Typology and synecology of aspen woodlands in the central-southern Apennines (Italy): new findings and synthesis
iForest - Biogeosciences and Forestry, Volume 13, Issue 3, Pages 202-208 (2020)
doi: https://doi.org/10.3832/ifor3315-013
Published: May 19, 2020 - Copyright © 2020 SISEF
Research Articles
Abstract
In order to review and complete our knowledge of the typology and synecology of the aspen communities from the central-southern Apennines, ten original relevés were performed on the Gargano plateau and a set of 35 relevés assigned to four community types (HP: Holco mollis-Populetum tremulae; MP: Melico uniflorae-Populetum tremulae; FP: Fraxino orni-Populetum tremulae; GP: Geranio versicoloris-Populetum tremulae) were assembled from literature. These relevés along with several environmental variables either measured or estimated were involved in cluster and ordination analyses. The relevés from Gargano formed a distinctive cluster and were assigned to a new community type (SP: Stellario holosteae-Populetum tremulae ass. nova), which can be considered an Adriatic synvicariant of HP that is distributed in similar habitats (doline bottoms) but on the Tyrrhenian escarpment. At low levels of floristic similarity, the grouping of relevés in two clusters induces a sharp separation between the aspen communities distributed in the central Apennines (MP and FP) and those from the southern Apennines (SP, HP and GP), which is mainly due to compositional differences in the regional species pool. The ordination scores of relevés were best related to terrain slope, soil nitrogen, elevation, air temperature, light availability and, to a lesser extent, to soil moisture and reaction. Unlike MP and GP that appear the most mesophilous, the FP stands display a slightly more xerophilous and acidophilous character induced by the steeper slopes on which they occur. The HP habitat is the driest and lightest very likely because of the open overlying canopy, in contrast to MP stands featuring a high shrub cover. The highest occurrence of nitrophilous species was observed in SP and MP. The management of these pioneer woods should be aimed at conservation, as they play an important role in the recovery of forest herb diversity along the ecological succession towards hardwood forests.
Keywords
Differential Species, Distribution Area, Environmental Variable Fitting, Gargano, Multivariate Analyses, Phytosociologic Classification, Secondary Succession, Stellario holosteae-Populetum tremulae
Introduction
Cultural landscapes not only represent a valuable legacy of human activity over millennia, but also a useful object of study for inferring patterns in vegetation dynamics ([15]). In particular, the long tradition of land cultivation and sheep farming in the Apennines has deeply shaped the vegetated landscape in these mountainous areas, a pattern which extends across the Eurosiberian and Mediterranean regions. Around the middle of the 20th century, the profound changes brought about by industrial development has led to extensive abandonment of arable fields, pasturelands and mown grasslands ([39]). As a consequence, the natural secondary succession, which has been developing over the following decades, has induced visible changes in the landscape through the development of patches of scrub and pioneer woodlands ([11], [24], [43]). They both play an important role in vegetation recovery, especially in facilitating the establishment of late successional tree species throughout the nemoral and boreal forest zones ([13]). On the positive side, this trend is beneficial for, among other processes, carbon sequestration ([19], [26]), reduction of soil erosion ([8]) and forest connectivity at landscape level ([17]). On the other hand, woodland encroachment that follows the abandonment of traditional land use has also proved a matter of some concern with respect to the decline in biodiversity ([10], [1], [2]).
Aspen (Populus tremula) stands have a fine-grained, patchy distribution in the central-southern Apennines, also reaching the island of Sicily. The understory of these stands is usually dominated by ubiquitous herbs from open habitats that are remnants of the past seral stages and persist due to the loose foliage of aspen crowns. In spite of such apparent floristic homogeneity, the composition of these forests varies more or less across ecoregions and along environmental gradients, given the wide variation in site (local) and geographical conditions in the central-southern Apennines. As a matter of fact, several forest community types (syntaxa) have been described in this geographical area: Melico uniflorae-Populetum tremulae ([28]), Fraxino orni-Populetum tremulae ([41]), Holco mollis-Populetum tremulae ([38]) and Geranio versicoloris-Populetum tremulae ([12]). In addition, a survey carried out recently on the Gargano peninsula ([40]) has revealed particular aspen stands that need to be analysed in the context of similar communities for a proper syntaxonomical assignment. To date no synthesis exists on the floristic composition and ecology of these aspen community types.
In the present study we aimed at: (i) characterising syntaxonomically and synecologically the aspen stands from Gargano; (ii) reviewing all aspen forest types from the central-southern Apennines; (iii) detecting the site/geographical factors that might be responsible for the variation in plant species composition of these woods.
Materials and methods
Study area
The Gargano peninsula is situated in the southern part of the Adriatic (eastern) side of the Italian peninsula and lies over a geological complex exclusively composed of limestones and dolomites. The area surveyed is located in the core part of the peninsula, that is on the High Plain of San Marco in Lamis and Cagnano Varano roughly between 600 and 1000 m a.s.l. The land topography is typical for karst areas, with many scattered dolines and lack of surface waters. At the lowest elevations (565-570 m) the mean monthly temperature ranges from 5.4 °C (in January) to 24.1 °C (in July), whereas the mean monthly rainfall varies between 37 mm (in August) and 125 mm (in December) summing up to a mean of 900 mm per year. According to Pesaresi et al. ([30]), the study area is included within a climatic unit characterised by a temperate oceanic bioclimate, lower supratemperate thermotype and upper subhumid ombrotype. Biogeographically the area investigated is part of the Mediterranean region, East-Mediterranean subregion, Adriatic province and Apulian sector ([35]).
