
Variation of wood and bark density and production in coppiced Eucalyptus globulus trees in a second rotation
iForest - Biogeosciences and Forestry, Volume 9, Issue 2, Pages 270-275 (2015)
doi: https://doi.org/10.3832/ifor1442-008
Published: Sep 08, 2015 - Copyright © 2015 SISEF
Research Articles
Abstract
Coppiced Eucalyptus globulus trees with 18 years of age and in a second rotation were analysed in relation to wood and bark density in a spacing trial with five initial plant densities. A total of 25 stumps, with a variable number of stems per stump, from one to three, were analysed. A comparison was made to the previous first rotation single stem trees, also harvested at 18 years of age. The average wood basic density at breast height of the eucalypt coppiced trees was 567 kg m-3, with lower wood density for the closest spacings. The wood of coppiced Eucalyptus globulus trees was 2.5% less dense than that of the single stem trees in the first rotation (average 582 kg m-3). Within the tree, the wood density decreased from stump level to breast height level and then gradually increased until 11.3 m and then decreased slightly. The bark density was, on average, 473 kg m-3, ranging from 455 to 487 kg m-3. The mean bark density was comparable to the bark density in the first rotation. The average bark content was 17.4% of the stem volume, providing 25 to 52 ton ha-1. Compared to the first rotation, the average tree volume in the coppice was lower because the individual trees were smaller by 40 to 66%, while the estimated volume production per ha of the coppice was 1.1 to 1.8 times more due to the increased number of trees that were left in each stump.
Keywords
Eucalyptus globulus, Wood Density, Plant Density, Coppice, 1st Rotation
Introduction
Wood density is an important property when assessing the raw material quality for pulping since it affects pulp yield and other aspects of the pulp and papermaking processes, as well as the economics of forest production and mill processing. Most breeding programmes have therefore taken wood density as one of the determining selection criteria. This has been the case of eucalypts used for the pulp industry, for which the breeding traits have included wood density in addition to those related to tree growth ([3], [9], [23], [38]).
Eucalyptus globulus is extensively cultivated for the pulp and paper industry in temperate regions due to its fast growth, good stem characteristics and exceptional pulping quality ([25]). In Europe, E. globulus plantations exist mainly in Portugal and Spain, where they cover approximately 1.3 million ha. Portugal was the first country to have pulp mills that were based on E. globulus plantations, in addition to developing a dedicated eucalypt silviculture ([1]).
The plantations are managed as a coppice system, with a first cycle of single-stem trees (first rotation) followed by two or three coppice cycles until the final harvest, stump removal and replanting. The species regenerates easily through stump coppice after harvesting. Since the coppice regrowth benefits from the established root system, the coppice rotations without replanting are advantageous, provided that the first rotation was well established and the stump regrowth was sufficient ([12], [15]). In the usual practice, a selective thinning of the sprouts is carried out in the beginning of the coppice cycle to maintain or increase the stand density up to about 20% ([39], [40], [37]).
The wood quality of E. globulus single stem trees has been extensively studied, including wood density and its variation within the tree or site, as compiled in Pereira et al. ([25]). At the age of harvest for the pulp industry in the temperate regions (10-14 years), E. globulus wood basic density varies in the range 470-650 kg m-3 and selection for improved dry weight production is therefore made in on-going improvement programmes ([2], [41], [25]). Most pulp mills adopt 650 kg m-3 as the upper limit for the eucalypt wood density to avoid liquor impregnation problems and pulping heterogeneity.
However, there is very little information on the quality of coppiced wood and the differences in relation to the previous single-stem rotation. Information on the influence of silvicultural practices on the technological quality of coppiced trees is also particularly sparse. Sharma et al. ([35]) investigated non-coppiced and coppiced wood of E. tereticornis and concluded that the wood quality of both is comparable, which means they can be used for the same purposes. However, as regards wood density, the few studies available report a decrease of wood density in the coppiced trees. Schonau ([33]) found that wood density of coppiced trees of E. grandis was lower than that of their parent trees, and Sesbou & Nepveu ([34]) confirmed that the first cutting decreased the basic density in E. camaldulensis. Zbonak et al. ([42]) found the same in clones of E. grandis, E. grandis x E. urophylla and E. grandis x E. camaldulensis, for which the wood density of the parent trees was higher than that of the coppiced trees. Zobel & Van Buijtenen ([43]) reported that the properties of coppiced wood were different from those of the original trees, namely they had a lower density.
