
Biomass production of young lodgepole pine (Pinus contorta var. latifolia) stands in Latvia
iForest - Biogeosciences and Forestry, Volume 6, Issue 1, Pages 10-14 (2013)
doi: https://doi.org/10.3832/ifor0637-006
Published: Jan 14, 2013 - Copyright © 2013 SISEF
Short Communications
Collection/Special Issue: IUFRO 7.01.00 - COST Action FP0903, Kaunas (Lithuania - 2012)
Biological Reactions of Forest to Climate Change and Air Pollution
Guest Editors: Elena Paoletti, Andrzej Bytnerowicz, Algirdas Augustaitis
Abstract
Biomass as a source of renewable energy is gaining an increasing importance in the context of emission targets set by the European Union. Large areas of abandoned agricultural land with different soils are potentially available for establishment of biomass plantations in the Baltic states. Considering soil and climatic requirements as well as traits characteristic for lodgepole pine (Pinus contorta Dougl. var. latifolia Engelm) and the scarcity of published knowledge, we assessed the above-ground biomass of Pinus contorta in comparison to that of native Scots pine (Pinus sylvestris L.) and factors affecting biomass production. Data were collected in 3 experimental trials, located in two sites in central part of Latvia: Zvirgzde and Kuldiga (56°41’ N, 24°28’ E and 57°03’ N, 21°57’ E, respectively). Trials were established with density 5000 tree ha-1, using seed material from Canada (50°08’-60°15’ N, 116°25’-132°50’ W) and two Pinus contorta stands with unknown origin growing in Latvia. Results reveal that absolute dry aboveground biomass of Pinus contorta reaches 114 ± 6.4 t ha-1 at age 16 on a fertile former arable land, 48 ± 3.6 and 94 ± 9.4 t ha-1 at age 22 and 25, respectively, on a sandy forest land (Vacciniosa forest type). The biomass is significantly (p < 0.01) and considerably (more than two-fold) higher than that of the native Pinus sylvestris and the productivity is similar (in fertile soils) or higher (on poor soils) than reported for other species in energy-wood plantations. Provenance was a significant factor affecting the above-ground biomass, and the ranking of provenances did not change significantly between different soil conditions. It provides opportunities for further improvement of productivity using selection.
Keywords
Introduced Species, Productivity, Provenance, Above-ground Biomass
Introduction
Use of biomass for energy has increased in Baltic Sea region countries in the last decade. It is partly a price-driven process and partly related to strategic goals of European Union to increase share of renewable energy in the total energy consumption ([6]). In Latvia this share has to reach 40% in year 2020. Increased use of renewable energy decreases the dependence on imported fossil materials and is an important aspect in climate change mitigation effort ([5]). In forest-rich countries as Fenoscandia and Baltic states, biomass from wood is an important component in the total renewable energy production. Use of logging residues and stumps from clear-cuts is rapidly increasing and is expected to rise in the next years as a consequence of strategic decisions of countries ([23], [39]). Technologies have been developed to facilitate the use of logging residues from thinning. However, this source of biomass has several limitations: in some areas soil depletion and loss of increment in next rotation cycles ([14]), quality of biomass, accessibility, transportation distance. Therefore establishment of plantations for biomass production has been considered in the Baltic states, where large areas of abandoned agricultural land suitable to this purpose are available. In Latvia, the above areas cover approximately 1.44 million ha ([37]). Currently, broadleaved trees - hybrid aspen (Populus tremuloides x P. tremula), grey alder (Alnus incana) and bushes (Salix spp. breeds) are used and recommended for the establishment of biomass plantations ([18]). All the above species require fertile soils and/or fertilization to ensure productivity ([26], [19]). However, the abandoned agricultural lands are often on poor soils, therefore looking at alternative species (including Pinus contorta) is a relevant objective for biomass production.
