Local neighborhood competition following an extraordinary snow break event: implications for tree-individual growth
iForest - Biogeosciences and Forestry, Volume 7, Issue 1, Pages 19-24 (2014)
doi: https://doi.org/10.3832/ifor1033-007
Published: Oct 14, 2013 - Copyright © 2014 SISEF
Research Articles
Abstract
Sustainable forest management practices and the increased interest of plantation forestry in species mixtures require an understanding of individual-tree growth in complex and diverse forests. Individual-tree growth has been found to be affected by factors such as species identity and size of the target tree as well as of neighboring trees, neighbor density and abiotic factors. However, most of these studies have been conducted in plantations or mixed forests with a very limited number of species. We conducted our study in a subtropical forest in China, which was very species rich and highly heterogeneous with regard to topography (slope inclination and aspect, elevation) and successional status. Prior to our study the forest was subjected to an exceptional snow break event. We asked whether individual-tree basal area increment (BAI) over three growing seasons following the snow break event was related to local biotic and abiotic conditions. We hypothesized that individual-tree BAI is negatively affected by local neighborhood competition, positively affected by local neighborhood diversity, negatively related to slope inclination, and positively related to aspect to south. Individual-tree BAI of four different species was mainly affected by tree size, local neighborhood competition and species identity. Contrary to our expectations, we did not observe significant effects of local neighborhood functional diversity and abiotic conditions. However, we did find a negative effect of plot functional diversity on BAI. This finding may be explained by a negative correlation of plot functional diversity and snow break damage, which was observed in mid- and late-successional stages. The disturbance event did considerably change the competitive local neighborhood interactions by reducing competition, which increased the BAI of target trees in more disturbed/more diverse plots.
Keywords
Basal Area Increment, Crown Projection Area, Functional Diversity, Gutianshan National Nature Reserve, Local Neighborhood Diversity
Introduction
The increasing interest in sustainable forestry necessitates studies of structurally complex and diverse forests in order to understand their ecological processes and to integrate this knowledge in management approaches such as “close to nature” or mixed-species plantation forestry ([25], [2]). In even-aged, simply structured monocultures, stand-based approaches are appropriate to explain and model tree growth. However, in structurally complex forest stands with a heterogeneous mixture of species, tree growth is strongly affected by intrinsic properties of individual trees and their local neighborhood ([9], [17], [51]). Accordingly, individual- tree approaches are much more appropriate than stand-based approaches for growth analyses, and individual-tree growth should be analyzed as a function of tree size, tree vitality, genotype, local neighborhood interactions, and local abiotic site conditions ([34], [27], [45]).
The local neighborhood conditions generally include biotic and abiotic factors. Among the most important biotic factors are tree-tree interactions. These local neighborhood interactions may occur as competition, but also as competitive reduction and facilitation (i.e., complementarity - [24], [18]). These interactions have been modeled as a function of the size, number, and identity of tree individuals. Negative effects on the growth of a target tree have been observed where neighboring trees were larger, denser, or closer. Specifically, since competition for light is known to be size-asymmetric, larger individuals have a disproportionate effect on smaller individuals by shading ([38]). Distance-dependent competition indexes account for the distance between neighboring tree individuals, for their density as well as for their size ratios ([3], [12]).
In mixed-species stands non-equivalent neighbor effects on individual-tree growth may not only be generated by distance and size ratios, but tree species may also differ in their competitive effects. It has been found that conspecifics are stronger competitors than heterospecifics ([41], [31], [35], [45]). This may be due to high competitive equivalence between individuals of the same species, whereas individuals of different species may use resources in a complementary, and thus more efficient way, and, thereby, reduce competition. At the stand level a positive effect of diversity on tree growth has been confirmed for forest systems of different biomes such as the neotropics ([37]), the Mediterranean region ([44]) as well as the temperate and boreal zone ([32]). Following the local neighborhood approach we expect that local diversity should positively affect individual- tree growth as well. However, this has rarely been studied for highly diverse and structurally complex (near-) natural forest ecosystems.
Furthermore, the effects of abiotic factors have to be included in individual-tree growth analyses, because resource availability is not only determined by the local biotic, but also by the local abiotic conditions. On sloping terrain, for example, topography (i.e., slope inclination and aspect) is one of the major factors influencing solar irradiation, temperature and soil moisture availability ([23], [46]).
