Wood production and nutritional status of Pinus taeda L. in response to fertilization and liming: a meta-analysis of the Americas
iForest - Biogeosciences and Forestry, Volume 16, Issue 4, Pages 195-201 (2023)
doi: https://doi.org/10.3832/ifor4296-016
Published: Jul 25, 2023 - Copyright © 2023 SISEF
Research Articles
Abstract
Loblolly pine (Pinus taeda L.) is one of the most planted forest species in the Americas. Since few studies have comprehensively assessed loblolly pine responses to fertilization, the present study performed a meta-analysis of the Americas based on 44 publications (1970-2022) of loblolly pine fertilization under field conditions. In general, fertilization increased root dry matter (+33%), litter (+21%), plant height (+6%), trunk diameter (+9%), wood yield (+30%), and needle concentrations of P (+9%), K (+36%), Ca (+17%), Mg (+14%), and S (+12%). Wood production was higher with residue fertilization, primarily with use of composite residues (cellulosic sludge + ash), compared to mineral fertilization. In regards to mineral applications, wood production was higher when multiple nutrients were added from fertilization and liming operations. Applications at planting (< 1 year) or on established trees (2-8 years), showed similar increases in wood production with higher responses occurring on sandy soils. These factors generally increased needle nutrient concentrations, except for no alteration or slight decreases in N under most conditions. The present study revealed loblolly pine responses to contrasting application strategies, which can help identify efficient fertility management practices for this commercially significant tree species.
Keywords
Loblolly Pine, Planted Forest, Waste, Sandy Soil, Needle Composition
Introduction
Loblolly pine (Pinus taeda L.) is the most planted conifer in the southeastern United States of America (USA) and is native to this region. In Brazil and other South American countries, loblolly is also widely cultivated in support of several timber industries ([26]). In the last few decades, natural regeneration of harvested forests in the southeastern USA has moved toward systematic introduction of improved seedlings and intensification of weed control, soil preparation, and fertilization ([21], [14], [15]). Where introduced in South America, similar silvicultural practices have been largely followed, except for fertilizer use, even if plantings occurred on low fertility soils ([31]). In Brazilian planted pine environments, soil nutrient depletion suggests a need for implementing fertilizer and lime applications ([42], [22]).
Nutrient application should be directed towards elements limiting pine growth, which can vary widely depending on soil parent material and can be strongly influenced by soil attributes such as texture and native fertility. In the US, nitrogen (N) responses have been reported for soils with organic matter levels of 1.33 and 0.37% for six sites under Pleistocene terraces and seventeen sites under other soil formations, respectively ([14]). These organic matter levels were much lower than the 5.2, 4.2, 3.3, and 2.4% noted for soils originating from basalt, granite, claystone, and sandstone in Araucaria angustifolia native forest of southern Brazil ([25]), which were replaced by pine production areas. Such observations may help explain why few studies have evaluated N fertilization in this region. In contrast, pine responses to phosphorus (P) application have frequently been reported for the Americas ([14], [2], [17], [18]). Unlike P, even soils with low available potassium (K) have shown lower response ([15]) or no response ([9], [18]). Use of micronutrients has garnered some interest since their application with macronutrients has been shown to increase tree production ([14]).
Use of limestone and alkaline organic residues to elevate soil pH and supply of calcium (Ca) and magnesium (Mg) has been evaluated since Mg deficiency (associated with or without Ca deficiencies) has been reported for pines in the USA and Brazil ([16], [38], [1]). Alkaline residues may also contain several macro- and micronutrients that increase the possibility of pine responses ([39], [41], [36]). In such cases, joint application of more than one nutrient could result in synergistic effects ([14], [2]).
As a function of fertilizer and soil pH corrective applications, changes in loblolly pine needle nutrient concentrations have exhibited variability between sites. Sypert ([43]) evaluated addition of 10 elements (N, P, K, Ca, Mg, S - sulfur, Mn - manganese, Zn - zinc, B - boron, and Cu - copper) at several locations and reported increased foliar concentrations for 9, 6, 4, and 1 elements for sites in Georgia, Texas, South Carolina, and Alabama, respectively. In this same study, a decrease was registered for 4 elements at South Carolina sites and 1 element at Texas sites, which was possibly related to a dilution effect and/or an interaction. Similarly, Pereira et al. ([33]) associated residue use with decreased foliar P, Ca, S, Fe, Mn, and B and increased tree growth, which suggests a significant dilution effect.
Given the long growth cycles of loblolly pine, time of application can also play a role in responses to nutrient and soil corrective applications. Such applications can commonly occur at planting or mid-rotation following first or second thinning operations. Application at planting can correct for deficient elements and favor initial growth ([30], [31]). However, applications at stand initiation can lead to slow tree growth and nutrient losses due to reduced ability to intercept and absorb highly mobile nutrients (i.e., less developed and inefficient root system). Mid-rotation fertilization can favor greater growth of trees with ample root systems and absorption capacity. The formation and accumulation of litter following nutrient and corrective applications can allow for retention within this horizon, thereby contributing to sources of Cu, Zn, and soil acidity correctives ([1]).
An integrated evaluation of factors influencing fertilization efficiency is a complex and onerous task when conducted using conventional experimentation. However, several investigations have taken an integrative approach of evaluating plant responses to different management practices through meta-analysis of data from the published literature base ([29], [11], [12]). For example, a meta-analysis study evaluating the effects of liming and wood ash on forest ecosystems showed that wood ash provided a greater increase in growth and wood production compared to liming alone ([37]).