Native range of Pinus contorta in northwestern America spreads from latitude 30 to 64° N and from 0 to 3900 m in elevation ([38]). Establishment of Pinus contorta plantations at commercial scale in Sweden started around 1970 ([8]), and was based on earlier research results on superior productivity of this species in comparison to native Pinus sylvestris ([21]). Natural regeneration of Pinus contorta occurs after forest fires or clear cuts and is often very dense. High density of stands can be retained throughout the rotation period, so that even at the age of 90 years there are still not less than 2000 trees ha-1 ([33]). Furthermore, greater survival, earlier exposure of new needles, longer needles with a larger surface area, fast root development and a higher productivity per unit of nitrogen ([27]) make Pinus contorta a suitable species for biomass production.
Biomass equations and estimates have been developed and published for stands in the native range of Pinus contorta ([28]). For Europe, a very limited amount of such studies is available and usually only stem biomass is calculated ([24], [25]). Comprehensive analysis of biomass equations by Zianis et al. ([41]) reveals only one available equation for total above-ground biomass of Pinus contorta, developed in Iceland.
Considering the potential of the species and the lack of comprehensive information on its biomass production, the aim of this study was to assess the above-ground biomass of Pinus contorta Dougl. var. latifolia Engelm. in comparison to the native Pinus sylvestris and factors affecting biomass production.
Material and methods
Study was based on 3 experiments (Tab. 1), located in two sites in central part of Latvia: Zvirgzde and Kuldiga (56° 41’ N, 24° 28’ E and 57° 03’ N, 21° 57’ E, respectively). Trials in Zvirgzde (No 82 and No 707) were located on poor, sandy soil, Vacciniosa forest type (according to classification by [4]), while the trial in Kuldiga (No 702) was on fertile clay soil, former arable land, approximately corresponding to the Oxalidosa forest type. Soil ploughing was carried out in rows prior to planting. Initial spacing in all trails was 2x1m (5000 trees ha-1), two-years-old bare-rooted plants were used, no thinning carried out prior to measurement. Experiments No 702 and No 707 consist of the same set of Pinus contorta provenances from Canada (50° 08’ - 60° 15’ N, 116° 25’ - 132° 50’ W), and one seed lot from a Pinus contorta stand from Latvia (Tab. 2). Experiment No 82 includes only 3 provenances (54° 24’ - 58° 38’ N, 122° 45’ W), each represented by 5 open-pollinated families.
Tab. 1 - Characteristics of the experimental plots studied.
| Exp. No | Site | Soil | Forest type | Pinus contorta provenances | Age |
|---|---|---|---|---|---|
| 82 | Zvirgzde | poor, sandy | Vacciniosa | 3 from Canada | 25 |
| 707 | Zvirgzde | poor, sandy | Vacciniosa | 14 from Canada; 2 from Latvia | 22 |
| 702 | Kuldiga | fertile, clay | Oxalidosa | 13 from Canada; 1 from Latvia | 16 |
Tab. 2 - Pinus contorta provenances represented in trials in Latvia. (*): seeds form unknow origin from Pinus contorta stands growing in Latvia.
| Provenance | Latitude | Longitude | Altitude | No702 | No707 | No82 |
|---|---|---|---|---|---|---|
| Teslin, Yukon | 60°15’ | 132°50’ | 750 | X | X | - |
| Fort Nelson | 58°38’ | 122°41’ | 495 | - | - | X |
| Pink Mountain | 57°00’ | 122°15’-45’ | 850 | X | X | X |
| Cecil Lake | 56°25’ | 120° | 700 | X | X | - |
| Hudson Hope | 56°00’-05’ | 121°35’-59’ | 500-800 | X | X | - |
| Moberly Lake | 55°40’ | 121°25’ | 750 | X | X | - |
| Muskey Lake | 55°30’ | - | 900 | X | X | - |
| Fort St. James | 54°30’ | 124°10’ | 900 | X | X | - |
| Summit Lake | 54°24’ | 122°37’ | 813 | - | - | X |
| Smithers | 54°15’ | 127°00’ | 650 | X | X | - |
| Valemount | 53° | 119° | 850 | X | X | - |
| Rocky Mountain House | 52°37’ | 115°20’ | 1000 | - | X | - |
| Nordegg | 52°20’ | 116°25’ | 1400 | X | X | - |
| Falkland | 50°32’ | 119°40’ | 1300 | X | X | - |
| Lac le Jeune | 50°29’ | 120°29’ | 1250 | X | X | - |
| Monashee | 50°08’ | 118°30’ | 950 | X | X | - |
| Skriveri* | 56°40’ | 25°10’ | 29 | X | X | - |
| Bukulti* | 57°01’ | 24°14’ | 12 | - | X | - |
Each seed lot (provenance or family) was represented by 50 to 60 tree block plots (in experiments No 702, 707 and 82, respectively), randomly distributed in 4 replications. One to two Latvia’s Pinus sylvestris provenances were used as controls in every experiment. Height and diameter was measured for every tree at age 16 (trial No 702), 22 (No 707) and 25 (No 82). To improve representation of Pinus sylvestris for comparison among species, sample plots (500m2) were established in 20 randomly selected stands (age: 22 years) in the same forest type and tree heights and diameters were measured. For comparison purposes, material for experiment No82 was increased using data from neighboring Pinus sylvestris trial No 19, established at the same year with the same spacing. Moreover, data from altogether 20 seed lots from phenotypically selected seed orchards, representing different regions of Latvia, were included in the analysis.
