
Nitrogen removal and its determinants in hybrid Populus clones for bioenergy plantations after two biennial rotations in two temperate sites in northern Italy
iForest - Biogeosciences and Forestry, Volume 8, Issue 5, Pages 668-676 (2015)
doi: https://doi.org/10.3832/ifor1254-007
Published: Feb 02, 2015 - Copyright © 2015 SISEF
Research Articles
Abstract
The sustainability of bioenergy coppice plantations is strongly affected by the Nitrogen (N) balance, whose removal is very high due to the frequent harvest of large quantities of biomass composed of small-sized shoots. Poplar bioenergy coppice plantations could have a Nitrogen removal comparable to herbaceous crops. In this study, five hybrid poplar genotypes (“AF2”, “AF6”, “Monviso”, “83.148.041”, “I214”) were compared for tree morphological traits related to yield, N removal in the harvested biomass and Nitrogen wood concentration (N%) after two biennial coppice rotations in two experimental plantations located in northern Italy. N removal was primarily influenced by biomass production, and linear positive relationships between biomass yield and N removal were established. N removal also varied greatly among genotypes due to clonal differences in yield and in N%, in relation to significant differences among clones for their branching and sprouting habits. In the first rotation, branchiness was positively correlated to N% with a significant coefficient of determination (R2=0.813), while at the end of the second rotation it was also significantly correlated to the shoots per stool ratio (R2=0.804). “Monviso” and “83.148.041” were the clones showing the highest yield, but also a high N% associated to an high level of branchiness and shoots per stool ratio. Our results highlight that poplar genotype selection for sustainable N management should be aimed at genotypes with low wood N concentration, coupling high yield with low branching and sprouting habits as in the case of the clone “AF2”.
Keywords
Branching Habit, Coppice Plantations, Fertilization, Growth Traits, Sprouting Habit
Introduction
Tree bioenergy plantations have the potential to produce large quantities of biomass to partially replace fossil fuels, as well as to mitigate the global warming ([11]), provided that they are environmentally and socially sustainable. Compared to herbaceous energy crops, tree bioenergy plantations may be more sustainable for biofuel conversion in terms of energy and CO2 balance ([7], [47]). In many temperate countries, bioenergy coppice plantations of fast growing trees have been established with a short harvesting period of 2-3 years ([10], [18]). Such plantations are frequently referred to as short rotation coppices (SRCs). Tree species most frequently used for temperate SRCs include hybrids of willows (Salix spp.), poplars (Populus spp), eucalyptus (Eucalyptus spp.) and robinia species (Robinia pseudoacacia L.). The use of clones of such species with improved yield traits is very common.
SRC plantations are most frequently established on agricultural arable soils, and their cultivation cycle is fully mechanized, from establishing trees to harvesting the biomass. Despite this high level of mechanization, their energy balance is generally very positive ([17], [35]), with more than 60% of the crop energy balance represented by nitrogen (N) fertilization.
The importance of N fertilization depends on two main factors. Firstly, similar to other plant nutrient elements, N is largely removed with the frequent and complete cyclical harvest of the aboveground biomass. Nutrient removal in SRCs is far greater than in conventional forestry ([33], [23], [53]), due to the high density of the shoot population, characterized by very small sized stems and a high bark volume (wood/bark ratio - [31], [2], [22]). Secondly, the chemical synthesis of N fertilizers is an industrial process requiring an enormous quantity of energy ([30]).
A third element highlighting the importance of nitrogen fertilization for bioenergy sustainability lies in its environmental impact ([19], [27]), in terms of groundwater pollution by nitrate leaching and ozone depletion by the emission of nitrous oxides after the combustion of biomass with a relative high N concentration. Low requirements of N fertilizer and high efficiency of N utilization are therefore desirable crop traits, especially for energy crops. This is relevant to achieve a favorable, positive energy balance for the SRC crops ([48]).
Nevertheless, little is known about the nutrient use and efficiency of poplar SRCs, and how they vary according to factors such as site conditions, biomass yield and genetic origin of the planting material. Significant variations in N uptake and N use efficiency (NUE) have been found among clones of important fast-growing tree species such as poplars ([5], [25]), willows ([1]), and eucalypts ([45]). According to Vitousek ([54]), NUE indicates the net primary productivity (NPP) to total N uptake in a certain period. However, studying N balance in fast growing tree plantations, the aforementioned authors calculated NUE as the ratio between the harvested biomass over its N content, concluding that NUE is an important factor for the selection of planting material for SRCs. No attempts were made to connect the NUE of clones with clonal biomass yield characteristics, such as the distribution among stem/ shoot populations with different dimensions, the allocation ratio between branch and stem woody biomass, the shoot density or the number of shoots on the stools. Indeed, all these parameters may vary significantly among poplar clones. Such variability could affect N removal and NUE at a clonal level in poplar hybrids, altering the percentage of bark and young woody tissues in the total harvested woody biomass ([39]). This resulted from a high absorption of N, along with many other plant nutrients, by poplar trees, resulting in a higher N concentration in the bark than wood ([34]), and the percentage of bark to the total aboveground woody biomass strongly increases with decreasing stem dimensions ([22], [31]) and increasing branchiness ([39]).
The objective of this work was to examine the effects of coppicing cycles and hybrid poplar clones on stool morphology yield traits, wood N concentration (N%), NUE and N removal, with the aim of assessing clone suitability for bioenergy production and helping to refine the criteria of N fertilization for the woody crop in temperate plantation forestry.
In 2003, a network of three experimental plantations on arable soils was established in Italy in order to test new clones of poplar hybrids for SRC, by studying their biomass productivity and growth in relation to site characteristics and genetic background. Data on the first three biennial rotations were published in Paris et al. ([40]) and Sabatti et al. ([44]). In two plantations, woody samples were collected during the first two harvests from representative trees/shoots in order to determine the N concentration in woody tissue and to estimate the amount of N removed with harvesting.
Our aims were: (1) to investigate the effect of coppicing on N removal in the harvested biomass; (2) to test whether the studied poplar clones differed in their N use; and (3) to determine whether the morphological parameters of trees could be determinants in affecting wood N% at the clonal level.
Material and ethods
Locations and experimental settings
This study was conducted in two experimental plantations established in spring 2003 aimed at comparing hybrid poplar clones under SRC conditions. Both plantations, Bigarello and Vinovo, are located in the Po valley, northern Italy, and their main characteristics are summarized in Tab. 1. Detailed information on the experimental plantations are reported by Paris et al. ([40]). The two plantations were subject to very similar establishment and cultural practices, as reported in Tab. 1. The list of poplar clones compared in this study is reported in Tab. 2. Clone I-214 is a traditional selection developed in Italy for timber production. The other clones are new cultivars recently developed in Italy, specifically for biomass production under SRC management. These clones are currently under evaluation in Italy, as well in other European countries with various site conditions, such as Spain ([49]) and Romania ([21]).