Finally, tree growth analyses have to consider that growth rates vary through time as a response to changes in environmental conditions (e.g., climate) and local neighborhood interactions, but also as a result of natural or human disturbances. These disturbances selectively remove or kill neighbors from the local environment of residual trees and can, therefore, influence interactions among individuals and the local environment-growth relationship ([20]). Important types of natural disturbances are snow and ice storms ([7]). The resulting tree damages, such as breakage of stems or crown, cause shifts of the light regime of the forest stand, predominantly towards an increased spatial heterogeneity. Damaged tree individuals have been shown to have lower increases in stem diameter than their undisturbed neighbors ([40], [1]). The growth reduction in turn has implications for local neighborhood interactions since this affects the competitive strength of tree individuals.
The aim of the present study was to predict individual-tree growth in a heterogeneous mixed forest stand by means of biotic and abiotic factors of the local neighborhood. The study was conducted in a subtropical broad-leaved evergreen forest in China. China aims at sustainable forest management and large areas have been afforested ([50], [10]). However, afforestations are mostly monocultures causing a variety of environmental problems ([11], [10]).
The studied forest is very species-rich and heterogeneous with regard to topography (slope inclination and aspect) and successional stages. Prior to our study the forest area experienced an exceptional snow break event. In this complex forest stand we wanted to know if individual-tree basal area increment (BAI) over three growing seasons following the snow break event is related to local biotic and abiotic conditions. More precisely we hypothesized:
- (H1): Individual-tree BAI is negatively affected by local neighborhood competition.
- (H2): Individual-tree BAI is positively affected by local neighborhood diversity.
- (H3): Individual-tree BAI is negatively related to slope inclination.
- (H4): Individual-tree BAI is positively related to aspect to south.
Material and Methods
Study site
The study was conducted in the Gutianshan National Nature Reserve (NNR), which is located in Zhejiang province, P.R. China (29º 8.18 - 29º 17.29 N, 118º 2.14 - 118º 11.12 E). The Gutianshan NNR has an area of about 81 km2 with elevations ranging from 250 to 1250 m a.s.l. It is situated in the subtropics with a warm temperate climate and a short dry season in winter. The mean annual temperature is 15.1 °C and the mean annual precipitation is about 1964 mm ([22]).
The potential natural vegetation is subtropical broad-leaved evergreen forest, with a similar number of coexisting evergreen and deciduous broad-leaved species, though evergreen species is dominating in abundance ([22]). The forest is species-rich as 59 woody species > 1m height have been identified on a plot of 30 x 30 m size ([8]). The area is heterogeneous with respect to the successional stages, with older stands dominated by Castanopsis eyrei (Champ. ex Benth.) Hutch. and Schima superba Gardn. et Champ. and younger dominated by Pinus massoniana Lamb. and Quercus serrata Murray var. brevipetiolata. Our study sites comprise intermediate to steep slopes (ranging from 10° to more than 50°) and small-scale changes of aspect. The study area was affected by an extraordinary snow break event in January/February 2008 ([52]) with many trees exhibiting stem breakage or crown loss of differing extent ([30]).
Study design
Field data have been collected in autumn of 2008 and 2011 on 27 plots of 30 x 30 m size within the framework of the BEF (Biodiversity and Ecosystem Functioning) China project. The plots were randomly chosen within the Gutianshan NNR (limited by inaccessibility and avoiding areas of high snow break damage) and stratified by successional stage: (1) <20 yrs; (2) <40 yrs; (3) <60 yrs; (4) <80 yrs; (5) ≥80 yrs ([8]). The elevation of plot locations ranged from 250 to 900 m.