Above-ground biomass (including whole trunk and all branches with needles) was estimated with equations based on measurements of 221 sample trees of Pinus contorta from experiment No 82 and 90 trees of Pinus sylvestris from experiment No 19 and forest stands (Jansons et al., unpublished - eqn. 1):
for Pinus sylvestris and (eqn. 2)
for Pinus contorta, where D is the diameter at breast height, and H is the total height of the tree.
Both equations provided precise estimates of above ground biomass for the respective species and tree dimensions in Latvia (R2 = 0.93 and R2= 0.96, respectively).
Equations were estimated using linear and non-linear regression analysis; significance of differences was determined using t-test and ANOVA when the assumptions of parametric tests were met, or using Mann-Whitney U-test and Kruskal-Wallis test when assumptions were violated. Pearson’s correlation coefficient was used as a measure of linear dependency. All the analyses were performed using the software R (R Development Core Team, version 2.12.1).
Results and discussion
Results reveal that above-ground biomass of Pinus contorta reaches 114 ± 6.4 t ha-1 at the age of 16 years on a fertile former arable land (Fig. 1). It is more than two-fold of that observed for Pinus sylvestris in the same conditions (50 ± 15.9 t ha-1), and significantly higher (p < 0.01) in comparison to older Pinus contorta trials on poor sandy forest soils at age 22 and 25 (48 ± 3.6 t ha-1 and 94 ± 9.4 t ha-1, respectively). Also in trials on forest land, above-ground biomass of Pinus contorta significantly exceeded (p < 0.05) the above-ground biomass of Pinus sylvestris.
Fig. 1 - Above-ground biomass of young Pinus contorta and Pinus sylvestris stands in Latvia. Pinus sylvestris biomass was recalculated assuming the same survival as for Pinus contorta in a particular trial. For more details, see text.
