
Long-term changes in surface-active beetle communities in a post-fire successional gradient in Pinus brutia forests
iForest - Biogeosciences and Forestry, Volume 10, Issue 2, Pages 376-382 (2017)
doi: https://doi.org/10.3832/ifor2140-009
Published: Mar 16, 2017 - Copyright © 2017 SISEF
Research Articles
Abstract
Fire is one of the most important ecological factors for many ecosystem types. Since prehistoric times, synergistic effects of fires and humans have led to changes in Eastern Mediterranean ecosystems. The effects of fire on different trophic levels, particularly regarding plants, have been examined intensively in fire-induced ecosystems. In this study, we aimed to study long-term changes in beetle community structure after fire in Pinus brutia Ten. forests. Five sites burned in different years and a control site unburned for at least 50 years were selected. Beetle sampling was conducted using four pit-fall traps in each of four transects in three replication plots at every successional site and in two plots at the control site. Microhabitat variables related to vegetation structure and litter layer were recorded and associated with abundances of beetles and feeding groups. The results showed that total, wood-eating, and predator beetles showed a decreasing trend of abundance along the successional gradient after fire. In contrast to these groups, herbivores tended to increase towards the late successional stages. Middle and late successional stages were important in terms of species richness, species diversity and evenness of beetle communities and feeding groups. The characteristics of vegetation and litter layer changed with successional gradient, playing a decisive role in the structure of beetle communities at successional sites. According to the data presented here, a mosaic structure consisting of different successional stages is very important to sustain high species diversity in beetle communities.
Keywords
Forest Fires, Coleoptera, Brutia Pine, Resilience, Autosuccession
Introduction
Pinus brutia is the dominant tree of old growth forests in the Aegean, Marmara, and Mediterranean regions of Turkey. Such forests range from the sea level up to 700-800 m in the Aegean region and 1500 m in the Mediterranean region ([15]). Fire is an integral part of many ecosystems such as the Mediterranean and it serves as a strong selective force on plant and animal communities ([45]). As one of the most important ecological factors, fire has been studied comprehensively in these ecosystem types. Much attention has been paid to vegetational changes and regeneration strategies of plant communities in Mediterranean forests. Because dynamic feedbacks between plants and their consumers influenced the response of the whole ecosystem, incorporating higher trophic level responses into the pool of knowledge regarding fire effects is essential ([16]).
Effects of fire on insects and other arthropods can operate through a variety of mechanisms at different temporal scales ([2]). Direct effects of fires include mortality, forced emigration ([50]), or immigration of pyrophilous insects ([53], [54]). The sensitivity of fire effects depends on biological characteristics such as trophic level, seasonal activity, and vertical distribution ([30]). Indirect or long-term effects of fire depend substantially on habitat changes. Following fire, changes in plant species’ composition, plant diversity ([20], [39], [38]), and plant architecture ([40], [20]) influence the diversity and richness of insect communities. In Mediterranean ecosystems, a general model concerning response to fire at population or community scale is not possible, but on a regional scale, two resiliency components, elasticity and inertia, along with the mosaic pattern of the fires and the possibilities of local emigration and emigration result in relative stability of the animal communities ([30]). At the functional scale, greater stability is achieved by replacement of functionally similar species that dominate under different environmental conditions in the heterogeneous parts of the region ([27]).
Beetles form the largest order in the animal kingdom, with over a third of a million described species ([5]). They are commonly used in ecological monitoring studies, as they display assemblage-level responses to habitat change and can be a more direct measure of the recommencement of some ecosystem functions ([21]). The aim of the present study was to investigate the long-term response of beetle communities to habitat change depending on successional gradient after fire. The existence of beetles in all consumer trophic levels allows evaluation of the responses of these groups. In the present study, beetle community composition was also assessed by feeding groups to determine whether the their abundance differed among post-fire successional sites.
Material and methods
Study area
The study was carried out in several Pinus brutia forests located in the Marmaris region (36° 50′ N, 28° 17′ E) and its surrounding areas, which is situated on the Mediterranean coast of southwestern Turkey. This area has a typical Mediterranean climate with hot and dry summers. The total precipitation is 1211.7 mm year-1 (between 1975 and 2006), with a dry period lasting 5 months from May to September. Monthly mean temperatures range from 10.6 °C in January to 28.3 °C in July. The dominant vegetation of the area is P. brutia forest and these forests are characterized by fire-created habitat mosaic consisting of many sites at different stages of succession.
The study sites were selected from P. brutia forests burned at different times in the area. The successional gradient consisted of five areas burned 3, 6, 9, 16, and 26 years ago and an area unburned for at least 50 years. The samplings were performed at three replication sites for every successional site and at two for the unburned forests. All sampling sites were selected in the central part of the burned areas, where crown fires formerly caused the complete destruction of vegetation and aboveground layer. The sites were located apart from unburned forest at least 1000 m.