The high plain is covered by secondary grasslands, abandoned fields and Turkey oak-dominated forests (Physospermo verticillati-Quercetum cerridis Aita et al., 1977 em. Ubaldi et al., 1987), the latter representing the natural potential vegetation. Dry grasslands and small Turkey oak woodlots occur on the doline slopes, whereas their bottoms are covered by aspen woods or abandoned crops invaded by Pteridium aquilinum.
Data collection in the field and from literature
The floristic survey of aspen stands from Gargano was carried out following the phytosociological approach ([44]). A single relevé (plant community sample) was performed on the bottom of each doline that was covered by a well-developed aspen canopy. In total, a number of ten relevés (SP01-SP10) were performed by visually estimating the relative cover of each vascular plant species.
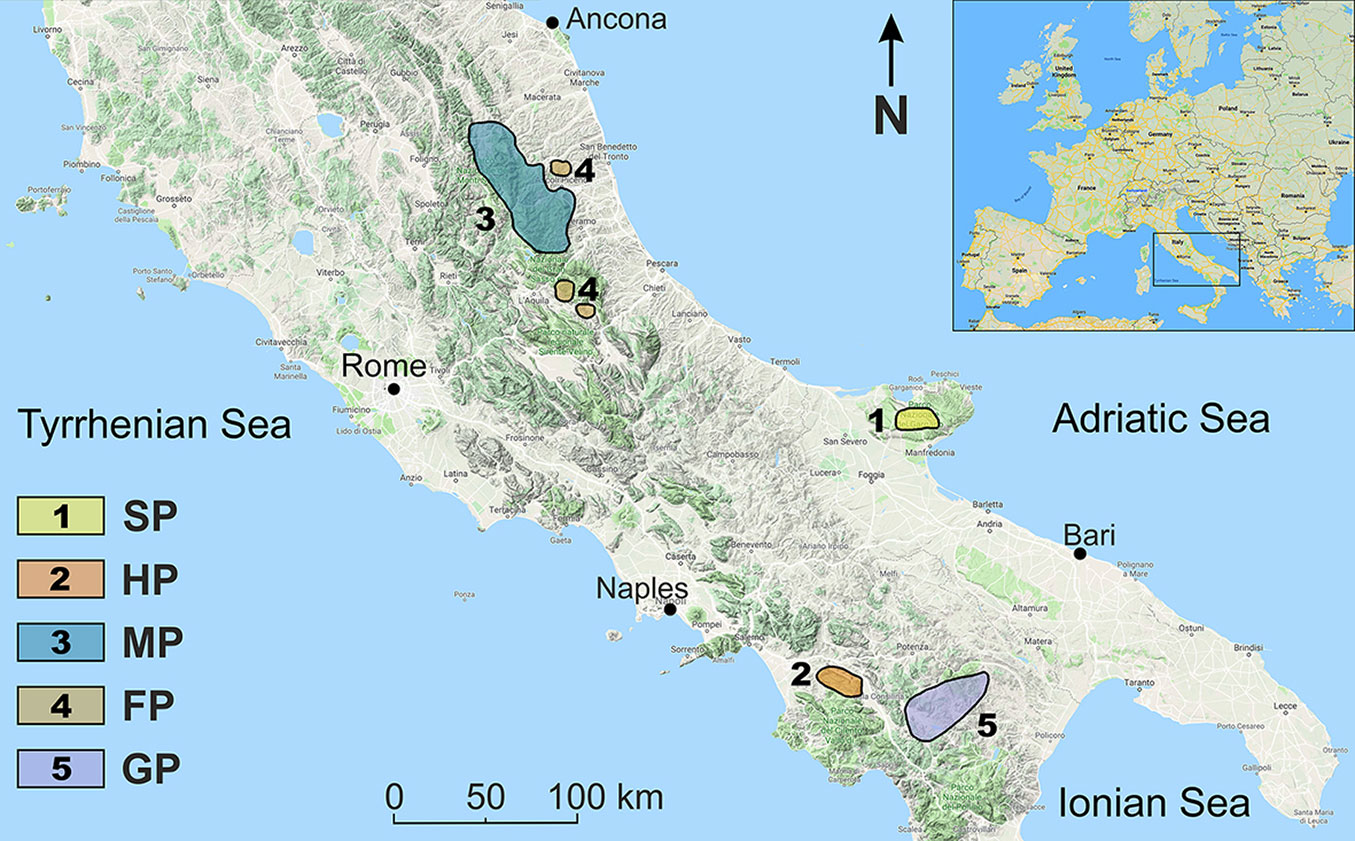
An additional set of 35 relevés, performed in aspen stands throughout the central-southern Apennines, were digitised from published literature as follows: ten relevés (MP01-MP10) of Melico uniflorae-Populetum tremulae ([28], [32]); eleven relevés (FP01-FP11) of Fraxino orni-Populetum tremulae ([41], [32]); four relevés (HP01-HP04) of Holco mollis-Populetum tremulae ([38]); ten relevés (GP01-GP10) of Geranio versicoloris-Populetum tremulae ([12]). The geographical distribution of the first two and the last two aspen syntaxa reflects to some extent a regional vicariance (Fig. 1). All aspen community types considered in this study are jointly presented in a synoptic table (see Tab. S2 in Supplementary material).
