals of the regression model. (ME): Mean Error; (MSE): Mean Standardized Error; (RMSSE): Root Mean Square Error Standardized; (RMSE): Root Mean Square Error; (ASE): Mean Standard Error. For the semivariogram model to be suitable for interpolation, ME and MSE must be close to 0; the RMSSE should be close to 1 e; the RMSE and the ASE should have similar values and the smallest possible ([34]).
| Statistics | Value |
|---|---|
| ME | -0.16 |
| MSE | 0.01 |
| RMSSE | 0.97 |
| RMSE | 6.39 |
| ASE | 6.82 |
The combination of the regression model estimates (Fig. 5a) and the kriging residuals (Fig. 3) resulted in the final prediction map of the biomass stock by kriging with regression (Fig. 5b). The global map and the map corrected by the kriging of residuals reveal similar behaviors of the biomass stocks; however, the greater detail given by the final map is remarkable. The biomass stock map interpolated from the ISD technique is represented in Fig. 5c. It is characteristic of this technique to preserve the amplitude of the original data, so the minimum and maximum values found on the map are those indicated in the descriptive statistics of the data.
Fig. 5 - Global map of biomass stock obtained by the regression model (A), interpolated by kriging with regression - RK (B) and interpolated by the Inverse Square of Distance technique - IQD (C).
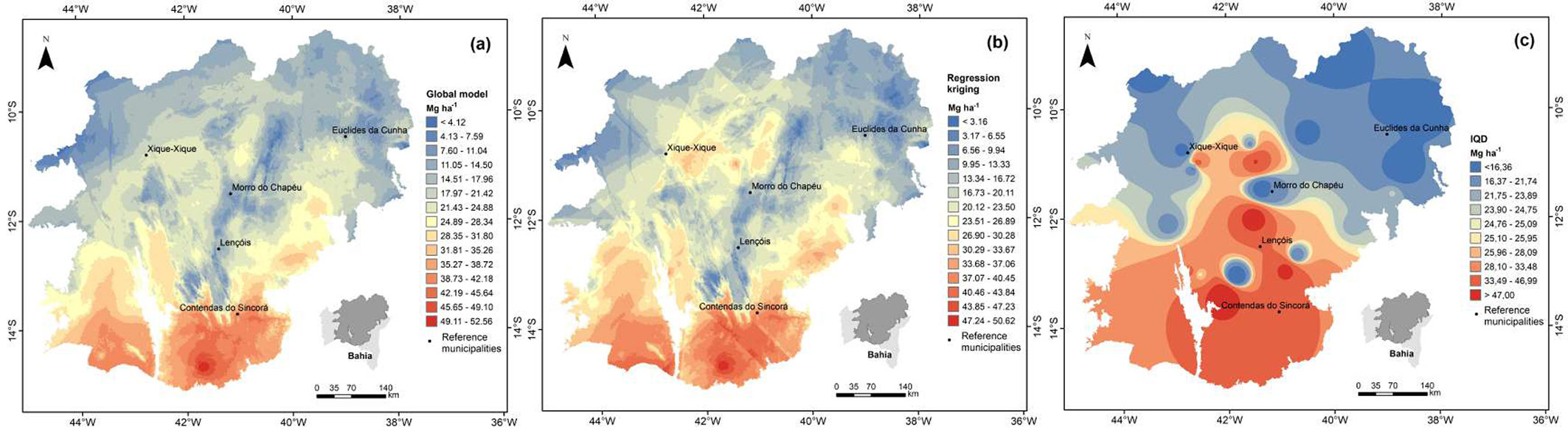
There is generally an increase in the biomass stock in the north-south direction. Biomass stocks below 20 Mg ha-1 occur in the north of the study area due to lower precipitation rates and higher temperatures. The region surrounding the municipality of Euclides da Cunha is classified by Ganem ([20]) as a priority area for creating an integral protection conservation unit due to the urgency of habitat loss and susceptibility to environmental degradation.