In the present study, a meta-analysis was conducted to evaluate the efficiency of fertilization on the production and nutritional status of loblolly pine under various growth conditions in the Americas. Our hypothesis was that fertilization and liming, whether with mineral or residue applications, favors the growth and nutrition of loblolly pine, especially when multiple nutrients were provided.
Material and methods
Literature search and data compilation
A search for publications that evaluated the response of loblolly pine to fertilization was performed on Google Scholar® between October 2021 and June 2022. The following combinations of terms were used in this search: “Pinus taeda”, “fertilization”, “America” and “Brazil”, or “USA”. We reviewed each publication to determine whether these studies met the following criteria: (1) study conducted under field conditions in the Americas; (2) contained treatments without fertilization (control treatment) and treatments with fertilization (experimental treatment); and (3) results could be directly extracted from the text, tables, and/or figures. After careful evaluation, 44 publications representing field trials conducted at 65 sites (Argentina n = 5; USA n = 20; Brazil n = 40), were selected for this meta-analysis (Tab. S1 in Supplementary material).
General information on these research efforts (location, soil texture, fertilization type, tree age at fertilization, and nutrients applied), means (X), standard deviations (SD), and number of replications (n) were extracted to compose the values for assessed tree attributes (production of wood, litter, and roots; tree height and trunk diameter; and needle concentrations of N, P, K, Ca, Mg, and S). In studies that reported only the coefficient of variation (CV%), eqn. 1 was used to obtain SD values:
For studies that did not report data variability information, the mean SD was calculated for the control and experimental treatments using all data of each study. All data were extracted and compiled into a spreadsheet.
Data categorization
To evaluate the response of loblolly pine to fertilization, three groups of control factors were considered: fertilization factor; plant factor; and soil factor. Only control factors with more than ten paired comparisons were considered. For the fertilization factor, fertilizer type (mineral or residue), number of nutrients applied (one, two, or more than three), type of nutrients applied (macronutrients; macro- + micronutrients), and mineral fertilization (with or without lime application) were considered. Due to the low number of paired comparisons, it is worth noting that it was not possible to analyze liming in isolation, but only in association with fertilizer applications. Plant factors were categorized based on tree age at the time of nutrient application (< 1 yr, 2-8 yrs, and 9-16 yrs). Soil factors were based on clay content (< 15%, 15-30%, and 31-65%).