We selected four target species of high abundance ([49]): Castanea henryi (Skan) Rehd. et Wills. and Q. serrata var. brevipetiolata (both deciduous), C. eyrei and S. superba (both evergreen). Within the plots 20 target trees per species (10 for C. henryi) were chosen randomly from all individuals complying with the following criteria: (i) single stemmed; (ii) diameter at breast height (dbh, 1.3 m above ground) >10 cm (intermediate and old plots) or dbh >3 cm (in young plots); (iii) crown position in the upper canopy; (iv) no or only slight crown damage caused by the snow break event; (v) each target species could only be selected once per plot. Following the local neighborhood approach we assume that important interactions take place between neighboring tree individuals ([34]). Target tree neighbors were defined by using the reversed cone method following Biging & Dobbertin ([3]) with an opening angle of 70° of the cone. This approach was used because in a native forest remnant of the same study region Von Oheimb et al. ([45]) found that the mode of competition in this forest type is primarily size-asymmetric, i.e., competition for light. For a tree on a slope, the crowns of neighboring trees in an uphill direction are situated at a higher position than those of the same-sized neighbors in a downhill direction and may, thus, impose a stronger influence on individual- tree growth at a given height. The search cone method takes this into account by giving more weight to tree height than distance: more neighbors are included in uphill than in downhill direction at a given opening angle. Each target tree together with its neighbors formed a target group. For a more detailed description we refer to Lang et al. ([27]).
Field data
We measured the dbh of all target trees in 2008 and 2011 by means of a diameter measurement tape. The measurement positions on the trees were permanently marked with white color. Radial increment was calculated as basal area increment per year (BAI in cm2 yr-1). The relative position of the neighbors to the target tree was determined by recording the azimuth and horizontal distance from stem base to stem base. For all individuals eight crown radii in the cardinal and ordinal directions were measured, each being the distance, determined by means of a densiometer, from crowntip to the stem. Crown projection areas (CPA) of all tree individuals were calculated as polygons based on the eight radii. Crown measurements took place in 2008 and 2011. The horizontal distances of all neighboring trees to their target tree and the crown radii were measured using a Forester Vertex Hypsometer (Haglöf, Sweden).
To describe the local competition experienced by each target tree, we used a distance dependent competition index (CI) modified after Castagneri et al. ([12]), with the CPAs to describe the size of tree individuals (eqn. 1):
where CPAj is the neighbor tree crown projection area (m2); CPAt is the target tree crown projection area (m2); disttj is the horizontal distance (m) between the neighbor j and the target tree t.
Two competition indexes were calculated. One was based on CPAs of 2008 (CI08) and refers to the situation immediately after the snow break event. The other competition index was based on CPA measurements of 2011 (CI11), thus including the growth reactions of crowns three growing seasons after the disturbance event.
Local and plot functional diversity was calculated as Rao’s quadratic entropy (Rao’s Q - [36], [5]) of eight functional leaf traits (these were specific leaf area; leaf dry matter content; content of nitrogen, aluminium, calcium and phosphorus; C/N ratio; stomata density) of either all neighboring trees of a target group (local functional diversity) or all individuals >1 m height of the plot (plot functional diversity, for trait data description see [26]). A detailed description of the calculation of Rao’s Q is provided by Lang et al. ([29]).
Furthermore, the local environmental parameters slope inclination and aspect and plot elevation a.s.l. were recorded. To quantify the severity of disturbance of the snow break event we first estimated the percentage of crown loss of former undisturbed crowns of each tree individual. Categories were defined as undamaged (0% loss) = 0, 1-25% loss = 1, …, crown completely broken 100% loss = 5. Plot level disturbance was then calculated as the summed mean category scores of all individuals divided by the number of observed individuals.
Statistical analyses
A total of 65 target trees entered the analyses. Between 2008 and 2011 one plot with two target trees was destroyed. Two of the small diameter target trees (10 cm and 11 cm dbh, respectively) showed the highest absolute BAIs, and the target tree with the largest dbh (56 cm in 2008) had an extremely low BAI. These five target trees were excluded from the analyses, resulting in 10 target trees of C. henryi, 19 target trees of C. eyrei, 18 target trees of Q. serrata and 18 target trees of S. superba which have been analyzed.
We tested the effects of the initial dbh, environmental parameters (slope inclination and aspect, elevation, damage caused by the snow break event), local neighborhood competition, local neighborhood and plot diversity and species identity on the BAI of target trees. Two full linear mixed effects models were compared with regard to the competition indexes used: CI08 vs. CI11. Plot was fitted as a random factor of the mixed effects models to account for the spatial nesting of data, namely the blocking of target groups within plots. Model simplification was done by stepwise backward selection of fixed factors, removing the least significant variables until only significant predictor variables were left (p < 0.05).
The response variable was square root transformed to meet the criterion of normality. Prior to the analyses, all predictor variables have been checked for collinearity (correlations between variables did not exceed R = 0.6) and all numerical variables have been centered and scaled to ensure comparability of results. We assessed by plotting that residuals were well behaved, that the response variable is a reasonable linear function of the fitted values and that errors are normally distributed within the plots ([14]).