Insect sampling
Beetles were sampled with four pit-fall traps that were placed along a transect at 10-m intervals. Four transects were used at every replication site. The traps consisted of plastic jars of about 7 cm in diameter buried in the soil up to the rim and half-filled with 30% ethylene alcohol. Sampling was carried out from March to October of 2006. At the end of each sampling period insects caught in the pitfall traps were removed and brought to the laboratory for preparation. All caught beetles were sorted into morphospecies according to their external morphology and identified to family level.
Habitat Measurement
Microhabitat structure of the sites was analyzed with the point quadrat technique ([41]). For every transect at each replication site, two trap locations were selected for measurement of microhabitat attributes. In each measurement location nine points were selected; one was at the center and the remaining eight were situated in four different directions around the trap. At each point, the plant species were identified, the height of plant at this point was measured, and the surface characteristics were recorded.
Habitat variables measured at each site were height and total cover of vegetation, plant species richness, cover of plant functional types (tree, shrub, subshrub, and grass) and cover of biotic and abiotic surface components.
Statistical analysis
To test the statistical significance of any changes in abundances of beetles and feeding groups among successional stages, the analysis of variance (ANOVA) was applied. Total counts during sampling were used. In the ANOVA, abundances were expressed as mean number of individuals per four traps in a transect. Square root and logarithmic transformations were used when the data did not meet the parametric assumptions. Overall, six successional stages were compared using means of abundances from transects at the replication sites. Linear regression was used to test change in beetle abundance along the successional gradient. Linear regression was also used to examine interactions between microhabitat variables and feeding groups. The following community parameters were estimated for each successional site: species richness (number of species), species diversity Shannon index (H′ = -∑ pi log pi, where pi is the relative abundance of the ith species) and evenness (E′= H′/ log S, where S is the number of species). Specialist species to a certain successional stage and generalist species were estimated. Species that made up over 90% of collected individuals in certain successional sites were evaluated as specialist species for that site.