Tab. 1 - Main characteristics of the Vinovo and Bigarello study sites with experimental plots of hybrid poplar clones under short rotation coppicing management in northern Italy. (a): Köppen & Greiger classification ([32])
| Characteristics | Vinovo | Bigarello |
|---|---|---|
| Long /Lat | 7° 38′ 20.40″ E 44° 57′ 25.03″ N |
10° 53’ 11″ E 45° 11′ 26.93″ N |
| Climate a | Cfb (Warm temperate, fully humid with warm summer) | Cfa (Warm temperate, fully humid with hot summer) |
| May to August Prec. (mm) | 60 mm in 2003; 143 mm in 2004; 233 in 2005; 198 in 2006 | 67 mm in 2003; 216 mm in 2004; 292 in 2005; 181 in 2006 |
| Soil (0-20 cm depth) |
(14.4% clay, 19.9% sand); pHwater 7; tot N soluble 10.1 mg N kg-1 | (43.3% clay, 16.8% sand); pHwater 8.34; tot N soluble 10.6 mg N kg-1 |
| Cultivation model |
Planting date: April 2003; planting density 5900 cuttings ha-1; biennial harvestings in Jan. 2005 and Dec. 2006; No fertilization |
Planting date: March-April 2003; planting density 5900 cuttings ha-1; biennial harvestings in Dec. 2004 and Jan. 2007; No fertilization |
Tab. 2 - List of hybrid poplar clones studied in northern Italy. (a): denotes where and by whom the crosses were made; (M) male; (F) female; (Yr): year of cross.
| Genotype | Sex | Parentage | Yr | Breedera | Selection for |
|---|---|---|---|---|---|
| AF2 | M | P. × canadensis | 1992 | F. Alasia, Savigliano (Italy) | biomass |
| AF6 | F | (P. × generosa) × P. nigra | 1993 | F. Alasia, Savigliano (Italy) | biomass |
| Monviso | F | (P. × generosa) × P. nigra | 1991 | F. Alasia, Savigliano (Italy) | biomass |
| 83.148.041 | M | P. × canadensis | 1983 | CRA-PLF, Casale M. (Italy) | biomass |
| I-214 | F | P. × canadensis | 1929 | G. Jacometti, Villafranca P. (Italy) | plywood |
The experimental design consisted in a randomized block with three-four replications per clone. Each replication was made up of homogenous plots with 60 trees organized in 5 rows (inter-row distance 280 cm), each with 12 trees (intra-row spacing 60 cm), with an inner sampling area of one row with 12 trees (20.16 m2).
No fertilization was supplied at any point along the cultural cycle of plantations, either before planting or during cultivation. Biomass harvesting, in biennial cycles, was performed mechanically in winter after leaf shedding.
Field sampling
Sampling and measurements were carried out at both plantations soon before mechanical harvesting in winter 2004/2005 and 2006/2007, at the end of the first and second biennial rotation cycle, respectively. Measurements were aimed at determining the main growth traits of the clones, i.e., stem dimensions and the number of shoots per stool. More details on the sampling procedures adopted and the allometric regression equations used are reported by Paris et al. ([40]).
The field sampling for nitrogen content was organized as follows. In each experimental site, the diameter at 100 cm above the ground (D) of all individual shoots was measured, and the shoot population of each clone recorded into diameter classes (diametric classes 1 cm wide; the numeric value of each class represented the central value, i.e., class 1 included all sections whose diameter ranged from 0.5 to 1.5 cm). Thereafter, 10-cm thick stem discs were obtained from 10-15 undamaged sample shoots (stem and branches with no evident damages such as wounds, pest and disease attacks) per each clone. The sampled shoots were randomly selected so that the diameter range in each experimental site and clone was well representing the whole variation across sites and clones.
For each sample shoot, D and total height (H) were recorded with a caliper (precision 0.1 mm) and tape (precision 0.1 cm), respectively, and the fresh weight of both stem and branches was directly measured in the field with an electronic portable hanging scale (HBC 20K 50, KERN, Germany - precision 50 g).
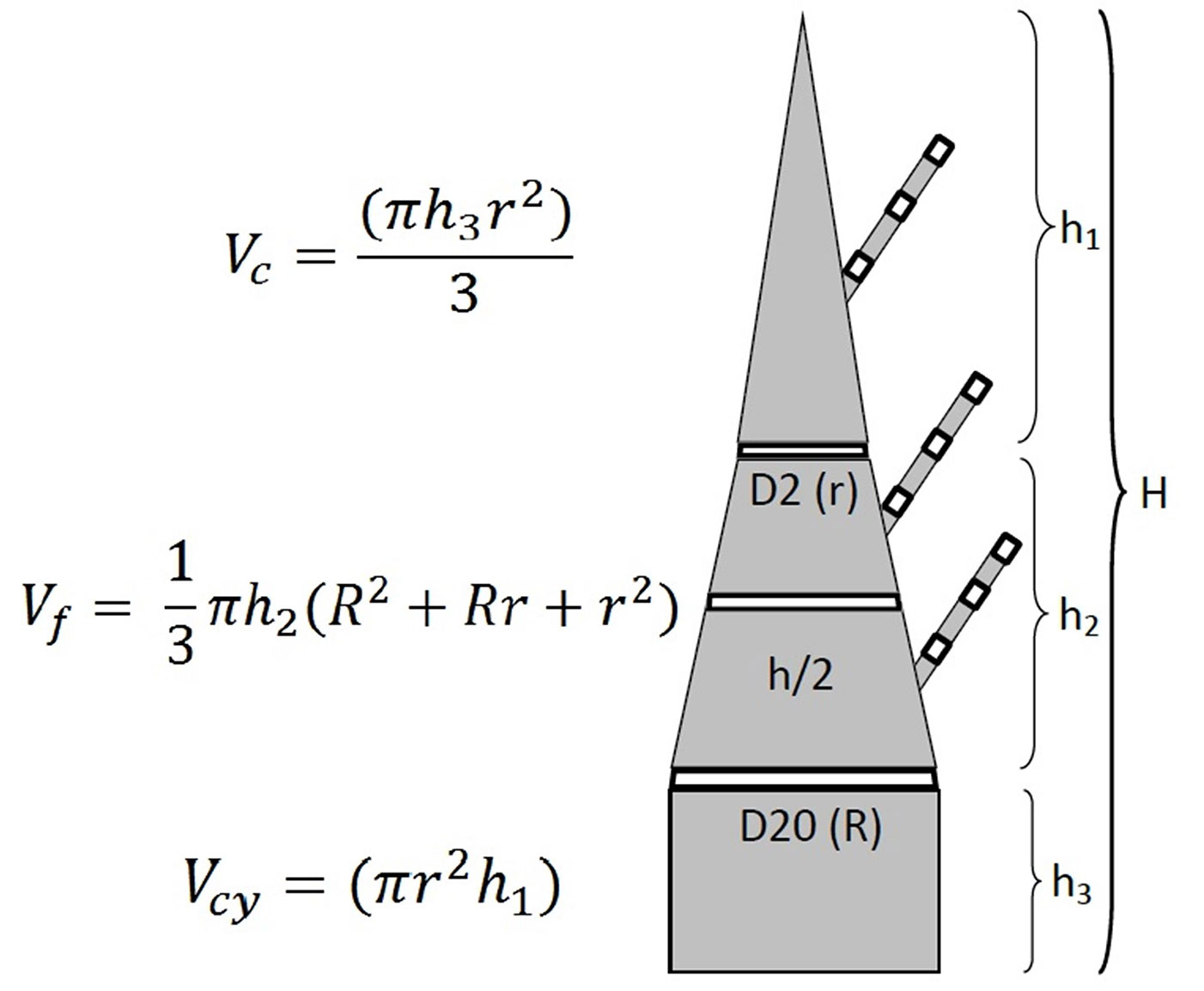
The stem discs were collected at three different heights above the ground (Fig. 1) measured with a tape (precision 0.1 cm): (i) 20 cm (disc D20, with R radius); (ii) at half of the total stem height (disc H/2); (iii) and at the height where the stem diameter was 2 cm (disc D2, with r radius). These discs represent 3 stem sections of the sample shoot: (i) the basal section, with a cylindrical shape and volume Vcy; (ii) the intermediate section (H/2), whose volume (Vf) can be estimated as a frustum of cone; (iii) the top section (D2) with a conic shape and volume Vc. Branches were sampled separately for each sample shoot, collecting 3 discs from their basal, median and apical part. Three average branches per shoot were selected along its entire height. Disc samples represented approx. 5% of the total weight of each individual sample shoot.