Fig. 1 - Regional distribution of the five aspen woodland types in the central-southern Apennines (Italy). Abbreviations: SP = Stellario holosteae-Populetum tremulae; HP = Holco mollis-Populetum tremulae; MP = Melico uniflorae-Populetum tremulae; FP = Fraxino orni-Populetum tremulae; GP = Geranio versicoloris-Populetum tremulae.
Elevation and terrain slope were measured at each sampled site. Other environmental variables were estimated indirectly by means of ordinal ranks of species ecological optima, namely ecological indicator values for light, air temperature, edaphic moisture, soil reaction and nitrogen ([31]).
The botanical and syntaxonomical nomenclature followed the latest online version of Euro+Med PlantBase ([9]) and respectively, the “Prodromus of the vegetation of Italy” ([3]).
Data analysis
Given the scale of observations, the emphasis on species turnover over species dominance, the inevitable bias in species cover estimates and the nature of multivariate methods subsequently applied on data, the binary community matrix based only on the presence-absence of species was employed in all analyses ([14], [33], [45]). The community dissimilarity matrix was built on the basis of the complement of the Sørensen index, which has many desirable properties ([18], [5]). Following many previous findings ([20]), the flexible beta algorithm was employed in the hierarchical clustering of relevés. The output corresponding to beta = -0.46 was finally retained as suggested by the largest value of the agglomerative coefficient (0.84). The optimal number of clusters was assessed based on the maximum value of the Calinski & Harabasz ([6]) index, as all the other internal validation criteria displayed monotonic relationships with the number of clusters. No further refinement of classification could be achieved by reallocating the relevés among groups through optimisation algorithms that maximise either the cluster average silhouette or the within-cluster / among-cluster similarity ratio ([36]). Finally, the cluster stability was estimated by the level of bootstrap mean similarity (BMS), with values larger than 70% being considered acceptable for the distinction of plant community types. The holotype of the newly described community-type was selected on the basis of the largest positive silhouette width in the target cluster. The plant species best associated with the distinguished community types were identified by means of the group-equalised, point biserial correlation coefficient (r.g), which accounts for differences in sample size (number of relevés) and enables the selection of potential diagnostic species.
The ordination of relevés in the reduced space of species was performed through local non-metric multidimensional scaling (NMDS), as it is a non-parametric, flexible technique making only few assumptions about the nature of the data. A number of three axes was extracted as suggested by the “elbow rule” applied to the distribution of the stress index with increasing dimensions ([23]). The correlation between the NMDS axes and the species occurrence was estimated through the point biserial coefficient. The importance of the environmental variables in terms of proportion of floristic variance explained (R-square) was assessed separately through non-parametric linear fitting with respect to all NMDS axes by using a modified function (“envfit.iv”), which addresses the bias in the relationship between mean species attributes (i.e., mean ecological indicator values) and relevé scores along ordination axes ([46]). The environmental variables displaying only marginal, linear effects on NMDS scores were involved in generalised additive models, in order to fit smooth surfaces through thin-plate splines in the ordination space.
The statistical significance of the above statistics was estimated by 104 random permutations. All numerical analyses were carried out in R ver. 3.6.1 ([34]) using the packages “vegan” ([27]), “cluster” ([22]), “fpc” ([16]), “optpart” ([37]) and “indicspecies” ([7]).
Results
Classification of all aspen stands
At first glance, the dendrogram of relevés revealed that these might have been reasonably grouped in two to six clusters (Fig. 2a). The classification in two clusters (distinguished at the nodes marked by black dots in Fig. 2a) would have matched exactly the regional distribution of relevés, that is in the southern and, respectively, the central Apennines (Fig. 1). Nevertheless, the maximum value of the Calinski-Harabasz index pointed to five clusters as the optimal classification of relevés (Fig. 2b). Considering also the fair to high stability of clusters, with the exception of HP cluster that included only four relevés, the solution with five groups was finally retained.
Fig. 2 - (a) Dendrogram of the relevés performed in aspen woods from the central-southern Apennines. The numbers refer to values of the stability index estimated for each of the five clusters distinguished. Abbreviations as in Fig. 1. (b) Distribution of the Calinski-Harabasz criterion by number of relevé groups distinguished via hierarchical clustering. The arrow points to the optimal number (5) of clusters.