The objective of the present study is to provide insight on the stem quality of coppiced E. globulus trees in a second rotation regarding the wood density and its variation within the tree, as well as of bark density, and to analyse the effect of the initial planting density (i.e., stump density) by using a spacing trial for the study. It is also an objective to compare data on wood density, tree volume and stand productivity of the coppice with the first single stem previous rotation, since this data is available from previous studies ([20], [21]). This will allow for estimating changes between the two cutting cycles and therefore better adapt the silvicultural management.
Material and methods
Field sampling
Eucalyptus globulus Labill. trees were harvested at 18 years of age, at the end of the second rotation from a spacing trial located in the site of Alto do Vilão, Furadouro (centre of Portugal, 10 km from the Atlantic Ocean - 39° 20′ N, 9° 15′ W, 50 m a.s.l.). The climate is of the Mediterranean type tempered by oceanic influence, with an annual rainfall of 608 mm and a mean temperature 15.2 °C. The predominant soil type is a sandy podzol associated with eutric cambisols.
The pulping company CELBI (now ALTRI) established the Alto do Vilão trial with commercial seeds in March 1975. The trial includes two blocks, each with five plots with different tree spacings (m×m): 3×2 (1667 trees ha-1), 3×3 (1111 trees ha-1), 4×3 (833 trees ha-1), 4×4 (625 trees ha-1) and 5×4 (500 trees ha-1). The blocks are located side by side and the spacings are distributed according to decreasing density in one of the blocks and the opposite in the other block.
The trial was harvested in March 1993, at 18 years of age, as the first rotation, and the stumps were left to sprout for a second rotation cycle. The thinning of the shoots took place in October 1995, 2.6 years after harvest, following the usual practice of leaving shoots with dimensions comparable to those of the first rotation of the same age ([37]). This led to a variable number of shoots per stump. The end of the second rotation was in 2008, at 18 years of age, when the trial was harvested. Before harvest, the diameter at breast height (dbh) and the total tree height were measured.
Sampling was conducted in a total of 25 stumps by randomly selecting five stumps in each spacing plot. As described, the stumps had a variable number of stems; seven stumps with three stems/stump, six stumps with two stems/stump, and 12 stumps with one stem/stump. From each sampled tree, 10 cm thick discs were cut at different stem heights: base, 1.30 m and every 2 m along the stem until the top, corresponding to a 7 cm diameter.
Measurements
The wood and bark were separated. Estimates of basic density were obtained using the water immersion method by determining the green saturated volume and the oven-dry weight (TAPPI Test Method - T258 om 98, TAPPI Press, Atlanta, GA, USA). The influence of the initial planting density and stump density on wood density at breast height was studied using a one-way ANOVA analysis. Statistical analysis was conducted using the software package SJHNASUAU© for Windows Version 2.0 (Jandel Corporation, San Jose, CA, USA), with α = 0.05.
Comparison with first rotation data
The trees harvested in the second cutting had been also measured at the end of the previous first cutting, therefore allowing for direct comparison between the two rotation cycles, since the trees were 18 year old in both cutting cycles. At the harvest of the first rotation of this trial, the overbark diameter at 1.3 m above the ground and the tree height were determined and the wood basic density was measured at dbh ([20]). These values were compared to those obtained for the same stumps in the second rotation.
For comparison of tree and stand volume production between the two cutting cycles, the individual tree total volume was calculated for the first and second rotations using the volume equation model developed for E. globulus in Portugal by Tomé et al. ([40]) based on dbh and total tree height (eqn. 1):
Bark volume was calculated as the difference between the tree and stemwood volumes.
Results and discussion
Wood basic density
Tab. 1 gives the mean values of wood basic density at dbh for the Eucalyptus globulus trees in the second cutting cycle, as coppiced stems from the five stumps per spacing, in addition to the same data published for the first rotation ([20]).