The relationship between beetle abundance among sampling sites and microhabitat variables was estimated using ordination techniques. A canonical correspondence analysis (CCA) using the CANOCO version 4.5 software package ([44]) was used to assess habitat preferences of microhabitat variables in relation to successional stages. Twelve microhabitat variables were used in the analysis. The statistical significance of the canonical axes was tested using Monte Carlo permutations. Microhabitat variables were added one by one to the model, starting from the most important one, until the subsequent variable was statistically nonsignificant. The analysis was constrained by microhabitat variables measured for each site, which are indicated with arrows that represent the direction strength of their correlations.
Results
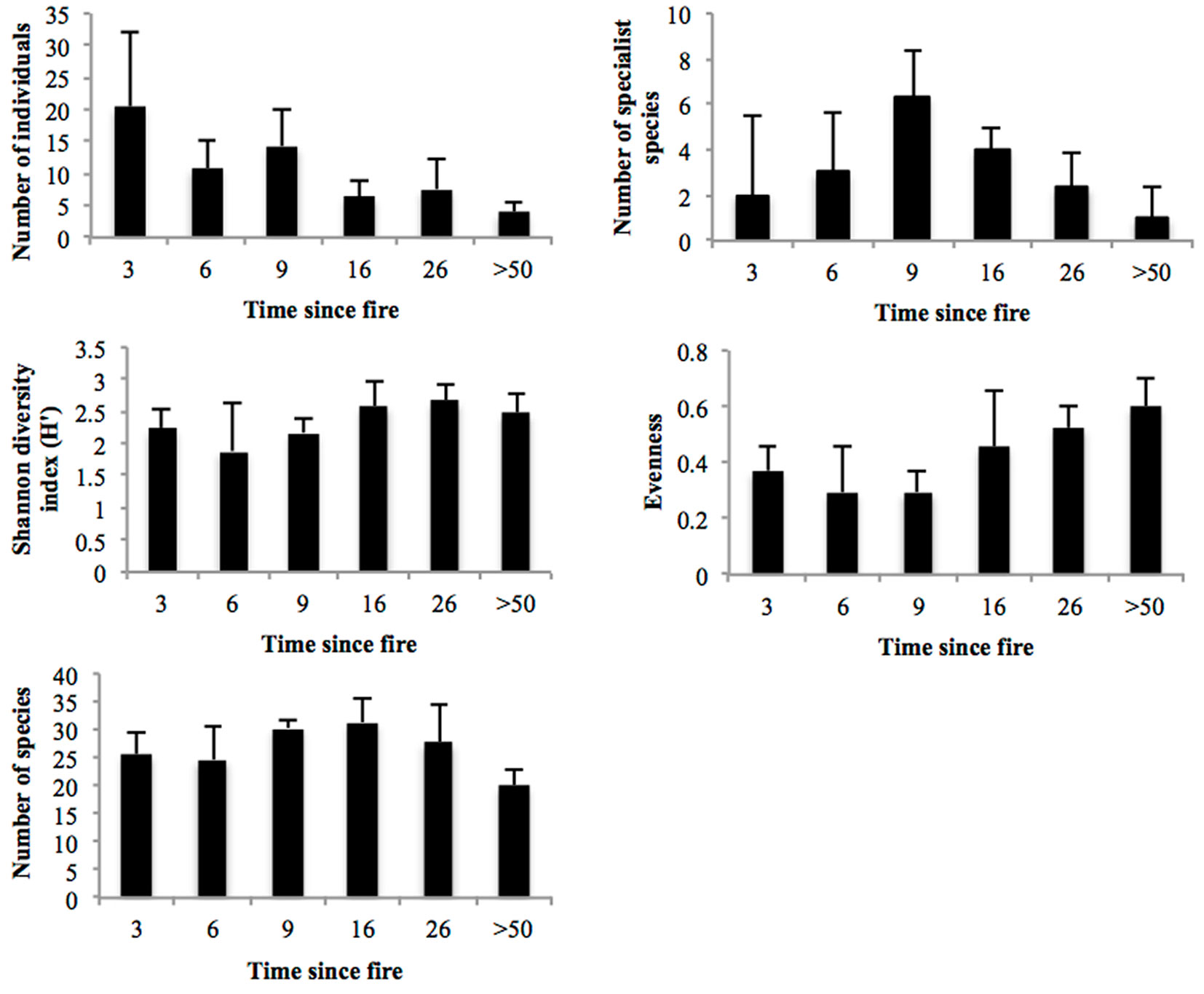
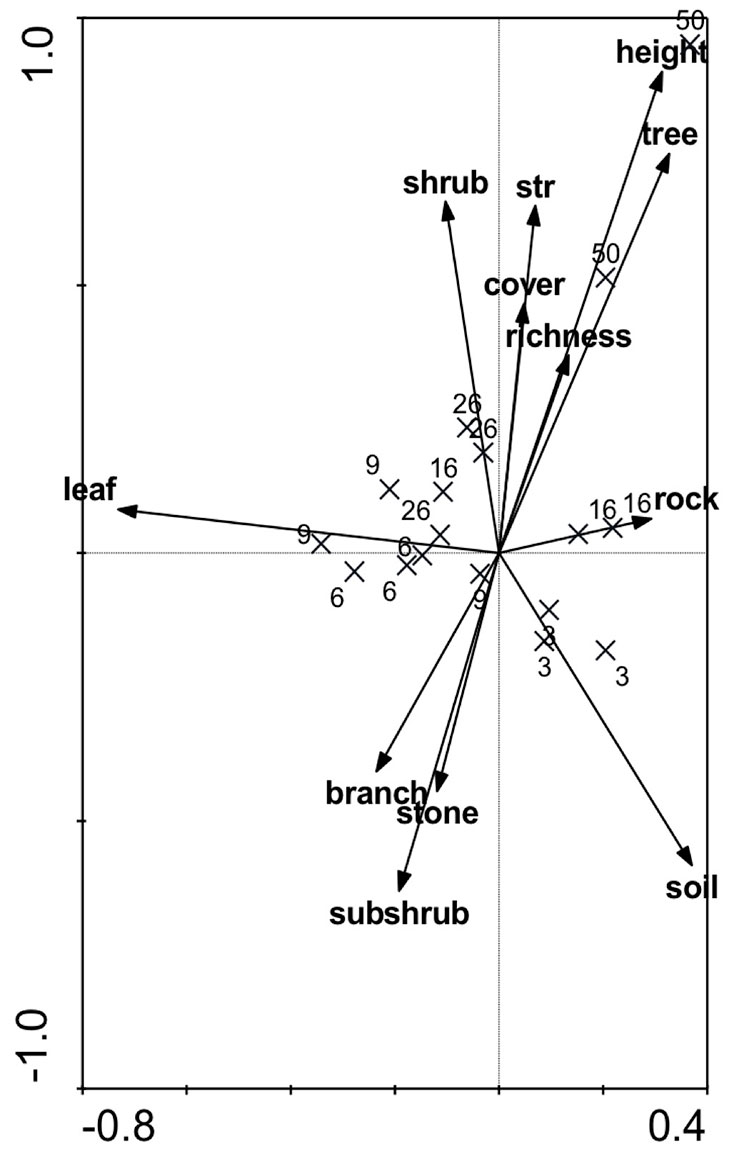
A total of 3425 beetles comprising 121 morphospecies from 26 families were collected (Tab. 1). The most abundant families were Buprestidae (41%) and Tenebrionidae (28%). The mean number of beetles per trap tended to decrease along the successional gradient (R2 = 0.298; df = 5; p < 0.0001), despite the rises at sites burned 9 and 26 years previously that deviated from the downward tendency (Fig. 1). The abundance value for the earliest stage was significantly greater than that of all other successional stages. The site burned 9 years previously had the second most abundant beetle community and it was significantly different than the site burned 16 years previously and the control site (F = 21.049; df = 5; p < 0.0001). However, species diversity and evenness values were higher in the late successional sites in comparison with the earlier stages (Tab. 2). The number of specialist species was highest at the site burned 9 years previously and declined towards the early and late successional stages (Fig. 1). Fifteen species of the total were generalist, well represented at all sites. CCA revealed interactions between microhabitat variables and successional sites in terms of beetle abundance (Fig. 2). The sum of eigenvalues was 2.071 and the eigenvalues of the first two axes were 0.338 and 0.254, respectively. These axes explained 28.6% of the cumulative variance of the species data and 36.7% of the species-environment relationship. The height of vegetation, tree cover, vegetation stratification, and plant species richness at the upper right side of the ordination and mature pine sites (sites unburned for at least 50 years) were highly correlated with these microhabitat variables. Most of the middle and late successional sites (from the site burned 6 years before to the site burned 26 years before) were associated with leaf and shrub cover. Vegetation height and leaf cover were the two significant variables associated with beetle abundance (F = 2.047, p = 0.006 and F = 1.875, p = 0.006, respectively). The earliest successional sites were just correlated with bare soil cover at the bottom right side.