Data analysis
Magnitude of the fertilization effect was calculated using the natural logarithm of the response ratio (lnRR - eqn. 2) as the effect size ([23]):
where Xe and Xc are the mean values for the experimental and control treatments, respectively. Variance (v) was calculated as (eqn. 3):
where SDe, ne, SDc, and nc represented standard deviations and numbers of replications for experimental and control treatments, respectively. Response ratio variance was required to obtain balanced response ratio values and 95% confidence intervals (CI). Thus, the effect of fertilization was considered significant when the 95% CI of the response ratio did not overlap zero. Mean response ratio and CI values were generated using random-effects method with restricted maximum likelihood estimation. To facilitate interpretation of variations between experimental and control treatments, the response ratio and CI of the treatments were transformed (eqn. 4):
All analysis were performed using OpenMEE software ([44]).
Results
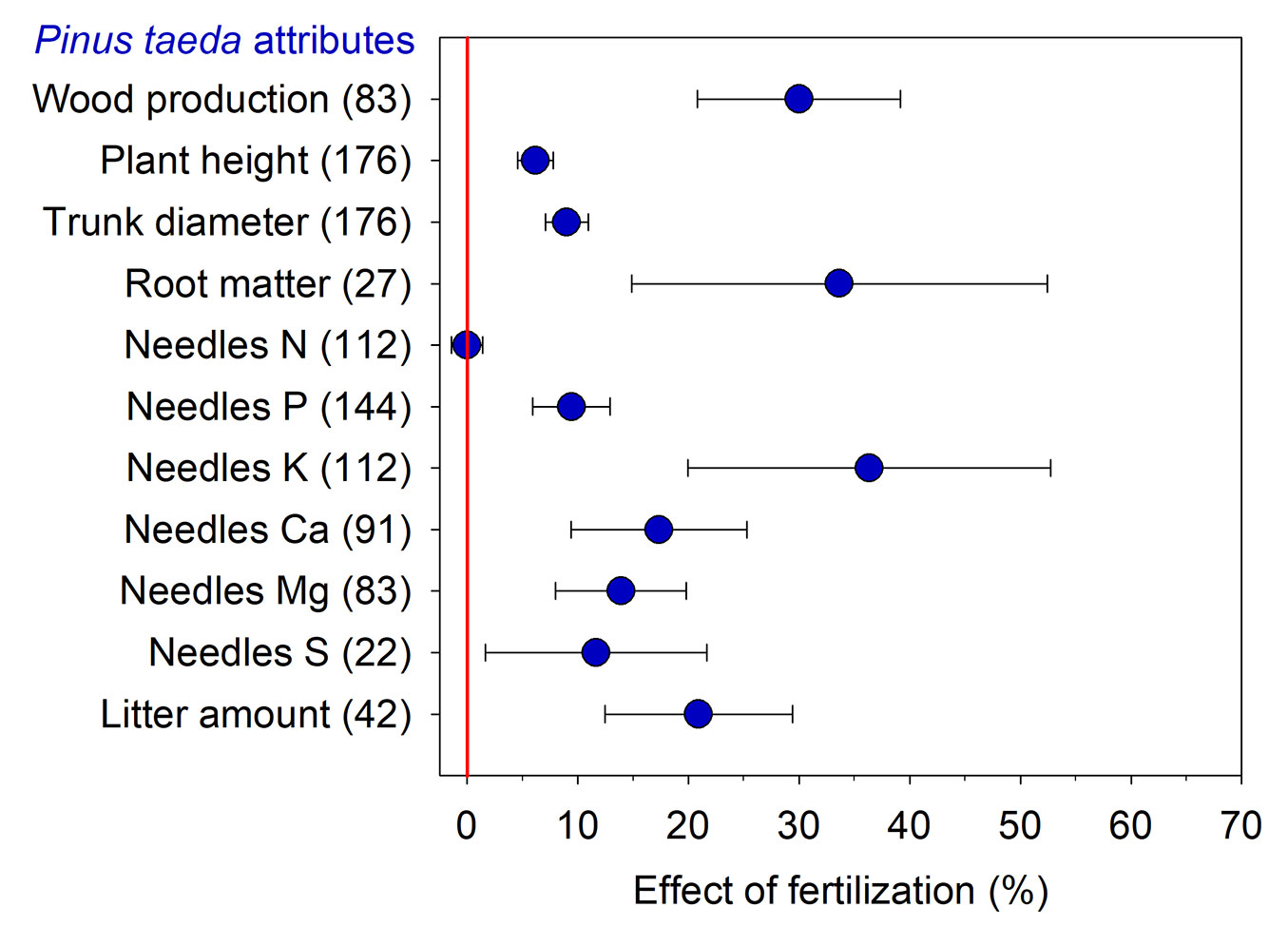
Fertilization provided a significant increase in several analyzed loblolly pine attributes, such as increases in wood production (+30%), root dry matter (+33%), and needle K concentration (+36% - Fig. 1). Fertilization increased plant height, trunk diameter, and litter amount by 6%, 9%, and 21%, respectively. Concentrations of other nutrients (i.e., P, Ca, Mg, and S) displayed increases between 9% and 17%, and only N concentrations were not affected.
Fig. 1 - Effect of fertilization on loblolly pine attributes compared to the absence of fertilization. Values are means ± 95% of the confidence interval (CI), and the number of comparisons used in the analysis of each attribute is presented in parentheses. Effect of fertilization is significant when the 95% CI response ratio value does not overlap zero.
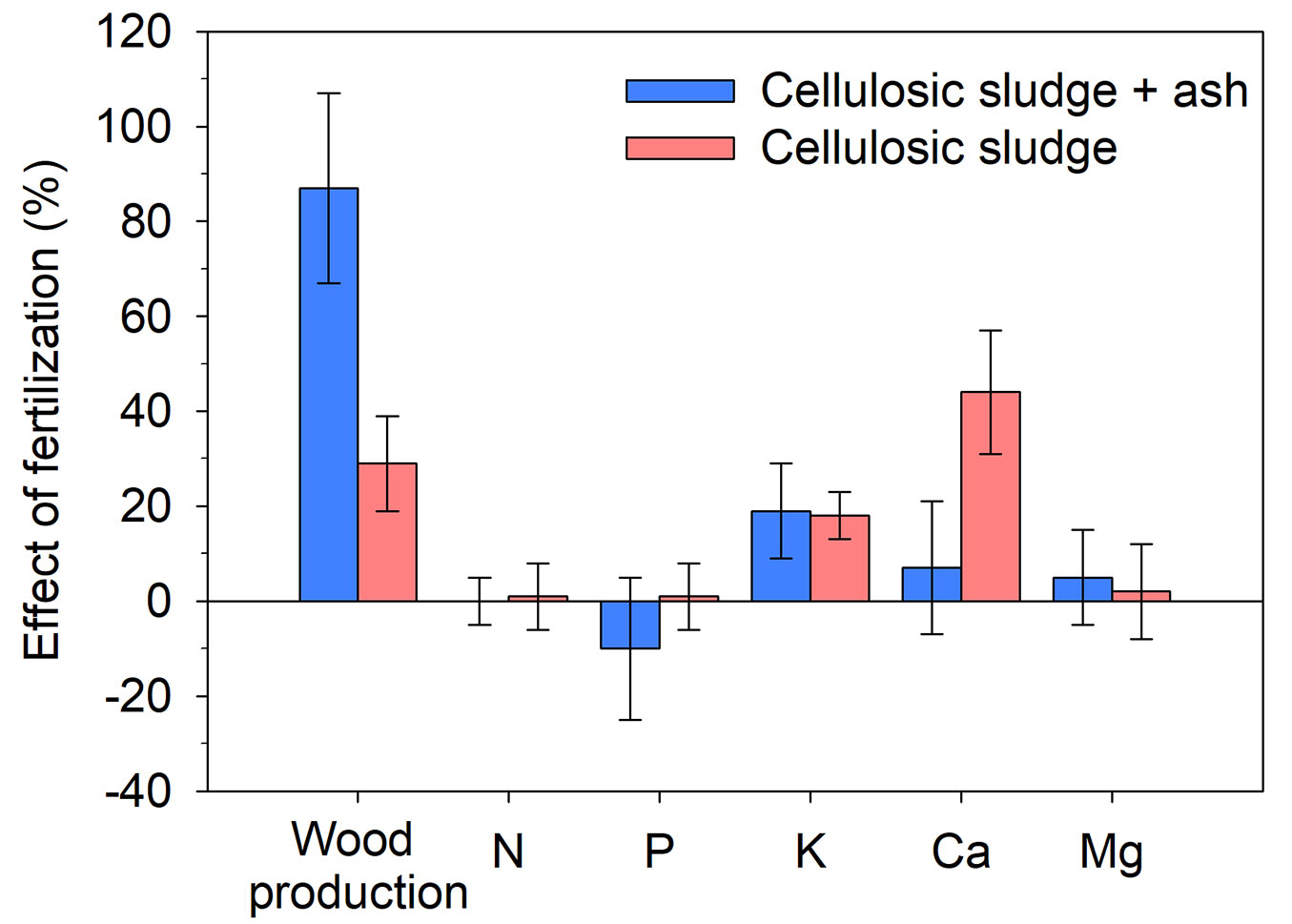
There was a direct relationship between increased loblolly pine wood production and fertilization, especially with residue applications (+66%) compared to mineral fertilization (+13% - Fig. 2). Analysis of residue type indicated that loblolly pine response was greater with applications of composite waste (cellulosic sludge + ash: +88%) compared to only applying cellulosic sludge (+29%), without differences in needle nutritional composition, except for higher Ca concentrations (+45%) when only cellulosic sludge was applied (Fig. 3). For mineral fertilization, wood production was greater with macro- and micronutrient applications (three or more nutrients applied) and when fertilization was combined with liming (Fig. 2).