A small area near the municipality of Xique-Xique stood out for exhibiting higher biomass values. The junction of the residual map shows the underestimation given by the global map in this region, reinforcing that the combination between a global interpolator and a geostatistician better represent the spatial distribution of a variable. The evident detail given in this specific area is due to the higher concentration of fragments sampled there, which in turn brought the regression kriging map (Fig. 5b) closer to the real stock in this region. In addition, a biomass gradient is observed in the mapping generated by kriging with regression which increases in the north-south direction. However, as this technique is dependent on the closest neighborhood points, the map did not detail the biomass for the subsampled regions and promoted the formation of islands around the sampled fragments.
The interpolated surface presented mean error (ME) values of -0.38, which suggests an overestimation of the biomass stock prediction by the ISD. The ME measures the model’s tendency to under or overestimate a variable of interest and should be close to zero for unbiased predictions ([41]). The Root Mean Square Error (RMSE) was 19.83 Mg ha-1. Although this type of interpolation is useful in the absence of spatial structure of the variable of interest, or even in the presence of weak dependence structure, the use of the ISD was relevant because it showed that the inconsistency in the mapping by kriging with regression was influenced by the subsampling in the study area.
Small isolated areas with smaller stocks can be observed in the south-central portion of the map where biomass stocks are greater than 25 Mg ha-1, which constitutes an effect of the low estimated biomass in the fragments sampled there. This shows that these fragments do not capture the real variation of biomass stock that occurs in the study area and reinforces that the underestimation of the Chapada Diamantina represented in the regression kriging map is due to the effect of these fragments. Despite this, in visually comparing the maps it is notable that the regression kriging is able to capture more specific details regarding the biomass distribution in the studied area than the ISD, which makes it more efficient in the mapping.
The largest biomass stocks are in the south of the map. The biomass in the municipality of Contendas do Sincorá was estimated at 37 Mg ha-1, while Virgens et al. ([51]) estimated an average value of 29.2 Mg ha-1. The authors classify the predominant vegetation in this area as arboreal Caatinga and in a late successional stage. Considering that these authors used the direct quantification method which is more accurate, it is stated that the biomass in the present study was slightly overestimated in this region.
The highest biomass stocks indicated in southern Bahia suggest that these are potential areas in the Caatinga to store carbon. Considering that the carbon contents in the biomass are generally around 50% ([27]), the forest carbon stock gradient in the area tends to be similar to that presented in Fig. 5b.
Low biomass values were found for the Chapada Diamantina region, where the municipalities of Morro do Chapéu and Lençóis are located. Chapada Diamantina is characterized by a set of mountains that reach more than 1000 meters in elevation, with high precipitation levels and milder temperatures (Fig. 4), constituting favorable conditions for forest productivity ([49]). Thus, this area tends to have larger trees, sustain denser forests and consequently store high stocks of plant biomass, as evidenced by the biomass mapping throughout the Caatinga carried out by Castanho et al. ([7]) using satellite images. The authors estimated values higher than 60 Mg ha-1 in this region and classified it as one of the areas with the highest plant biomass in the entire Caatinga.
However, this region does not support a homogeneity of dense vegetation. Chapada Diamantina is a set of communities which form a mosaic rich in physiognomies, depending on the topography, nature and depth of the soil. The high altitudes and busy relief result in a high erosion and occupation rate of the areas by rocky outcrops, which is limiting to vegetation development, with formations having different physiognomies from the general dominant context of this region occurring in these soils ([5]).
Therefore, it is likely that the low biomass values in this area on the map (Fig. 5b) are due to fragments sampled under these conditions. A higher sampling intensity over denser forest areas would allow regression kriging to more adequately estimate the spatial behavior of the biomass stock in Chapada Diamantina, respecting its different physiognomies. Thus, the conclusions about the spatial distribution of the biomass stock in the study area are moderate, recognizing that the database is limited in the number and distribution of fragments.
However, lower mapping accuracy was expected due to the richness of physiognomies of the Caatinga in Bahia, as characterizing the biomass in heterogeneous environments such as tropical forests is a challenge ([47]). But the result was satisfactory considering the database and the richness of detail achieved in the final regression kriging map.
There is no consolidated database available which covers the entire territory of the state of Bahia, which validates the data compiled in this study. Although not as consistent, the data provide the potential for biomass in each patch. A base of inventories capable of capturing the entire spectrum of vegetation variation in Bahia and showing its plurality of biomes, successional stages, anthropization degrees and phytophysiognomies is necessary. The existence of a consistent basis would enable developing techniques and studies capable of supporting the conservation and sustainable exploitation of the state’s forest resources with greater precision.