Fig. 1 - Schematic layout of wood discs sampled from the stem (D20; h/2, D2) and branches of sample shoots and the three main sections for the stem volumetric determination (basal, median and apical sections). (Vc): volume of the apical cone; (Vf): volume of the intermediate frustum of cone; (Vcy): volume of the basal cylinder; (h1, h2, h3): heights of the three stem sections considered; (H): total stem height; (R): lower radius; (r): upper radius.
Percentage branchiness (branch %) of each sample shoot was calculated as the branch weight to total shoot weight ratio.
Immediately after sampling, the radius of each disc was recorded by cross-measurement with a digital caliper (precision 0.1 mm). Each disc was then placed into a plastic bag for minimizing moisture loss.
Laboratory procedures and nitrogen analysis
One week after field collection, sample disc volume over bark was estimated by the water displacement method, thereby obtaining individual density estimates. Each disc was then split into two equal parts, one used for dry weight determinations (at 105 °C, to constant weight), and the other for N determinations.
Discs for N determinations were dried at 40 °C to constant weight, and then ground by a laboratory mill (M10 Basic Microfine Grinder, IKA, Staufen, Germany), obtaining a homogeneous dust which was dried again overnight at 40 °C, and subsequently analyzed by CHN gas chromatography (Carlo Erba NA 1500, Italy) to determine the relative percentage N concentration (N%).
Stem volume over bark was estimated for each sample shoot by summing the volumes of the 2 stem sections and the top section, using the formulas for the basal cylinder (Vcy = πr2 h1), for the intermediate frustum of a cone (Vf = 1/3 π h2 [R2 + Rr + r2]), and for the top cone (Vc=[πh3 r2]/3), respectively, where V is the volume, h1-3 are the height of the three stem sections, R the lower radius, and r the upper radius (see also Fig. 1). Conversion of fresh stem volume to stem dry weight was done using the recorded density of each stem disc.
Regarding the N content of three 3 stem sections, N concentrations were averaged over the discs at the basal (D20), intermediate (DH/2) and top (D2) sections, respectively. For the remaining branches, the branch N concentration was multiplied for the branch weight. The N content for each sample shoot was obtained by summing the N amounts of the three stem sections, including the branches wood nitrogen. The N uptake was then calculated per weight of each sample shoot according to its diameter class, and used for the estimation of total N uptake for each clone and site (N uptake, kg ha-1). NUE (kg dm kg N-1) was finally obtained dividing the above-ground woody biomass for each clone by its N uptake.
Statistical analysis
In each site, analysis of variance (ANOVA) was used to analyze the main effects of clone and cycle along with their interaction on growth parameters (D; H; Shoots/stool ratio; branch %) and N use parameters (N%; N uptake, NUE). This was carried out using a randomized complete block design with site, clone, cycle and clone × cycle as fixed factors and block as a random factor. Plot means for D, H, N shoots/stool ratio, N%, N uptake and NUE were the units of replication. Individual branchiness of test shoots after angular transformation (α = arcsin√%), was the unit of replication for branchiness. For significant ANOVA effects, post-hoc Duncan’s test (p ≤ 0.05) was used to test for significant differences between treatment means. Pearson’s correlation among growth and yield parameters (D; H; shoots/stool ratio; branch.%; Yield) and the different N use parameters (N%; N uptake) was also carried out. Average clonal values stratified per site, cycle and clone (n = 3-4) of the sampled parameters were used in this analysis. Finally, average plot values were used to analyze the relationship between Yield and N uptake, according to the two rotation cycles studied.
Results
Growth parameters
Results of the ANOVA carried out on growth parameters are shown in Tab. 3. At both sites, clone and cycle effects on stem or shoot dimensions (D and H) were generally highly significant after ANOVA. Coppicing significantly decreased both stem dimensions (D and H) in the two sites, with a more evident effect on D, which was reduced almost by half from the first to the second rotation at both sites (Tab. 4). Clone AF2 had consistently higher values in stem dimensions across the two rotation cycles, in both sites.
Tab. 3 - ANOVA F and p-values for morphological growth traits and Nitrogen (N) status of shoots of hybrid poplar clones after two rotations in the study sites (northern Italy). (1): Total shoot height; (2): Shoot diameter, measured at 100 cm above the ground; (3): percentage allocation ratios between stem and branch woody biomass;. (4): number of shoots on the stool; (5): percentage concentration of N in the above ground woody biomass, including wood and bark; (6): N content in the harvestable above ground woody biomass; (7): Nitrogen Use Efficiency.
| Site | Source of variation |
H1 | D2 | Branchiness 3 | Shoots/stool Ratio4 | N%5 | N Uptake6 | NUE7 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | F | p | F | p | F | p | F | p | ||
| Vinovo | Clone | 16.4 | <0.0001 | 5.3 | <0.01 | 4.6 | <0.01 | 0.6 | 0.696 | 313.2 | <0.0001 | 16.5 | <0.0001 | 208.3 | <0.001 |
| Cycle | 429 | <0.0001 | 396 | <0.0001 | 64.2 | <0.0001 | 360 | <0.0001 | 954.6 | <0.0001 | 34.4 | <0.0001 | 638.2 | <0.001 | |
| Site × Clone | 1.1 | 0.383 | 0.7 | 0.596 | 0.8 | 0.512 | 1 | 0.414 | 107 | <0.0001 | 8.1 | <0.01 | 48.4 | <0.0001 | |
| Bigarello | Clone | 27 | <0.0001 | 6.8 | <0.01 | 6.1 | <0.01 | 6.9 | <0.01 | 45.8 | <0.0001 | 16.5 | <0.0001 | 33.1 | <0.0001 |
| Cycle | 258 | <0.0001 | 350 | <0.0001 | 129.3 | <0.0001 | 846.3 | <0.0001 | 38.2 | <0.0001 | 34.4 | <0.0001 | 40 | <0.0001 | |
| Site × Clone | 12.3 | <0.01 | 0.7 | 0.602 | 5.5 | <0.01 | 7.3 | <0.01 | 8.2 | <0.01 | 8.1 | <0.01 | 9.9 | <0.0001 | |
Tab. 4 - Means and standard errors (in parenthesis) of total shoot height (H) and diameter (D), shoots/stool ratio, and percentage branchiness of hybrid poplar clones after two biennial rotations in the study sites in northern Italy. (§): Shoot diameter measured at 100 cm above the ground; (†): number of shoots on the stool; (‡): percentage allocation ratios between stem and branch woody biomass. Values within each column followed by different letters are statistically different after Duncan’s MR Test (P ≤ 0.05). (ns): not significant; (*): P ≤ 0.05,(**): P ≤ 0.01 ; (***): P ≤ 0.001, levels of significance of the differences between the average value of the first and second rotation cycle of each parameter in each site.