Tab. 1 - Beetle families from pitfall catches, their feeding habits and mean abundance per replication site in successional sites.
| Families | Feeding habits |
Time since fire (years) | |||||
|---|---|---|---|---|---|---|---|
| 3 | 6 | 9 | 16 | 26 | >50 | ||
| Anobiidae | xylophagous | - | 0.3 | - | 0.3 | - | - |
| Apionidae | herbivore | 0.3 | 1.0 | - | - | - | - |
| Bolboceratidae | saprophagous | - | - | 0.7 | - | - | - |
| Bruchidae | granivore | 0.3 | - | 0.3 | 0.7 | - | - |
| Buprestidae | xylophagous | 220.0 | 26.0 | 78.3 | 91.0 | 55.7 | 18.0 |
| Carabidae | predator | 41.0 | 5.0 | 30.0 | 5.0 | 5.0 | 2.0 |
| Cerambycidae | xylophagous | - | - | 0.7 | 0.7 | 0.3 | - |
| Chrysomelidae | herbivore | 0.3 | 0.3 | 0.3 | - | - | - |
| Cleridae | predator | 80.3 | 3.3 | 4.0 | 16.7 | 11.0 | 5.5 |
| Cucujidae | saprophagous | - | 0.3 | - | 1.0 | 0.7 | 0.5 |
| Curculionidae | herbivore | - | - | 0.7 | 2.0 | 0.7 | - |
| Dermestidae | saprophagous | 0.3 | 0.7 | 0.3 | - | - | 0.5 |
| Elateridae | various | - | 1.0 | 0.7 | 0.3 | - | - |
| Glaphyridae | saprophagous | 0.7 | 0.7 | 8.0 | 1.7 | 2.3 | - |
| Geotrupidae | saprophagous | 0.3 | 0.7 | 0.3 | 1.0 | 0.7 | 1.0 |
| Lucanidae | various | - | - | - | - | 0.3 | - |
| Meloidae | herbivore | 4.7 | 1.3 | 4.0 | 7.0 | 1.0 | 0.5 |
| Mordellidae | herbivore | 4.3 | 2.7 | 3.7 | 9.0 | 13.7 | 0.5 |
| Nitidulidae | saprophagous | - | - | 1.0 | - | - | - |
| Oedemeridae | herbivore | 0.3 | - | 1.0 | 1.7 | 0.3 | 1.0 |
| Orphnidae | herbivore | - | - | - | 1.0 | 4.0 | - |
| Rutelidae | herbivore | 2.0 | 1.0 | 1.3 | - | 1.0 | - |
| Scarabaeidae | saprophagous | 5.0 | 4.0 | 3.0 | 2.0 | 1.0 | 3.0 |
| Scolytidae | xylophagous | 7.3 | 2.0 | 3.0 | 11.0 | 2.7 | 13.5 |
| Staphylinidae | predator | 8.7 | 2.7 | 4.0 | 4.7 | 11.3 | 10.0 |
| Tenebrionidae | various | 65.3 | 118.7 | 104.3 | 18.0 | 23.7 | 3.5 |
Fig. 1 - Number of individuals, species and specialist species, species diversity and evenness values for beetle communities at successional sites (mean ± standard deviation).
