The cork oak in the Mountains of Palermo (Italy): ecological insights from the south-eastern edge of its distribution range
iForest - Biogeosciences and Forestry, Volume 13, Issue 4, Pages 336-344 (2020)
doi: https://doi.org/10.3832/ifor3360-013
Published: Aug 07, 2020 - Copyright © 2020 SISEF
Research Articles
Abstract
The uneven presence of the cork oak (Quercus suber L.) within its distribution range is not only determined by its climatic requirements but also by specific edaphic needs. Although most of the natural populations thrive in acidic soils deriving from metamorphic or volcanic rock outcrops, some cork oak populations are found growing in soils deriving from calcareous bedrock, which are considered less suitable. We carried out a multidisciplinary investigation at the south eastern edge of the Q. suber distribution range (Mountains of Palermo, NW Sicily), including soil, floristic, and vegetation surveys, aimed at: (i) assessing the native or introduced origin of some peculiar cork oak populations; (ii) describing the associated plant communities and soils; (iii) identifying the ecological factors which could explain the local adaptation to soils deriving from calcareous bedrock; (iv) discussing the ecological role played by this species in the study area and within its distribution range. The collected information suggest that the detected cork oak stands are native to these areas. We hypothesized that the co-occurrence of high fire frequency, high soil calcium content and erosion, which caused the intermixing of different parent materials, might favor its competitive interactions with other Mediterranean tree species, thus accounting for the local presence of Q. suber. The study of cork oak populations thriving in peculiar substrates at the driest end of the range could be of great importance for the future conservation of this species, which is expected to face growing threats in the coming decades.
Keywords
Mediterranean Evergreen Forest, Soil Chemistry, Vegetation Science, Wildfire, Landscape, Tree Species, Quercus suber
Introduction
The cork oak (Quercus suber L.) is one of the most important tree species in the Mediterranean Basin, where its presence is of high ecological, landscape and socio-economic importance ([53], [28]). Cork oak forests cover altogether more than 2 million ha ([43]), almost 1.5 million of which in Europe and 700,000 ha in North Africa ([34]). Its distribution area encompasses the western portion of the Mediterranean Basin and the southern portion of the European Atlantic coasts ([34]). The cork oak shows a discontinuous geographical distribution, much more restricted and fragmented than might be expected based on its climatic requirements. However, compared to other evergreen oaks, Q. suber is characterized by a more restricted ecological niche, as particularly demanding in terms of soil humidity ([50]). The species grows in warm-humid Mediterranean areas with oceanic features, receiving at least 450 mm of mean annual rainfall, from sea level up to 2000 m a.s.l., although most cork oak stands are found below 800 m a.s.l. ([34]).
Soil physical-chemical characteristics are regarded as being among the ecological factors most correlated to the occurrence of the cork oak in its natural range ([50]). Q. suber generally prefers sandy soils derived from siliceous rocks with acid reaction, while usually avoids soils from base-rich, calcareous bedrocks such as dolomitic limestones, calcareous sandstones, and marls ([58]). Indeed, the importance of various substrate-related variables, particularly lithology and soil type ([18]), for the distribution of cork oak has been repeatedly acknowledged. The preference for well drained soils with coarse textures is known ([38]). Conversely, reduced organic matter availability or soil acidification have negative effects on cork oak regeneration in the longer term ([24]).
There is increasing evidence of a declining trend in cork oak populations, especially at the edge of the species’ natural distribution range ([64], [43]). Many factors have been invoked to explain the ongoing process, which is bound to increase in the forthcoming decades based on current climate change scenarios ([43]), and due to other growing threats, such as overgrazing, soil pathogens and land-use changes ([21]). In the Mediterranean region, climatic forecasting seem to indicate a rise in temperatures and a reduction in precipitation, which will probably decrease the potentially suitable areas for cork oak, especially in the southern edge and at lower altitudes; accordingly, northward and upward shifts could occur ([64]). Indeed, the expected reduction in soil water availability could favor other co-occurring and more drought-tolerant woody species, such as holm oak (Quercus ilex L. - [50]). Aridity has been found to play a key role in Q. suber regeneration ability, markedly hindering seedling recruitment chance and originating over time an aged demographic structure ([43]).
In Italy, the most extensive cork oak forests occur in Sardinia (82.7% of total national area), followed by Sicily, Tuscany and Calabria, while more discontinuous and smaller nuclei are found in Campania, Lazio and Apulia ([35]). In Sicily, Quercus suber is a native species which covers approx. 19,000 ha, from sea level up to 1000 m a.s.l. It accounts for approx. 7% of total regional forest cover ([15]), in some cases characterizing old growth stands ([5]). Only 41.3% of the Sicilian cork oak stands are currently managed for production and the progressive abandonment of cork cultivation is considered to be a threat to the conservation of these ecosystems on a regional scale. Furthermore, since these populations grow at the driest end of the species’ range, they are particularly prone to the effects of climate change, and could suffer from a marked reduction and local extinction in the next future ([64]). For this reason, they are of particular interest for the future dynamics of cork oak in the whole Mediterranean Basin. In specific reference to the cork oak, according to Aronson et al. ([4]): “Knowledge of species requirements and constraints can be used to promote conservation and restoration efforts or to expand its current distribution limits in view of the current climate and land use changes”.
In this study, we aimed to gain ecological insights from the cork oak stands occurring in NW Sicily in soils deriving from calcareous bedrock, which are generally considered to be less suitable for the species. In particular, we characterized soil and plant communities associated to cork oak stands, occurring at the southeastern edge of its natural distribution range. The main objectives of the study were to: (i) identify and quantify the area covered by cork oak populations in the Mountains of Palermo (NW Sicily); (ii) investigate the effective origin of these populations (whether natural or artificial) based on historical and floristic information; (iii) identify the ecological factors responsible for the presence of the species in soils that developed on calcareous substrates; (iv) propose a synthetic interpretation of the ecological role played by the cork oak within local potential vegetation.