| Site | Clone | H (cm) | D (cm) § | Shoots/stool ratio † | Branchiness (%) ‡ | ||||
|---|---|---|---|---|---|---|---|---|---|
| 1st rotation | 2nd rotation | 1st rotation | 2nd rotation | 1st rotation | 2nd rotation | 1st rotation | 2nd rotation | ||
| Vinovo | AF2 | 844.5 (17.3) a | 628.0 (10.3) a | 6.3 (0.3) a | 3.5 (0.1) a | 1.2 (0.04) ns | 3.9 (0.29) ns | 15.5 (1.1) b | 8.5 (1.4) |
| AF6 | 764.2 (22.6) bc | 587.5 (2.7) ab | 5.4 (0.3) b | 3.1 (0.03) ab | 1.5 (0.06) ns | 4.2 (0.24) ns | 16.9 (1.3) b | 10.6 (1.3) | |
| Monviso | 731.0 (11.5) c | 510.7 (15.6) c | 5.4 (0.2) b | 2.9 (0.2) b | 1.3 (0.07) ns | 4.3 (0.34) ns | 24.1 (1.2) a | 13.2 (1.9) | |
| 83.148.041 | 816.5 (14.1) ab | 589.7 (17.4) bc | 5.2 (0.2) b | 3.1 (0.1) b | 1.1 (0.03) ns | 4.6 (0.31) ns | 19.6 (1.5) b | 9.8 (1.8) | |
| I-214 | 742.5 (3.6) bc | 559.2 (10.5) b | 5.5 (0.1) b | 2.9 (0.1) b | 1.4 (0.1) ns | 4.2 (0.41) ns | 17.5 (1.8) b | 10.9 (1.8) | |
| mean | 780.0 (11.9)*** | 575.0 (10.3) | 5.6 (0.1) *** | 3.1 (0.1) | 1.3 (0.05) *** | 4.2 (0.15) ns | 18.7 (0.7) *** | 10.6 (0.8) | |
| Bigarello | AF2 | 536.5 (13.8) a | 434.0 (9.1) c | 4.4 (0.2) b | 2.4 (0.2) a | 1.0 (-) d | 4.1 (0.09) c | 20.7 (1.3) b | 11.3 (1.5) e |
| AF6 | 476.2 (10.7) b | 319.2 (1.4) d | 3.8 (0.1) ab | 2.0 (0.1) b | 1.3 (0.06) d | 4.6 (0.33) c | 24.0 (2.3) bc | 19.3 (1.4) bcd | |
| Monviso | 504.8 (5.4) ab | 259.5 (10.1) e | 4.2 (0.1) a | 2.1 (0.1) ab | 1.2 (0.06) d | 5.3 (0.3) b | 30.2 (1.4) a | 14.9 (1.6) de | |
| 83.148.041 | 512.0 (7.8) ab | 420.5 (17.5) c | 3.8 (0.1) ab | 2.0 (0.1) b | 1.0 (0.02) d | 6.1 (0.22) a | 31.3 (1.7) a | 11.5 (1.2) e | |
| I-214 | 425.5 (18.1) c | 338.5 (11.9) d | 3.4 (0.3) b | 1.8 (0.3) b | 1.1 (0.04) d | 4.6 (0.25) c | 31.0 (1.7) a | 17.3 (2.4) cd | |
| mean | 491.0 (10.1)*** | 354.3 (15.4) | 3.9 (0.1) *** | 2.0 (0.1) | 1.1 (0.03) *** | 4.9 (0.18) | 27.5 (1) *** | 14.7 (0.8) | |
At both sites, clone and cycle effects on branchiness were generally highly significant. Coppicing strongly reduced plant branchiness in both sites, i.e, the first coppicing strongly decreased the amount of aboveground woody biomass allocated to branches. At both sites, clones “Monviso”, “83. 148.041” and “I-214” (at Bigarello) had the highest values of branchiness after the first biennial rotation, while the clone “AF2” had the lowest values. After the second rotation, the clone effect on branchiness was not significant at Vinovo, while a significant clone × cycle interaction in Bigarello, with clones “AF2” and “83.148.041” having the lowest values of branchiness.
At both sites, the shoots/stool ratio was affected by the rotation cycle, strongly increasing from the first to the second rotation as a consequence of coppicing. After the first rotation, no significant differences among clones were found for this parameter at both sites. After the second rotation, the clone effect was very significant in Bigarello. Clones “Monviso” and “83.148.041” had the highest values of the shoots/stool ratio in both sites, while clone “AF2” had the lowest.
Nitrogen use
Results of the ANOVA carried out on N use parameters are shown in Tab. 3. At both sites, clone and cycle effects were highly significant for N%, with a significant increment from the first to the second rotation (Tab. 5). “Monviso” and “83.148.041” clones had the highest values of wood N% across the two sites after the first cycle. These two clones showed the highest N% after the second cycle at Bigarello, while at Vinovo the clone “83.148.041” had the highest N% value along with clone “AF6”. After the first rotation, “Monviso” had the highest N uptake in both sites. After the second rotation, “83.148.041” had by far the highest N uptake.
Tab. 5 - Means and standard errors (in parenthesis) of Nitrogen (N) status in the shoots of hybrid poplar clones after two biennial rotations in the study sites (northern Italy). (§): percentage concentration of N in the above ground woody biomass, including wood and bark; (†): N content in the harvestable above ground woody biomass; (‡): Nitrogen Use Efficiency. Values within each column followed by different letters are statistically different after Duncan’s MR Test (P ≤ 0.05). (*): P ≤ 0.05; (**): P ≤ 0.01; (***): P ≤ 0.001, levels of significance of the differences between the average value of the first and second rotation cycle of each parameter in each site.
| Site | Clone | N % § | N uptake (Kg ha-1) † | NUE ‡ | |||
|---|---|---|---|---|---|---|---|
| 1st rotation | 2ndrotation | 1st rotation | 2nd rotation | 1st rotation | 2nd rotation | ||
| Vinovo | AF2 | 0.59 (0.016) f | 0.80 (0.003) d | 165.7 (4.4) de | 210.5 (16.1) cd | 168.8 (43) a | 125.2 (0.4) d |
| AF6 | 0.75 (0.013) c | 1.05 (0.003) b | 225.9 (3.7) c | 286.0 (15.8) b | 133.2 (2.2) bc | 96.3 (0.3) f | |
| Monviso | 0.96 (0.020) e | 0.96 (0.060) c | 245.5 (5.1) bc | 250.3 (16.7) bc | 104.2 (2.2) e | 103.8 (0.6) e | |
| 83.148.041 | 0.73 (0.016) e | 1.15 (0.002) a | 164.5 (3.6) de | 340.8 (18.3) a | 137.8 (3.0) b | 87.0 (0.2) g | |
| I-214 | 0.60 (0.003) f | 0.78 (0.005) d | 132.2 (0.7) e | 172.2 (10.6) de | 165.4 (0.9) a | 127.5 (0.8) cd | |
| mean | 0.73 (0.040) *** | 0.95 (0.030) | 186.8 (11) *** | 252.0 (15) | 141.9 (6.2) *** | 108.0 (3.6) | |
| Bigarello | AF2 | 0.65 (0.024) f | 0.86 (-) cd | 73.1 (2.7) bc | 129.3 (14.9) bc | 155.4 (5.61) a | 116.6 (0.02) cd |
| AF6 | 0.73 (0.036) e | 0.89 (0.001) bc | 84.2 (4.2) b | 102.6 (10.5) bc | 138.0 (7.23) b | 112.7 (0.08) cd | |
| Monviso | 0.91 (0.031) bc | 0.93 (0.015) b | 123.1 (4.2) a | 175.1 (29.8) ab | 110.7 (3.91) de | 107.4 (1.89) de | |
| 83.148.041 | 1.00 (0.004) a | 1.00 (0.019) a | 111.3 (0.4) a | 222.6 (42.5) a | 100.2 (0.41) e | 99.8 (2.18) e | |
| I-214 | 0.75 (0.002) e | 0.82 (0.012) a | 54.4 (0.8) c | 69.8 (14.0) c | 133.3 (1.85) b | 121.9 (2.13) c | |
| mean | 0.81 (0.030) *** | 0.90 (0.020) | 89.2 (7.4) ** | 139.9 (16.5) | 127.5 (5.5) *** | 111.7 (1.8) | |
NUE was significantly affected by clone and cycle effects at both sites, with a significant decrease from the first to the second rotation.