Tab. 2 - Species richness, species diversity and evenness values for feeding groups of beetle communities at successional sites (mean ± standard deviation).
| Parameter | Feeding Groups | Time since fire (years) | |||||
|---|---|---|---|---|---|---|---|
| 3 | 6 | 9 | 16 | 26 | >50 | ||
| Species richness | Herbivores | 4.500 ± 3.5 | 4.500 ± 1.5 | 6.000 ± 1.7 | 6.667 ± 2.1 | 4.333 ± 2.1 | 2.000 ± 1.4 |
| Xylophagous | 8.000 ± 1.0 | 8.333 ± 1.5 | 7.333 ± 1.5 | 9.000 ± 1.7 | 7.333 ± 1.2 | 5.000 ± 0.0 | |
| Predators | 8.333 ± 1.5 | 4.667 ± 2.1 | 8.333 ± 0.6 | 9.000 ± 2.6 | 8.667 ± 0.6 | 7.500 ± 0.7 | |
| Saprophagous | 2.667 ± 2.1 | 4.000 ± 2.6 | 4.667 ± 1.2 | 4.667 ± 1.5 | 5.000 ± 3.5 | 3.500 ± 0.7 | |
| Species diversity | Herbivores | 1.165 ± 0.4 | 0.995 ± 0.5 | 1.552 ± 0.4 | 1.514 ± 0.2 | 0.925 ± 0.2 | 0.401 ± 0.6 |
| Xylophagous | 1.204 ± 0.3 | 1.681 ± 0.2 | 1.355 ± 0.4 | 1.627 ± 0.5 | 1.575 ± 0.2 | 1.198 ± 0.2 | |
| Predators | 1.366 ± 0.1 | 1.312 ± 0.4 | 1.388 ± 0.5 | 1.690 ± 0.4 | 1.834 ± 0.3 | 1.644 ± 0.2 | |
| Saprophagous | 0.566 ± 0.8 | 1.190 ± 0.6 | 1.167 ± 0.1 | 1.307 ± 0.3 | 1.373 ± 0.5 | 1.221 ± 0.2 | |
| Evenness | Herbivores | 0.848 ± 0.2 | 0.792 ± 0.1 | 0.735 ± 0.2 | 0.706 ± 0.1 | 0.685 ± 0.3 | 0.872 ± 0.2 |
| Xylophagous | 0.427 ± 0.1 | 0.652 ± 0.0 | 0.549 ± 0.1 | 0.648 ± 0.3 | 0.679 ± 0.2 | 0.672 ± 0.2 | |
| Predators | 0.479 ± 0.1 | 0.851 ± 0.1 | 0.531 ± 0.3 | 0.645 ± 0.2 | 0.741 ± 0.2 | 0.693 ± 0.1 | |
| Saprophagous | 0.830 ± 0.2 | 0.946 ± 0.0 | 0.724 ± 0.2 | 0.834 ± 0.1 | 0.916 ± 0.1 | 0.979 ± 0.0 | |
Fig. 2 - Canonical correspondence analysis (CCA) biplot for the beetles communities at 17 sites constrained by twelve microhabitat variables.
Of the beetles that could be placed into feeding guilds, there were more xylophages (55.6% of individuals) than any other guild. Predators (25.9%), herbivores (10.3%), saprophages (8.1%), and granivores (0.2%) were also represented. Predator and xylophagous beetles were more abundant in the earliest successional stages in comparison with the other successional stages (df = 5, p < 0.0001, F = 12.503 and F= 13.617, respectively). Concerning interactions with microhabitat variables, abundances of both feeding groups was associated with only bare soil cover (xylophagous: R2 = 0.140; F = 10.736; p = 0.002; predators: R2 = 0.286; F = 26.398; p < 0.0001).
Another guild, herbivores had a contrary trend to xylophagous and predator beetles over the successional gradient. Their abundance tended to increase with successional gradient, whereas it was smallest at the control site. This site differs significantly from sites burned 16 and 26 years previously (F = 3.180, df = 5, p = 0.013). Herbivore abundance was positively correlated with rock cover (R2 = 0.160, F = 12.598, p = 0.001 - Fig. 3).
Fig. 3 - The changes of feeding groups of beetles depending on successional gradient and interactions between abundance of feeding groups and significant microhabitat variables.