Differences in crown loss due to the snow break event between plots of different successional stages were assessed by ANOVA. The relation between plot functional diversity and crown loss of older plots (successional stages 3-5) was analyzed by a linear model.
All statistical analyses were performed using R 2.15.3. Linear mixed effects models were analyzed by means of the package “nlme” ([33]) and multiple comparisons were performed using the package “multcomp” ([21]).
Results
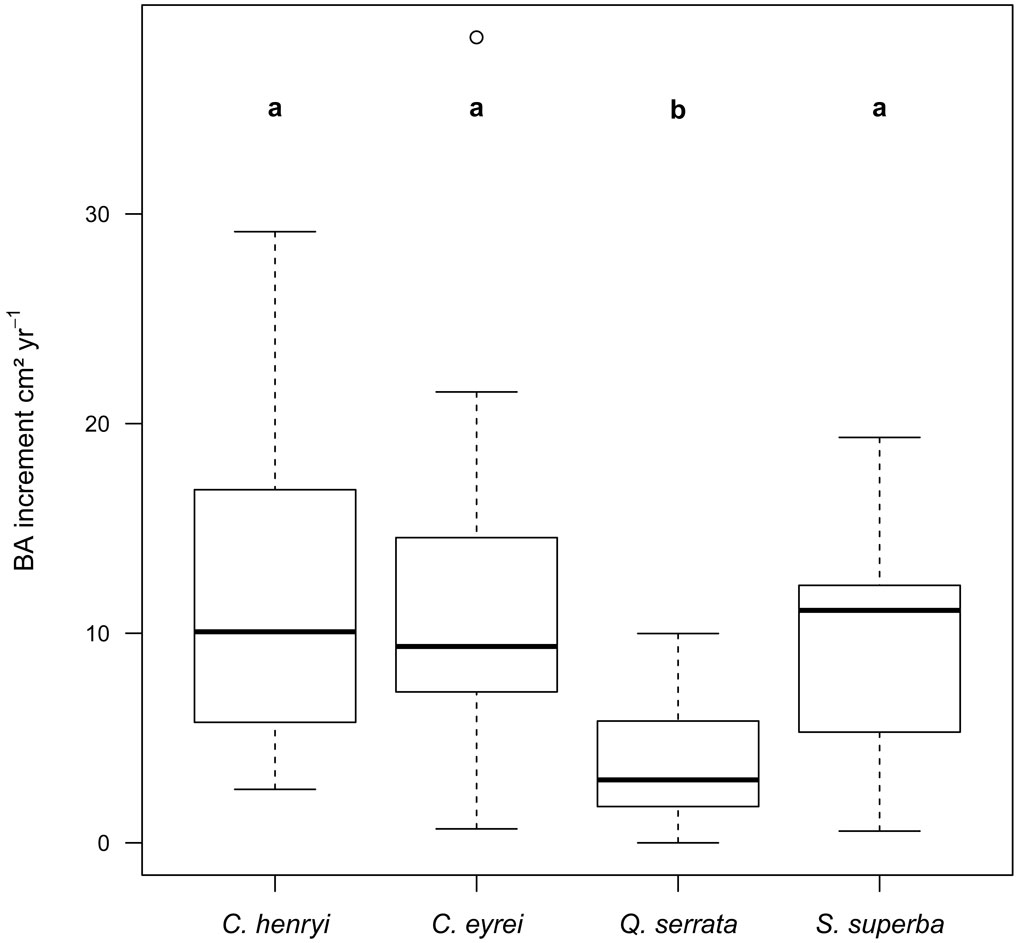
The variability in target tree BAI was mainly explained by initial dbh (effect size: 0.34, model A and 0.42, model B - Tab. 1). The thicker the stem of an individual-tree, the higher was its BAI. Furthermore, BAI was significantly different between the tree species. With a mean initial diameter of 18 cm BAI amounted to 12.1 ± 0.1 cm2 yr-1 in C. henryi, to 9.0 ± 0.1 cm2 yr-1 in C. eyrei, to 7.0 ± 0.1 cm2 yr-1 in S. superba and to 4.6 ± 0.1 cm2 yr-1 in Q. serrata, as calculated from species-specific regressions (for observed means see Fig. 1). The BAI of Q. serrata was significantly lower than those of the other species (post-hoc Tukey Test, p < 0.05).