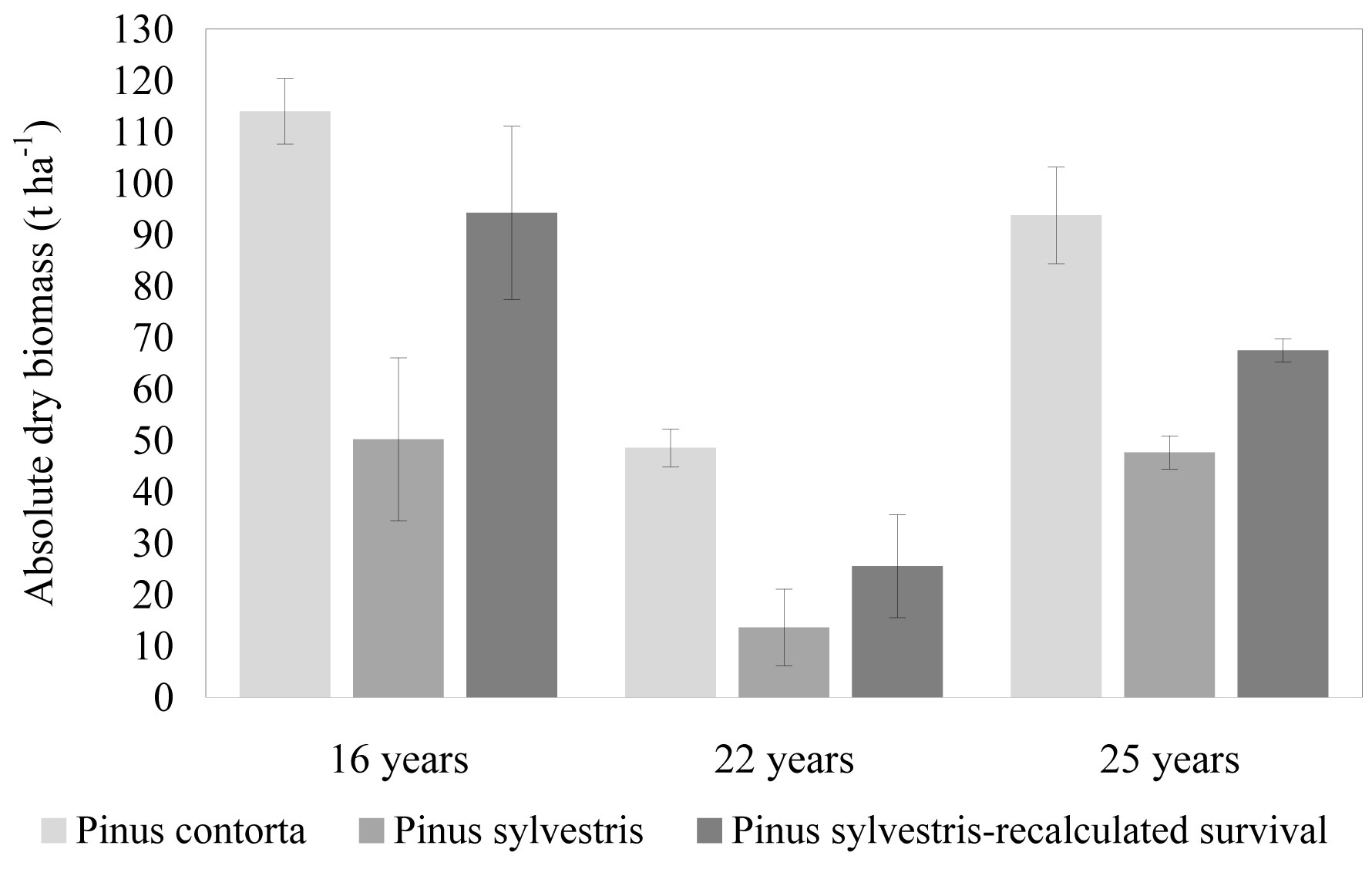
Higher productivity of Pinus contorta in comparison to Pinus sylvestris has been found in extensive trials in Sweden. For example, on average two-fold yield difference between the above-mentioned species was detected at the age of 23 years ([11]) and 36-38% difference in wood dry mass was found at age 20 ([7]) in trials located in similar climatic conditions as in Latvia. These results were the basis for an intensive introduction of this species in Sweden ([8], [22]).
Part of the detected differences in biomass production on per ha basis can be explained by differences in survival rate, which was 81 ± 3.1 % for Pinus contorta and only 44 ± 11.8 % for Pinus sylvestris at the age of 16 years. Similar significant differences in survival among pine species were observed also in older trials: 72 ± 4.7 % vs. 37 ± 8.7 % and 58 ± 4.2 vs. 41 ± 2.4 % at age 22 and 25, respectively.
High survival of Pinus contorta provenances from latitude above 50° N was found also in Estonia at the age of 19 years (67 %) and in Sweden at the age of 18 years (71-92 % on average) in trials with climatic conditions similar to those in Latvia ([20], [10]). A general trend of higher survival for Pinus contorta in comparison with Pinus sylvestris was also reported from a comprehensive analysis of sets of 10 to 20-years-old stands in Sweden ([9], [7]). Difference in survival between species was ranging from 15 to 45 % at the age of 23 years ([11]). The observed differences could be partly explained by the influence of needle cast (Lophodermium seditiosum Minter, Staley & Millar) reducing the survival of young Pinus sylvestris ([2], [1], [12]), but not of Pinus contorta. Differences between species may also be due to the self-thinning rate, since the initial density in trials is rather high (5000 trees ha-1).