Tab. 1 - Variation of wood and bark basic density at breast height and bark content in percentage of stem volume in the 18-year-old coppiced E. globulus trees grown in the different spacing plots, as well as data for the first rotation (published by [20]). Mean of all coppiced trees per spacing and standard deviation.
| Rotation | Variable | Spacing (m × m) | ||||
|---|---|---|---|---|---|---|
| 4×5 | 4×4 | 4×3 | 3×3 | 3×2 | ||
| 2nd | Wood density (kgm-3) | 582 ± 26 | 597 ± 37 | 574 ± 32 | 557 ± 24 | 553 ± 38 |
| Bark density (kgm-3) | 456 ± 88 | 487 ± 21 | 483 ± 44 | 482 ± 25 | 455 ± 34 | |
| Bark content (% of stem volume) | 17.4 ± 0.1 | 17.4 ± 0.1 | 17.4 ± 0.1 | 17.4 ± 0.2 | 17.4 ± 0.1 | |
| 1st | Wood density (kgm-3) | 594 ± 43 | 583 ± 44 | 590 ± 40 | 580 ± 34 | 563 ± 42 |
| Bark content (% of stem volume) | 14.4 ± 0.5 | 13.4 ± 2.5 | 14.0 ± 0.7 | 13.8 ± 0.7 | 13.4 ± 1.1 | |
The average wood basic density at breast height for all the eucalypt coppiced trees was 567 kg m-3, ranging from 553 to 597 kg m-3 among spacings. For the same trees at 18 years of age in the first rotation, the range of wood density was between 563 and 594 kg m-3, with an average of 582 kg m-3 ([20]). These values are within the range (425-668 kg m-3) reported for the wood basic density of E. globulus trees at harvest age for pulping of 9-15 years in the first rotation ([24], [18], [19], [26], [30], [4], [11], [38]).
The differences in wood density between the coppiced and the single stem trees were of a small magnitude. On average, the wood from coppiced trees was 2.5% less dense than that of the single stem trees. The difference was statistically significant (P=0.039) only for the 3×3 m spacing (Fig. 1).
Fig. 1 - Average wood density of parent and coppiced trees for the different spacings. Bars represent the standard deviation.
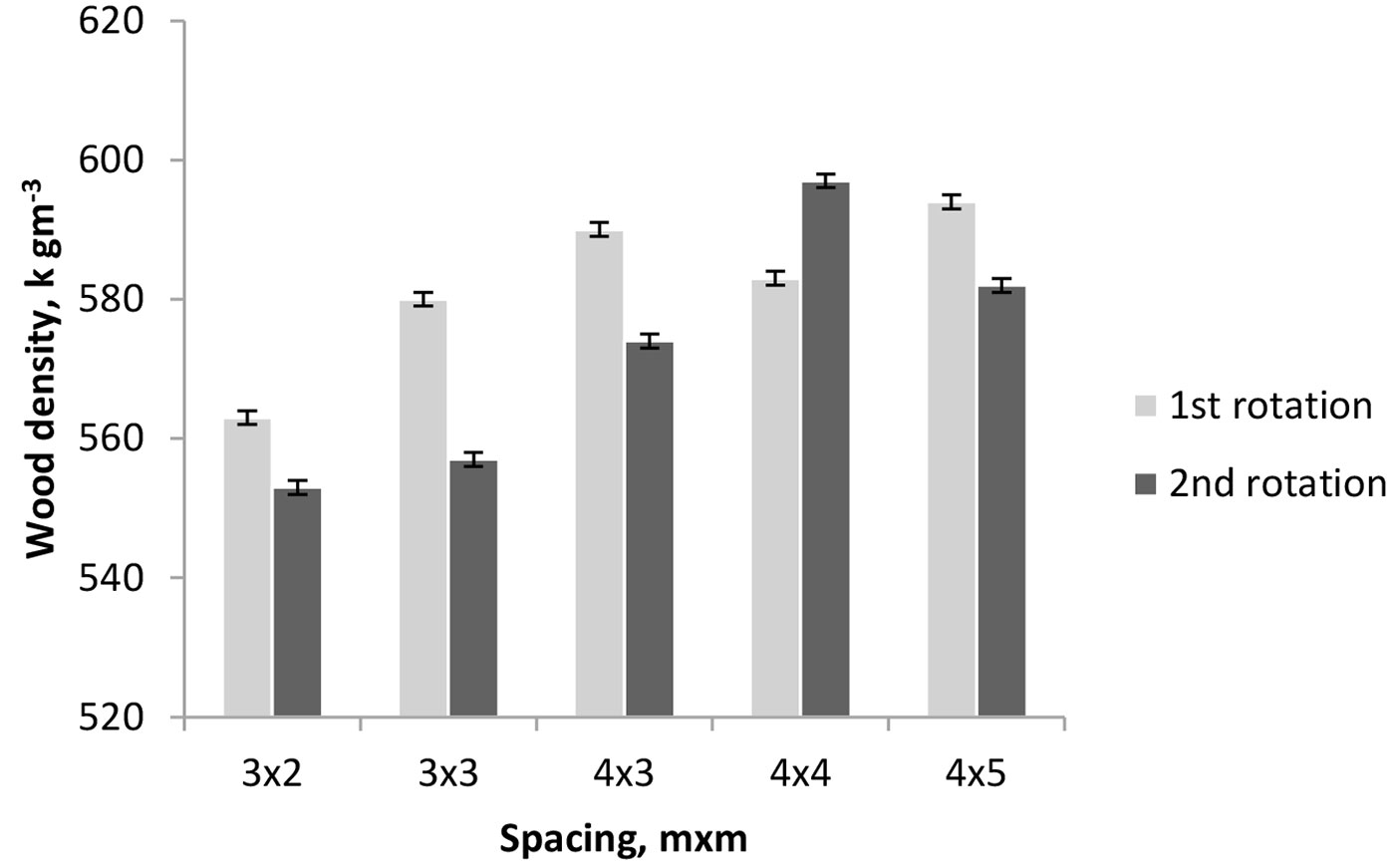
