Tissue carbon concentration of 175 Mexican forest species
iForest - Biogeosciences and Forestry, Volume 10, Issue 4, Pages 754-758 (2017)
doi: https://doi.org/10.3832/ifor2421-010
Published: Aug 05, 2017 - Copyright © 2017 SISEF
Research Articles
Abstract
Reliable calculations of carbon stocks in forest ecosystems are crucial for proper implementation of global warming mitigation policies. Accurate estimations depend upon applying the correct factor of carbon (C) concentration for different forest species and tissues instead of the often assumed 50% carbon content. Despite the high forest species richness in Mexico and the increasing CO2 emissions, data on carbon concentrations in forest plant tissues are scarce. In this study, we determined variation in C concentration of different tissues for 175 plant species common in Mexican forests. C contents were estimated and contrasted for plant distribution, taxa, and plant structure (main stems, branches, twigs, bark, leaves, buds, fruits, roots and root cuticles). The mean C concentration across species was 44.7%. Species significantly differed in C concentration by tissue, environment and taxa. These multi-species data contribute to improve precision on estimates of C balance in terrestrial ecosystems, reducing the uncertainty in C inventories in Mexico and elsewhere.
Keywords
Carbon Sink, Plant Tissue C, Multi-species C, Global Warming
Introduction
Estimates of carbon stocks of terrestrial ecosystems are complex ([27]), due to a large species and ecosystem diversity caused by the convergence of Nearctic and Tropical regions ([31]), climate heterogeneity ([32]), and complex geological and climatic history ([13]).
Knowledge on carbon (C) concentration of different plant species and plant tissues is essential to accurately estimate C stocks in forest ecosystems ([38]) and thus understanding of C dynamics. Studies in different parts of the world and in Mexico often use a constant value for C concentration (usually 50%) to model C stocks in different ecosystems and taxa ([28], [40], [30]). Other authors, however, argue that such generalization can under- or over-estimate real C stocks ([43], [19]), causing uncertainty in the potential of forests as C sinks ([22]).
Variations in C concentration between species is influenced by phylogeny and the environment (biotic and abiotic) in which plant species grow ([37]). Different plant species may have a specific chemical composition and carbon compounds due to their metabolism, as physiology and morphology are linked to an optimal functioning under the ecological conditions where they have evolved ([35]). Within a given individual, C concentration varies between tissues ([42]), depending to a larger extent on the chemistry of such tissues ([34]) than on plant age or size ([4]). This study presents C concentration values for 175 plant species in 18 families from temperate, tropical, subtropical, arid and semiarid zones in Mexico to establish whether and how biomass C concentration differed across environments, taxa and plant tissues. Such dataset will help determine if the use of the generalized assumption of 50% C content is applicable for Mexican forests and similar ecosystems and species in other parts of the world.
Material and methods
Experimental design and C concentration determinations
A selective sampling of 175 representative forest species ([29]) was carried out across Mexican environments. Sites were selected trying to encompass the large environmental variation but restricted by logging and accessibility. Temperate, tropical/subtropical and arid/ semiarid ecosystems were targeted as per González-Medrano ([16]). In 68 localities in 17 states (Fig. S1 in Supplementary materials), at least three individuals from each of the 175 species, a sample of ca. 50 g (fresh mass) was taken from different plant parts following Henry et al. ([18]): leaves (L), buds (Bd), fruits (F), branches (B), bark from branches (Bb), twig (< 5 cm in diameter - T), bark from twig (Bt), stem (S), bark from stem (Bs), roots (R) and root cuticles (Rc). Only undamaged trees with no deformities were included in the study.
Samples were taken to the lab following the procedures described in Karlik & Chojnacky ([20]), and immediately (<24 hours) were placed and dried in the lab at room temperature to avoid loss of volatile substances ([2], [23]). Following the procedure by Lamlom & Savidge ([22]), we broke down the samples into small particles to better estimate C content using a pulverizing mill, (Fritsch Pulverisette 2®, Idar-Oberstein, Germany), which yields fractions smaller than 10 μg. Total carbon concentration (TCC) was obtained using a Solids TOC Analyzer (model 1020A®) with catalytic combustion method from O·l·Analytical (College Station, TX, USA) in the Soils and Fertility lab in Postgraduate College.