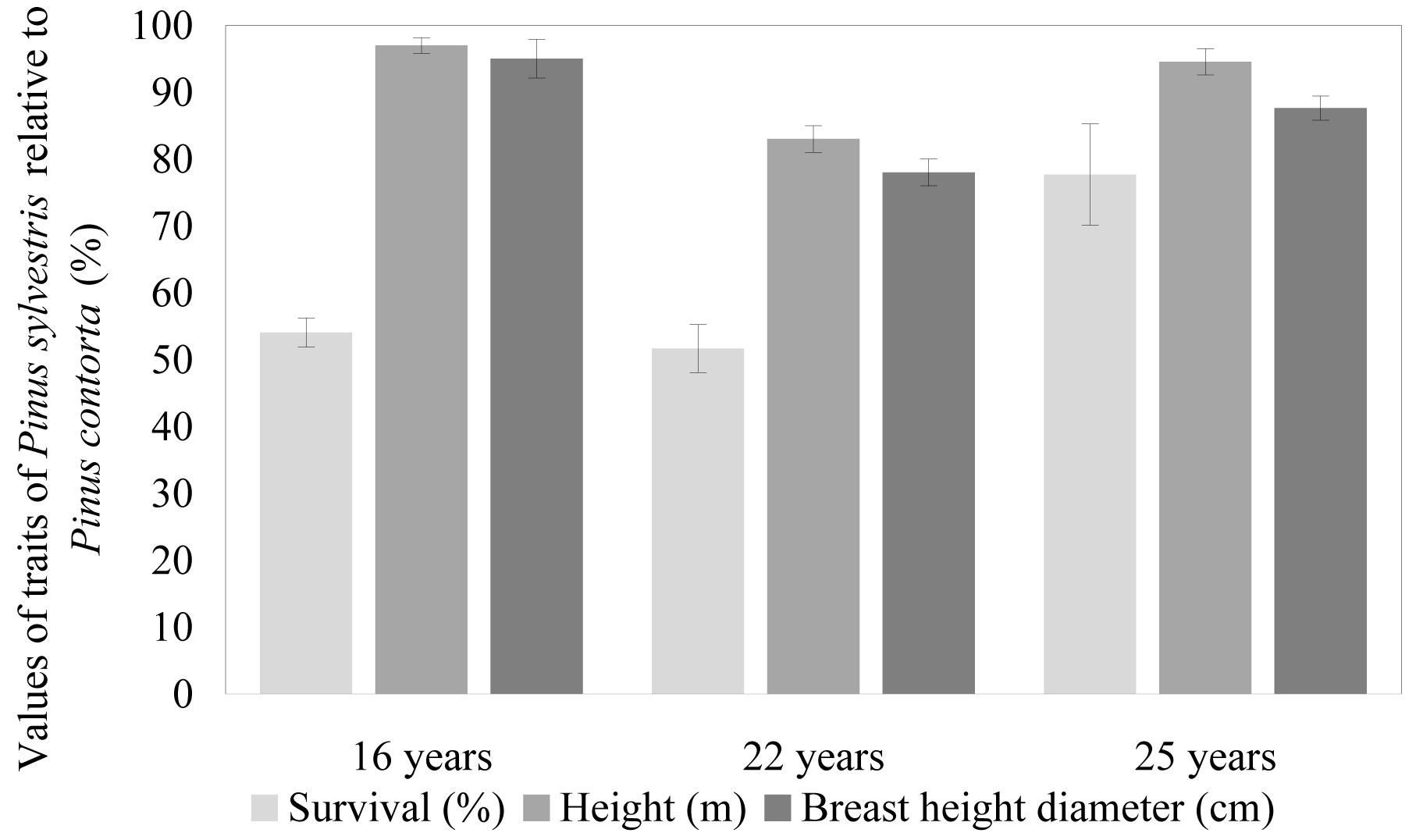
If the biomass of Pinus sylvestris in trial No 702 is recalculated using average survival rate of Pinus contorta, it reaches 90.4 t ha-1, i.e., on average 80 % of that found for Pinus contorta. Even larger relative differences among species remain in older trials on forest land: survival-adjusted biomass of Pinus sylvestris reached only 45 % of that estimated for Pinus contorta (Fig. 1). Moreover, differences in average tree dimensions between species were more pronounced in older trials (Fig. 2). Increasing differences in average tree dimensions were found also from the analysis of sample trees from trial No 82 (Jansons et al. - unpublished). It can be concluded that Pinus contorta has a remarkably higher above-ground biomass than Pinus sylvestris on both poor and fertile soils and superiority is related to differences in both survival and growth rate.