A few studies on Eucalyptus species also reported very small differences in wood density between the seed- and sprout-originated trees. A decrease in basic density was reported in E. tereticornis coppice (696 kg m-3 for coppice and 710 kg m-3 for non-coppiced trees - [35]). Sesbou & Nepveu ([34]) found that the basic density of coppice material in E. camaldulensis was 5% less dense than that of the first rotation material (502 and 527 kg m-3, respectively). Ferrari ([8]) compared coppice shoots with first rotation samples of E. bicostata, E. globulus and E. viminalis, and found that eucalypt coppice wood was 8% less dense.
An effect of stump density on the wood basic density was found (P = 0.036), though the differences were statistically significant only between the widest (4×4) and the closest (3×3 and 3×2) spacings. The between-tree variability in each spacing was small, with coefficients of variance of the means below 5% in all cases. However, the stand density did not significantly influence wood density in the first rotation ([20]).
Wood density did not show a correlation with growth rate, as measured by tree diameter (R² = 0.0154, P = 0.433), confirming previous reports that growth rate is not correlated with wood density in E. globulus ([18], [19], [26]). Raymond & Muneri ([29]) also reported this lack of relationship between growth rate and wood density for E. globulus.
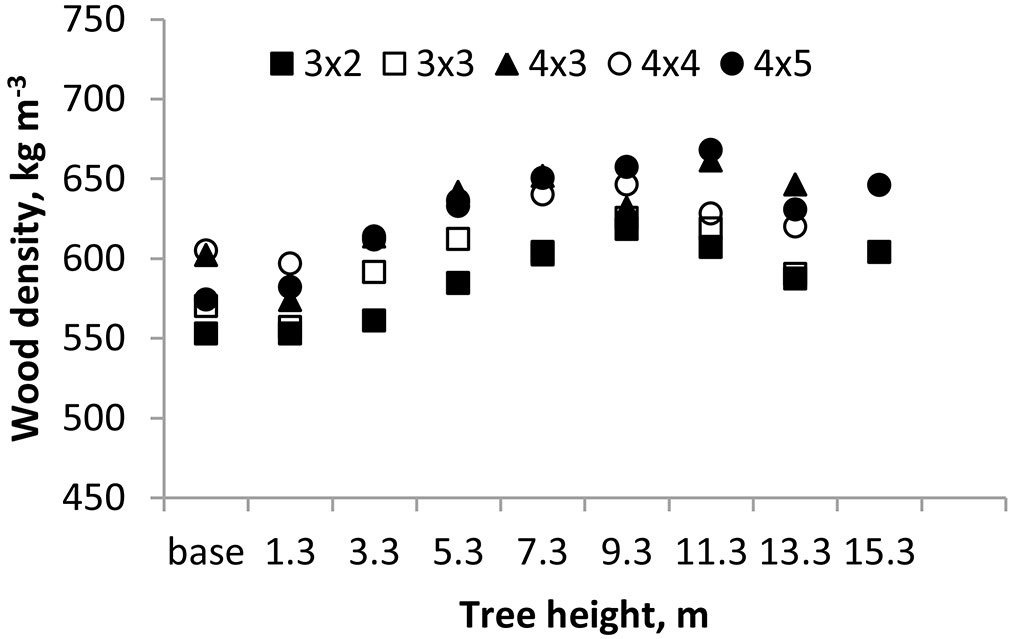
The axial variation of the wood basic density showed in all cases a similar profile in the coppiced trees, increasing along the stem from 581 to 636 kg m-3, respectively at the base and top (Fig. 2). Initially the wood density declined from stump level to breast height level and then gradually increased until 11.3 m and then slightly decreased. The vertical pattern of basic density variation was similar across all spacings. For eucalypts, the predominant trend is a reduction in density from the base of the stem to 10% of the total tree height, followed by a progressive increase in density, as has been described for E. globulus ([6], [31], [5], [26], [30], [13], [10], [4]).