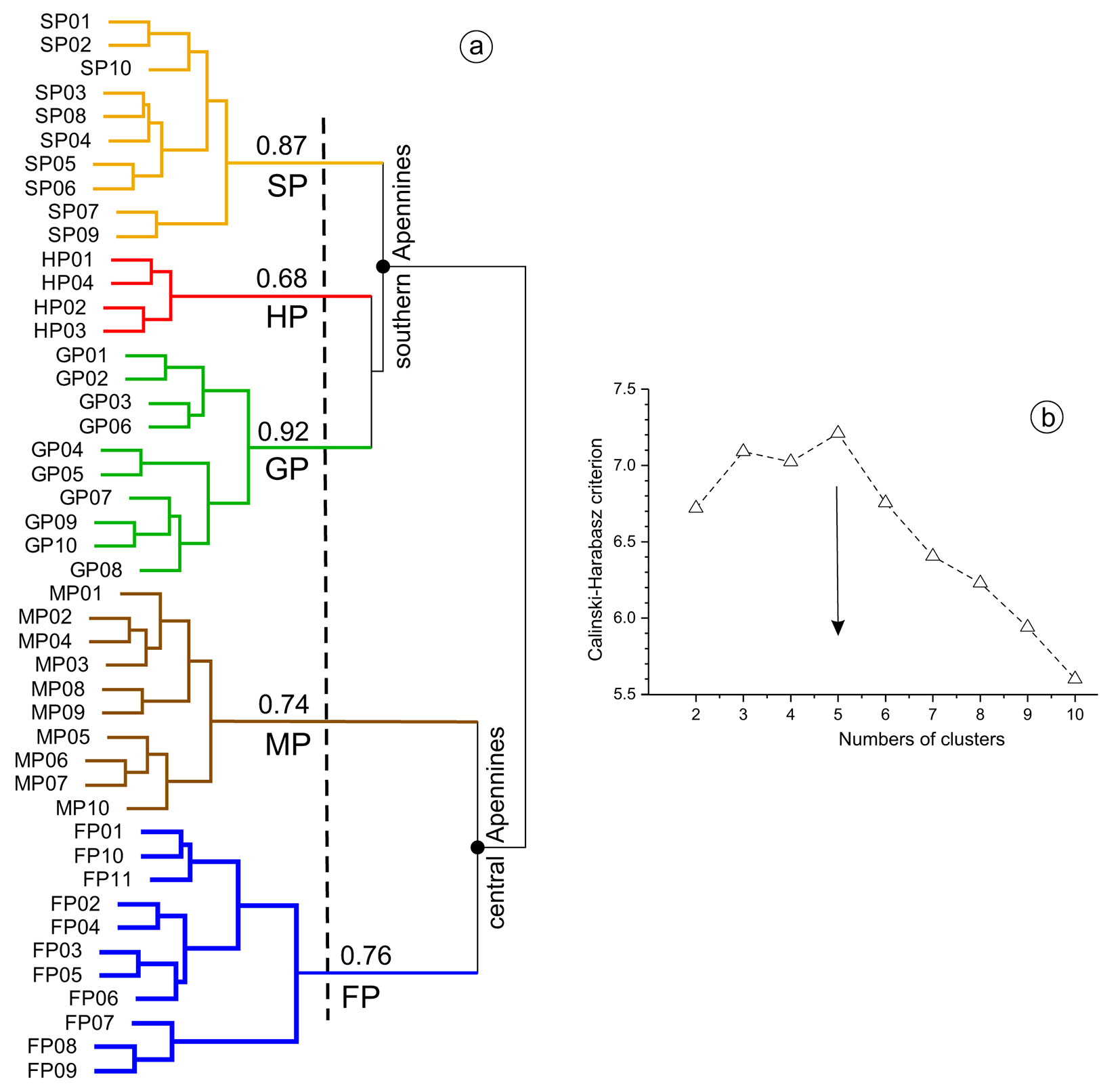
All the ten relevés performed in Gargano were gathered in one cluster (SP), whereas the other four clusters of relevés (HP, MP, GP and FP) were perfectly circumscribed to the aspen wood types described in the literature (Fig. 2a).
Typology of aspen stands from Gargano
The ten relevés from the cluster SP were assigned to a new syntaxon named Stellario holosteae-Populetum tremulae ass. nova hoc loco, within the Aceri obtusati-Populenion tremulae [41] suballiance, Corylo avellanae-Populion tremulae (Br.-Bl. ex O. Bolòs, 1973) Rivas-Martínez & Costa, 1998 alliance, Betulo pendulae-Populetalia tremulae Rivas-Martínez & Costa, 2002 order and Querco-Fagetea Br.-Bl. & Vlieger, 1937 class (Tab. S1 in Supplementary material). The relevé SP08, displaying both the largest positive silhouette and the highest mean similarity to the SP cluster, was selected as the holotype of the new syntaxon (Tab. S1). The most important understory species in terms of relative cover are Pteridium aquilinum and Rubus caesius.
Within the context represented by the other aspen wood types from the central-southern Apennines, several species displaying significant correlation with the Stellario-Populetum group of relevés were detected (Tab. 1). Of these discriminant species only three are shade-tolerant understory species (Stellaria holostea, Carex depauperata and Allium pendulinum).