Fig. 2 - Productivity and survival of Pinus sylvestris relative to that observed for Pinus contorta in a particular trial.
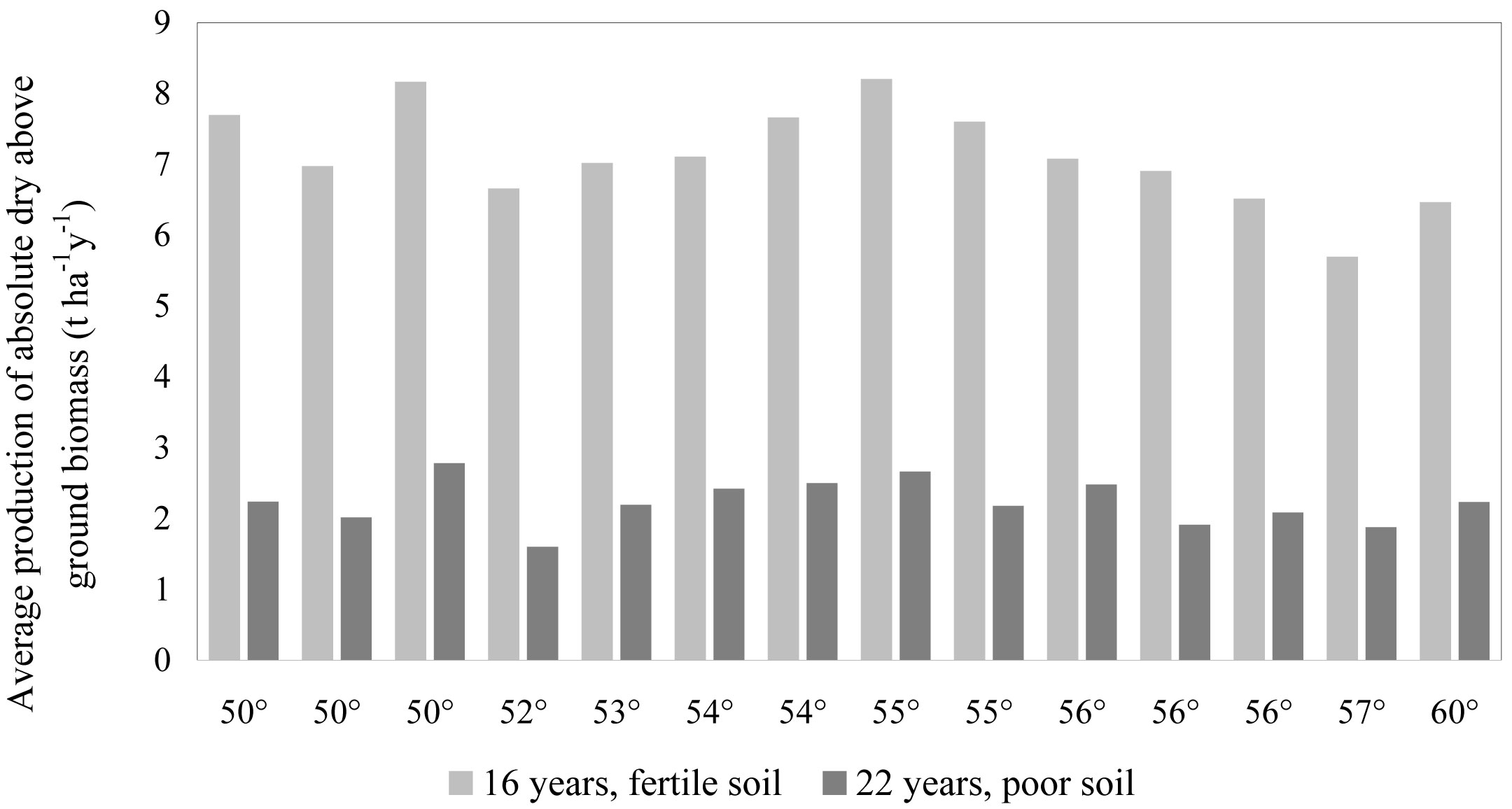
Mean annual dry biomass production of Pinus contorta on fertile soil at the age of 16 years reached 7.1 ± 0.40 t ha-1, but on poor soils it was only 2.2 ± 0.17 t ha-1 at age 22 and 3.8 ± 0.36 t ha-1 at age 25. In all trials, biomass production of Pinus contorta significantly exceeded (p < 0.01) that of Pinus sylvestris. Provenance had also a statistically significant influence (p < 0.01) on biomass production. In experiment No 702 production was ranging from 5.7 to 8.2 t ha-1 y-1, while in experiment No 707 was 1.6-2.8 t ha-1 y-1 (Fig. 3). Ranking of provenances was stable in both sites, with notably different growth conditions: the mean correlation among provenance was r = 0.74 (p < 0.01).