Tab. 1 - Comparison of the most parsimonous models explaining BAI, original full models have been fitted with competition based on crown data of 2008 (Model A) and based on crown data of 2011 (Model B).
| Model | Parameters | DF | F-value | P-value |
|---|---|---|---|---|
| Model A | Initial dbh | 35 | 16.24 | <0.001 |
| Species identity | 35 | 3.8 | 0.011 | |
| Model B | Initial dbh | 34 | 17.72 | <0.001 |
| Competition | 34 | 5.97 | 0.021 | |
| Plot functional diversity | 25 | 5.39 | 0.021 | |
| Species identity | 34 | 2.88 | 0.037 |
Fig. 1 - Boxplots of BAI of target species. Different characters denote significant differences.
Although our study area was highly variable with regard to topography, neither slope inclination and aspect nor elevation was included among the significant predictors in the final models. Likewise, we did not find an effect of snow damage on target tree BAI.
The competitive strength of neighboring trees, expressed as CI11, negatively affected BAI (effect size: -0.19 - Tab. 1). However, only CI11 could be retained in the final model, whereas CI08 had no predictive power. This was further confirmed by a comparison of the two models containing initial dbh, CI08 (model 1) or CI11 (model 2), plot functional diversity and species identity (Akaike’s Information Criterion - AIC: 166.15 vs. 165.89).
Local functional diversity had no predictive power, whereas plot functional diversity was retained in the final model with CI11 (Tab. 1). Plot functional diversity negatively affected BAI and had a similar effect size as CI11 (effect size: -0.25).
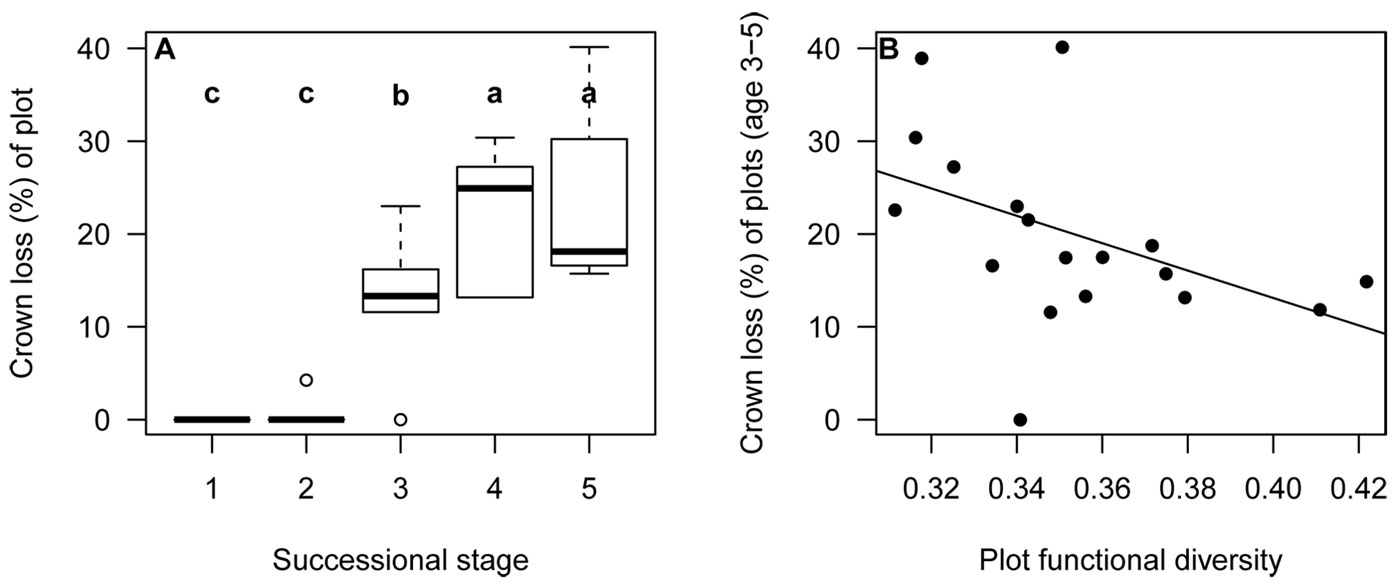
When comparing the damage due to snow break between plots of different successional stages, we found that only plots of the intermediate and late stages have been affected (ANOVA: F-value = 100.1; p < 0.001). Young stages did not show any damage at all (Fig. 3). Using the plots of the intermediate and late stages we found that the degree of disturbance by snow break was significantly less in plots of higher functional diversity (linear model: estimate = -0.43; p < 0.001 - Fig. 3).