Statistical analyses
As classification resulted in different sample sizes, data were subjected to an unbalanced ANOVA, using the general linear model (GLM) procedure of SAS version 9.1 ([36]). Differences between taxa (division, family and genus) and ecosystems were tested by one-way unbalanced ANOVAs, in which only groups with at least three samples were considered. Differences in concentration of C between tree tissues were evaluated, analyzing separately the two divisions: Magnoliophyta and Pinophyta. In all analyses the carbon concentration values were transformed with the arcsine function to satisfy the assumptions of normality and homogeneity of variances. A multiple comparison between means was carried out with the post-hoc Tukey’s test at P = 0.05 level.
Results
The mean C concentration across species and plant parts was 44.7% (Tab. S1 in Supplementary material), varying from 44.1% to 45.8% for temperate zone species, 42.8% to 44.3% for species from tropics and sub-tropics and from 43.8% to 44.2% for species in arid and semiarid zones (Tab. 1). Tree species from different biomes significantly differed in C concentration (F = 9.07, p = 0.0002). Species from the temperate zone had the highest C concentration (45.8%) and those from tropical and subtropical zones the lowest (42.8%).
Tab. 1 - Carbon concentration for Mexican forest species studied by region and moisture regime. (n): number of species. Different letters indicate significantly different means (p<0.05) after Tukey’s test.
| Region | Moisture regime |
n | C Concentration (%, mean ± sd) |
|---|---|---|---|
| Temperate Zone | Semidry | 9 | 44.14 ± 0.54 ab |
| Semihumid | 52 | 45.81 ± 0.36 a | |
| Tropical and Subtropical zones | Humid | 72 | 44.35 ± 0.23 ab |
| Semidry | 6 | 42.79 ± 0.37 b | |
| Subhumid | 4 | 44.16 ± 0.30 ab | |
| Arid and semiarid zones | Semidry | 22 | 43.95 ± 0.34 ab |
| Dry | 10 | 44.23 ± 0.50 ab |
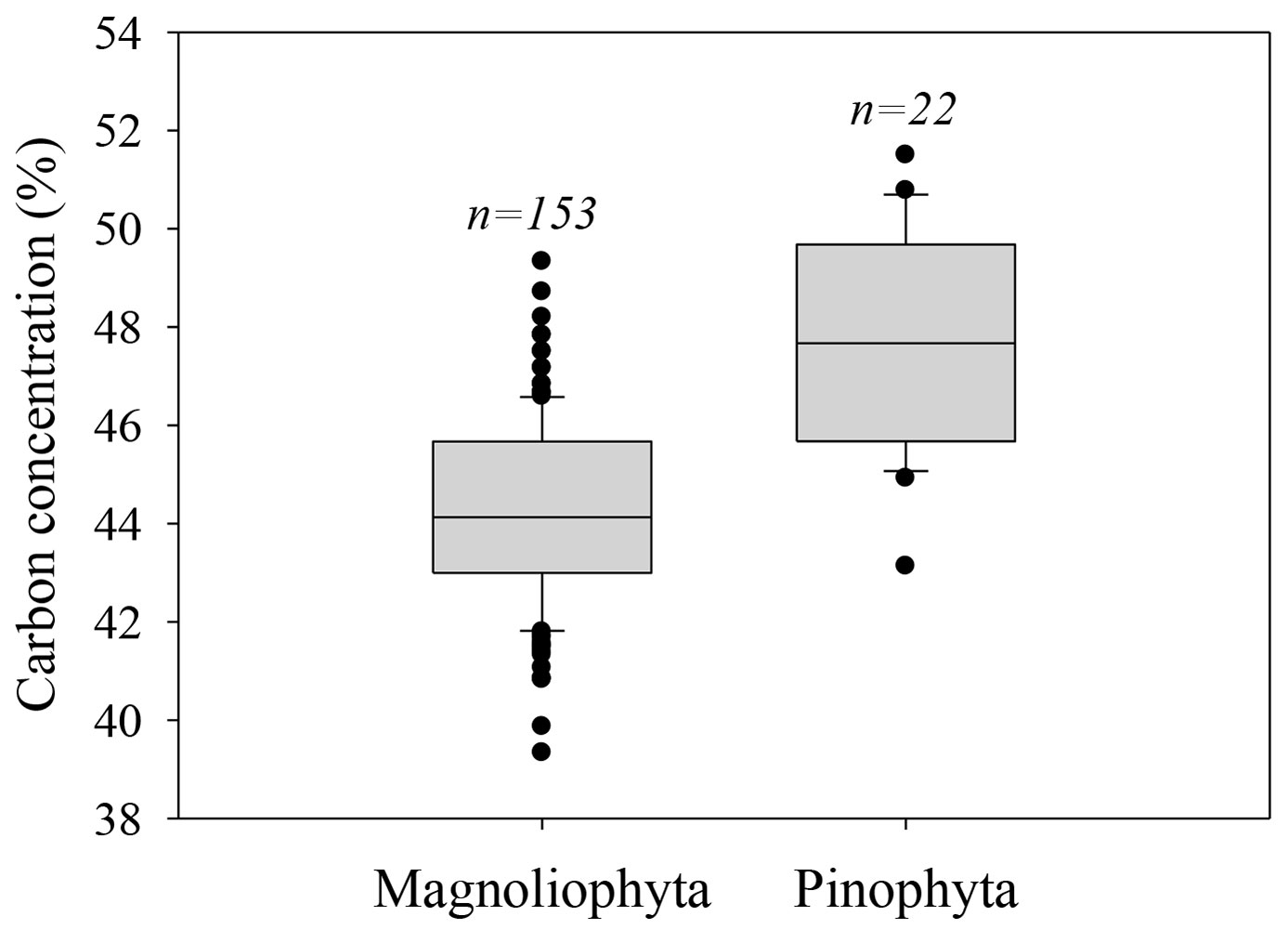