Fig. 2 - Axial variation of wood basic density of 18-year-old E. globulus coppiced trees. Mean of all coppiced trees per spacing.
Bark content and density
The bark content in 18-year-old E. globulus coppiced trees represented, on average, 17.4% of the total stem volume, while for the first rotation at the same age, the average bark content was somewhat smaller, at 13.8% (Tab. 2). Both values are consistent with the few available studies: 15.2% of bark content for 7- and 11-year-old E. globulus trees ([28], [22]), values between 13.6% and 15.9% in 4-year-old E. grandis ([32]), and 20.2% for E. urophylla at seven years of age ([14]).
Tab. 2 - Biometric data for Eucalyptus globulus trees grown with different planting densities in the second cutting cycle as coppiced trees, as well as data published for the first rotation ([37]). The values refer to five stumps per spacing. Mean values and standard deviation are reported.
| Rotation | Variable | Spacing (m × m) | ||||
|---|---|---|---|---|---|---|
| 4×5 | 4×4 | 4×3 | 3×3 | 3×2 | ||
| 2nd | Tree volume (m3) | 0.30 ± 0.07 | 0.20 ± 0.08 | 0.30 ± 0.15 | 0.25 ± 0.17 | 0.35 ± 0.13 |
| Volume per stump (m3) | 0.64 ± 0.30 | 0.48 ± 0.39 | 0.55 ± 0.34 | 0.38 ± 0.30 | 0.40± 0.16 | |
| Volume over bark (m3 ha-1) | 320.0 | 300.0 | 458.2 | 422.2 | 666.8 | |
| Bark content (% of stem volume) | 17.4 ± 0.1 | 17.4 ± 0.1 | 17.4 ± 0.1 | 17.4 ± 0.2 | 17.4 ± 0.1 | |
| Volume of bark (m3 ha-1) | 55.7 | 52.2 | 79.7 | 73.5 | 116.0 | |
| Wood production (ton ha-1) | 155 | 149 | 218 | 195 | 306 | |
| Bark production (ton ha-1) | 25 | 25 | 38 | 35 | 52 | |
| Stem bark content (% weight) | 13.63 | 14.19 | 14.64 | 15.06 | 14.32 | |
| 1st | Tree volume (m3) | 0.67 ± 0.32 | 0.51 ± 0.26 | 0.45 ± 0.19 | 0.38 ± 0.20 | 0.32 ± 0.11 |
| Volume over bark (m3 ha-1) | 292.2 | 284.6 | 321.7 | 341.6 | 378.9 | |
| Bark content (% of stem volume) | 14.4 ± 0.5 | 13.4 ± 2.5 | 14.0 ± 0.7 | 13.8 ± 0.7 | 13.4 ± 1.1 | |
| Volume of bark (m3 ha-1) | 48.2 | 45.9 | 54.0 | 60.8 | 76.8 | |
The bark density at breast height was, on average, 473 kg m-3, ranging from 455 to 487 kg m-3 among different spacings (Tab. 1). Bark density at breast height did not show significant differences among spacings (P=0.269). No correlation was found between radial growth and bark density (R2 =0.019, P=0.394).