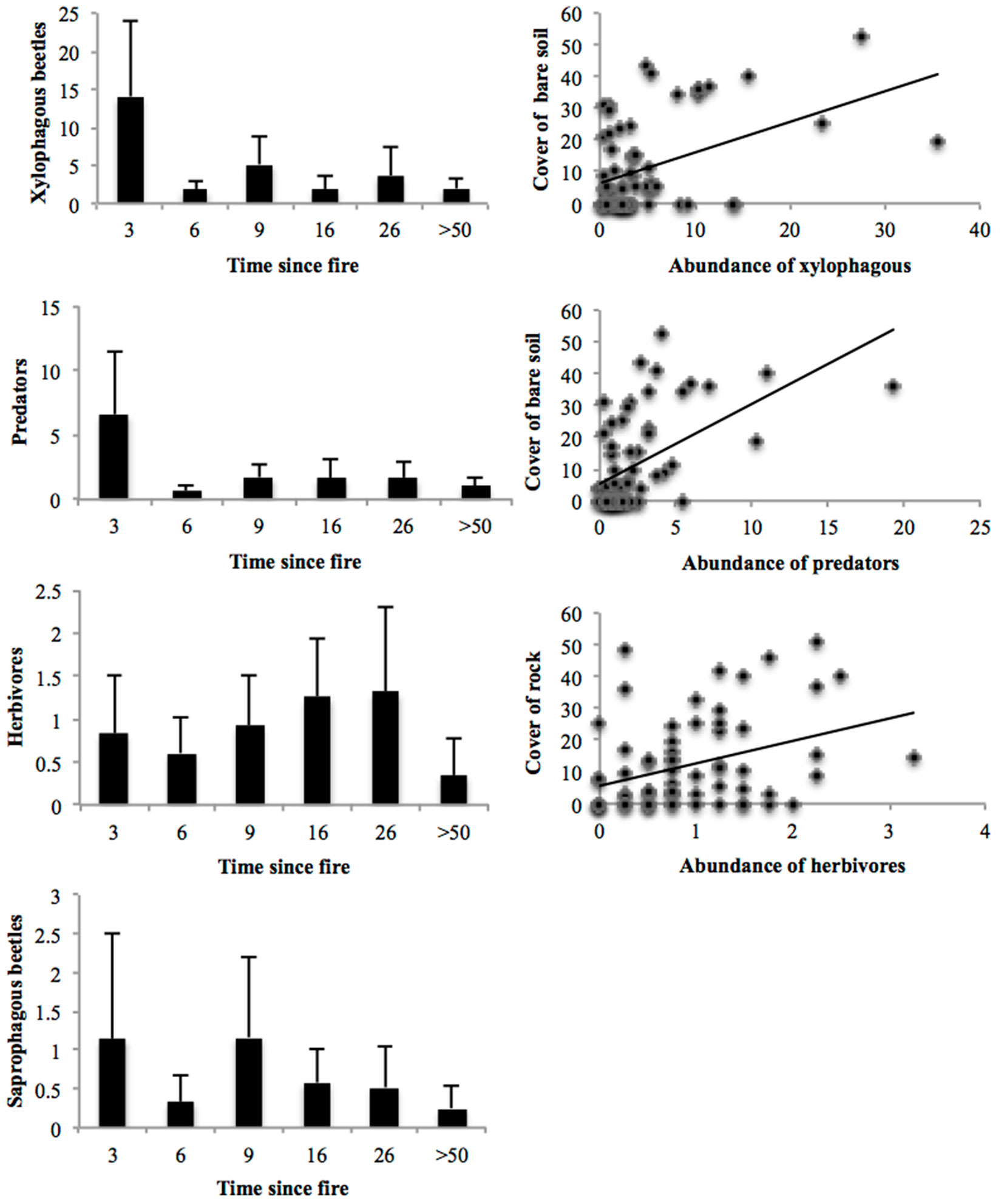
Abundance of saprophagous beetles was higher at sites burned 3 and 9 years previously and lower at site 16 years old and later successional stages. No significant differences were found among successional stages in terms of abundance (Fig. 3). No microhabitat variables were related to the abundance of saprophagous beetles.
Discussion
Fire destroyed stands in a very short time, causing drastic changes in the habitat structure of faunal communities. Forest ecosystems characterized by a large extension of pine trees and a layer of humus and dead leaves on the ground are transformed to areas with a simpler vegetation structure with no tree cover and a large extension of bare ground after fire ([23]). For resilient ecosystems such as low-altitude Mediterranean forests, plant communities have some adaptive traits that enable them to regenerate rapidly after fire ([17], [11]). Some studies from these ecosystems show that faunistic groups recover within a short time after fire due to their high resilience as well ([35], [42]).
According to several studies conducted in the Mediterranean basin, the seasonality of the Mediterranean climate ([36]) and post-fire age of the stands ([31]) are two important factors that affected arthropod fauna. The activity rates of certain insect groups after fire increase rapidly and time since fire is one of the main factors determining the dynamics and structure of the community ([28], [32]). The results obtained in this investigation are in agreement with these studies that beetle communities displayed change depending on the successional gradient after fire. The earliest successional stage supported a greater number of individuals. At this stage, beetle abundance was the highest and it decreased depending on the successional gradient. In studies performed in fire-induced ecosystems, similar results were obtained ([13], [26]). The higher abundancy in earlier stages could arise from different groups’ attraction to post-fire areas. Some groups, such as heliophilous and floricolous insects, profit from the reduced tree cover and the number of flowering plants ([26]). Muona & Rutanen ([28]) stated that many groups of beetles in different guilds benefited from the fire-caused changes in availability of microhabitats and foods. Campbell et al. ([7]) stated about pollinator beetle groups that floral visitors increased in abundance and species richness in burn treatments because of the reduced density of overstorey trees and the increased amount of herbaceous plant growth.