Materials and methods
Identification and characterization of the study areas
In order to detect any cork oak population, either as forest stand, small nucleus or isolated individual, the main botanical literature concerning the Mountains of Palermo (the mounts surrounding Palermo city) was consulted ([42]). The historical information gathered was used to identify potential areas covered by Q. suber. After being validated through dedicated field surveys, the detected areas were characterized by adopting the hierarchical classification developed by Blasi et al. ([9]); this method entailed overlapping the pedological map (scale 1:250.000 - [29]) with the geological map (scale 1:250.000 - [39]), the bioclimatic map (scale 1:250.000 - [8]), and the map of Sicilian potential vegetation (scale 1:250.000 - [1]; see Tab. S1 in Supplementary material). Three macro-areas, geographically distinct from one other and fairly homogeneous in terms of substrate (geology and soil types), were identified (Tab. 1): (i) macro-area A: “Pianetto” (municipalities of Altofonte and Santa Cristina Gela); (ii) macro-area B: “Cozzo Secco” (municipality of Borgetto); (iii) macro-area C: “Colline della media valle del fiume Oreto e del Monrealese” (municipalities of Altofonte and Monreale).
Tab. 1 - Main characteristics of the study areas. Sources: (1): Fierotti ([29]); (2): Lentini & Carbone ([39]); (3): Bazan et al. ([8]).
| Characteristics | Macro-area | ||
|---|---|---|---|
| A | B | C | |
| Locality | Pianetto (Cozzo Giammeri and Costa Zolia) | Cozzo Secco | Colline della media Valle del fiume Oreto e del Monrealese |
| Stand description | Small, isolated and discontinuous, grazed, pure stand or mixed with holm and downy oak | Scattered and discontinuous stands, grazed, mixed with evergreen thermophilous sclerophyllous species or with downy oak | Isolated plants and small nuclei, discontinuous, pure stands or mixed with evergreen thermophilous species |
| Municipality | Altofonte, Santa Cristina Gela | Borgetto | Monreale, Altofonte, Marineo, Montelepre, Giardinello |
| Regional Technical Map 1:10.000 (Section no.) |
595130 and 608010 | 594140 | 594120 and 594160 |
| Maximum range of UTM Coordinates |
E 2373518-2374629 N 4206720-4208280 |
E 2356966-2357797 N 4209665-4210804 |
E 2370475-2371368 N 4213457-4217808 |
| Area (ha) | 251 | 50 | 240 |
| Mean altitude (m a.s.l.) | 720 | 560 | 480 |
| Altitudinal range (m a.s.l.) | 597-835 | 432-659 | 240-680 |
| Slope (%) | 20 | 20 | 30 |
| Main aspect | NO-SE | SW | None |
| Soil type1 | Leptosols, Regosols, Phaeozem (Association 7) | Leptosols (Association 6) | Leptosols, Regosols, Phaeozem, Cambisols (Associations 21 and 29) |
| Lithotype2 | Calcilutites and marls, calcarenites grading up to doloarenites and dolorudites (Geology 42) | Silicoclastic deposits, yellowish graded quartzarenites and brown clays (Geology 8 and 30) | Radiolarites and Spongolites with redeposited calcarenites and calcirudites, and red marls (Geology 41) |
| Bioclimate3 | thermo-Mediterranean upper sub-humid | thermo-Mediterranean lower sub-humid | thermo-Mediterranean upper sub-humid |
The occurrence of the cork oak had already been reported in macro-area A (at Dingoli site) by Marcenò & Trapani ([42]), while cork oak forests have been present in macro-area B at least since XIII century, when the territory was owned by the Abbey of “Santa Maria di Altofonte” ([12]). Conversely, no previous historical data was available on the occurrence of Q. suber in macro-area C.
Subsequently, we searched for local data on wildfires, as they are an important factor explaining the natural distribution of the cork oak in the Mediterranean, as well as in our study areas. Information collected by the Special Office of the Firefighting Forest Fire Service ([63]), for the period preceding our field surveys, and by the Command of the Forest Service of Sicily for more recent data (2007-2019 - ⇒ https://sifweb.regione.sicilia.it/arcgis/rest/services/Censimento_Incendi/MapServer), was considered. The oldest data are sorted by municipality and they provide information on the occurrence of wildfires in Sicily in the years 1986-2002 ([63]). In detail, in the framework of the Regional plan for the protection of vegetation from fires, a regional map of fire risk was developed, not only considering the factors which enhance the likelihood of wildfire (e.g., vegetation types or climate traits), but also taking into account the frequency of wildfires occurring in a given area, as well as the average affected area and the average affected wooded area, thus including a quantitative assessment. In particular, this map was elaborated through the overlapping of different base maps, including a statistical risk map, a vegetation risk map, a climate risk map, and a morphological risk map.
For a climatic and bioclimatic characterization of the study areas, the thermopluviometric stations of Partinico (189 m a.s.l.) and Monreale (310 m a.s.l. - [27], SIAS, ⇒ http://www.sias.regione.sicilia.it/) were considered the most suitable (Tab. 2). Taking into account their location, the Partinico station was used to characterize macro-area B, while the Monreale station was used for macroareas A and C. Annual precipitation ranged from 706 mm (Partinico) to 991 mm (Monreale), while mean temperature ranged from 16.2 °C (Monreale) to 17.8 °C (Partinico). Raw climatic data were used to identify the duration of the drought-stress period, which generally spans from the third ten-day period of April to the third ten-day period of September in both stations. According to Rivas-Martínez ([55]), as well as local classification schemes ([8]), the bioclimate for macro-area B is thermo-Mediterranean lower sub-humid, while for macro-areas A and C, the bioclimate is thermo-Mediterranean upper sub-humid.
Tab. 2 - Precipitation and temperature data in the considered thermo-pluviometric stations.
| Variable | Parameter | Meteo Stations | |
|---|---|---|---|
| Partinico | Monreale | ||
| Precipitation (mm) | Mean annual | 706 | 991 |
| Monthly maximum | 97 | 129 | |
| Monthly minimum | 4.3 | 4.7 | |
| Temperature (°C) | Mean annual | 17.8 | 16.2 |
| Average of the maximum | 22.6 | 21.3 | |
| Average of the minimum | 12.7 | 11.4 | |
| Maximum average of the hottest month | 31.8 | 31.1 | |
| Minimum average of the coldest month | 6.6 | 5.3 | |
The cork oak distribution map
A distribution map of the cork oak, either found as forest stand, small nucleus and/or isolated individual, was made in a GIS environment. Field data were registered by means of a Trimble® GPS (Trimble Inc., Sunnyvale, CA, USA). Data referring to the UTM plane coordinate system, timed to share information, latitude and longitude, were exported as shapefiles to be treated with QGIS® ver. 3.10 (QGIS Development Team) and were overlapped with sections no. 594140, 595130, 594120, 594160 and 608010 of the Regional Technical Map (scale 1:10.000). The shape relative to punctual distribution files of cork oak was then overlapped with the soil, geological and bioclimatic maps.