Fig. 3 represents the axial variation of bark density in the five spacings. Bark density was highest at the stump level (488 kg m-3), decreasing along the stem up to 3.3 m (462 kg m-3), after which it remained rather constant, at an average of 465 kg m-3. Within each spacing, no significant differences in bark density were found (P = 0.269).
Fig. 3 - Axial variation of bark basic density of 18-year-old E. globulus coppiced trees.
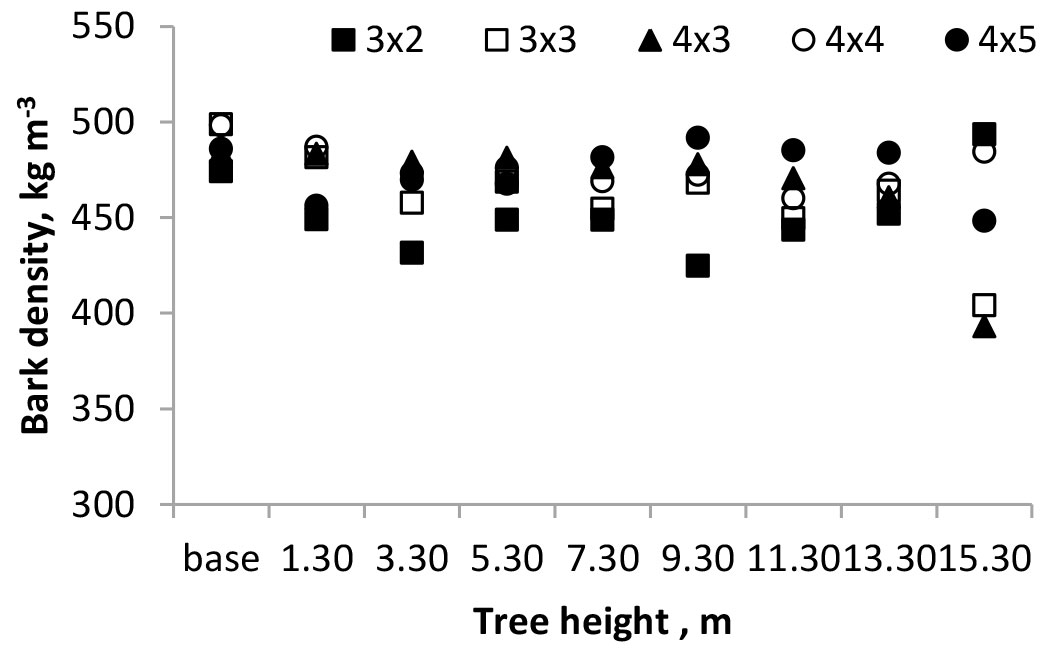
Very few data exist on E. globulus bark density. Quilhó & Pereira ([26]) reported a range from 302 to 454 kg m-3 in 12-year-old E. globulus trees. Macfarlane & Adams ([16]), for 6-year-old E. globulus from two sites, found values of 397 and 401 kg m-3, and Farrington et al. ([7]) reported 370 kg m-3 for 2- to 6-year-old E. globulus trees. It is expectable that bark density will increase with tree age, as the present study shows, due to the evolution of bark anatomy with age, e.g., the increase of sclerified cells and collapsed phloem ([27]).
Wood and bark production
Tab. 2 summarizes the production data for the E. globulus trees in the second cutting cycle, as coppiced stems from the five stumps per spacing, for individual tree volume, volume per stump and production values per hectare in volume and dry weight of wood and bark. As referred above, the stumps had a variable number of trees, from one to three, thus the mean volume per stump was derived from the sum of its individual stems. A comparison of volume production with the first rotation is shown in Fig. 4 using a 2nd:1st rotation volume ratio.
Fig. 4 - Ratio of volume production (m3 ha-1) between the 2nd and 1st rotations of E. globulus for five different spacings.