Tab. 1 - Group-equalised point biserial correlation coefficients (r.g.) between species and the group of Stellario-Populetum tremulae relevés in the context of all aspen woodland types from the central-southern Apennines. Only the highest ranked ten species in terms of strength of positive correlation are listed.
| Differential species |
r.g | P-value (>r.g) |
|---|---|---|
| Vicia cassubica | 0.873 | <0.0001 |
| Stellaria holostea | 0.807 | 0.0003 |
| Poa trivialis | 0.750 | <0.0001 |
| Cruciata glabra | 0.694 | <0.0001 |
| Rubus caesius | 0.591 | 0.0003 |
| Carex depauperata | 0.590 | 0.0043 |
| Allium pendulinum | 0.590 | 0.0051 |
| Peucedanum oreoselinum | 0.590 | 0.0052 |
| Aristolochia pallida | 0.590 | 0.0054 |
| Poa pratensis | 0.590 | 0.0063 |
The refined list of differential species associated with each of the five aspen wood types is highlighted in the synoptic table featuring the percentages of occurrence of species by community type (Tab. S2 in Supplementary material). The Geranio-Populetum and Fraxino-Populetum were poorly represented by differential species compared with the other forest types.
Ordination of all aspen stands
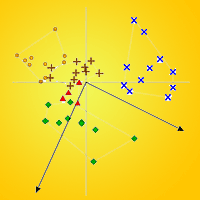
Overall, the indirect ordination of relevés in the tridimensional NMDS space revealed a fair between-group separation that matched the five clusters previously distinguished (Fig. 3). The first NMDS axis was best correlated (negatively and positively) with Geum urbanum (r = -0.79) and Juniperus communis (r = +0.57), which pointed to a nitrogen-based fertility