Fig. 3 - Percentage of crown loss due to a snow break event per plot for different successional stages (A) and correlation of crown loss of older plots and plot functional diversity (B). Different characters denote significant differences.
Discussion
This study provided evidence that in heterogeneous species-rich forests BAI of target trees of four species was mainly determined by tree size, local neighborhood competition and species identity. Surprisingly, we found no effect of local neighborhood functional diversity but a negative effect of plot functional diversity on BAI. For plots of mid- and late-successional stages this may be explained by the stronger snow break damage with higher plot functional diversity.
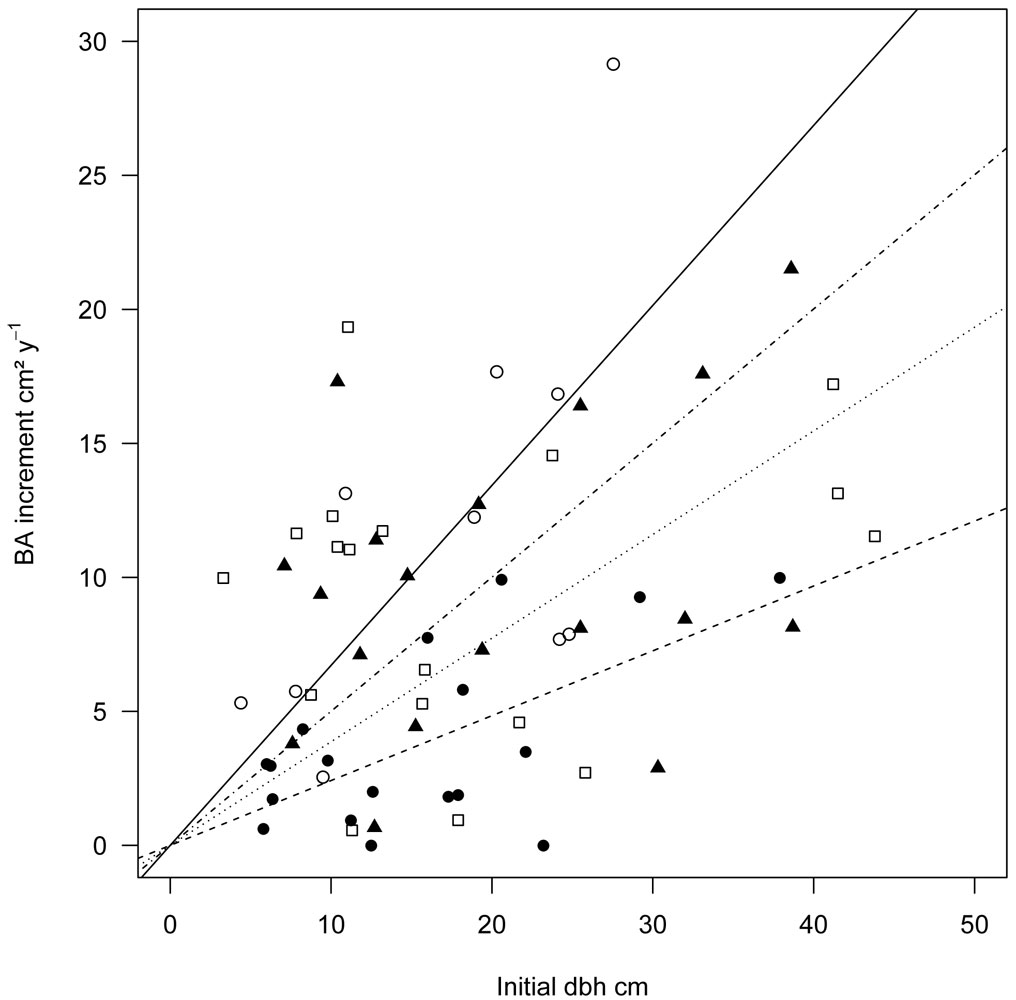
Our models showed that the initial dbh had a strong positive effect on BAI. This is in accordance with other studies, which emphasize the importance of size factors in predicting individual-tree radial growth ([42], [16], [6]). However, there is considerable variation in the diameter-BAI relationship, as displayed in Fig. 2. In a natural mixed-species and multi-age forest the diameter-BAI relationship is obscured by the coexistence of slow-growing older trees and fast-growing younger ones ([6]). Additionally, species-specific differences in BAI contribute to this finding. These differences indicate that there is interspecific variation in functional traits between the four tree species. For three of the four species Lang et al. ([28]) found species-specific differences in sapling growth in the same study region. Furthermore, Chen et al. ([15]) observed high growth rates for saplings of S. superba in forest gaps, and Q. serrata was recorded to be very slow growing ([47]). However, further studies are needed to elucidate the relationships between functional traits and growth rates in these subtropical forests.