Fig. 3 - Above-ground biomass production vs. latitude of origin for different Pinus contorta provenances in Latvia.
Significant effect of genotype (clones, breeds) and soil conditions on above-ground biomass production has been detected also for other species grown or proposed for biomass production in the Baltic Sea region: Alnus glutinosa, Populus tremula, Salix spp., Populus spp. For example, two-fold differences in dry above-ground biomass production of Salix breeds on fertilized and unfertilized plots were detected in Estonia in trials with 4-year rotation period ([13]). Analysis of common aspen stands on former agricultural lands at the age from 5 to 24 years reveals notable variation of dry biomass production: 2.9 to 9.2 t ha-1 y-1, with the lowest productivity for stands growing on sandy soils ([15]). Depending on Salix breed, biomass production in 4-5 year rotation period on fertilized soils reaches from 8 to 14 t ha-1 y-1 in southern part of Sweden ([40]), with similar climatic conditions as in Latvia. In the same region, biomass production at the age of 6 years on former arable land for poplars was 7.5 t ha-1 y-1, for hybrid aspen, for grey alder 4.5 t ha-1 y-1 and for birch 2.3 t ha-1 y-1 ([35]). These figures are similar or lower than those found for Pinus contorta in experiment No 702. Above-ground biomass production in 4 to 40-years-old grey alder stands, typically located on fertile soils, in Scandinavian and Baltic States reaches 3-5 t ha-1 y-1 ([3], [30], [31], [16], [36], [26]). These figures are consistent with our results for Pinus contorta on poor, sandy soils (trials No 707 and No 82). Higher biomass production (4.5 to 10 t ha-1 y-1) was found for hybrid aspen and poplars, where regeneration by sprouting was carried out ([15], [35], [17], [29]). Productivity is mostly (but not always) higher also in Salix plantations, where fertilizer is used: for example, in Latvia and Lithuania it reaches 2.2-13 t ha-1 y-1 ([32], [19]). Results reveal that Pinus contorta has similar or higher productivity than other species recommended for establishment of biomass plantations in the Baltic Sea region, especially on poor, sandy soil where fertilization is not applied.
In the plantation biomass could be obtained not only by clear felling, but also by thinning. Results reveal that light thinning from below (according to current legislation in Latvia), leaving 1000 tallest trees ha-1, would provide on average 33.3 t ha-1 dry biomass in experiment No 707 and 61.8 t ha-1 biomass in experiment No 702. Biomass production of suppressed trees on poor sandy soil at the age of 22 years ranges from 1.4 to 2.6 t ha-1 y-1 (1.9 ± 0.20 t ha-1 y-1 on average) and could ensure notable income in thinning.
The analysis carried out failed to reveal any specific trend in productivity of Pinus contorta (biomass, height, diameter) in relation to the geographical location of provenance (latitude, longitude, altitude). Results are coherent with the earlier findings in Sweden from sites with similar climatic and photoperiodic conditions as in Latvia: best growing provenances originated from latitudes 51°-56° N, and no trend between provenance origin and its height growth was found ([11], [34]). Within this geographical range, individual seed source has an important impact on the plantation success (productivity, survival - [20]).