gradient (i.e., from Fraxino-Populetum to Stellario-Populetum). The second NMDS axis was best positively correlated with Melica uniflora (r = +0.84) and negatively correlated with Agrimonia eupatoria (r = -0.60) and Dactylis glomerata (r = -0.59), suggesting a weak, canopy-openness gradient (i.e., from Melico-Populetum to Holco-Populetum). Finally, the NMDS axis 3 was best negatively correlated with Geranium versicolor (r = -0.57) but positively correlated with Acer campestre (r = +0.55), which alluded to an altitudinal gradient (i.e., from Stellario-Populetum/Fraxino-Populetum to Geranio-Populetum).
Fig. 3 - NMDS ordination of all the relevés in the reduced, bidimensional space determined by the axes 1 and 2 (a), and by the axes 1 and 3 (b). Convex hulls corresponding to each aspen woodland type are drawn by dotted lines. The environmental predictors explaining significant fractions of floristic variation are represented through vectors. Abbreviations as in Fig. 1.
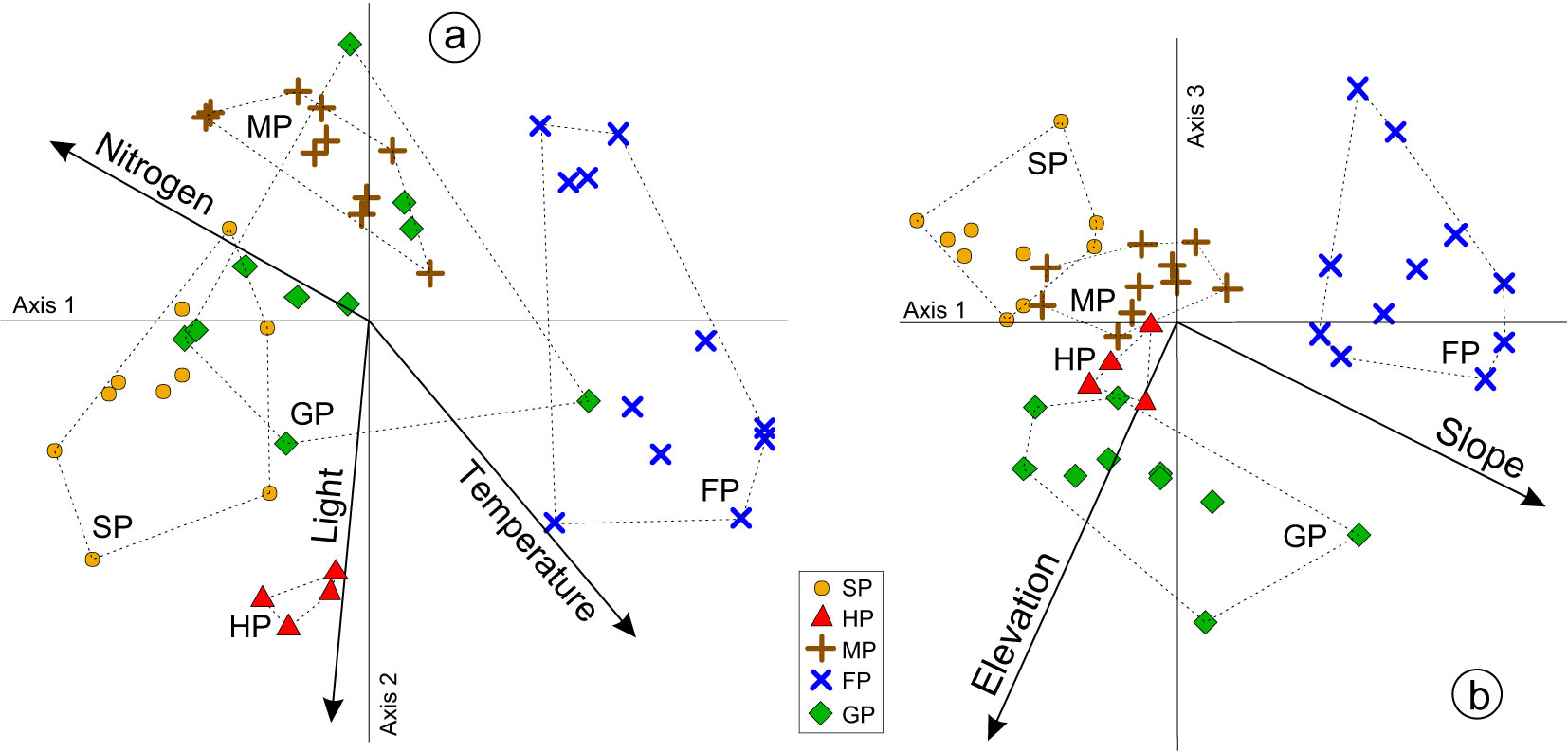
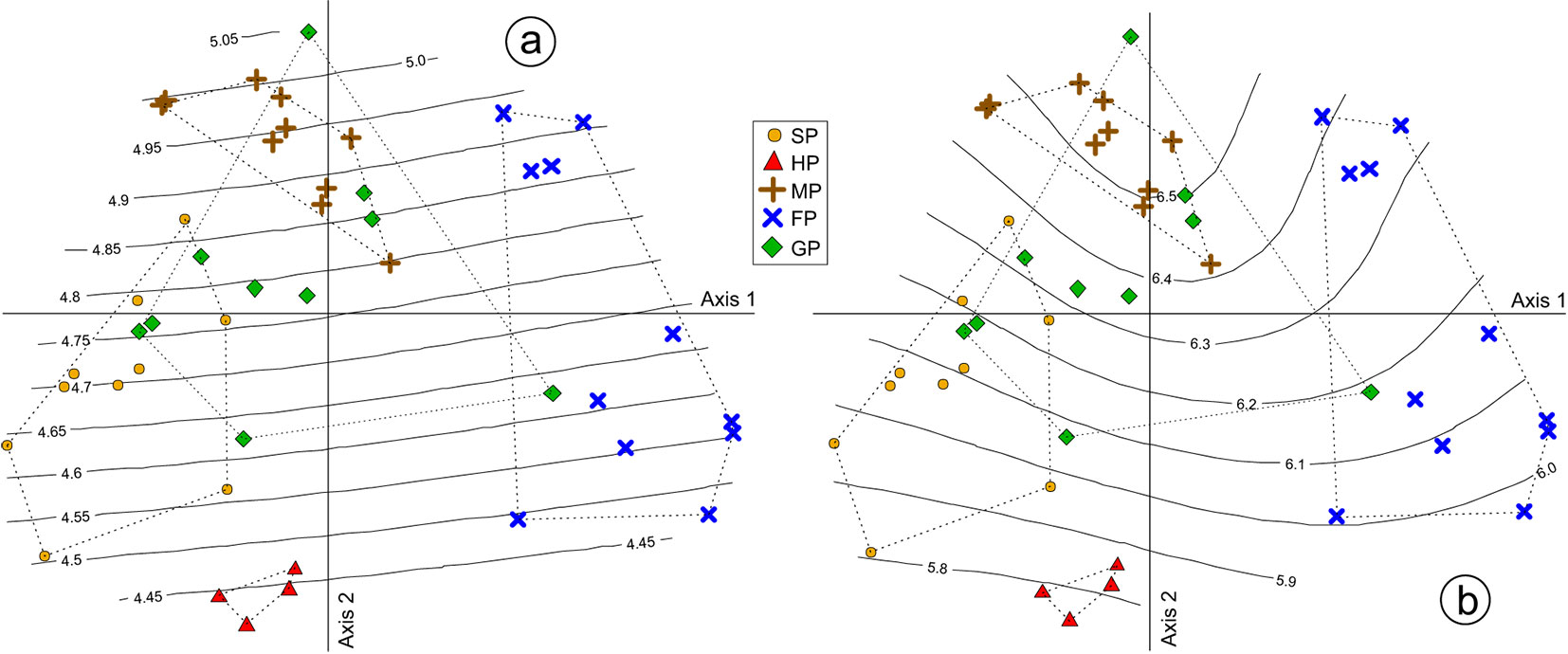
Soil nitrogen and terrain slope were linearly but oppositely related to the NMDS axis 1, whereas light and temperature were both linearly and concordantly related to the NMDS axis 2 (Tab. 2, Fig. 3a). Among the environmental variables, only elevation was linearly and significantly related to the NMDS axis 3 (Tab. 2, Fig. 3b). Although non-linear, the responses of NMDS scores to soil reaction and moisture were statistically significant and mostly oriented along the axis 2 (Tab. 3, Fig. 4).