Correlations, ield and N uptake
N% and NUE were never correlated with the shoot dimensions (Tab. 6), meaning that D and H did not affect the N use parameters of the studied clones. On the contrary, several significant correlation coefficients were found between the branching and sprouting habits and the N use parameters. Fig. 2 shows the linear regressions between branchiness and wood N% at both sites and across the two rotation cycles. The regression coefficients were never significant after the second rotation cycle, when branchiness remarkably decreases as a consequence of the strong increase of shoot density after coppicing. At the end of the first cycle, with higher values of branchiness, the regression coefficient was R2 = 0.813 (p ≤ 0.05) at Vinovo and R2 = 0.621 (p ≤ 0.10) at Bigarello. In both cases, clone “AF2” showed the lowest values of branchiness and the lowest values of wood N%. In contrast, clones “Monviso” and “83.148.041” showed high values of branchiness and high wood N%.
Tab. 6 - Pearson’s correlation coefficients (R2) between Nitrogen use parameters and morphological yield traits of hybrid poplar clones after two biennial rotations in the study sites (northern Italy). (1): Shoot diameter, measured at 100 cm above the ground; (2): total shoot height; (3): percentage allocation ratios between stem and branch woody biomass;. (4): number of shoots on the stool; (5): percentage concentration of N in the above ground woody biomass, including wood and bark; (6): N content in the harvestable above ground woody biomass; (7): Nitrogen Use Efficiency.
| Variable | Vinovo | Bigarello | ||||||
|---|---|---|---|---|---|---|---|---|
| 1st rotation | 2nd rotation | 1st rotation | 2nd rotation | |||||
| N%6 | NUE7 | N% | NUE | N% | NUE | N% | NUE | |
| D1 | 0.298 | 0.385 | -0.055 | 0.065 | 0.040 | -0.076 | 0.004 | 0.001 |
| H2 | -0.299 | 0.271 | -0.006 | 0.018 | 0.014 | 0.0003 | 0.012 | -0.006 |
| Branchiness (%)3 | 0.813* | -0.747** | 0.017 | -0.039 | 0.622 | -0.701** | -0.205 | 0.192 |
| Shoots/stool ratio4 | 0.041 | 0.045 | 0.558 | -0.505 | -0.012 | 0.001 | 0.804* | -0.761* |