Different taxa also significantly differed in C concentration (F = 58.41, p < 0.0001). The highest C concentration was found for Pinophyta (47.5%), while Magnoliophyta had 3.3% less C concentration (44.2% - Fig. 1). Asparagaceae and Arecaceae had the lowest C concentration (43.1%).
Fig. 1 - Differences in Carbon concentration between Magnoliophyta and Pinophyta. A higher C concentration for species of Pinophyta was observed.
Carbon concentration varied between families (F = 6.72, p < 0.0001), from 42.8% for Asparagaceae to 48.3% for Pinaceae (Tab. 2). Within family variation in C concentration was relatively low with a mean standard deviation of 1.45%. Carbon concentration also differed between genera (F = 17.43, p < 0.0001). Two main groups could be identified (Tab. 3): the first included the genus Pinus, with the highest concentration (48.96%), while Acacia, Quercus, Bursera, Cordia, Pithecellobium and Ficus were in the second group.
Tab. 2 - Mean and standard error of carbon concentration for 18 families. Only families with enough replicates were used in this analysis. Different letters indicate different means (p<0.05) after Tukey’s test.
| Order | Family | Number of species |
Carbon concentration (%, mean ± sd) |
|---|---|---|---|
| Pinophyta | Cupressaceae | 5 | 45.68 ± 0.83 bcd |
| Pinaceae | 16 | 48.29 ± 0.50 a | |
| Magnoliophyta | Asparagaceae | 3 | 42.79 ± 1.01 e |
| Anacardiaceae | 7 | 45.01 ± 0.69 bcde | |
| Arecaceae | 5 | 43.58 ± 0.74 de | |
| Bignoniaceae | 4 | 44.40 ± 0.74 cde | |
| Boranginaceae | 4 | 43.10 ± 0.56 de | |
| Burseraceae | 4 | 43.54 ± 0.72 de | |
| Combretaceae | 3 | 42.82 ± 0.52 e | |
| Ericaceae | 4 | 47.15 ± 0.77 | |
| Fabaceae | 39 | 44.53 ± 0.28 ab | |
| Fagaceae | 17 | 44.29 ± 0.41 cde | |
| Malvaceae | 4 | 43.00 ± 0.81 e | |
| Meliaceae | 3 | 43.41 ± 1.21 de | |
| Moraceae | 8 | 43.41 ± 0.55 de | |
| Rubiaceae | 5 | 44.81 ± 0.49 bcde | |
| Salicaceae | 3 | 45.68 ± 0.83 bcd | |
| Sapotaceae | 3 | 48.29 ± 0.50 a |
Tab. 3 - Mean and standard error of carbon concentration of some of the genera sampled. Different letters indicate different means (p<0.05). (*): Genera with at least tree sampled species were included to comply with assumptions of analysis of variance.