Tab. 2 - Independent testing of the effects of different environmental variables on NMDS relevé scores by permutational linear fitting.
| Response variable |
NMDS axis 1 |
NMDS axis 2 |
NMDS axis 3 |
R2 | P-value (>R2) |
|---|---|---|---|---|---|
| Elevation | -0.373 | 0.422 | -0.827 | 0.488 | <0.0001 |
| Terrain slope | 0.758 | 0.532 | -0.376 | 0.454 | <0.0001 |
| Light | -0.088 | -0.978 | 0.189 | 0.684 | 0.0025 |
| Air temperature | 0.633 | -0.749 | 0.194 | 0.727 | 0.0007 |
| Soil moisture | -0.292 | 0.955 | -0.051 | 0.485 | 0.0673 |
| Soil reaction | 0.538 | 0.839 | 0.084 | 0.442 | 0.1009 |
| Soil nitrogen | -0.871 | 0.491 | -0.024 | 0.543 | 0.0342 |
Tab. 3 - Summary statistics of the generalised additive models employed for fitting smooth surfaces referring to indicator values of soil reaction and moisture within the space determined by the first two NMDS axes.
| Response variable |
Smooth term s(NMDS1, NMDS2) | Intercept | Adj-R2 | Deviance explained (%) |
||
|---|---|---|---|---|---|---|
| F value | P-value | t value | P-value | |||
| Soil moisture |
4.685 | <0.0001 | 168.5 | <0.0001 | 0.489 | 51.6 |
| Soil reaction |
5.466 | <0.0001 | 174.0 | <0.0001 | 0.528 | 57.9 |
Fig. 4 - Contour lines of indicator values of soil moisture (a) and soil reaction (b) along with convex hulls of aspen woodland types, overlaid on the ordination of relevés within the reduced space determined by the first two NMDS axes. Abbreviations as in Fig. 1.
Discussion
The classification of relevés drawn from the central-southern Apennines confirmed the compositional separation of the four previously described aspen community types as well as the floristic distinction of the forest stands inventoried in Gargano. It is also worth mentioning that, at low levels of compositional resemblance, the grouping of relevés in two clusters induced a sharp separation between the aspen communities distributed in the central Apennines (Melico-Populetum and Fraxino-Populetum) and those from the southern Apennines (Stellario-Populetum, Holco-Populetum and Geranio-Populetum). Such distinction at a large spatial scale was mainly due to compositional differences in the regional species pool.
The newly described “plant association” (community type), Stellario holosteae-Populetum tremulae, was assigned to the class Querco-Fagetea as these stands developed in Gargano were physiognomically tall, almost closed canopy woods and hosted several mesophilous forest herbs as well as sporadic Quercus cerris, Castanea sativa and Acer obtusatum saplings. As Stellario-Populetum tremulae belongs undoubtedly to the Apenninic suballiance Aceri obtusati-Populenion tremulae, we had to make reference (for the time being) to the upper ranked syntaxon, i.e., the alliance Corylo-Populion, despite it bears a nomen ambiguum ([25]). These authors proposed the Astrantio-Corylion avellanae Passarge, 1978 as an alternative valid name but the latter syntaxon encompasses only scrub communities, being part of the order Prunetalia spinosae Tx., 1952 and the class Crataego-Prunetea spinosae (syn. Rhamno-Prunetea). We argue that the aspen communities of Aceri obtusati-Populenion tremulae do not fit into the latter upper ranked vegetation units and therefore, the syntaxonomical framework herein should be maintained until the nomenclatural issues are sorted out.
Syndynamically, the stands of Stellario-Populetum tremulae are likely to be replaced by Turkey oak stands (Physospermo verticillati-Quercetum cerridis), which are acknowledged as the mature stage of that dynamic series ([4]). The bottoms of the shallower dolines were covered by Pteridium aquilinum-dominate communities, which presumably represented an early seral stage. Similar Turkey oak woods, but including few, more thermo-xerophilous species, should potentially succeed the Holco-Populetum tremulae stands ([38]). In contrast, the Melico-Populetum tremulae and Geranio-Populetum tremulae woods should successionally develop toward Fagus sylvatica forests ([28], [12]), whereas the Fraxino-Populetum tremulae woods should develop towards acidophilous Castanea sativa forests ([41]).
Stellario-Populetum tremulae is geomorphologically and floristically related to Holco-Populetum tremulae, as both syntaxa develop on the bottom of karstic dolines and their understory is dominated by Pteridium aquilinum. Besides, Stellario-Populetum can be considered an Adriatic synvicariant of Holco-Populetum that is distributed on the Tyrrhenian escarpment. The compositional dissimilarity between these two aspen forest types was mainly determined by some thermophilous species occurring in the understory and tree canopy of the Holco-Populetum stands. Probably on these grounds, Rosati et al. ([38]) assigned the Holco-Populetum tremulae community type to a different alliance, i.e., Teucrio siculi-Quercion cerridis.
Unlike the Melico-Populetum and Geranio-Populetum stands that possessed a larger number of mesophilous species typical of Fagetalia, the Fraxino-Populetum stands displayed a slightly more xerophilous and acidophilous character induced by the steeper slopes and expressed by the preferential presence of Carpinus orientalis, Genista tinctoria and Juniperus communis. The aspen stands, distributed on the north-eastern slope of the Turchio Mountain (Abruzzo National Park) and inventoried by Pedrotti ([29]), are also circumscribed to Fraxino orni-Populetum tremulae.