| Biennial Yield5 | 0.014 | -0.018 | 0.708^ | -0.694 | 0.097 | -0.074 | 0.839* | -0.844* |
Fig. 2 - Linear regressions between the percentage woody biomass allocated to the branches (branchiness, in %) as the independent variable, and the percentage N concentration in the wood (N%) as the dependent variable, for the five clones in the two study sites (northern Italy), in relation to first and second biennial rotations. Each point represents the mean value of 3 replications. (*): P ≤ 0.05; (^): P ≤ 0.1. See Tab. 2, Tab. 4 and Tab. 5 for additional information.
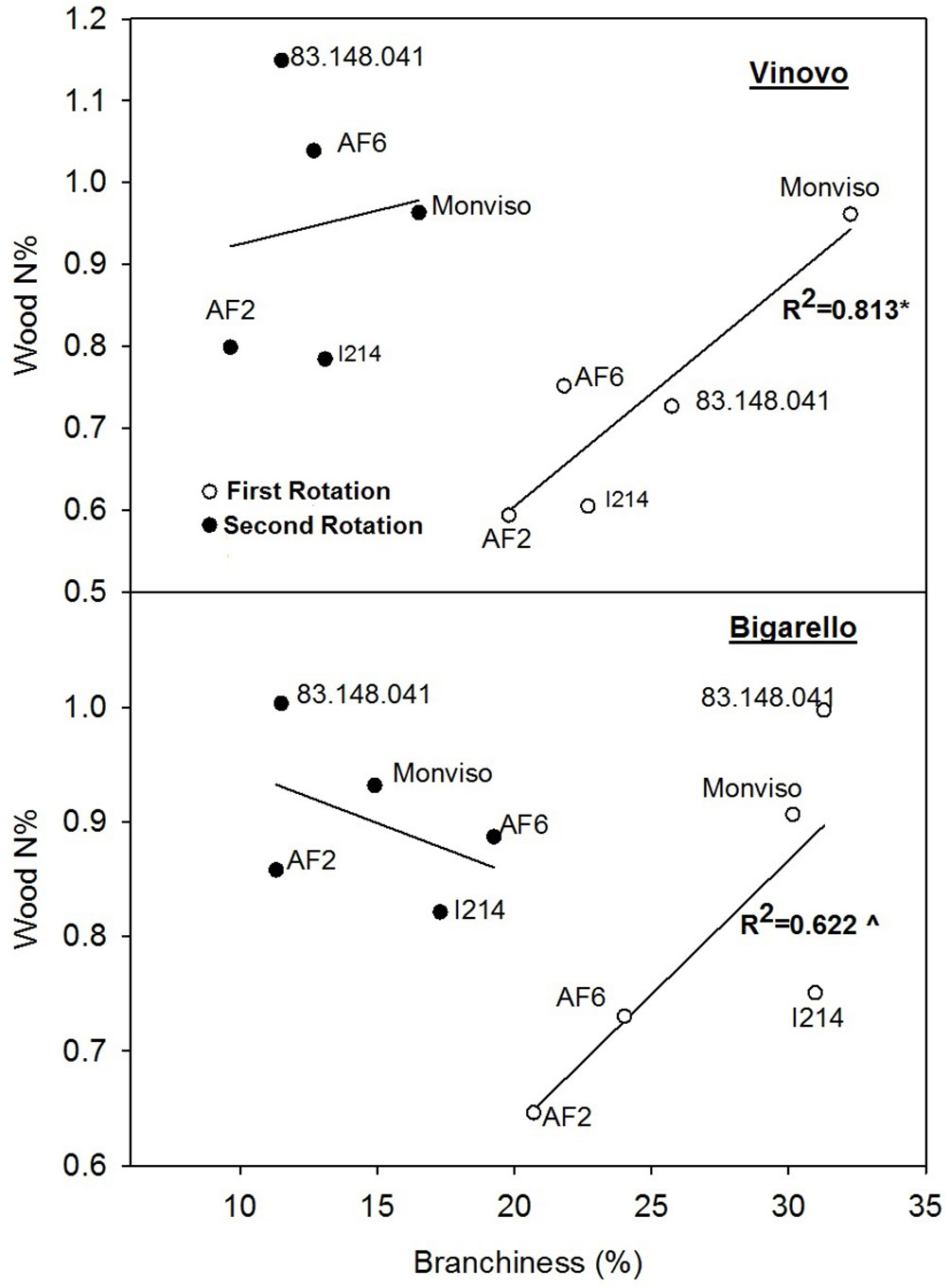
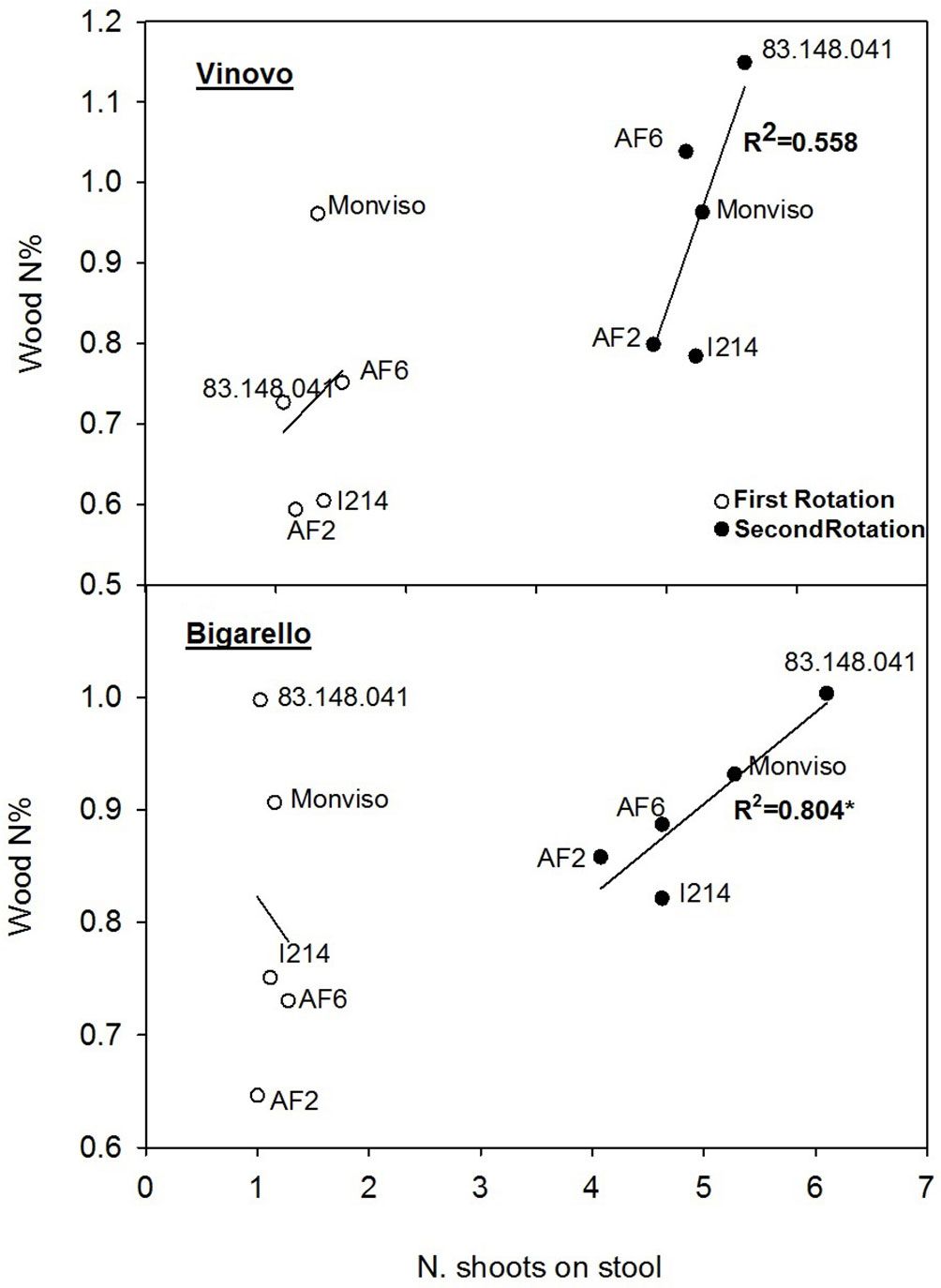
Fig. 3 shows the linear regressions between the shoots/stool ratio and wood N% in the two sites and across the two rotation cycles. The regression was never significant after the first rotation cycle. After the second cycle, with higher values of shoots/stool ratio, the regression coefficient were R2 = 0.804 (p ≤ 0.05) at Bigarello and R2 = 0.56 (p ≤ 0.10) at Vinovo. In both cases, the lowest values of shoots/stool ratio and wood N% were observed for clones “AF2” and “I214”. In contrast, “83.148.04”1 had the highest values of shoots/stool ratio and the highest wood N%.
Fig. 3 - Linear regressions between the shoots/stool ratio (the number of shoots on the stool) as the independent variable, and the percentage N concentration in the wood (N%), as the dependent variable, for the five clones in the two study sites (northern Italy), in relation to first and second biennial rotations. Each point represents the mean value of 3 replications. (*): P ≤ 0.05; (^): P ≤ 0.1. See Tab. 2, Tab. 4 and Tab. 5 for additional information.
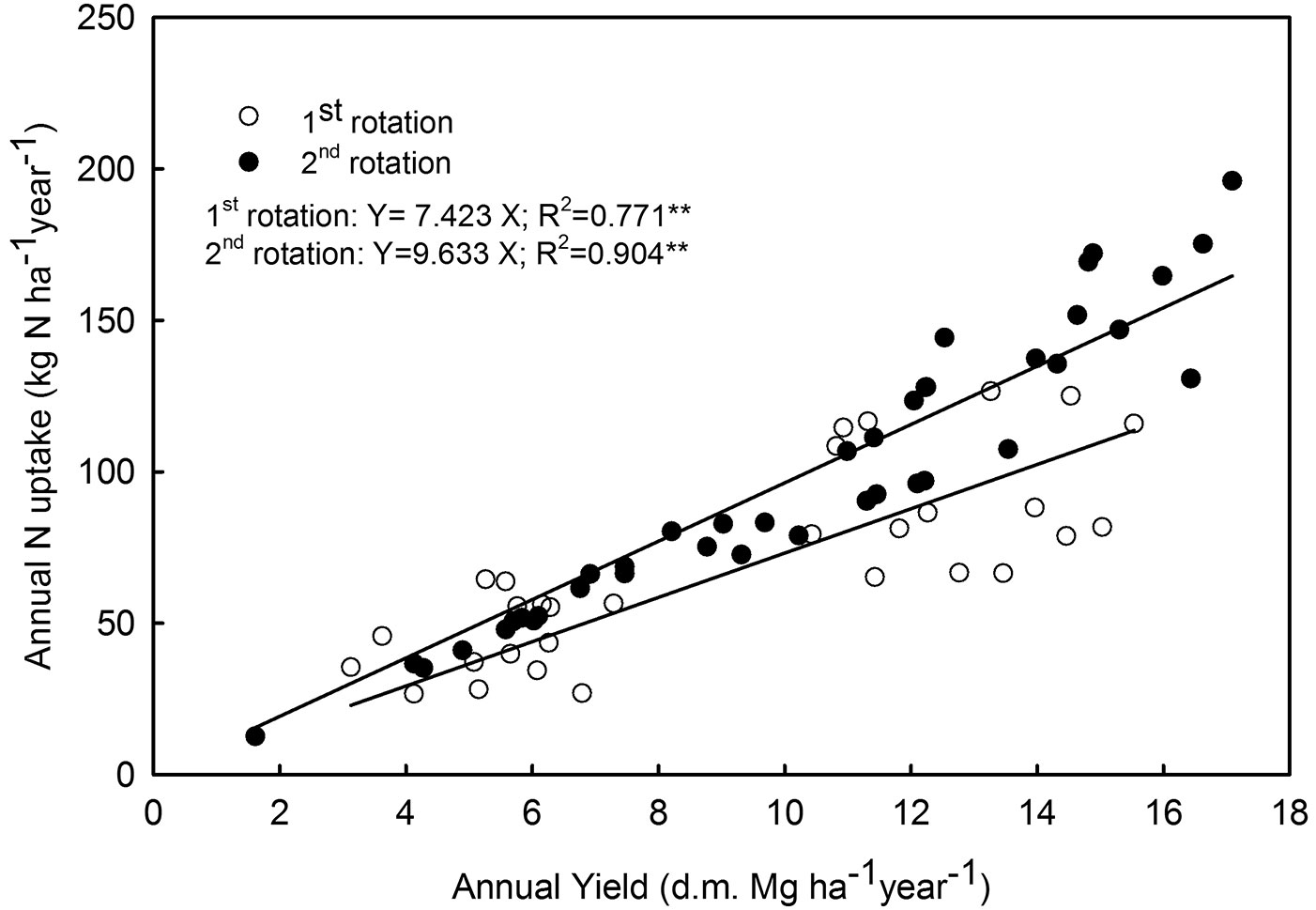
Yield data as aboveground woody biomass of the studied poplar clones are reported in Tab. 7. N uptake in the aboveground woody biomass of the five clones in relation to the corresponding annual yield across the two study sites and the two rotation cycles are shown in Fig. 4. Data are reported as single replication values. These data are well stratified into two different linear regressions for the first and the second rotation cycle.