This information would provide greater detail in the final map in the present study and would promote a substantial improvement in mapping. Silveira et al. ([47]) and Scolforo et al. ([46]) achieved satisfactory results when mapping the spatial distribution of biomass and carbon stock, respectively, by regression kriging for the state of Minas Gerais, and close to what was observed in the field. However, both authors used a robust database with greater sampling intensity and well-distributed forest fragments in the study area.
Due to the heterogeneity of the Caatinga vegetation in Bahia, it is suggested to consider the different phytophysiognomies, ecoregions or climatic zones of the biome as independent variables in the biomass modeling in future works. This distinction has been used to reduce the spatial variation common in vegetation data over large areas ([4], [46], [47]). Barni et al. ([4]) considered the different phytophysiognomies that occur in the state of Roraima when modeling the biomass for the state, classifying them into four forest groups that were used as input variables in the regression model. With this, the authors managed to improve the performance in the interpolation, as the model was able to better capture the vegetation variations, and thus generate a better representation of the biomass stocks in the state.
Similarly, Scolforo et al. ([46]) considered the different biomes that occur throughout the state of Minas Gerais and included them as a categorical variable in the regression model used to spatialize the carbon stock in the state. The authors obtained a consistent, more flexible model that adequately estimated the spatial behavior of carbon, with a reduction of possible trends in specific areas.
Study limitations
Our analysis explored a variety of data sources and analytical tools that can be applied to develop predictive maps that incorporate the observed spatial pattern of biomass in landscapes at the scale of the Caatinga biome in the state of Bahia. However, the context of available data still lacks more excellent coverage or prescription for some areas, therefore some limitations remain.
Firstly, it is common to face problems using inventory data collected differently in each location, mainly associated with taxonomically reliable data and correctly defined technical protocols in a given location. As a result, some trees may be measured differently, and individual biomass estimates may show under- or over-estimate trends. For example, many local, regional, and pan-tropical equations use the diameter at 1.30 m ground level (dbh) in biomass estimates, and many inventories only have a diameter at 0.30 m ground level (diameter at base). Furthermore, not all Caatinga forest fragments or land use forms can be equally represented, leading to high standard deviations or region-biased biomass estimates. Although the locations were spatially well distributed for the Caatinga biome in Bahia, and some locations were close to each other, an effort was made to correct and standardize dendrometric measurements in specialized protocols and to use forest land-use maps at a scale of finer detail. The analysis of the biomass stocks discussed is also based on accurate classifications of land cover and markedly anthropized gradients, which may not be available or may be misaligned with the inventoried plots. This helps to correct errors about the representativeness and potential distribution of biomass at the plot level and the biome scale.
Second, different scales of environmental data (10 × 10 km tables) and inventory form another complication, as plots covering a few hectares are unlikely to represent an area of 10 × 10 km ([48]). In addition, some regions or areas contained more inventoried trees, plots, or surveyed surface area than others, resulting in different levels of captured biomass stock by the site. Although attempts have been made to correct the diversity values for these sampling biases, likely, they have at least partially influenced the results. Ideally, meta-analyses like this should be performed on standardized plot layouts in combination with high-resolution environmental data ([21]). Unfortunately, such an approach is still complex to carry out in more remote dry tropical areas in northeastern Brazil, where reliable inventory data is already a problem ([19]), not to mention reliable high-resolution data on soils and climate ([18], [23]).
Thirdly, and related to the available environmental layers and their potential to explain the biomass in Caatinga vegetation, it was noted that the functional importance of many environmental variables suggests poor results among all the variables tested based on characteristics related to predictive importance. The high collinearity may help to explain this fact; however, other modeling approaches, as well as other environmental variables, should be tested, mainly related to the soil of the biome. For example, edaphic factors are vital in explaining many attributes and ecological patterns of the forest, mainly related to productivity and diversity in tropical forests ([49]). In addition, the low predictive values obtained for the other climatic variables may be explained by the available GIS layers in which the spatially explicit grid (interpolated values) does not correctly explain the inherent values at the plot level, requiring, therefore, an effort of data collection at local scales. Detailed investigation of these factors locally would be critical, as it could identify other ecological functions of the edaphoclimatic information set for potential biomass at a more detailed scale in the Caatinga biome of Bahia. However, the results obtained corroborate with studies of the same tropical geographic regions ([43]) with highly varied species composition ([12]), which leads to the main conclusion being satisfactory. Finally, more direct studies on the set of ecological forces acting to define tree biomass patterns are needed to clarify these answers.