| Genus* | Number of species |
Carbon concentration (%, mean ± sd) |
|---|---|---|
| Acacia | 7 | 43.93 ± 0.42 b |
| Bursera | 3 | 42.89 ± 0.44 b |
| Cordia | 3 | 43.08 ± 0.79 b |
| Ficus | 6 | 43.08 ± 0.53 b |
| Pinus | 12 | 48.96 ± 0.51 a |
| Pithecellobium | 3 | 43.92 ± 0.96 b |
| Quercus | 17 | 44.29 ± 0.41 b |
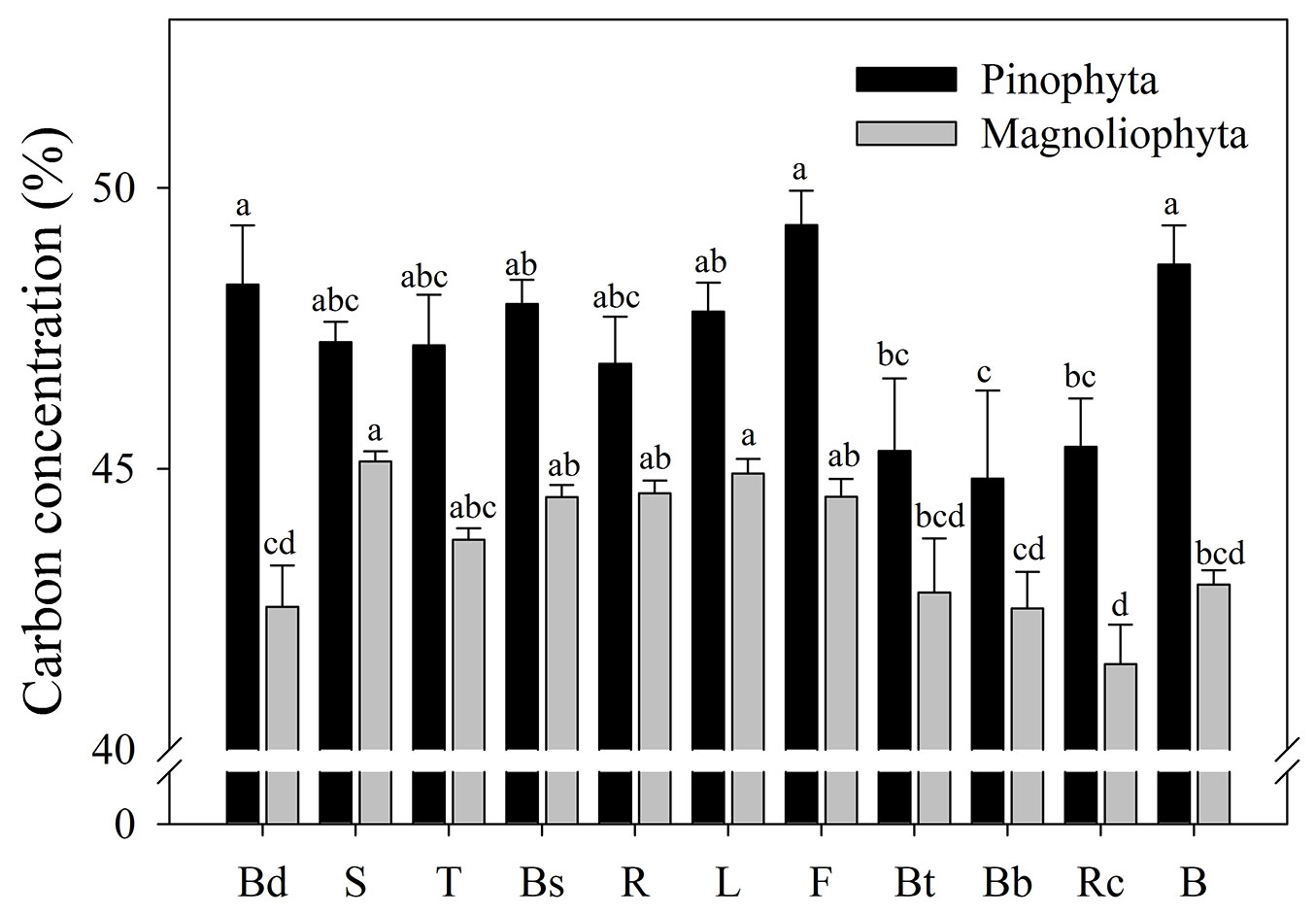
Carbon concentration between plant structure differed for Pinophyta (F = 2.36, p = 0.0128) and Magnoliophyta (F = 10.36, p < 0.0001). For conifers, fruits, buds and bark had carbon concentrations close to 50% (Fig. 2), which were significantly higher than those of dead tissue of roots and branches (44.8% to 45.3%). For broad-leaved species, C concentration was higher in stem and leaves (Fig. 2).