Tab. 7 - Means and standard errors (in parenthesis) of dry matter (d.m.) biennial above ground woody biomass yield (Mg dm ha-1) of hybrid poplar clones after two biennial rotations in the study sites in northern Italy. Readapted from Paris et al. ([40]). Within each column, values followed by different letters are statistically different according to Duncan’s MR Test (P ≤ 0.05). (ns): not significant difference between the yield average values of the first and second rotation cycle in each site.
| Clone | Vinovo | Bigarello | ||
|---|---|---|---|---|
| 1st rotation | 2nd rotation | 1st rotation | 2nd rotation | |
| AF2 | 27.93 (3.81) ab | 26.37 (2.40) a | 11.31 (1.78) ab | 15.07 (2.01) ab |
| AF6 | 30.07 (2.50) a | 27.56 (1.86) a | 11.52 (1.58) ab | 11.57 (1.36) ab |
| Monviso | 25.53 (3.26) ab | 26.02 (2.13) a | 13.58 (1.73) a | 18.62 (3.26) a |
| 83.148.041 | 22.64 (1.63) b | 29.66 (1.86) a | 11.16 (0.50) ab | 22.24 (8.87) a |
| I214 | 21.86 (1.58) b | 21.93 (1.43) b | 7.25 (1.69) b | 8.40 (1.87) b |
| Mean | 25.60 (2.55) ns | 26.31 (1.94) | 10.96 (1.46) ns | 15.18 (3.46) |
Fig. 4 - Linear regressions between annual yield in dry matter (d.m.) woody biomass as independent variable and Nitrogen (N) uptake in the harvestable biomass as dependent variable for the 5 clones in the two study sites in northern Italy, in relation to first and second biennial rotations. Each point represents a single replication (plot) value. (**): P ≤ 0.01.
Discussion
Nitrogen removby harvest and its implications for assessing N fertilization in SRCs
Bioenergy plantations of hybrid poplars, under short rotation coppicing (SRC) management are intensive systems of forestry plantation, requiring optimal site conditions in terms of soil quality, nutrient availability and soil moisture balance, as well intensive cultural practices such as weed control, irrigation and fertilization ([24], [22], [41], [4], [44]). Consequently, poplar SRC should be considered and managed as a system closer to agriculture than to silviculture. The importance of N in promoting poplar tree nutrition, growth and biomass yield has often been demonstrated in different site conditions and management options ([13], [26], [8], [12], [6]). In this study, we obtained high values of N removal in the woody biomass harvested in the first and second biennial cycles (Fig. 4). Comparison with similar studies in the literature is difficult ([1], [29], [52], [52]), mainly because N removal with SRC harvesting is affected by many factors, such as the yield, the rotation cycle, the fertilization and irrigation regime. Indeed, all of the aforementioned authors did not develop a relationship between annual yield and N removal.
The two plantations considered in this study were established on alluvial arable soils with contrasting soil qualities within a larger network of plantations aimed at testing SRC poplar clones across Italy ([40], [44]. Annual clonal yields ranged between 5-25 dry matter (dm) Mg ha-1year-1 ([36], [40]). The clones chosen for this study are among the best performing in SRC in Italy, along with the clone “I214” which is the most used for timber plantations across Italy, mainly in the continental northern part. Both the study sites are located in the Po plain, the largest and most fertile agricultural area in northern Italy. Bigarello was the poorest site in the plantation network, with soil texture (43.3% clay) and pH (8.34) outside of the optimal range for hybrid poplar cultivation in Italy. Contrastingly, Vinovo was an average site in terms of biomass yield and site characteristics. Total soluble N in the soil of the two study sites is quite similar, and no fertilization was carried out at both sites during the study.
Using the data collected in these experiments, we established a relationship between the annual yield of aboveground biomass and N removal for poplar SRC in Italy (Fig. 4). This relationship may be considered a useful tool for assessing the fertilization balance of poplar SRC plantations for nitrogen. However, these data must be used with caution and fertilization needs to be tailored to the specific poplar SRC plantation. Although N removal by harvest is relevant, it is just one factor affecting N cycling, which mainly depends on the balance between N losses (leaching, denitrification and N uptake in stumps/root biomass) and inputs (N from natural depositions, annual leaf fall and fine root mortality - [24]). Generalization is hard to achieve both for N inputs and outputs, since many of these factors are site- and/or crop-specific.
Three main general observations regarding tailoring N fertilization in poplar SRC plantations may be highlighted here. Firstly, N uptake values in the aboveground woody biomass of poplar SRCs are comparable to those of many temperate agricultural crops ([37]). Secondly, at the same level of productivity, the need for N input increases considerably between the first and the second cycle of poplar SRC plantations (Fig. 4). In fact, shoot density increased dramatically during the second cycle due to coppicing, thus reducing shoot dimensions (Tab. 4). This has also been observed by other authors on shoot density and shoot diameter after coppicing, both in hybrid poplar and willow SRC plantations ([1], [3]). The third general observation is that in SRCs coppicing led to a significant shift in shoot population density and dimensions, thus altering the bark and nutrients percentages in the above ground woody biomass, with a concomitant increment in N% in the harvest.
Contribution of yield traits and determinants N removal
We found an extremely high variability in average annual removal of N amongst the studied clones (Tab. 5). This clonal variability was determined more by annual yield than N%. For each clone, the coefficient of variation (CV%) for biomass yield was always higher than 28.5%, while the CV% for N% was never higher than 18.2% (data not shown). This is in agreement with Adegbidi et al. ([1]) who studied biomass yield and nutrient uptake and concentrations on the woody biomass of willow and poplar clones for SRC in the USA. In our study, clone “I214” had the lowest N uptake always and the lowest yield. The clone with the highest N uptake was “83.148.041”, which was also the most productive clone in our experiments. This means that N% is a secondary parameter as compared with biomass yield in affecting clonal variability in N uptake amongst the hybrid poplar clones for SRC. N% varied between a minimum of 0.59% for “AF2” in Vinovo during the first rotation to a maximum of 1.15% for “83.148.041” in Vinovo during the second rotation. These values are extremely high when compared with those reported by other authors who studied N% in the woody biomass of Salicaceae clones under SRC management. For example, Jug et al. ([29]) reported a variation range in N% between 0.27-0.69% for three clones (two poplars and one willow) in Germany under a short rotation management with a 5-year coppicing cycle and biomass yield ranging between ca. 10-14 Mg dm ha-1 year-1 ([28]). In this case, the low N% in woody biomass may be explained by a rotation cycle longer than that adopted in our study. Moreover, Adegbidi et al. ([1]) also investigated nutrient concentrations in the woody biomass of five willow and one hybrid clones in the New York State, with N% ranging from 0.38 to 0.93, according to 1-3 year coppicing rotation cycles. Such values were surprisingly lower as compared with our study, mainly considering the shorter rotation cycles and the higher number of sprouts per stool of willow SRCs. However, no detailed information on sampling for chemical analysis were reported therein.