Regarding the interaction between beetle community parameters and microhabitat variables, our results showed that the characteristics of the vegetation and litter layer play an important role in the community structure of beetles along the succession. As an autosuccession process in pine forests of the Mediterranean basin, plant species composition does not differ among successional stages ([48], [43]). After fire, plant communities return to a stage similar to that existing before the fire ([47]). Thereby habitat change in the successional process is much more dependent on the characteristics of the vegetation and litter layer. The changes in beetle community structure and dynamics in successional gradient substantially depend on these variations among successional sites. The variables that increased with successional gradient and reached their maximum in the mature forest, such as height of vegetation, stratification, tree cover, and plant species richness, were important for beetle communities in the climax community. At the middle and late successional sites, beetle communities were correlated with shrub and leaf cover. The importance of leaf litter and shrub cover for beetle communities was revealed in the study performed by York ([55]).
Xylophagous and predator beetles, which were two guilds that benefited from the post-fire conditions, had the highest abundance in the earliest stage as well. Since wood-eating insects prefer damaged or recently dead trees in burned areas, this group is frequently associated with fire ([34], [24], [4]). In some studies significant outbreaks caused by xylophagous insects were recorded in burned areas ([33]). This higher activity could be due to immigration of helio- and anthophilus species from surrounding intact stands ([25]). Open surfaces with woody materials and higher temperature than the surrounding areas attracted many species to recently burned areas ([46], [51]). Another abundant group, predators, consist of the families Carabidae, Staphylinidae, and Cleridae. The higher activation level of predators in the earliest stage resulted substantially from the families Carabidae and Cleridae. Carabidae is a group that has been frequently focused on in studies about habitat changes after disturbances ([12], [22], [49]). Cook & Holt ([8]) stated that fire might not have dramatic effects on ground beetles on prairies. They were represented in greater numbers in recently burned areas through colonization by opportunistic species ([9]). Cleridae are a group known as predators on wood-boring beetle larvae ([5]). It is not surprising to find them in greater numbers at the earliest successional site in correlation with wood-eating beetles. Saint-Germain et al. ([32]) found similar results regarding these groups and stated that saproxylic species (Buprestidae, Cerambycidae, and Scolytidae) depend on largely dead wood, and subcortical predators (Cleridae) were represented in great number of individuals in the earliest successional stages. In this study we found that abundance of Coleoptera guilds was substantially associated with surface characteristics. In particular, bare soil cover was determined the most important microhabitat variable because it was associated with abundance of total beetles, xylophagous species, and predators. Apigian et al. ([3]) stated similarly that increased bare mineral soil resulting from loss of duff and litter layers was the most significant variable explaining the change in beetle communities.
Contrary to xylophagous and predator beetles, herbivore abundance exhibited a rising trend with successional gradient, surpassing the higher value in the earliest stage. Species richness and diversity were highest in the middle successional stage. Herbivore abundance was associated with just one microhabitat variable, rock cover. Leaf quality, habitat structure, and habitat heterogeneity are variables reported to affect the herbivore community structure ([16]), but the positive interaction with rock cover is surprising. Rock cover is not a microhabitat variable that can associate with the herbivore abundance directly. Both herbivore abundance and rock cover had higher values in site burned 16 and 26 years ago. Detected correlation may have resulted from analogous changes of these two independent variables with successional gradient. But then, if we look at different point of view, high cover of rock in late successional stages may have contributed to increase of herbivore abundance. The study area has a higher productivity in comparison with similar sites in the eastern Mediterranean region because of peculiar local climatic conditions ([43]). Bare rock areas can represent forest openings in dense closed vegetation of late successional stages. Herbivores may have positively responded to these forest openings.
Middle and late successional stages with high abundance of herbivores, at the same time, were very important for species richness, species diversity and evenness of all guilds and total beetles. The number of specialist beetle species was also higher in the middle successional stages as compared with the other stages. Kaynas ([14]) stated in her study performed at the same sites that plant species richness, height of vegetation, and stratification increase along the successional gradient. The middle and late successional stages are more complex habitats including diverse microhabitats compared to the earlier ones, as the architectural complexity increases with vegetation height ([20]). Habitat permanence, habitat complexity, resource availability, and diversity increase with successional gradient ([6]). Habitat complexity was a powerful predictor of the species richness and abundance of pitfall-trapped beetles ([19]). Gardner et al. ([10]) stated that arthropod diversity was substantially influenced by the habitat components related to plant architectural and vertical diversity. The middle and late successional stages had an advantage over the climax community because they were lacking a crown layer. Complex habitat structure as well as the absence of a closed crown layer increased species richness of these groups in these habitats. Lower species diversity for some insect groups in closed canopy mature forest, as compared to successional sites burned at different times, was reported in several studies ([18]).