Soil surveys
The choice of the most suitable areas for the pedological surveys was based on two steps: after superimposing the cork oak distribution map with the geological map, direct field observations of the soil features and plant species assemblages (acidophilous vs. basiphilous) were carried out. The benchmark pedons (=soil profiles) representative of the main soil types (Tab. 1) were described in the field after direct soil excavation and according to the genetic horizons defined by Schoeneberger et al. ([56]). Overall, 5 pedons, sampled in bulk and undisturbed soil horizons, were described: four pedons belonging to macro-area A, and one pedon belonging to macro-area B. No pedon was considered in macro-area C because it was not possible to access the privately owned areas. Pedon 1 (Code Ag1) and pedon 2 (Code Ag2) were located at Cozzo Giammeri (38° 00′ 23.80″ N, 13° 20′ 34.41″ E), pedon 3 (Code Az3) and pedon 4 (Code Az4) were located at Costa Zolia (37° 59′ 50.75″ N, 13° 20′ 49.64″ E), and pedon 5 (Code B) was located at Cozzo Secco (38° 01′ 31.15″ N, 13° 09′ 07.21″ E). For each pedon, three soil samples were collected at several soil depths. Bulk soil samples were air dried, crushed and 2-mm sieved. Particle size distribution was determined by sieving and sedimentation (ISO 11277:2009): total carbonates volumetrically, pH potentiometrically (ISO 10390:2005) and soluble ions by ion chromatography of saturated-paste water extracts (Dionex DX-120®, Sunnyvale, CA, USA). Soil classification followed the IUSS Working Group ([36]).
Floristic and vegetation surveys
Floristic and phytosociological surveys were carried out during spring 2004. Analysis of cover-value ranges available from vegetation surveys on evergreen forests in Sicily (see [13] and literature therein) suggested the use of 20% as a valuable threshold for the cover of Q. suber. Full vegetation surveys, within 100 m2-wide plots, were carried out only within the sub-populations where cork oak cover exceeded 20%. Four plots were established in macro-area A, and seven plots were established in each of the macro-areas B and C, which included the most interesting cork oak stands. For each plant species, the cover/dominance scale proposed by Braun-Blanquet ([11]) was considered, with modifications subsequently proposed by Pignatti & Mengarda ([52]). The Shannon diversity index (H’ - [59]) and Evenness index (Pielou J) were calculated after converting Braun-Blanquet values according to Tüxen & Ellenberg ([62]). Classification of the vascular plants followed Pignatti et al. ([51]) and hierarchical classification of the plant communities followed Guarino et al. ([31]). Each plant species surveyed was included in one of the following phytosociological alliances: (i) Erico-Quercion ilicis, including pure evergreen or mixed ever- and summer-green oak forest species linked to neutrocline-acid soils (EQi); (ii) Quercion ilicis, including pure evergreen or mixed ever- and summer-green oak forest species (Qi); Oleo-Ceratonion siliquae, including maquis species (OCs); (iii) Pruno-Rubion ulmifolii, including mantle species (PRu); (iv) Cisto-Ericion multiflorae, including garrigue species (CEm); (v) Hyparrhenion hirtae, including xerophilous grassland species (Hh); (vi) Other species belonging to other syntaxa.
Where only a few scattered individuals of cork oak occurred in open and discontinuous plant communities (cork oak cover <20%), we considered only the presence/ absence of diagnostic vascular plants typical of forest and pre-forest communities (maquis, mantle, garrigue and grassland), as well as of species characteristic of rocky and scree habitats. In these areas, we aimed to highlight whether a high number of species characteristic of Erico-Quercion ilicis could support the past occurrence of cork oak forests. The authors of all scientific epithets concerning plants mentioned in the text are provided in Tab. S2 (Supplementary material).
Statistical analysis
To test the effects of the environmental variables, namely altitude, main aspect (north or south) and lithology (calcareous or flysch) on richness, diversity (Shannon H’) and evenness (Pielou J), we used a general linear model (GLM). Before running the model, we first checked that the three dependent variables followed a normal distribution (Shapiro-Wilk test: p = 0.55, 0.23 and 0.85, respectively). Spatial autocorrelation across the sampling plots using Moran’s I index, calculated from three different weight matrices (i.e., distance to the nearest neighbor) was also checked ([25]). We set the distance range at 2300 m, which was the minimum distance to allow all plots to have at least one neighbor. We then set the range at 4300 m to connect all plots inside the same macro-area, and finally we used a “queen” matrix, searching for the first closest neighbor without a maximum distance threshold, thus connecting at least one neighbor from all macro areas. To test for differences in richness, diversity, and evenness between the three macro-areas, we used one-way ANOVA, after verifying the homogeneity of variances in the residuals of the model (Levene’s test: p = 0.69, 0.50 and 0.12, respectively). All statistical analysis was performed in R v. 3.6.1 ([54]) and spatial autocorrelation was assessed with GEODA 1.8.16 ([2]).
Results
Distribution and characterization of the study areas
The distribution map of the areas where cork oak occurs in the Mountains of Palermo is shown in Fig. 1. Macroarea A is mainly characterized by calcilutites, marls, and calcarenites grading up to doloarenites and dolorudites, and Leptosols, Regosols, and Phaeozem ([29], [39]). Macroarea B is mainly characterized by Silicoclastic deposits, Numidian Flysch, and mainly Leptosols ([29], [39]). Macroarea C is mainly characterized by Radiolarites and Spongolites with redeposited calcarenites and calcirudites, red marls, and Leptosols, Regosols, Phaeozem, and Cambisols ([29], [39]).
Fig. 1 - The cork oak distribution map in the Mountains of Palermo. The different letters indicate the three macro-areas (A, B and C). Up on the left, the natural distribution range of Quercus suber is reported ([28]).