Our study highlighted the importance of productive determinants in Salicaceae clones such as branching and the shoots/stool ratio in affecting N% in the woody biomass produced by SRC plantations ([9], [46], [55], [15]). We found significant and positive regression coefficients between Branch% and N% during the first rotation cycle at both study sites (Tab. 6). After coppicing (second rotation), N% became strongly related to the shoots/stool ratios, with an increment in shoot density and a decrease in branch%. Such evidence can be easily explained by the fact that both branch% and shoots/stool ratio should increase the bark percentage, thus increasing N% in the total woody biomass ([39]).
Two ideotypes of hybrid poplar clones with successful yield performance, but with opposite N% have been identified by this study. Clones like “AF2” combined low N% with low values of branch% and shoots/stool ratio during the first and second cycles, respectively. In contrast, clones like “Monviso” and “83.148.041” presented high N% with high values of the aforementioned morphological traits. Ceulemans et al. ([9]) and Scarascia-Mugnozza et al. ([46]) demonstrated the importance of sylleptic branches in improving growth and yield of hybrid poplar clones. More recently, several pedigree studies showed that successful genotype ideotypes in growth and yield can exhibit opposite morphological characteristics as for branching ([50], [51], [42], [15]) and sprouting habits ([42]). Our experiments showed that high yielding clones, such as “Monviso” and “83.148.041” ([40], [44]), combine high branching and sprouting habit with low NUE. In contrast, “AF2” presents moderate to good yield performance ([40], [44]), low branching and sprouting habits, as well as high NUE.
Conclusions
Nitrogen balance is a factor of considerable importance for the sustainability of bioenergy crops, in terms of their energy balance, greenhouse gas emission, environmental contamination, and biofuel quality ([11]). Selecting bioenergy crops with a low need of N fertilization is therefore crucial, as well as a low level of N content in the harvested and processed biomass. Tailoring N application to the real need of the crop is also a viable strategy for reducing N contamination in the environment and N content in the harvested biomass.
The N removal by harvesting was studied in two poplar coppice bioenergy plantations during the first two rotation cycles of new hybrid clones, specifically selected for intensive management with high planting density (about 6000 plants ha-1) and a very short harvesting cycle (2 years). These kinds of agroforestry systems are intensive forms of bioenergy plantations of increasing importance in many countries with temperate climatic conditions ([10], [18]). The studied clones are currently used at a commercial level in Italy and other European countries.
N removal was found to be primarily influenced by biomass production. A significant relationship between biomass yield and N removal was detected. Such relationship can be used to refine N fertilization balance for the woody crop, both using chemical fertilizers and organic waste for disposal in bioenergy plantations (phytoremediation - [43], [16]). Coppicing improved the amount of N removal dramatically, as a consequence of strong changes in the morphological traits and density of shoot populations. Morphological shoot traits were found to be correlated at a clonal level to N concentration in the woody tissues, as well as to the NUE of the poplar clones. Low branchiness was related to low N concentration in the harvested wood. A low number of shoots per stool after coppicing was also found to be correlated with low N% and high NUE. Clone “AF2” had high NUE, low branchiness and a low shoots/stool ratio, combined with moderate to good biomass production in the present study. Clone “AF2” is also a productive clone with a stable yield across sites and coppicing cycles, s shown by other experiments in Italy ([38], [20], [14], [44]).
Our results demonstrate the feasibility of cultivar selection in poplars for SRC aimed at improving the NUE of genotypes, along with their biomass yield, the low branchiness and a low number of shoots per stool, as in the case of clone “AF2”.
Acknowledgments
This study was funded by the Consiglio per la Ricerca e la Sperimentazione in Agricoltura, Italy, “BIOENERLEGNO Project 2008-09”, and by the European transnational Project “AgroCop: Maximizing Timber and Energy Wood Production by Innovative Agroforestry Systems with Short Rotation Coppice as Intercrop, 2012-14”, WoodWisdom-Net - ERA-NET Bioenergy. L. Tosi’s PhD fellowship was funded in equal parts by the project AgroCop and by the University of Tuscia, Viterbo, Italy. The authors acknowledge two anonymous reviewers for their useful comments on the manuscript.
References
CrossRef | Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Authors’ Info
Authors’ Affiliation
Luca Tosi
C.N.R.- Istituto di Biologia Agroambientale e Forestale, v. G. Marconi 2, I-05010 Porano (TR - Italy)
Corpo Forestale dello Stato, v. A. Pacinotti 5, I-01100, Viterbo (Italy)
Luca Tosi
Giuseppe Scarascia-Mugnozza
Di.B.A.F.- Università degli Studi della Tuscia, v. S. Camillo de Lellis, I-01100 Viterbo (Italy)
Corresponding author
Paper Info
Citation
Paris P, Mareschi L, Sabatti M, Tosi L, Scarascia-Mugnozza G (2015). Nitrogen removal and its determinants in hybrid Populus clones for bioenergy plantations after two biennial rotations in two temperate sites in northern Italy. iForest 8: 668-676. - doi: 10.3832/ifor1254-007
Academic Editor
Gianfranco Minotta
Paper history
Received: Jan 28, 2014
Accepted: Oct 30, 2014
First online: Feb 02, 2015
Publication Date: Oct 01, 2015
Publication Time: 3.17 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2015
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 56011
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 46787
Abstract Page Views: 3719
PDF Downloads: 3922
Citation/Reference Downloads: 29
XML Downloads: 1554
Web Metrics
Days since publication: 4163
Overall contacts: 56011
Avg. contacts per week: 94.18
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2015): 11
Average cites per year: 1.00
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Wood production and nutritional status of Pinus taeda L. in response to fertilization and liming: a meta-analysis of the Americas
vol. 16, pp. 195-201 (online: 25 July 2023)
Research Articles
Variation of wood and bark density and production in coppiced Eucalyptus globulus trees in a second rotation
vol. 9, pp. 270-275 (online: 08 September 2015)
Research Articles
Preliminary study on genetic variation of growth traits and wood properties and superior clones selection of Populus ussuriensis Kom.
vol. 12, pp. 459-466 (online: 29 September 2019)
Research Articles
Contrasted growth response of hybrid larch (Larix × marschlinsii), jack pine (Pinus banksiana) and white spruce (Picea glauca) to wood ash application in northwestern Quebec, Canada
vol. 14, pp. 155-165 (online: 06 April 2021)
Research Articles
Nursery fertilization affected field performance and nutrient resorption of Populus tomentosa Carr. ploidy levels
vol. 15, pp. 16-23 (online: 24 January 2022)
Review Papers
Pathways to increase supply of sustainable wood from planted forests and trees to address bioeconomy needs: a review
vol. 19, pp. 168-185 (online: 12 May 2026)
Research Articles
Combined pre-hardening and fall fertilization facilitates N storage and field performance of Pinus tabulaeformis seedlings
vol. 9, pp. 483-489 (online: 07 January 2016)
Research Articles
Effects of thinning and pruning on stem and crown characteristics of radiata pine (Pinus radiata D. Don)
vol. 10, pp. 383-390 (online: 16 March 2017)
Research Articles
Physical, chemical and mechanical properties of Pinus sylvestris wood at five sites in Portugal
vol. 10, pp. 669-679 (online: 11 July 2017)
Research Articles
Adaptability and interspecific variability in growth and leaf traits of eucalypt
vol. 14, pp. 560-568 (online: 09 December 2021)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