All aspen forest types are distributed on substrates moderately to rich in base cations, such as mixtures of marls and sandstones or limestones, except for Fraxino-Populetum that can also occur on slightly acidic sandstones. Despite that, the soils underlying the Holco-Populetum stands appeared the poorest in bases (and also the driest), as inferred from the plant indicator values. This is in part due to the sinkholes that have enhanced the water drainage and base leaching ([21]). On the other side, the Melico-Populetum woods are the most mesophilous, very likely because of the finer soil texture determined by the underlying marls ([42]). The second most mesophilous aspen forest type is Geranio-Populetum of which stands occur on average at higher elevations than the others.
Apart from the main environmental drivers responsible for the differentiation of the five aspen forest types, there are other factors (anthropogenic and developmental) that have induced more subtle floristic differences between these communities. First, a higher amount of soil nitrogen, either remnant from old crops or washed through rainfall run-off from the surrounding pastures, was probably responsible for the preferential occurrence of nitrophilous species such as Rubus caesius and Poa trivialis in Stellario-Populetum stands, or Aegopodium podagraria and Heracleum sphondylium in Melico-Populetum stands. Second, the larger tree canopy openness in the Holco-Populetum stands, as opposed to the relatively high cover of Corylus avellana in the Melico-Populetum stands, has determined the exclusive occurrence of shade-intolerant, thermophilous species such as Asphodeline liburnica and Aristolochia lutea in the former aspen woods.
Conclusions and implications for management
On the basis of current knowledge, five types of aspen wood can be distinguished in the central-southern Apennines. Differences in topography, edaphic conditions and light availability are the main factors responsible for the observed variation in the floristic composition of these woods. The present findings are based on a relatively low number of relevés (especially those circumscribed to Holco-Populetum tremulae) and, therefore, some observed patterns may slightly change once new data are added and analysed.
Aspen stands have little economic value because of the low density of their timber. As a consequence, the forest management should be aimed at conservation, as these pioneer woods play an important role in the recovery of forest herb diversity along the ecological succession towards hardwood forests. To address the concerns regarding the decline in biodiversity due to aspen woodland encroachment, an adequate fraction of meadows and pastures should be continuously maintained in the landscape and managed appropriately.
Acknowledgements
GR and FP carried out the field survey. DG conceived the study, performed the numerical analyses and wrote the first draft. All authors contributed to refining the initial draft.
The authors are grateful to John Akeroyd for critically commenting on and editing the latest draft of the manuscript, and to Murrja Edmir for providing the colour background of the distribution map.
Conflict of interest
The authors declare that they have no conflict of interest.
References
Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Department of Biosciences, University of Camerino, Camerino (Italy)
Department of Taxonomy and Ecology, Babes-Bolyai University, Cluj-Napoca (Romania)
Corresponding author
Paper Info
Citation
Russo G, Pedrotti F, Gafta D (2020). Typology and synecology of aspen woodlands in the central-southern Apennines (Italy): new findings and synthesis. iForest 13: 202-208. - doi: 10.3832/ifor3315-013
Academic Editor
Michele Carbognani
Paper history
Received: Dec 06, 2019
Accepted: Mar 28, 2020
First online: May 19, 2020
Publication Date: Jun 30, 2020
Publication Time: 1.73 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2020
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 40834
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 34422
Abstract Page Views: 2966
PDF Downloads: 2576
Citation/Reference Downloads: 1
XML Downloads: 869
Web Metrics
Days since publication: 2237
Overall contacts: 40834
Avg. contacts per week: 127.78
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2020): 3
Average cites per year: 0.50
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Towards a functional phytosociology: the functional ecology of woody diagnostic species and their vegetation classes in Northern Italy
vol. 14, pp. 522-530 (online: 22 November 2021)
Research Articles
Classification and mapping of Spanish Mediterranean mixed forests
vol. 12, pp. 480-487 (online: 14 October 2019)
Short Communications
Biodiversity inventory of trees in a neotropical secondary forest after abandonment of shaded coffee plantation
vol. 10, pp. 303-308 (online: 23 February 2017)
Research Articles
Growing at the forest edges: how natural regeneration develops under fragmentation
vol. 15, pp. 248-255 (online: 19 July 2022)
Commentaries & Perspectives
The role of plant sociology in the study and management of European forest ecosystems
vol. 6, pp. 55-58 (online: 21 January 2013)
Research Articles
Changes in tree layer and altitudinal distribution of herbaceous species in temperate old-growth forests over 30 years
vol. 15, pp. 206-212 (online: 11 June 2022)
Research Articles
The cork oak in the Mountains of Palermo (Italy): ecological insights from the south-eastern edge of its distribution range
vol. 13, pp. 336-344 (online: 07 August 2020)
Research Articles
Evaluation of estimation methods for fitting the three-parameter Weibull distribution to European beech forests
vol. 15, pp. 484-490 (online: 01 December 2022)
Technical Reports
Diversity pattern of vegetation in and around proposed Kotlibhel hydroelectric project along the Alaknanda River in Garhwal Himalaya (India)
vol. 4, pp. 38-43 (online: 27 January 2011)
Research Articles
Drought effects on the floristic differentiation of Greek fir forests in the mountains of central Greece
vol. 8, pp. 786-797 (online: 08 April 2015)
iForest Database Search
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords