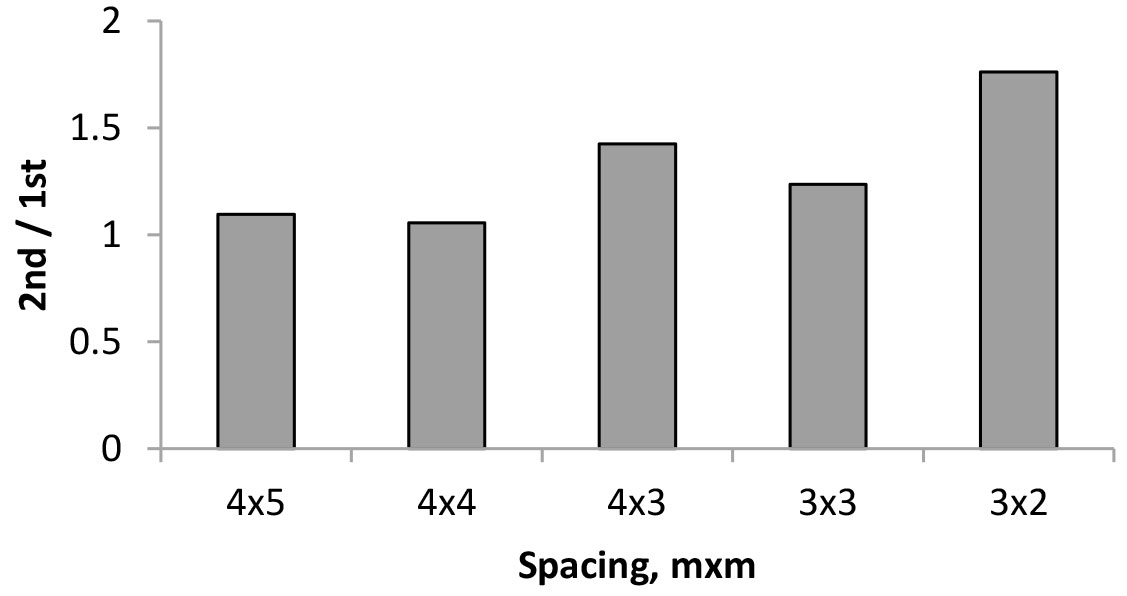
The average tree volume in the coppice was lower than in the first rotation because the individual trees were smaller by about 40 to 66% (Tab. 2). However, when the comparison is made at the stump level, the tree volume obtained per stump was similar to the volume obtained by the single parent tree. At the stand level, considering the estimated volume production per ha, the coppice crops produced 1.1 to 1.8 times more than the original parent trees (Fig. 4) due to the increased number of coppiced trees in each spacing.
Stand density was an important factor in stand productivity during the first rotation. In the closest spacing of 3×2 m, the trees were smaller and increased regularly with the widening of the spacing, until the 4×5 m spacing, respectively 0.32 m3 and 0.67 m3. However, the tree dimensions did not compensate for the difference in the number of trees in the different spacings, and the volume production per ha was highest in the 3×2 spacing (666.8 m3 ha-1) and lowest in the 4×4 (300 m3 ha-1) and 4×5 (320 m3 ha-1) spacings.
On the contrary, the spacing effect in the second rotation was not observed at tree level. This shows that a coppiced stand somewhat loses the effect of the initial planting density and has a more irregular space occupation by the tree crowns, probably in relation to the specific local situation, i.e., the number of stems per stump. In fact, the tree stand density changed between the two rotations due to the variable number of sprouts that were left in each stump. The between-stump variation was lower for the closest 3×2 spacing, while all other spacings had a more irregular tree distribution (data not shown).
In coppiced eucalypt plantations, it is usually assumed that peak production occurs in the second rotation, as a result of the favorable effect of a fully developed root system, and explaining the faster initial growth of shoots as compared to that of seedlings ([17]). Sims et al. ([36]) also found that total biomass yields of the coppice harvest were significantly higher compared to the single stem harvest of 19 Eucalyptus species after six years of growth. The present study shows that this is certainly true, and for the initial spacing of 3×2 m, there was an increase of production of 76% as compared with the first rotation (Fig. 3). For the 4×3 spacing, the production increase was 42%. However, for the other spacings, the production increase was not so high (e.g., by 9.5% for the 4×5 m spacing) and certainly depended on the number of sprouts left in each stump, as already discussed. This highlights the importance of the sprout thinning operation.