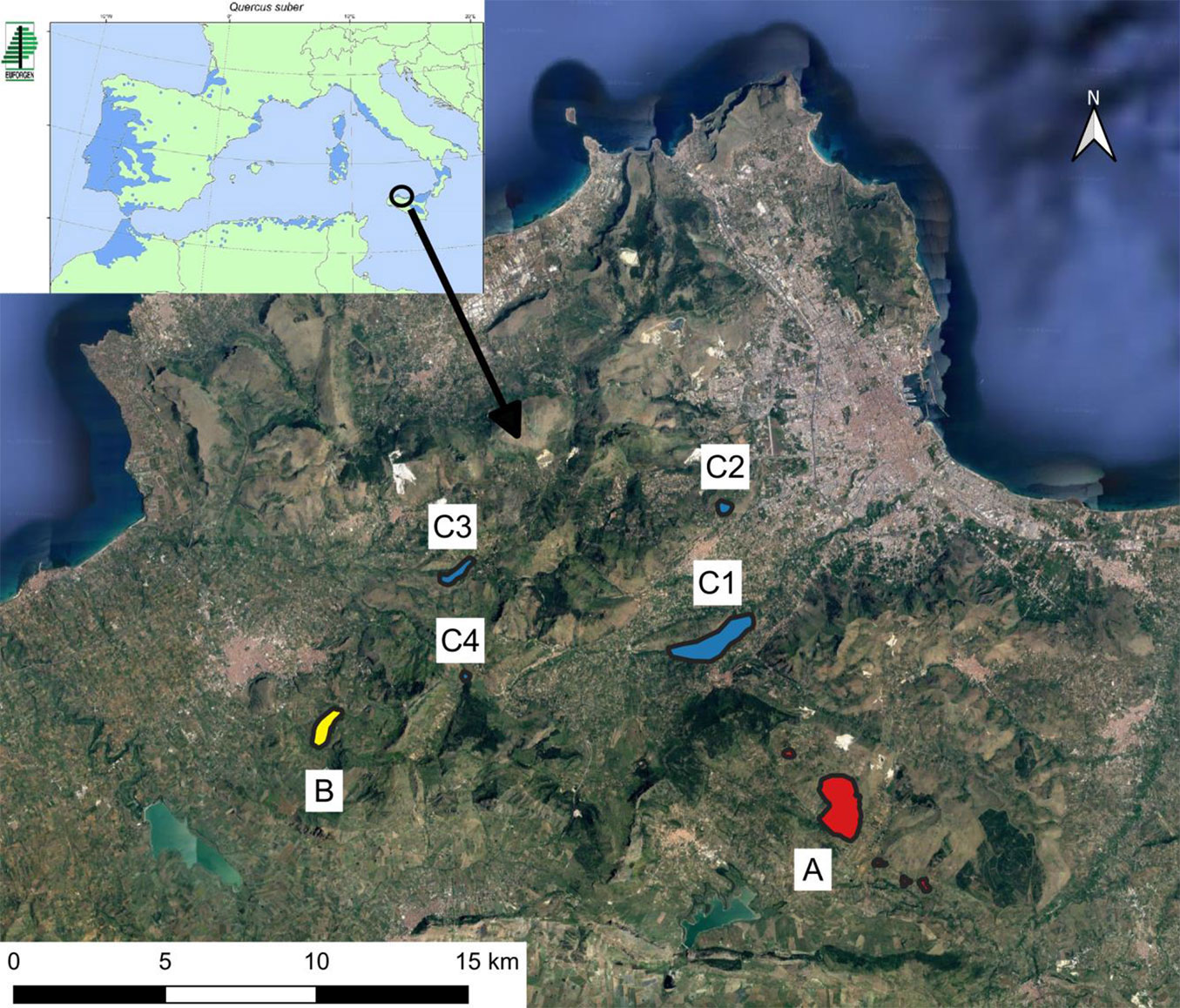
Macro-area A has a surface of about 251 ha and is localized in the south-east sector of the map. Macro-area B is around 50 ha wide and is localized in the south-west sector of the map. Macro-area C is approx. 240 ha wide, encompassing much more fragmented and isolated stands, localized in the northern sectors of the map. Overall, the areas under investigation, covered by more or less dense cork oak forests, cover approximately 540 ha. Concerning fire occurrence, all investigated areas fall within areas classified as high or very high fire risk, according to the regional classification scheme ([63]). This means that these areas not only have a high likelihood of suffering from wildfire but they have been effectively burned with high frequency at least at the end of the last century ([63]). Such a trend has been confirmed by recent data (2007-2019), showing a high mean annual frequency of wildfire passage, ranging from one wildfire every two years elsewhere in macro-area B to one wildfire per year elsewhere in macro-area C. Hence, although the total area is considerably large, the intense human pressure may represent a serious threat for the conservation of these forest ecosystems.
Soil surveys
Most of the investigated soil profiles showed marked limitations in root growth, mostly belonging to Leptosols. The five pedons showed a taproot plus shallow horizontal roots. The main portion of root biomass is located in the root crown just below the trunk base at a soil depth < 0.3 m. As we sampled by horizontal profiles, the horizontal extension of the whole root system, including roots thinner than 1 cm, is unknown. In macro-area A, pedon 1 and pedon 2 (Ag1 and Ag2) belonged to Leptosols / Regosols (sporadically Phaeozems) soil associations, while pedon 3 and pedon 4 (Az3 and Az4) belonged to Leptosols. These soils evolved from lithological substrates made of siliceous shales, radiolarites and marls, with interbedded lenticular resedimented breccias. Graded and laminated calcareous sandstone with elements from Middle-Upper Triassic Cretaceous platforms occurred in pedon 3 and pedon 4. Cozzo Giammeri profiles (Ag1 and Ag2) evolved on slopes with a gradient of 15 and 25%, respectively, while Costa Zolia profiles (Az3 and Az4) evolved on slopes with a gradient of 10%. The pedon differed in terms of stoniness and rockiness. The main land use was overgrown pastureland. In macro-area B, pedon 5 belonged to Leptosols, which evolved on lithological substrates made of graded doloarenites and dolorudites, and re-sedimented dolomite breccias with megabreccias from Triassic carbonate platforms. This profile was open on one side in a slope with a gradient of 35%, stoniness of 25%, rockiness of 20% and moderately rapid drainage. A morphological description of all pedons is given in Tab. S3 (Supplementary material). Average soil pH ranged between 6 and 7 in all study sites. Variability of soil pH increased almost linearly with a soil depth of up to 80 cm (ranging from 5.0 to 7.8); conversely, it decreased progressively at higher soil depths (Fig. 2). Overall, the large variability in soil pH (from 4.6 to 7.8) showed good plasticity of cork oak to this edaphic factor. Na/Ca molar ratios were constantly higher than 1, Mg/Ca ratio exceeded 0.6, while Na/K ratio exceeded 20 (Fig. 3).
Fig. 2 - Soil pH variation with soil depth (average of pedons 1-5). The dashed line represents the median of pH values while whiskers indicate the range. The shaded area represents the range of soil pH in Quercus suber stands found in literature.