Fig. 2 - Linear regressions of BAI over initial dbh (2008) for each target species. Open circles; unbroken line: C. henryi; triangles; dotdashed line: C. eyrei; bullets; dashed line: Q. serrata; quadrats; dotted line: S. superba.
Topography (i.e., slope inclination and aspect) has important implications for the direction and intensity of incoming radiation ([23]). In the northern hemisphere, south-facing slopes receive much more solar irradiation than north-facing slopes, creating strong gradients of light, temperature and soil moisture: higher light levels and temperatures as well as lower soil moisture corresponds with south-facing slopes. Furthermore, slope inclination may correlate with edaphic conditions, in particular soil depth, soil moisture and nutrient availability. Steeper slopes may be shallower, and therefore may limit rooting space in one dimension. Slope inclination has been shown to affect the stature of a tree in terms of stem inclination and crown asymmetry ([43], [27]). Contrary to our third and fourth hypothesis, however, topography had no effect on BAI in our study. High morphological plasticity of the target species may be the reason for the low sensibility of radial growth towards topographic conditions in these highly variable environments ([27]).
Competition
The results of our study confirm our first hypothesis. Local neighborhood competition (expressed as CI11) was an important predictor of individual-tree BAI. We calculated the CI with tree data from two different points in time, but, interestingly, we found that only CI11 was significantly related to BAI, whereas CI08 was not. Furthermore, the full model 2 including CI11 had stronger predictive power than the full model 1 in which CI08 was included. CI08 was calculated based on the conditions directly after the snow break event, but did not contribute to explain the variability in BAI over the three growing seasons following this event. The extreme snow break event of January 2008 is clearly an important natural disturbance of these subtropical forests, occurring with a mean frequency of 50 to 100 years ([39]). When establishing the permanent plots in summer 2008, we selected plots with comparatively low damage by the snow storm. Nevertheless, our results show that this rare disturbance event reduced competition also in the less damaged forest patches, which may considerably change the competitive interactions.
Diversity
In contrast to our second hypothesis, we did not find any effect of local neighborhood functional diversity on BAI. There might be two possible explanations for this finding. Firstly, it has been found that complementary effects vary over time, with stand development and local neighborhood species composition ([4], [13], [19]). Since this is an observational study we could not strictly control the different factors. Future research in a newly established large-scale forest BEF experiment of the BEF-China project is designed to systematically analyze species interactions under controlled conditions ([48]). Secondly, the disturbance caused by the snow break event had a strong effect on local neighborhood interactions. These effects may have overruled local diversity effects. Future investigations of the local neighborhood interactions in the permanent study plots will elucidate whether there is a longer term influence of the snow break event on tree growth.
Surprisingly, we did find a negative effect of plot functional diversity on BAI. We suspect that this is not a direct effect. We found that young plots did not suffer from damage caused by snow break. This is in general accordance with Man et al. ([30]) who observed the largest proportion of undamaged trees in the lowest dbh class included in their study in the Gutianshan NNR (i.e., 8 - 10 cm). Small trees are presumably less prone to snow break damage due to their relatively smoothness of stems and restricted CPA. Including only plots from intermediate and late successional stages we found that snow break was reduced in plots of higher functional diversity (Fig. 3). Thus, we hypothesize that a higher crown loss in less diverse plots, increased incoming sunlight and canopy space, for enhancing individual-tree growth. In our study sites, Lang et al. ([29]) found that higher plot functional diversity was correlated with higher horizontal heterogeneity of the canopy. Furthermore individual tree CPA has been found to increase with local functional diversity ([27]). Thus, the heterogeneous light conditions in more diverse stands may promote optimal crown growth and stability, leading to a decrease of snow break damage.