Fig. 2 - Carbon concentration for plant tissues grouped by taxonomic division. Different letters between bars of the same color indicate different means (p<0.05). Leaves (L), buds (Bd), fruits (F), branches (B), bark from branches (Bb), twig (<5 cm in diameter -T), bark from twig (Bt), stem (S), bark from stem (Bs), roots (R) and root cuticles (Rc).
Discussion
The common C-concentration constant used for the aerial parts of the trees is 50%. This is still broadly used in large scale models of C fluxes and sinks ([3], [24], [25], [15]). In our study, all values for C concentration of 175 Mexican forest plants were below 50% with an average value of 44.7%. In agreement with other multi-species studies ([46], [23], [7]), we suggest to use specific estimations for different genera and ecosystems instead of the 50% assumption. We also recommend to test the overestimation stemming from the adoption of such assumption in other ecosystems and species.
C concentration was higher in temperate than in tropical tree species, which could be a result of C rich volatile substances ([37], [15]), and lignin ([22]) in species of temperate environments. In contrast, tropical trees might have less C due to differences in cellulose and lignin contents ([12], [23]).
C concentration was higher for Pinophyta than for Magnoliophyta, likely due to high contents of resins and other C rich organic compounds in the former ([33]). Variations in C concentration between biomes were small, suggesting that C concentration is relatively constant for species across environments, as already reported in other studies ([23], [41]). Moreover, a large within-family variations in C content was observed. This is in agreement with findings that C concentration is not a strongly phylogenetically-conserved trait ([23]). In our study the larger within-family variation was found in Pinaceae, coinciding with the findings of Yerena-Yamallel et al. ([44]). This is possibly due to differences in chemical composition of cellulose, lignin, hemicellulose, and starches ([24]). The high C content found in leaves compared with other tissues might be the result of higher rates of volatile compounds, as suggested by Yerena-Yamallel et al. ([45]). Other studies have also found variations of C concentration between plant tissues, with higher values in leaves ([46], [10], [11]). Because leaves differ in their longevity, their role as C stocks must vary between evergreens and deciduous species ([23], [24]). The high values of C (ca. 50%) found in bark and cones of Pinophyta in this study, could be the result of C-rich lignin and suberin in these structures ([14]).
Other studies have observed variations in C concentration in dead or decaying tissue. For instance Harmon et al. ([17]) found that uncertainties associated to forest C inventories may be reduced using specific detritus C coefficient for each taxon, instead of using the generalized 50% assumption. In dead conifer trees, as decomposition takes place, a slight increase in C concentration occurs ([8], [21]). Other authors argue about the relative importance of intra- vs. interspecific variation in wood C for forest C assessments. For instance, Martin et al. ([25]) suggest that variations in C concentration between tissues of the same species are less important than variations between species for modelling C dynamics in forest ecosystems. Improvement and validation of models for given ecosystems could be a viable solution, using wood density as an independent variable as suggested by some authors ([39], [9]).
Reliable C concentration estimations are crucial for determining the role of forests in the global C cycle ([5]). The results from this study contribute to more accurate estimations of carbon concentration of plant tissues across a large number of tree species in Mexico. Even though differences between the 50% common assumption and actual findings are small, these differences would mean gross C overestimations at the scale of stands, forests or biomes ([15]). These could lead to C content overestimations in studied Mexican forests in the order of 1.5 Mg ha-1 ([45]) or 3.9 Mg ha-1 ([1]). Taxa-specific results contribute to a more realistic assumptions of C sinks in different ecosystems ([6], [26]) with implications for other countries where the studied species occur and challenges the current assumption of a 50% C concentration of plant tissues across forests worldwide.
Conclusions
The C concentration of plant tissues for Mexican tree species differed between environments, taxa and plant tissues. Our results show that the generalized assumption of 50% C content is consistently slightly above real values for the species studied. This overestimation, though small in some cases, could lead to errors of 3.9 Mg ha-1 in some Mexican forests. To improve estimations of C reserves, our values should be used for the studied genera. Given the large number of species studied, it would be important to test whether the generalized 50% assumption does apply in other environments. It is not practical to use C concentration for each plant tissue, therefore more research that models C concentration indices is still necessary, especially to distinguish dead or decaying tissues. Our study contributes to a better understanding of C concentration in forest ecosystems and provides a large dataset for Mexican species that can be contrasted across environments elsewhere.