Conclusions
We integrate in situ tree measurements and remote sensing data to map the potential distribution of biomass in the Caatinga biome of Bahia with detailed spatial resolution. We primarily use free and open access data; therefore, our approach can be adopted and improved in other regions to assess total biomass stocks. While ecologists have an ever-expanding toolbox for extrapolating from local surveys to larger-scale patterns, it has long been assumed that the well-known barriers to reliable tree-level sampling and non-destructive estimates make sense extrapolations across the entire spectrum impossible in the Caatinga domain in the state of Bahia. Our methods enrich the information already available on biomass stocks at regional and global scales, as well as generate new perspectives for understanding and conserving tropical dry forests.
The quality of the regression model suggests that three main environmental factors possibly govern aboveground biomass. Increases in temperature and altitude suggest a reduction in biomass. However, the combination of average annual precipitation and moderate altitude conditions is the ideal climate to support the highest biomass stocks. Additional studies with a larger sample population and other variables can improve the model. Compared to ISD, regression kriging revealed in more detail the spatial variability of biomass. The adoption of a higher sampling intensity has the potential to maximize this detail given by the technique. A robust base of forest inventories is needed for Bahia in order to enable studies that allow for a more accurate understanding of the potential of the state’s forest resources.
Finally, we mapped the potential biomass distribution of the Caatinga biome in Bahia, but we do not guarantee that the conservation or preservation of these is established. The overwhelming effect of human activities (deforestation, fires, and the advance of agriculture and livestock) has led to substantial reductions in biomass and carbon stocks. Given the enormous carbon-storage capacity of tropical dry forests, the next avenue for research is to model the effects of climate change and future land-use changes on biomass patterns to refine conservation prospects in a changing world. These efforts are urgently needed in light of the constant and increasing rates of deforestation in the Caatinga biome in recent years and potentially threaten the planet’s high biodiversity hotspots.
Acknowledgments
We thank the ForestPlots.net, Inema and Caatinga forest management network teams for permission and availability of data to conduct the research. This project was supported by ForestPlots.net approved Research Project #142: “Mapping the biomass and carbon stock, volume and diversity in tropical forests", and the UK NERC Newton Fund project “Nordeste” (NE/N012550/1). The authors thank the Coordination for the Improvement of Higher Education Personnel (Capes) and the National Council for Scientific and Technological Development (CNPq) for financial support in a research grant for the first and third authors of this work.
The conceptualization, data curation and formal analysis: Santos HKV, Lima RB. The Investigation, Methodology, Validation, Visualization and Project administration: Santos HKV, Lima RB, Silva TTS, Souza RLT, Oliveira CP, de Paula A, Barreto-Garcia PAB, Veenendaal E, Queiroz L, Moonlight P, Rodrigues PMS, Santos RB, Sarkinen T, Pennington T, Domingues T, Cardoso D, Phillips O. Writing-original draft: Viana HK, Lima RB. Writing review and editing: Viana HK, Lima RB, Moonlight P, Phillips O, Cardoso D.