Fig. 2 - Effect of fertilization on wood production and nutrient concentration in loblolly pine compared to the absence of fertilization. (N): nitrogen; (P): phosphorus; (K): potassium; (Ca): calcium; (Mg): magnesium. Values are means ± 95% of the confidence interval (CI), and the number of comparisons used in the analysis of each attribute is presented in parentheses. Effect of fertilization is significant when the 95% CI response ratio values does not overlap zero.
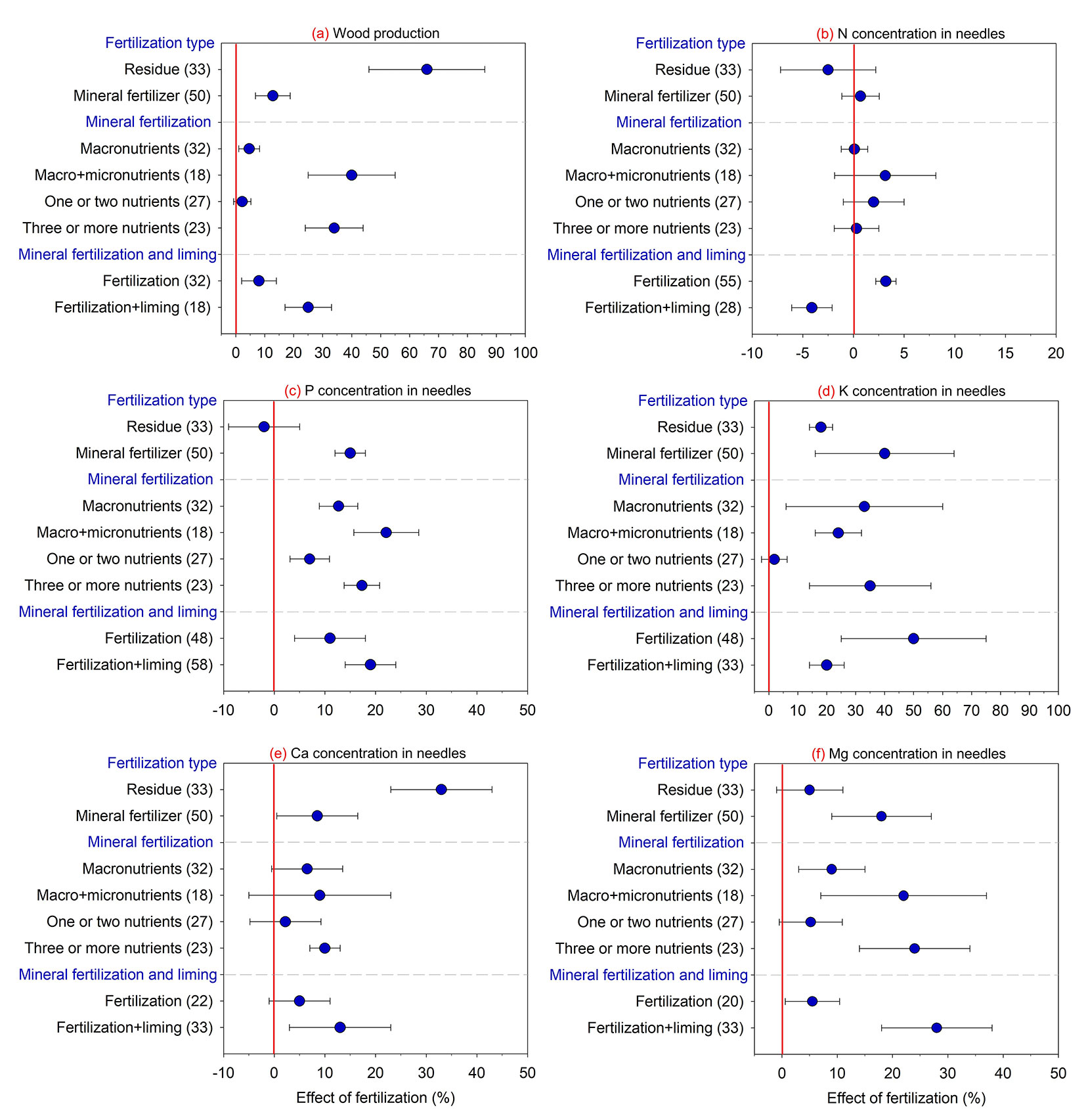
Fig. 3 - Effect of fertilization with different organic residues on wood production and nutrient concentration in loblolly pine compared to the absence of fertilization. (N): nitrogen; (P): phosphorus; (K): potassium; (Ca): calcium; (Mg): magnesium. Values are means ± 95% of the confidence interval (CI). Effect of fertilization is significant when the 95% CI response ratio values does not overlap zero.