Conclusions
The pronounced differences between CI08 and CI11 point to the relevance of multiple assessment of competition in time ([20]). We conclude that the influence of snow break on local neighborhood interactions, rather than the direct effect of the event, affected the growth response of surviving trees. These strong effects may impact stand structure and dynamics in the long run. Thus, we may ask if in future the disturbances of these extreme weather events, even more than today, may represent the main driver of forest growth and structure.
Acknowledgments
We are grateful to the Gutianshan NNR administration for the access permit to the forest reserve. In particular, we are indebted to Teng Fang for species identification. This work was funded by the German Research Foundation and carried out within the research group BEF China (DFG FOR 891/1), as well as by travel grants by the Sino-German Center for Research Promotion in Beijing (GZ 524, 592, 698 and 699). We are grateful for this support. The manuscript was much improved by the comments of two anonymous reviewers.
References
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
CrossRef | Gscholar
Authors’ Info
Authors’ Affiliation
Werner Härdtle
Goddert von Oheimb
Leuphana University Lüneburg, Faculty Sustainability, Institute of Ecology, Scharnhorststr.1, D-21335 Lüneburg (Germany)
University of Halle, Institute of Biology / Geobotany and Botanical Garden, Am Kirchtor 1, D-06108 Halle (Germany)
German Centre for Integrative Biodiversity Research (iDiv), Deutscher Platz 5d, D-04103 Leipzig (Germany)
Corresponding author
Paper Info
Citation
Lang AC, Härdtle W, Bruelheide H, von Oheimb G (2014). Local neighborhood competition following an extraordinary snow break event: implications for tree-individual growth. iForest 7: 19-24. - doi: 10.3832/ifor1033-007
Academic Editor
Emanuele Lingua
Paper history
Received: May 03, 2013
Accepted: Jul 07, 2013
First online: Oct 14, 2013
Publication Date: Feb 03, 2014
Publication Time: 3.30 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2014
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 59555
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 50086
Abstract Page Views: 3526
PDF Downloads: 4286
Citation/Reference Downloads: 34
XML Downloads: 1623
Web Metrics
Days since publication: 4634
Overall contacts: 59555
Avg. contacts per week: 89.96
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2014): 1
Average cites per year: 0.08
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Relationship between tree growth and physical dimensions of Fagus sylvatica crowns assessed from terrestrial laser scanning
vol. 8, pp. 735-742 (online: 11 June 2015)
Research Articles
Relationships between overstory and understory structure and diversity in semi-natural mixed floodplain forests at Bosco Fontana (Italy)
vol. 9, pp. 919-926 (online: 21 August 2016)
Technical Advances
Improved estimates of per-plot basal area from angle count inventories
vol. 7, pp. 178-185 (online: 17 February 2014)
Research Articles
Advantages of the point-intercept method for assessing functional diversity in semi-arid areas
vol. 8, pp. 471-479 (online: 31 October 2014)
Research Articles
Allometric models for the estimation of foliage area and biomass from stem metrics in black locust
vol. 15, pp. 281-288 (online: 27 July 2022)
Research Articles
Functional turnover from lowland to montane forests: evidence from the Hyrcanian forest in northern Iran
vol. 8, pp. 359-367 (online: 16 September 2014)
Research Articles
Scots pine’s capacity to adapt to climate change in hemi-boreal forests in relation to dominating tree increment and site condition
vol. 14, pp. 473-482 (online: 18 October 2021)
Research Articles
Tree aging does not affect the ranking for water use efficiency recorded from δ13C in three Populus deltoides × P. nigra genotypes
vol. 12, pp. 272-278 (online: 21 May 2019)
Research Articles
High resolution biomass mapping in tropical forests with LiDAR-derived Digital Models: Poás Volcano National Park (Costa Rica)
vol. 10, pp. 259-266 (online: 23 February 2017)
Research Articles
Effects of different silvicultural measures on plant diversity - the case of the Illyrian Fagus sylvatica habitat type (Natura 2000)
vol. 9, pp. 318-324 (online: 22 October 2015)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