The average stem bark content, in percentage of stem volume, was 17.4%, providing 25 to 52 ton ha-1 of bark respectively for the widest (4×5 m) and closest spacing (3×2 m). Macfarlane & Adams ([16]) reported, for 6-year-old E. glubulus trees grown in two sites with planting density of 1250 trees ha-1, bark volumes of 17.9 and 10.1 m3 ha-1, corresponding to 7.0 and 4.0 ton ha-1 of bark, respectively. The volume of bark was always higher in the second rotation, especially for the 3×2 m spacing, with a bark volume of 116 m3 ha-1.
Conclusions
The wood of coppiced Eucalyptus globulus trees in the second cutting cycle was 2.5% less dense than that of the single stem trees in the first rotation. The between-tree variation was low and the within-tree pattern of axial variation was similar for all trees, with an increase from base to top. The wood density was not influenced by tree growth, but it was lower in the trees of the closest spacing. The bark density from Eucalyptus globulus trees in the second cutting cycle was similar to that of the single stem trees in the first rotation. The stand production of the coppice, in relation to the single stem, depends on the number of sprouts left on the stump, and therefore on the sprout thinning operation. In all cases, the stand production of bark was higher for the coppice.
Overall, the differences between the first and second cutting cycles refer to individual tree volume and stand productivity, which are largely dependent on the stand density resulting from the number of sprouts left per stump. The silvicultural operation of coppice thinning should take this into account.
Acknowledgements
The work was carried out with the base funding to Centro de Estudos Florestais given by Fundação para a Ciência e Tecnologia, Portugal, under the PestOE/AGR/UI02 39/2014 programme. The authors acknowledge the support of the pulp company CELBI (now ALTRI) for the supply of samples and for stand and tree data.
References
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Helena Pereira
Centro de Estudos Florestais - Instituto Superior de Agronomia, Universidade de Lisboa - Tapada da Ajuda 1349-017 - Lisboa (Portugal)
Corresponding author
Paper Info
Citation
Miranda I, Pereira H (2015). Variation of wood and bark density and production in coppiced Eucalyptus globulus trees in a second rotation. iForest 9: 270-275. - doi: 10.3832/ifor1442-008
Academic Editor
Gianfranco Minotta
Paper history
Received: Sep 09, 2014
Accepted: May 06, 2015
First online: Sep 08, 2015
Publication Date: Apr 26, 2016
Publication Time: 4.17 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2015
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 62926
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 48783
Abstract Page Views: 6538
PDF Downloads: 5936
Citation/Reference Downloads: 26
XML Downloads: 1643
Web Metrics
Days since publication: 3988
Overall contacts: 62926
Avg. contacts per week: 110.45
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Jul 2026)
Total number of cites (since 2016): 14
Average cites per year: 1.27
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Impact of rotation length of Eucalyptus globulus Labill. on wood production, kraft pulping, and forest value
vol. 15, pp. 433-443 (online: 20 October 2022)
Research Articles
Validation of models using near-infrared spectroscopy to estimate basic density and chemical composition of Eucalyptus wood
vol. 17, pp. 338-345 (online: 03 November 2024)
Research Articles
Interaction between planting spacing and wood properties of Eucalyptus clones grown in short rotation
vol. 14, pp. 12-17 (online: 02 January 2021)
Short Communications
Effect of intensive planting density on tree growth, wood density and fiber properties of maple (Acer velutinum Boiss.)
vol. 9, pp. 325-329 (online: 22 October 2015)
Research Articles
Effect of stand density on longitudinal variation of wood and bark growth in fast-growing Eucalyptus plantations
vol. 12, pp. 527-532 (online: 12 December 2019)
Research Articles
NIR-based models for estimating selected physical and chemical wood properties from fast-growing plantations
vol. 15, pp. 372-380 (online: 05 October 2022)
Research Articles
Earlywood vessel features in Quercus faginea: relationship between ring width and wood density at two sites in Portugal
vol. 8, pp. 866-873 (online: 27 April 2015)
Technical Advances
Technical properties of beech wood from aged coppices in central Italy
vol. 8, pp. 82-88 (online: 04 June 2014)
Research Articles
Physical, chemical and mechanical properties of Pinus sylvestris wood at five sites in Portugal
vol. 10, pp. 669-679 (online: 11 July 2017)
Research Articles
Density, extractives and decay resistance variabilities within branch wood from four agroforestry hardwood species
vol. 14, pp. 212-220 (online: 02 May 2021)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