Regarding needle nutrient composition, mineral fertilization increased concentrations of various nutrients (i.e., P, K, Ca, and Mg) from 10% to 40%, while fertilization with residue only increased K (+20%) and Ca concentrations (+34% - Fig. 2). Mineral fertilization with three or more nutrients was more efficient in increasing concentrations of P, K, Ca, and Mg in needles (increases between 10% and 35%). Application of macronutrients alone or in association with micronutrients increased needle P, K, and Mg concentrations. Likewise, fertilization alone or in association with liming increased needle P, K, and Mg concentrations. However, Ca (+12%) and Mg concentrations (+28%) were higher as a result of liming with the opposite being observed for N concentrations (-4%).
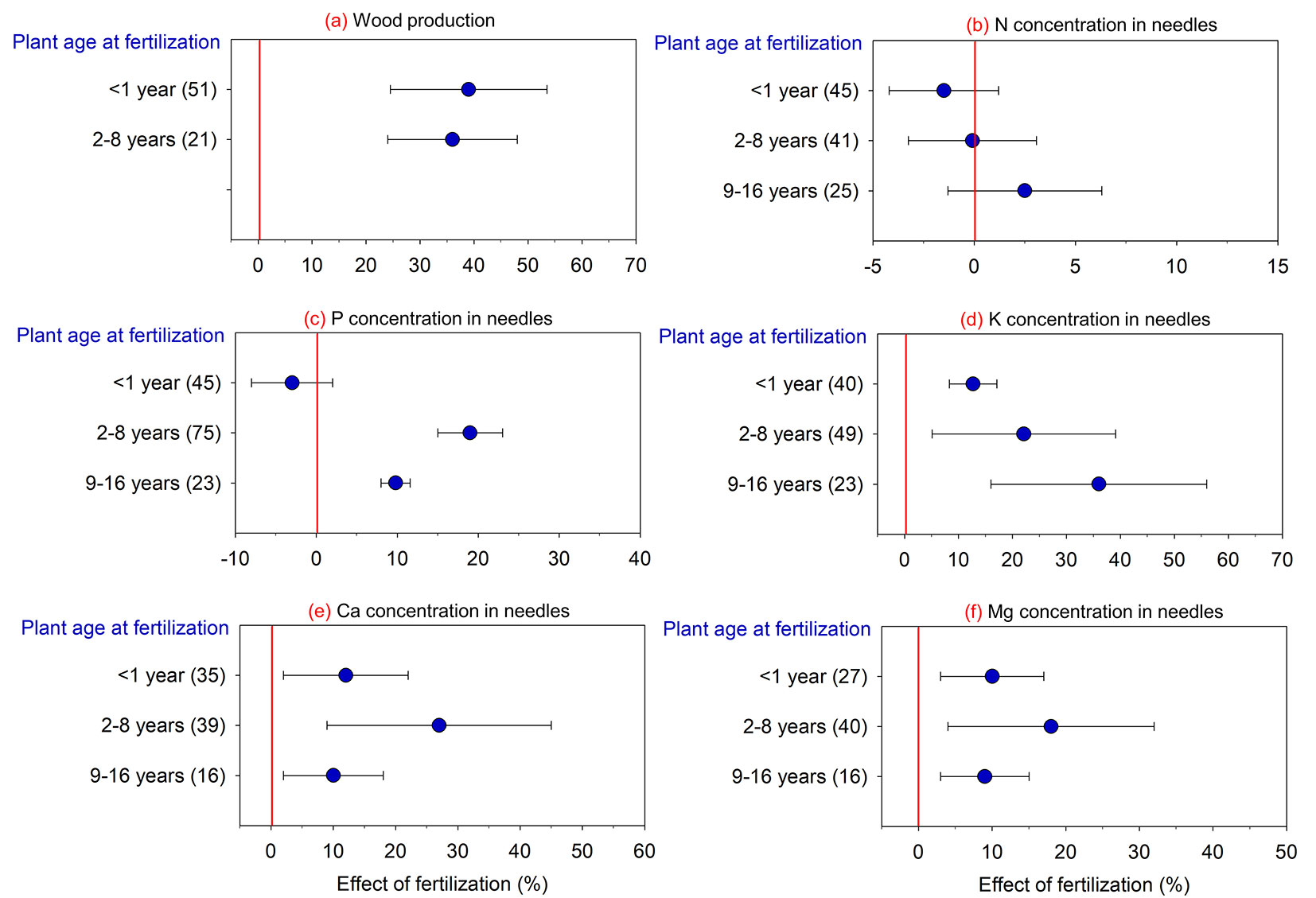
Increased loblolly pine wood production in response to fertilization occurred regardless of plant age at application since similar efficiency patterns were observed for trees less than one year-old up to trees 8 years-old (Fig. 4). On the other hand, this control factor had contrasting results based on the nutritional composition of loblolly pine needles: no effect for N; increases in P concentrations only when fertilization was performed on trees aged between 2-8 yrs (+19%) and 9-16 yrs (+10%); and increases in concentrations of other elements (i.e., K, Ca, and Mg) for all tree ages with fertilization, varying from 10% to 35%.
Fig. 4 - Effect of fertilization on wood production and nutrient concentration in loblolly pine compared to the absence of fertilization in response to plant age at fertilization. (N): nitrogen; (P): phosphorus; (K): potassium; (Ca): calcium; (Mg): magnesium. Values are means ± 95% of the confidence interval (CI), and the number of comparisons used in the analysis of each attribute is presented in parentheses. Effect of fertilization is significant when the 95% CI response ratio values does not overlap zero.
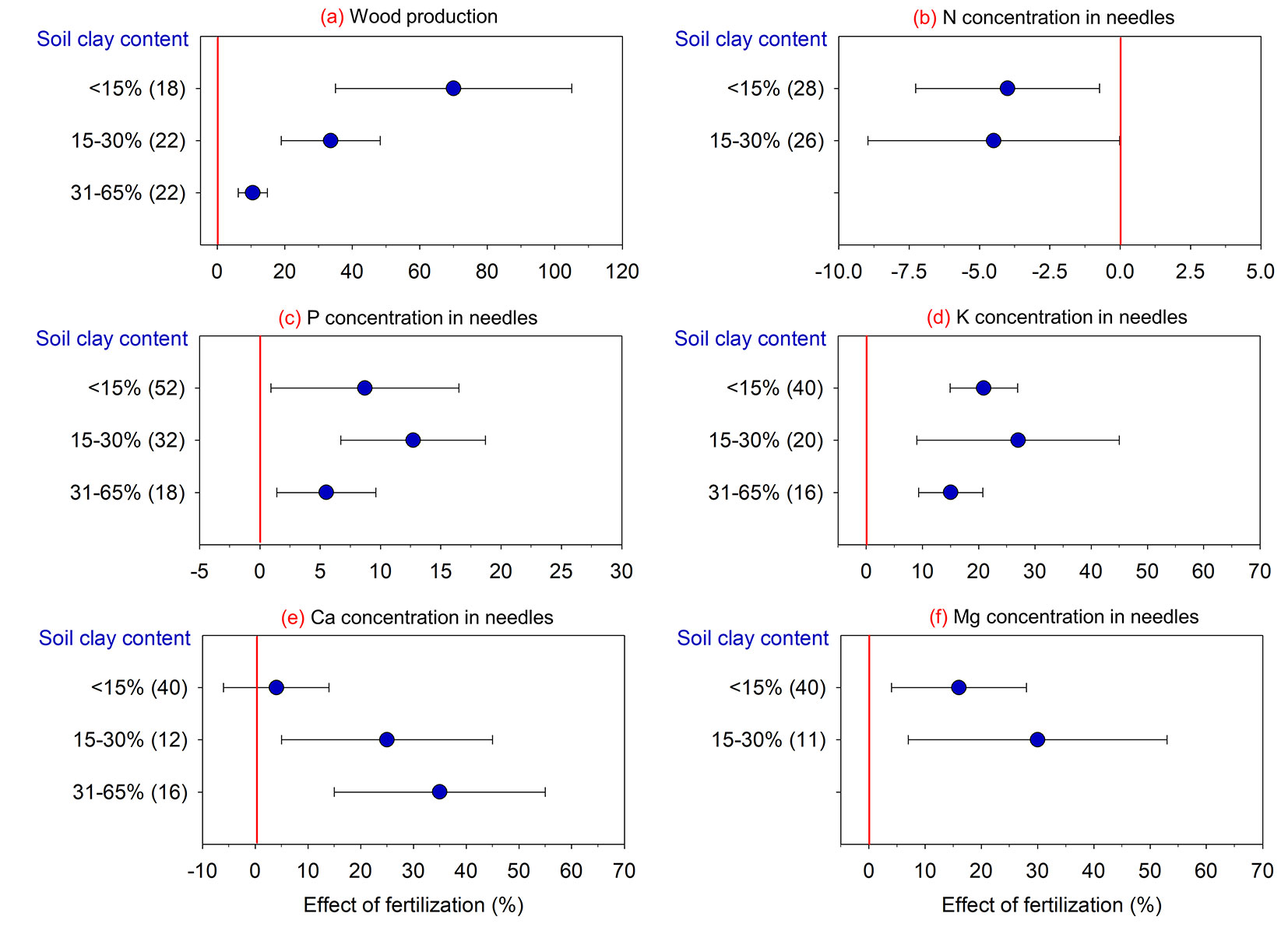
Soil granulometry was an important factor in understanding variations in fertilization efficiency on loblolly pine wood production (Fig. 5). Although fertilization increased wood production on soils with different clay values, the greatest tree response (+70%) occurred for sandy soils (clay contents < 15%) compared to more clayey soils, and were represented by wood production increases between 5% and 35%. Regarding nutritional composition of needles, fertilization decreased N concentrations and increased P, K, and Mg concentrations regardless of soil clay content. The only exception was Ca concentration in needles, which only increased (from 25% to 35%) for soils with clay contents between 15% and 65%.
Fig. 5 - Effect of fertilization on wood production and nutrient concentration in loblolly pine compared to the absence of fertilization in response to soil clay content. (N): nitrogen; (P): phosphorus; (K): potassium; (Ca): calcium; (Mg): magnesium. Values are means ± 95% of the confidence interval (CI), and the number of comparisons used in the analysis of each attribute is presented in parentheses. Effect of fertilization is significant when the 95% CI response ratio values does not overlap zero.
Discussion
General effect of fertilization
In intensively managed pine plantations, fertility practices targeting adequate nutrient levels in soil will allow for better productivity and increased wood production ([42], [17], [18]). Although companies in the forestry sector follow strict rules at various stages of cultivation, primarily aimed at certification of wood or derivatives for exports ([10]), adequate replacement of exported nutrients (from harvests) is often not considered ([31]). Thus, to avoid low tree growth and soil degradation from nutrient depletion, fertilization and liming could be important practices ensuring greater sustainability of loblolly pine production systems.
A significant increase in wood production of loblolly pine in response to fertilization, as revealed in the present study and supported by findings of multiple studies ([21], [14], [1], [15], [33]), occurs due to a series of effects related to the improvement of plant nutrition and belowground and aboveground growth. Fertilization favored enhanced root growth probably due to improved nutritional status ([8], [17]). Increases in pine root systems were reported as an effect of fertilization ([4]). In a 16-year-old forest, Albaugh et al. ([3]) found increases of 100% and 130 % in total aboveground and belowground biomass, respectively; this corroborates findings obtained in the present study. In addition to the direct effect of nutrients, the Ca/Al ratio of roots can also influence root growth in response to fertilizer and limestone use ([1]). Thus, a more vigorous loblolly pine root system translates into greater water and nutrient acquisition efficiencies, which along with higher nutrient supply via fertilization and liming, contributes to greater aboveground growth (in relation to height and trunk diameter) that favors wood production.
Litter accumulation may be associated with at least three factors: quantity of deposited material, quality of deposited material, and soil fertility at forest sites ([35], [31]). Our study revealed that higher litter deposition due to increased tree growth in response to fertilization and non-alteration of needle N (a primary nutrient influencing decomposition processes - [40]) may help explain litter maintenance. After evaluating needles samples from 110 studies of P. taeda regions in the USA, Albaugh et al. ([5]) ranked N as the least variable (Ca > K > Mg > P > S > N), suggesting low sensitivity to soil variation and possibly fertilization. Also, symptoms of N deficiency in Brazil are rare and restricted to areas having very shallow soils with growth occurring directly on the C horizon ([31]). However, response to N use has been observed under different conditions in the USA ([43], [14]). Since N acts as a great promoter of vegetative growth, it is likely that the needle response to N use is influenced by the dilution effect, which causes an apparent decrease in response. The decrease in N concentration in needle fall and increases in litter fall from P fertilization reported by Wienand & Stock ([45]) were used to explain decreased litter decomposition and litter accumulation in a P. elliotti system.
A significant increase in needle K may be associated with soil type in pine cultivations; these soils generally have low effective CEC and no K fixation ([14], [31], [13]), which decreases K adsorption capacity and allows for ready availability when applied. In relation to the plant, K is a nutrient required in high concentrations and associated with luxury consumption ([28]), which together favors greater K concentration increases in loblolly pine needles. In contrast to K, smaller increases in P concentration may be related to the specific adsorption of this nutrient to soil colloids, especially in very acidic soils ([31], [34]). Additionally, increases in P concentration were less frequent and to a lesser extent than K in Brazil ([18]) and USA sites ([43], [5]). Limestone use increased Ca and Mg concentrations; in comparison, these values were lower than K and higher than P. Commonly observed in pine systems, Mg deficiencies could promote tree responses to fertilization and/or liming ([20]). However, the smaller variation in needle Ca and Mg (compared to K concentration) can be explained by slow mobility from roots to needles since high adsorption within xylem tissue can act as a chromatographic column ([24]).
Effect of fertilization strategies
A consistent response to fertilization has been reported for mid-rotation applications ([21], [14], [15]), regardless of pine age in mid-term application. Although adequate response to fertilization at mid-rotations have been documented, early fertilization could be justified on sites displaying low initial growth. This may be the case for low fertility soils commonly used for pine plantations. In southern Brazil, ample nutrient reserves can be associated with very clayey soils derived from basalt parent material, while low nutrient reserves in sandstone derived soils ([32]) helps explain greater fertilization responses on sandy soils ([13], [41], [1], [17]). In the USA, large areas originating from marine sediments formed low fertility sandy soils that reflect a wide response to fertilization ([6]).
Greater tree response when more than one nutrient was supplied by mineral fertilization is due to synergistic effects reported for N and P ([2]) or NPK plus micronutrient additions ([14], [15]). Fertility experiments that add only one element (without addressing other required nutrients) can limit loblolly pine response on soils lacking multiple nutrients. Positive responses to fertilizer and lime use can be associated with the addition of three or more nutrients since lime can be a Ca and Mg source ([13], [1], [38]). The prominent lack of Mg observed in the USA ([3]), Brazil ([1], [38]), and worldwide ([20]) forest systems clearly suggests that liming can complement fertilizer applications.
Compared to mineral fertilization, the greater response to residue applications can be related to multi-element additions and higher rates of nutrients and organic compounds ([33]). A further consideration regarding organic residue use is the slow release of nutrients such as N, P, and S ([46]) and how this benefits soil biota, contributes to nutrient cycling, and reduces pathogenic actions ([19], [27]). Furthermore, it is interesting to note that some residues common in the cellulose industry (cellulose sludge, rich in Ca) can have a much more expressive effect on pine wood production by mixing with other residues that contain more nutrients, such as the mixture of cellulose sludge and ash ([41], [33]). The low-cost residue inputs represent a viable fertility option since the high cost of mineral fertilizer can be an impediment factor ([7]). Albaugh et al. ([6]) reported that increases in fertilizer consumption from 1969 to 1999 were followed by decreases in 2016, primarily due to higher costs. Thus, the use of residues in fertilization brings a series of economic and environmental advantages and should be used as a fertilization strategy in sites with pine plantations.
Regarding needle nutrient concentrations, mineral fertilization increased P, K, Ca, and Mg, reinforcing the idea that trees quickly absorb readily available nutrients from mineral fertilizer applications ([13], [17]), while residue use primarily increases K and Ca concentrations. When applying residues from the cellulose industry, increases in needle Ca were expected since the material composition contained high amounts of this nutrient ([39], [33]).
Although fertilization can generally lead to improved K concentration, results clearly showed that lime added with fertilizer can lead to decreased K concentration since other nutrients can compete for transporters and active absorption sites on roots. Excess Ca2+ and Mg2+ ions can lead to lower absorption of the K+ ion ([18]). Batista et al. ([13]) observed that Ca and Mg had reduced absorption in response to K fertilization; this antagonistic effect demonstrates the importance of calibrating soil correctives and fertilizer rates used in forest plantations.
Decreases or no major changes in N concentrations were verified in the present study, which is very different from that observed for the P, K, Ca, and Mg concentrations in loblolly pine needles. Even with N application, growth stimulation can lead to a dilution effect with lower levels of N in needles ([28]). Nitrogen is the nutrient most commonly limiting to loblolly pine growth and fertilization can increase availability, which increases leaf area/sunlight capture and tree growth ([21]), thereby lowering N concentrations.
Conclusion
This meta-analysis indicated that fertilization and liming increased loblolly pine wood production due to increases in plant height and trunk diameter. These production benefits were associated with increased root growth and improved nutritional status of trees. As a result of increased tree growth, there was also an increase in litter deposition that likely impacted forest nutrient cycling.
Although fertilization efficiency in loblolly pine was similar for applications at planting and on established sites of 2 to 8 years (especially for wood production), we verified slight variations in function of other controlling factors. In this regard, the most expressive tree response occurred with residue applications (vs. fertilization with mineral sources), with emphasis on residues with more balanced amounts of nutrients. When evaluating only mineral fertilization, multiple nutrients supplied by the joint application of fertilizer and lime was the most beneficial strategy for tree growth, possibly related to addressing lack of Ca and Mg. Regarding soil attributes, loblolly pine on sandy soils (<15% clay) was more responsive to fertilization. In summary, results obtained in this meta-analysis could be a useful guide for fertilization and liming practices in areas destined for reforestation with loblolly pine, aiming to guarantee greater sustainability of these production systems.
References
CrossRef | Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Antônio Carlos Vargas Motta 0000-0001-9117-1881
Tamires Maiara Ercole 0000-0003-4623-1042
Department of Soils and Agricultural Engineering, Federal University of Paraná, 1540 Funcionários St., Curitiba, PR, 80035-050 (Brazil)
Federal Institute of Southeast of Minas Gerais, 204 Monsenhor José Augusto St., Barbacena, MG, 36205-018 (Brazil)
USDA-ARS National Soil Dynamics Laboratory, 411 South Donahue Drive, Auburn, AL 36832 (USA)
Corresponding author
Paper Info
Citation
Rodrigues VDS, Motta ACV, Barbosa JZ, Ercole TM, Prior SA (2023). Wood production and nutritional status of Pinus taeda L. in response to fertilization and liming: a meta-analysis of the Americas. iForest 16: 195-201. - doi: 10.3832/ifor4296-016
Academic Editor
Daniela Baldantoni
Paper history
Received: Dec 28, 2022
Accepted: May 01, 2023
First online: Jul 25, 2023
Publication Date: Aug 31, 2023
Publication Time: 2.83 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2023
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 22335
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 16757
Abstract Page Views: 2966
PDF Downloads: 2155
Citation/Reference Downloads: 0
XML Downloads: 457
Web Metrics
Days since publication: 1065
Overall contacts: 22335
Avg. contacts per week: 146.80
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2023): 1
Average cites per year: 0.33
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Effect of seedling stock on the early stand development and physiology of improved loblolly pine (Pinus taeda L.) seedlings
vol. 9, pp. 690-695 (online: 12 May 2016)
Review Papers
Should the silviculture of Aleppo pine (Pinus halepensis Mill.) stands in northern Africa be oriented towards wood or seed and cone production? Diagnosis and current potentiality
vol. 12, pp. 297-305 (online: 27 May 2019)
Review Papers
Pathways to increase supply of sustainable wood from planted forests and trees to address bioeconomy needs: a review
vol. 19, pp. 168-185 (online: 12 May 2026)
Research Articles
Contrasted growth response of hybrid larch (Larix × marschlinsii), jack pine (Pinus banksiana) and white spruce (Picea glauca) to wood ash application in northwestern Quebec, Canada
vol. 14, pp. 155-165 (online: 06 April 2021)
Research Articles
Differences in early dynamics and effects of slope aspect between naturally regenerated and planted Pinus sylvestris woodland on inland dunes in Poland
vol. 9, pp. 875-882 (online: 17 June 2016)
Research Articles
Effects of planted European beech on the understory in Scots pine forests of Lithuania
vol. 7, pp. 12-18 (online: 07 October 2013)
Research Articles
Effect of family, crown position, number of winter buds, fresh weight and the length of needle on rooting ability of Pinus thunbergii Parl. cuttings
vol. 9, pp. 370-374 (online: 11 January 2016)
Research Articles
Study on the chemical composition of teak wood extracts in different organic solvents
vol. 14, pp. 329-336 (online: 09 July 2021)
Research Articles
Wood-soil interactions in soil bioengineering slope stabilization works
vol. 2, pp. 187-191 (online: 15 October 2009)
Research Articles
Impact of rotation length of Eucalyptus globulus Labill. on wood production, kraft pulping, and forest value
vol. 15, pp. 433-443 (online: 20 October 2022)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword