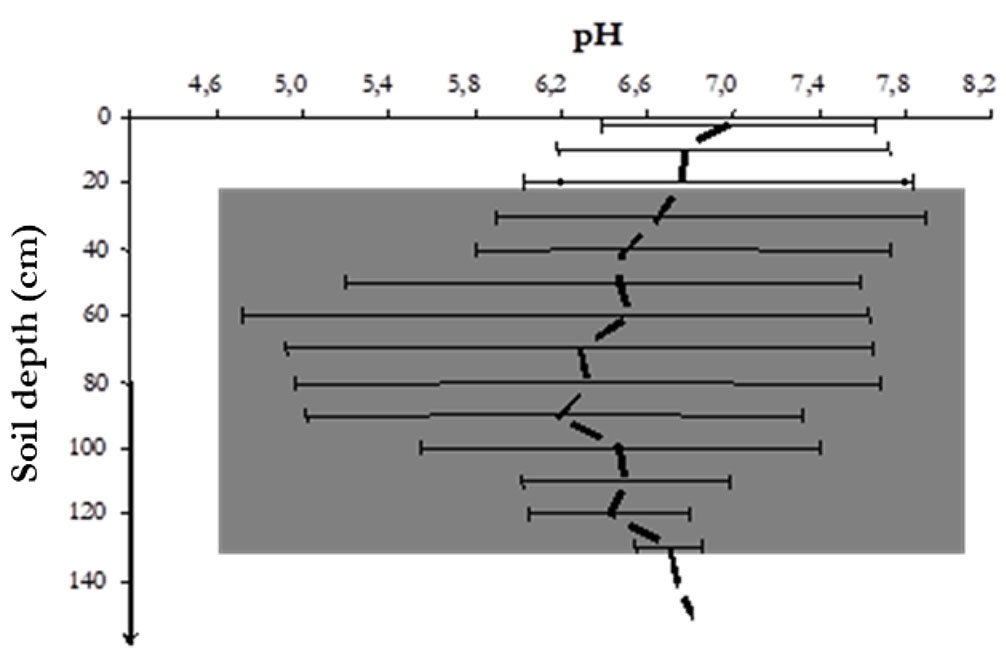
Fig. 3 - Ionic species within the saturated extracts from soils. Average concentrations in nM (x-axis) and depth (y-axis) in cm.

Floristic and vegetation surveys
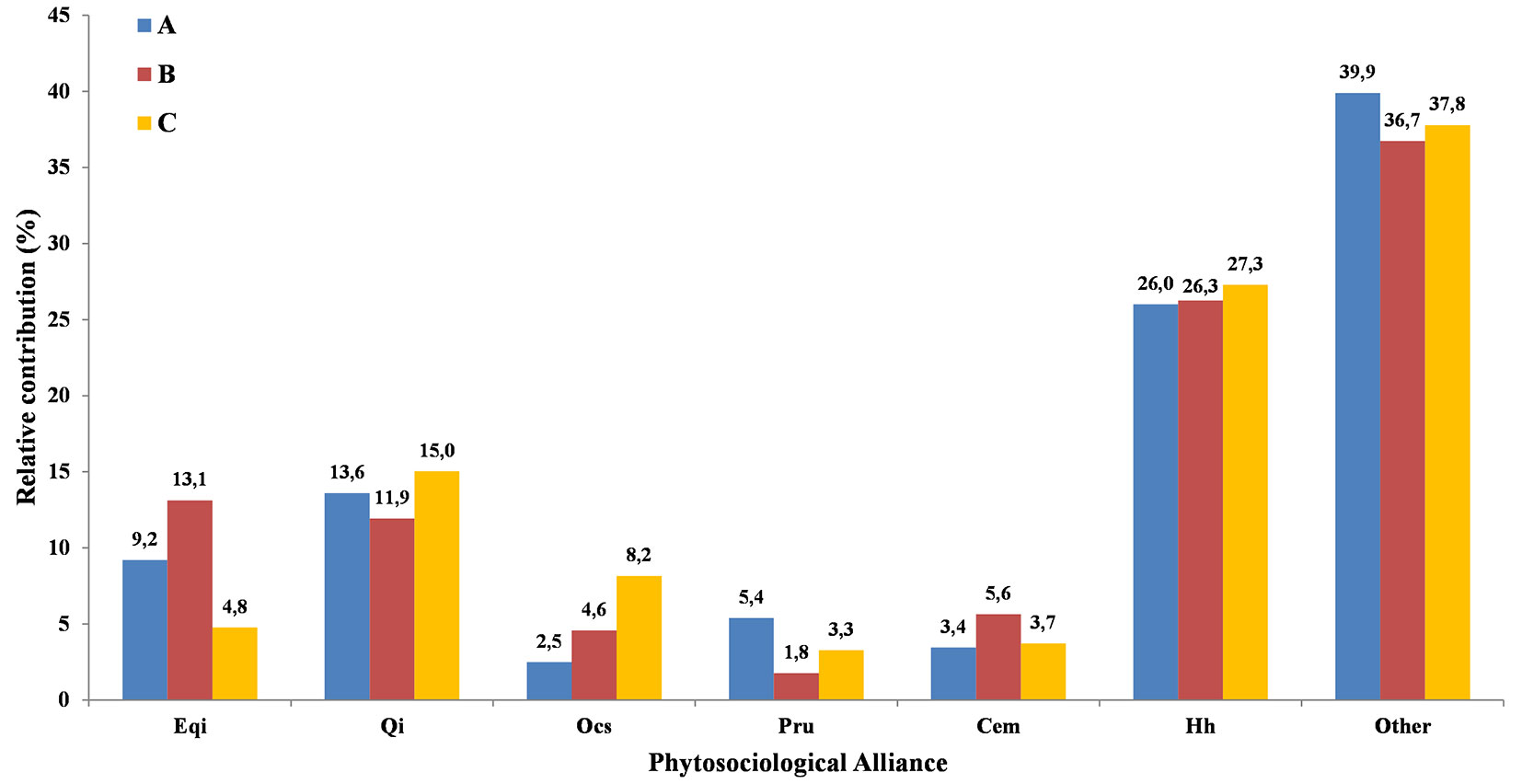
Despite evenness being autocorrelated in the two close range matrices, source variable richness as well as diversity were not autocorrelated in all matrices (Moran’s I p > 0.05 in all cases - Tab. S4), suggesting high local scale variability. GLM indicated that richness, diversity and evenness are not predicted by altitude, aspect and lithology (p > 0.05 in all cases - Tab. 3), and they did not differ between the macro areas (richness: F = 0.24, p = 0.78; diversity: F = 0.74, p = 0.49; evenness: F = 1.01, p = 0.38). However, significant differences (F = 7.92, p < 0.01) were found in the richness of species belonging to the alliance dominated by Q. suber (EQi), which was significantly higher in macro-area B than in C, although not differing from A (after Tukey HSD test). High degradation and fragmentation of forest habitats in the surveyed areas can be inferred from the number of plant taxa not framed within any alliance (see “Other Species” in Fig. 4 and Tab. S2 in Supplementary material), as well as from those belonging to the alliance Hh. In all macro-areas, plant taxa belonging to “other species” represented the dominant component of plant communities, ranging from 38.6% in macro-area B, 39.8% in C and up to 42.5% in A (Fig. 4).
Tab. 3 - Summary of the results of the GLM testing the effects of the three environmental variables on richness and diversity.
| Variable | Richness (R² = 0.22) | Diversity (R² = 0.13) | Evenness (R² = 0.05) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Estimate | t | p | Estimate | t | p | Estimate | t | p | |
| Intercept | 35.506 | 3.1 | <0.001 | 2.019 | 5.72 | <0.001 | 0.558 | 8.26 | <0.001 |
| Altitude | 0.028 | 1.36 | 0.2 | 0.001 | 0.83 | 0.42 | 0 | 0.49 | 0.63 |
| Aspect | -0.027 | 0.02 | 0.98 | 0.003 | 0.03 | 0.98 | 0.002 | 0.11 | 0.91 |
| Lithology | 6.64 | 1.86 | 0.08 | 0.155 | 1.41 | 0.18 | 0.018 | 0.83 | 0.42 |
Fig. 4 - The relative contribution (in %) of each surveyed phytosociological alliance within each macro-area (A, B and C). (EQi): Erico-Quercion ilicis; (Qi): Quercion ilicis; (OCs): Oleo-Ceratonion siliquae; (PRu): Pruno-Rubion ulmifolii; (CEm): Cisto-Ericion multiflorae; (Hh): Hyparrhenion hirtae; (Other): species belonging to other syntaxa.
Macro-area A is characterised by a complex and heterogeneous landscape, due to extreme variability in local topography (aspect and slope), continuous shifts from carbonate (compact calcarenites and calcilutites) to acid (quartz sandstones) outcropping rocks, as well as to major disturbance factors (mostly cattle grazing and fires). Here, cork oak stands, pure or mixed with a number of evergreen or deciduous trees, are intermingled with more open thermophilous maquis on S-facing ridges and summits. Quercus suber does not seem to play a dominant role in the absence of human disturbance. These forest communities show excellent potential for recovery and future development, although the possibility of establishing the potential vegetation (Quercetalia pubescenti-petraeae) with downy oak as the dominant tree species ([1]) is currently low. In fact, the current floristic composition of local communities is rich in plants linked to rocky habitats and overgrazed pasturelands, showing clear signs of soil degradation and thinning processes. Some cooler and shady microhabitats are suitable for Cephalanthera longifolia, Quercus congesta and Iris foetidissima, which are rare species on the Mountains of Palermo ([48]).
In macro-area B, the cork oak stand is uneven-aged, almost pure, mostly sparse and discontinuous, and rather plant species poor. High wildfire frequency is witnessed by the extremely high number of species tied to open patches of perennial grassland dominated by Ampelodesmos mauritanicus or of acidophilous garrigue dominated by Cistus spp. and Lavandula stoechas, while the current intensity of cattle grazing is underlined by high numbers of mantle and grassland species. Only near rocky outcrops, especially along ridges and on hilltops, a higher frequency of Mediterranean maquis species was recorded. In small valleys, where soils are deeper and the effect of wind is slightly mitigated, some scattered individuals of downy oak (Quercus pubescens Willd. s.l.) co-occur with cork oak, while the shrubby understory is dominated by Cytisus villosus. This plant community may be framed within the Erico-Quercion ilicis, due to the high number of species referred to this alliance, in addition to acidophilous species such as Lavandula stoechas, Limodorum abortivum, Pteridium aquilinum, and Tuberaria guttata. We may hypothesize that local forest communities belonged in the past to the Erico arboreae-Quercetum virgilianae, rather widespread in the Tyrrhenian sector of Sicily ([13]).
Macro-area C is subdivided into four sub-areas geographically disjoint from each other (Fig. 1). The most interesting cork oak stands occur in the southeast sub-area, where the semi-natural landscape appears rather uniform: a series of ridges whose S-facing slopes usually host thermophilous taxa and assemblages, while N-facing slopes host some remnant nuclei of Mediterranean oak forests. It is therefore quite difficult to assign local plant assemblages to a plant association. On more acidic substrates, like quartz sandstones and argillites, local forest communities probably belonged to the Erico-Quercion ilicis alliance, or alternatively to the Erico arboreae-Quercetum virgilianae plant association. Elsewhere, the high frequency of woody species typical of Oleo-Ceratonion, which represents the potential vegetation of this area ([1]), supports the hypothesis that cork oak locally belonged to the Oleo sylvestris-Quercetum virgilianae. Most of the cork oak stands in this macro-area are very scattered and open, forming a patchwork with xeric perennial grasslands dominated by Ampelodesmos mauritanicus or with mantle communities physiognomized by Crataegus monogyna, Cytisus infestus, Osyris alba, Daphne gnidium, Spartium junceum, Pistacia terebinthus and P. lentiscus. Intense and widespread soil erosion processes, and frequent wildfires caused extreme soil and habitat degradation. The high degree of soil thinning may also be inferred by the high frequency of species adapted to cliffs, ledges and screes, as observed in the small stand of “Contrada Loghi Vecchi” (Monte Caputo - see Tab. S5 in Supplementary material).
Discussion
Our study provides ecological insights from Quercus suber stands occurring on the Mountains of Palermo, at the southeast edge of its distribution range. The pedological characteristics obtained by direct soil excavation and pedon observation ([17]) allowed to infer some interesting indications on the ecological range of Q. suber in terms of soil variables. Recent research has emphasized the need for very detailed field data when studying cork oak stands ([64]). The same is true for our study sites, as the calcareous nature of the substrates is different from the typical siliceous rocks with acid reaction, where most of the cork oak stands occur.
Our study sites fall within areas considered to be of particular interest for the future dynamics of cork oak, as they could face growing risk of local extinction, or a significant reduction in the population size ([64]) due to many interacting detrimental factors (climate change, fire frequency, etc.) driven by man ([43]). Our field observations and wildfire data clearly proved that the surveyed cork oak stands are highly exposed to these risks.
The occurrence of species typical of the Erico-Quercion ilicis (EQi) may represent an effective proxy for assessing the native status of local Q. suber populations. A high frequency of base-avoiding plant species typical of the EQi was recorded in the understory of the surveyed cork oak stands. Since many of these species show high fidelity to cork oak stands throughout the whole island ([13]), all the macro-areas have very good potentiality for Q. suber. As historical data ([12], [33], [40], [46]) testify the local occurrence of Q. suber at least since the Middle Ages, and considering that the first afforestations in Sicily started around 1920s, cork oaks should be considered locally native in these areas and the hypothesis of a past introduction in pre-existing woodlands should be discarded.
Cork oak stands growing on alkaline or neutral-alkaline soils have been already observed in western and southeastern Sicily ([44]), as well as in southern Latium (central Italy), where they have been interpreted as steady “disclimax” communities favoured by century-long fire disturbance to the detriment of local “climax” forests dominated by holm and/or downy oak ([10]). In our study areas, three main factors, probably acting in synergy, could account for the scattered but persistent occurrence of Q. suber: (i) local fire disturbance; (ii) the peculiar characteristics of local soils; and (iii) the effect of erosion processes on the local geological framework.
High fire frequency, as observed in all macro-areas, could have hindered evolution towards more mature forest ecosystems, favouring the more fire-resistant cork oak, once a minor component of pristine mixed oak forests ([6]). Indeed, Q. suber is a fire-resistant tree species with a thick bark acting as an insulating material and protecting the inner woody structures. After fire passage, the cork oak is also able to resprout both from stem and basal buds ([49]), showing a notably high survival rate ([61]). The marked effect of human disturbance on local plant communities can also be derived by the fact that none of the abiotic factors assessed (altitude, aspect and lithology) proved to be a predictor of the observed richness, diversity and evenness of the cork oak stands.
Regarding pedological traits, we found large plasticity in Q. suber to substrate conditions, including the pH of the rhizosphere and the geological substrate. Average soil pH ranged between 6 and 7 in all study sites, which is consistent with values reported in the literature ([19]). Our results also match the preference of Q. suber for soils with acid or sub-acid reaction, together with a tendency to avoid limestone soils ([26]). However, the total calcium content in soils and its active portion should be distinguished. The cork oak, like the sweet chestnut (Castanea sativa Mill.) and the common bracken (Pteridium aquilinum (L.) Kuhn), may tolerate a rather high content of soluble limestone ([45]). Interestingly, recent research has found natural cork oak populations growing in soils with an active lime content below 26% and soil pH close to 8 ([26]). Q. suber rarely develops in podzols and regosols ([60]), whereas more frequently is found in cambisols ([3]) with a moderately thick humic horizon and subacidic soil pH. In the pedon profiles, the presence of nitrates proves that rhizosphere activity is mostly concentrated in the first 30 centimeters of soil depth. Subsequently, the scarcity of phosphorus, the excess of calcium and sodium, counterbalanced by similar concentrations of chlorides and sulphates, characterize all the pedon profiles. The average depth of the described pedons falls within the range reported in the literature ([22]). Soluble ions, particularly sodium, do not disturb the cork oak, though excess calcium most likely does ([41]). In fact, at each depth, molar ratios Na/Ca were always higher than 1, while Mg/Ca ratio was at maximum 0.6, and Na/K ratio was permanently higher than 20. It is plausible that the high Ca content, deriving from parent material, may represent a major obstacle for the colonization of forest species typical of more mature stages, thus favouring the cork oak. Furthermore, cork oak may produce changes in soil nutrient content, particularly N and Ca, affecting the composition of soil microbial community ([3]). This would explain the good establishment and natural regeneration even on dolomitic limestones found on Mt. Gibilmesi, Mt. Petroso and Costa Sant’Anna near Mt. Cuccio, where Q. suber was planted during the second half of the twentieth century ([20]). In agreement with its general ecological behaviour, on the Mountains of Palermo Q. suber seems to play a dominant role only in macro-area B, characterized by acidic soils and rocky outcrops. This finding suggests some peculiar adaptation to local conditions in the other macro-areas; for instance, the ability of cork oak seedlings to withstand iron deficiency has been proved ([30]).
As far as soil erosion is concerned, the calcareous substrates colonized by Q. suber are in tight contact with (and once were underneath) the flysch outcrops, mainly quartz sandstones and argillites ([16]). Due to the heavy impact of human activities since prehistoric and protohistoric times, the upper lithological layers (flysch) may have been eroded, remaining obliterated or reduced to small pockets in cracks of the underlying carbonate layers. In any case, if the general conditions for establishment of the species exist, the plasticity of the cork oak ([41]) may induce deep transformations on soil characteristics over time.
The issue of how to interpret the dynamic role played by cork oak in the vegetation of the thermo- and meso-Mediterranean is still under debate ([13]). In the progressive succession of the Mediterranean vegetation, many authors ([23]) are in agreement when considering the cork oak stands as an edaphic and stable climax, linked to loose soils with an acid reaction, and assigning them to the Quercion ilicis alliance. Conversely, other authors framed them within the Erico-Quercion ilicis, which includes all the evergreen western Mediterranean forests dominated by evergreen oaks ([13], [32]). Within its natural distribution range, including Sicily, the cork oak seems chiefly to characterize sub-optimal facies compared to local climax forests, whereas only rarely playing a primary role ([57]). However, due to the total lack of examples of undisturbed forest vegetation within the whole range of the Mountains of Palermo ([47]), forecasting the possible dynamic evolution of cork oak stands is rather complex, making impossible to assess which were the most mature “final” forests adapted to local edapho-climatic constraints. Mixed oak woods must have been the rule before man favoured holm oak, cork oak and manna-ashes (Fraxinus ornus) to the detriment of deciduous oaks (e.g., Quercus pubescens and Quercus cerris) by coppicing native woodlands. Conversely, it is well known that regressive processes affect these ecosystems when the disturbance is more intense and/or more frequent than the cork oak’s regeneration and recovery ability. Due to the combined effects of overgrazing and wildfires, cork oak stands are initially replaced by species-poor and structurally simplified mantle and shrubland communities, dominated by fast resprouters such as Cytisus infestus, Spartium junceum and Euphorbia characias, but also by species that are largely dispersed by wild animals, such as Pistacia lentiscus, Pistacia terebinthus, Osyris alba, Prunus spinosa, Pyrus spinosa, Rubus ulmifolius and Crataegus monogyna ([37]). If the disturbance regime is further intensified, perennial grasslands dominated by Ampelodesmos mauritanicus are the most likely fate of Q. suber stands, as has largely occurred in other areas of the western Mediterranean basin ([65]). Contrarily, open and scattered cork oak stands could evolve towards more complex woodlands if grazing pressure and fire frequency decrease ([14]). However, a similar scenario, consistent with the hypothesis of many authors who attribute the meaning of steady disclimax to cork oak stands, maintained by moderate disturbance regimes, especially wildfires ([7]), should be supported by further qualitative and quantitative investigations.
As far as our study sites are concerned, they have been particularly prone to intense and frequent human pressure (mostly wildfires and overgrazing) for millennia. Natural woodlands, including cork oak stands, have been exploited or cleared since proto-historical times. The land patches between these forests have been historically devoted to other agro-pastoral land uses (leaving no tree species), because agroforestry systems like dehesa do not belong to local Sicilian farming tradition. Accordingly, the surveyed cork oak stands can be interpreted as the last remnant nuclei of once more widespread forest patches. Such worrying conditions of the territory under investigation are further exacerbated by the lack of effective protection regime as none of the surveyed forest stands falls within protected sites, and by the still high impact of wildfires. Hence, we deem that these cork oak stands would deserve full protection as their survival in a medium to long-term perspective is far from guaranteed.
Conclusions
Compared to other evergreen oaks, cork oak has a more restricted ecological niche, particularly concerning soil humidity. Moreover, a decline of Q. suber is foreseen, especially at the edge of its natural range, due to overgrazing, high fire frequency, soil pathogens and land-use changes. We studied several cork oak populations on soils deriving from calcareous bedrock, which are generally less suited to the species. Based on our results, we hypothesized that the local presence of Q. suber could depend on high fire frequency, the peculiar biogeochemistry of local soils (e.g., high Ca content), and erosion, which caused the re-emergence of calcareous substrates. The evolution of these stands is hardly predictable as strictly linked to disturbance frequency and intensity (mostly wildfires). Monitoring activities and protection measures are highly necessary to contrast the likely future shrinkage of these populations, which are threatened by extremely high fire frequency and overgrazing, as well as the likely negative effects of climate change on regeneration ability and survival rate.
Acknowledgements
The individual contributions of authors are the following: RS, TLM and SP conceived the study and established methodology; EB, RS, TLM, RSB, GS, GMP, AG and SP collected and analyzed data; GMP, AG and SP carried out field surveys; RSB performed the statistical analysis; EB, RS, TLM, RSB, GMP and SP wrote the manuscript.
This work was financially supported by the PRIN2010-11 “CARBOTREES” (Climate change mitigation strategies in tree crops and forestry in Italy - national coordinator: R. Valentini), funded by the Italian Ministry of Education, University and Research, and by Tommaso La Mantia’s funding Pj autf 008567.
We thank the reviewers for significantly improving the quality of the manuscript and for providing us with useful suggestions. Thanks are due to Broni Hornsby for the revision of the English.
References
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Online | Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Online | Gscholar
Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Riccardo Scalenghe 0000-0001-7614-4842
Tommaso La Mantia 0000-0002-7494-742X
Rafael da Silveira Bueno 0000-0001-8964-8572
Giovanna Sala 0000-0002-4813-7052
Giuseppe Maria Pizzurro
Andrea Giaimo 0000-0001-6158-2778
Department of Agricultural, Food and Forest Sciences, University of Palermo. v.le delle Scienze Ed. 4, I-90128 Palermo (Italy)
Institute of BioSciences and BioResources (IBBR), Italian National Council of Research (CNR), Unit of Palermo, c.so Calatafimi 434, I-90129, Palermo (Italy)
Corresponding author
Paper Info
Citation
Badalamenti E, Scalenghe R, La Mantia T, Bueno RS, Sala G, Pizzurro GM, Giaimo A, Pasta S (2020). The cork oak in the Mountains of Palermo (Italy): ecological insights from the south-eastern edge of its distribution range. iForest 13: 336-344. - doi: 10.3832/ifor3360-013
Academic Editor
Francisco Lloret Maya
Paper history
Received: Jan 27, 2020
Accepted: Jun 06, 2020
First online: Aug 07, 2020
Publication Date: Aug 31, 2020
Publication Time: 2.07 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2020
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 44675
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 37992
Abstract Page Views: 3134
PDF Downloads: 2727
Citation/Reference Downloads: 3
XML Downloads: 819
Web Metrics
Days since publication: 2152
Overall contacts: 44675
Avg. contacts per week: 145.32
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2020): 3
Average cites per year: 0.50
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Towards a functional phytosociology: the functional ecology of woody diagnostic species and their vegetation classes in Northern Italy
vol. 14, pp. 522-530 (online: 22 November 2021)
Research Articles
Is cork oak (Quercus suber L.) woodland loss driven by eucalyptus plantation? A case-study in southwestern Portugal
vol. 7, pp. 193-203 (online: 17 February 2014)
Research Articles
Assessing the habitat conservation status by soil parameters and plant ecoindicators
vol. 7, pp. 170-177 (online: 14 February 2014)
Research Articles
Carbon storage in degraded cork oak (Quercus suber) forests on flat lowlands in Morocco
vol. 9, pp. 125-137 (online: 08 August 2015)
Research Articles
Classification and mapping of Spanish Mediterranean mixed forests
vol. 12, pp. 480-487 (online: 14 October 2019)
Research Articles
Evergreen species response to Mediterranean climate stress factors
vol. 9, pp. 946-953 (online: 07 July 2016)
Research Articles
SimHyb: a simulation software for the study of the evolution of hybridizing populations. Application to Quercus ilex and Q. suber suggests hybridization could be underestimated
vol. 11, pp. 99-103 (online: 31 January 2018)
Technical Reports
Diversity pattern of vegetation in and around proposed Kotlibhel hydroelectric project along the Alaknanda River in Garhwal Himalaya (India)
vol. 4, pp. 38-43 (online: 27 January 2011)
Research Articles
Using field and nursery treatments to establish Quercus suber seedlings in Mediterranean degraded shrubland
vol. 13, pp. 114-123 (online: 26 March 2020)
Commentaries & Perspectives
The role of plant sociology in the study and management of European forest ecosystems
vol. 6, pp. 55-58 (online: 21 January 2013)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