The high species richness at the middle and late successional sites might also be related to the pattern of shrub patchiness in the landscape. Shelef & Groner ([37]) stated that shrubs create the main driving force behind the beetle movement pattern and spatial distribution. Shrub shading might be an important factor for their thermoregulation in the Mediterranean climate, characterized by hot and dry summer seasons. Middle and late successional stages dominated by shrubs and short trees might create optimal conditions for insect species, providing protection against adverse environmental conditions and predation, as well as greater levels of prey items ([19]).
Conclusions
No negative effects on the community structure of beetles caused by habitat changes after fire were determined. Beetle communities of P. brutia forests appear to be resilient to fire as in many other fire-induced ecosystems ([29], [1], [2]). P. brutia forests in southwest Anatolia comprise a mosaic of forest patches of different ages after fire. Earlier successional stages were gaps of different size among mature forests, thus ensuring the maintenance of open habitat species in the area. Middle and late successional stages had high habitat heterogeneity and are of particular importance to sustain the species richness of beetles. Beside successional stages, unburned, mature P. brutia forests act as source areas for colonization of species ([52]). The protection of this mosaic structure of the landscape is very important to sustain high species richness of P. brutia forests.
Acknowledgements
Many thanks to Çagatay Tavsanoglu, Anil Soyumert, Oksal Macar, Filiz Yeni, Yasin Ilemin, Okan Ürker, Hüseyin Yilmaz, Sinan Kaynas and many undergraduate students for their help during the fieldwork, Atilla Küçükala, the Director of Marmaris National Park for his logical support, and Forest Management Directorate of Marmaris, Directorate of Marmaris National Park, and Aksaz Military Marine Base Command for allowing to conduct this study in the areas under their responsibility. The study was financially supported by Hacettepe University Scientific Research Unit (Project no: 02.02.601.004).
References
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Mehmet Akif Ersoy University, Faculty of Science and Arts, Department of Biology, Istiklal Campus, Burdur (Turkey)
Corresponding author
Paper Info
Citation
Kaynas BY (2017). Long-term changes in surface-active beetle communities in a post-fire successional gradient in Pinus brutia forests. iForest 10: 376-382. - doi: 10.3832/ifor2140-009
Academic Editor
Massimo Faccoli
Paper history
Received: Jun 16, 2016
Accepted: Oct 12, 2016
First online: Mar 16, 2017
Publication Date: Apr 30, 2017
Publication Time: 5.17 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2017
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 51728
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 43197
Abstract Page Views: 3770
PDF Downloads: 3522
Citation/Reference Downloads: 21
XML Downloads: 1218
Web Metrics
Days since publication: 3348
Overall contacts: 51728
Avg. contacts per week: 108.15
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2017): 6
Average cites per year: 0.67
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Post-fire recovery of the plant community in Pinus brutia forests: active vs. indirect restoration techniques after salvage logging
vol. 11, pp. 635-642 (online: 04 October 2018)
Research Articles
Identification of the ambrosia beetle Anisandrus dispar (Fabricius) (Coleoptera Curculionidae Scolytinae) using TaqMan™ probe assay on biological samples
vol. 16, pp. 182-187 (online: 30 June 2023)
Research Articles
Fuel characterization and crown fuel load prediction in non-treated Calabrian pine (Pinus brutia Ten.) plantation areas
vol. 15, pp. 458-464 (online: 03 November 2022)
Short Communications
Effective monitoring as a basis for adaptive management: a case history of mountain pine beetle in Greater Yellowstone Ecosystem whitebark pine
vol. 2, pp. 19-22 (online: 21 January 2009)
Review Papers
Increasing resistance and resilience of forests, a case study of Great Britain
vol. 17, pp. 69-79 (online: 21 March 2024)
Research Articles
Analysis of forest fires causes and their motivations in northern Algeria: the Delphi method
vol. 6, pp. 247-254 (online: 13 June 2013)
Research Articles
Influence of soil and topography on defoliation intensity during an extended outbreak of the common pine sawfly (Diprion pini L.)
vol. 10, pp. 164-171 (online: 19 November 2016)
Research Articles
Wildfire cause analysis: four case-studies in southern Italy
vol. 3, pp. 8-15 (online: 22 January 2010)
Research Articles
Distribution and abundance of the alien Xylosandrus germanus and other ambrosia beetles (Coleoptera: Curculionidae, Scolytinae) in different forest stands in central Slovenia
vol. 12, pp. 451-458 (online: 29 September 2019)
Research Articles
Allometric equations to assess biomass, carbon and nitrogen content of black pine and red pine trees in southern Korea
vol. 10, pp. 483-490 (online: 12 April 2017)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