References
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Online | Gscholar
Gscholar
Online | Gscholar
Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Authors’ Info
Authors’ Affiliation
Thaine Teixeira Silva 0000-0001-6082-371X
Alessandro De Paula 0000-0003-3676-3846
Patrícia Anjos Bittencourt Barreto-Garcia 0000-0002-8559-2927
Universidade Estadual do Sudoeste da Bahia, Departamento de Ciências Florestais, 45031-900 Vitória da Conquista, BA (Brazil)
Cinthia Pereira De Oliveira
Francisco Tarcísio Alves Júnior
Universidade do Estado do Amapá, Departamento de Engenharia Florestal, 68900-070 Macapá, AP (Brazil)
Universidade de São Paulo, Escola Superior de Agricultura Luiz de Queiroz, 13418-900, Piracicaba, SP (Brazil)
Instituto de Biologia, Universidade Federal da Bahia, 40.026-010, Salvador, BA (Brazil)
Tropical Diversity Section, Royal Botanic Garden Edinburgh, EH3 5NZ, Edinburgh (United Kingdom)
Wageningen University, Plant Ecology and Nature Conservation Group, 6700 AK, Wageningen (Netherlands)
Departamento de Ciências Biológicas, Universidade Estadual de Feira de Santana, 440369-00, Feira de Santana, BA (Brazil)
Colegiado de Ecologia, Universidade Federal do Vale do São Francisco, 563049-17, Senhor do Bonfim, BA (Brazil)
Universidade Federal de Lavras, 37200-900, Lavras, MG (Brazil)
Tropical Diversity Section, Royal Botanic Garden Edinburgh, EH3 5NZ, Edinburgh (United Kingdom)
School of Geography, University of Leeds, LS2 9JT, Leeds (United Kingdom)
Corresponding author
Paper Info
Citation
Viana Santos HK, Borges De Lima R, Figueiredo De Souza RL, Cardoso D, Moonlight PW, Teixeira Silva T, Pereira De Oliveira C, Alves Júnior FT, Veenendaal E, Paganucci De Queiroz L, Rodrigues PMS, Dos Santos RM, Sarkinen T, De Paula A, Barreto-Garcia PAB, Pennington T, Phillips OL (2023). Spatial distribution of aboveground biomass stock in tropical dry forest in Brazil. iForest 16: 116-126. - doi: 10.3832/ifor4104-016
Academic Editor
Emanuele Lingua
Paper history
Received: Mar 24, 2022
Accepted: Feb 14, 2023
First online: Apr 17, 2023
Publication Date: Apr 30, 2023
Publication Time: 2.07 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2023
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 30197
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 21868
Abstract Page Views: 5142
PDF Downloads: 2646
Citation/Reference Downloads: 14
XML Downloads: 527
Web Metrics
Days since publication: 1209
Overall contacts: 30197
Avg. contacts per week: 174.84
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Jul 2026)
Total number of cites (since 2023): 4
Average cites per year: 1.00
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Review Papers
Integration of forest mapping and inventory to support forest management
vol. 3, pp. 59-64 (online: 17 May 2010)
Commentaries & Perspectives
Benefits of a strategic national forest inventory to science and society: the USDA Forest Service Forest Inventory and Analysis program
vol. 1, pp. 81-85 (online: 28 February 2008)
Research Articles
Using self-organizing maps in the visualization and analysis of forest inventory
vol. 5, pp. 216-223 (online: 02 October 2012)
Research Articles
Comparing land use registry and sample based inventory to estimate forest area in Podlaskie, Poland
vol. 10, pp. 315-321 (online: 23 February 2017)
Research Articles
Simplified methods to inventory the current annual increment of forest standing volume
vol. 5, pp. 276-282 (online: 17 December 2012)
Technical Advances
Improved estimates of per-plot basal area from angle count inventories
vol. 7, pp. 178-185 (online: 17 February 2014)
Research Articles
Geostatistical techniques for estimating aboveground biomass in eastern Amazonia
vol. 19, pp. 85-93 (online: 13 March 2026)
Research Articles
Methods to inventory and strip thin in dense stands of aspen root suckers
vol. 8, pp. 590-595 (online: 22 April 2015)
Research Articles
Are we ready for a National Forest Information System? State of the art of forest maps and airborne laser scanning data availability in Italy
vol. 14, pp. 144-154 (online: 23 March 2021)
Research Articles
Integrating area-based and individual tree detection approaches for estimating tree volume in plantation inventory using aerial image and airborne laser scanning data
vol. 10, pp. 296-302 (online: 15 December 2016)
iForest Database Search
Google Scholar Search
Citing Articles
Search By Author
- HK Viana Santos
- R Borges De Lima
- RL Figueiredo De Souza
- D Cardoso
- PW Moonlight
- T Teixeira Silva
- C Pereira De Oliveira
- FT Alves Júnior
- E Veenendaal
- L Paganucci De Queiroz
- PMS Rodrigues
- RM Dos Santos
- T Sarkinen
- A De Paula
- PAB Barreto-Garcia
- T Pennington
- OL Phillips
Search By Keywords