Acknowledgments
Funding was provided by CONACYT (Consejo Nacional de Ciencia y Tecnología - grant CB-2013/222522). MPG, JASR carried out field and lab work; JASR, MPG, JF and EJ performed statistical analysis; MPG and EJ conceived the study and helped draft the first version manuscript. All authors wrote the manuscript. Many thanks to the landholders and foresters who allowed data collection. Direccion General de Vida Silvestre, SEMARNAT (Secretaría de Medio Ambiente y Recursos Naturales), Mexico, gave technical permit. Adrian Herrera and his staff assisted with field work. We are grateful to the anonymous reviewers for their useful comments and suggestions.
References
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Gscholar
CrossRef | Gscholar
Authors’ Info
Authors’ Affiliation
Universidad Juárez del Estado de Durango, Facultad de Ciencias Forestales, Río Papaloapan y Blvd. Durango s/n, Colonia Valle del Sur, C.P. 34120 Durango, Dgo. (México)
Campo Experimental Valle del Guadiana, Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias, Carretera Durango-Mezquital km 4.5, C.P. 34170, Durango, Dgo. (México)
Universidad Autónoma de Nuevo León, Facultad de Ciencias Forestales, A.P. 41, Carretera Nacional No. 85, Km 145, C.P. 67700, Linares, N.L. (México)
Instituto Potosino de Investigación Científica y Tecnológica, División de Ciencias Ambientales, Camino a la Presa San José No. 2055, Colonia Lomas 4a. Sección, San Luis Potosí, S.L.P., C.P. 78216 (México)
Corresponding author
Paper Info
Citation
Pompa-García M, Sigala-Rodríguez JA, Jurado E, Flores J (2017). Tissue carbon concentration of 175 Mexican forest species. iForest 10: 754-758. - doi: 10.3832/ifor2421-010
Academic Editor
Ana Rey
Paper history
Received: Mar 08, 2017
Accepted: Jun 15, 2017
First online: Aug 05, 2017
Publication Date: Aug 31, 2017
Publication Time: 1.70 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2017
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 50123
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 40871
Abstract Page Views: 3954
PDF Downloads: 4189
Citation/Reference Downloads: 18
XML Downloads: 1091
Web Metrics
Days since publication: 3246
Overall contacts: 50123
Avg. contacts per week: 108.09
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2017): 20
Average cites per year: 2.22
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Review Papers
Wood anatomy of boreal species in a warming world: a review
vol. 13, pp. 130-138 (online: 09 April 2020)
Review Papers
Comparative assessment for biogenic carbon accounting methods in carbon footprint of products: a review study for construction materials based on forest products
vol. 10, pp. 815-823 (online: 25 September 2017)
Research Articles
Soil C:N stoichiometry controls carbon sink partitioning between above-ground tree biomass and soil organic matter in high fertility forests
vol. 8, pp. 195-206 (online: 26 August 2014)
Research Articles
Assessing the carbon sink of afforestation with the Carbon Budget Model at the country level: an example for Italy
vol. 8, pp. 410-421 (online: 02 October 2014)
Research Articles
Do the rubber plantations in tropical China act as large carbon sinks?
vol. 7, pp. 42-47 (online: 21 October 2013)
Research Articles
Climate impacts on tree growth in a Neotropical high mountain forest of the Peruvian Andes
vol. 13, pp. 194-201 (online: 19 May 2020)
Research Articles
An approach to estimate carbon stocks change in forest carbon pools under the UNFCCC: the Italian case
vol. 1, pp. 86-95 (online: 20 May 2008)
Research Articles
Effects of warmer and drier climate conditions on plant composition and biomass production in a Mediterranean shrubland community
vol. 1, pp. 39-48 (online: 28 February 2008)
Research Articles
Voluntary carbon credits from improved forest management: policy guidelines and case study
vol. 11, pp. 1-10 (online: 09 January 2018)
Review Papers
Carbon neutrality of forest biomass for bioenergy: a scoping review
vol. 16, pp. 70-77 (online: 05 March 2023)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