Conclusions
Currently, most of the biomass consumed in the growing number of modern district heating plants in cities and municipalities is supplied by logging residuals, but the share of wood chips from clearing of naturally over-grown abandoned agricultural lands is increasing. It is likely that this source will need to be replaced with wood from plantations in the future. Our results suggest that properly selected provenances of Pinus contorta can be the best alternative for the establishment of such plantations on poor soils, since the biomass is significantly (p < 0.01) and considerably (more than two-fold) higher than that of the native Pinus sylvestris. On fertile soils, establishment of plantations for biomass production is currently carried out at small scale by Populus spp., and Alnus incana (less than 30 ha in year 2011), but predominantly by Salix spp. clones (152 ha). Our results prove that Pinus contorta is a competitive alternative also in these conditions, producing similar amount of biomass (7.1 ± 0.40 t ha-1 y-1) and not requiring fertilization and intensive management.
Acknowledgements
This study was part of the European Social Fund’s project: “Importance of genetic factors on formation of forest stands with high adaptability and qualitative wood properties” (2009/0200/1DP/1.1.1.2.0/09/APIA/ VIAA/146).
References
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
U Neimane
J Rieksts-Riekstins
Latvian State Forest Research Institute “Silava”, Rigas Street 111, LV-2169 Salaspils (Latvia)
Forestry Faculty, Latvia University of Agriculture, Akademijas Street 14, LV-3001 Jelgava (Latvia)
Corresponding author
Paper Info
Citation
Jansons A, Sisenis L, Neimane U, Rieksts-Riekstins J (2013). Biomass production of young lodgepole pine (Pinus contorta var. latifolia) stands in Latvia. iForest 6: 10-14. - doi: 10.3832/ifor0637-006
Academic Editor
Marco Borghetti
Paper history
Received: Jul 13, 2012
Accepted: Oct 31, 2012
First online: Jan 14, 2013
Publication Date: Feb 05, 2013
Publication Time: 2.50 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2013
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 58959
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 48083
Abstract Page Views: 4481
PDF Downloads: 4820
Citation/Reference Downloads: 26
XML Downloads: 1549
Web Metrics
Days since publication: 4923
Overall contacts: 58959
Avg. contacts per week: 83.83
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2013): 7
Average cites per year: 0.54
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Technical Reports
Biomass equations for European beech growing on dry sites
vol. 9, pp. 751-757 (online: 17 June 2016)
Research Articles
Estimation of above-ground biomass and sequestered carbon of Taurus Cedar (Cedrus libani L.) in Antalya, Turkey
vol. 6, pp. 278-284 (online: 01 July 2013)
Research Articles
Estimation of forest biomass components using airborne LiDAR and multispectral sensors
vol. 12, pp. 207-213 (online: 25 April 2019)
Research Articles
Estimation of aboveground forest biomass in Galicia (NW Spain) by the combined use of LiDAR, LANDSAT ETM+ and National Forest Inventory data
vol. 10, pp. 590-596 (online: 15 May 2017)
Research Articles
Effects of planting density on the distribution of biomass in a douglas-fir plantation in southern Italy
vol. 8, pp. 368-376 (online: 09 September 2014)
Research Articles
Climate-wise models of biomass productivity for hybrid poplar clones in Europe
vol. 16, pp. 188-194 (online: 30 June 2023)
Research Articles
Methods for biomass stock estimation in Mediterranean maquis systems
vol. 10, pp. 108-114 (online: 21 August 2016)
Research Articles
Allometric equations to estimate above-ground biomass of small-diameter mixed tree species in secondary tropical forests
vol. 13, pp. 165-174 (online: 02 May 2020)
Research Articles
Growth, spring phenology and stem quality of four broadleaved species assessed in provenance trials in the Netherlands - Implications for seed sourcing
vol. 18, pp. 242-251 (online: 22 September 2025)
Research Articles
Above ground biomass estimation from UAV high resolution RGB images and LiDAR data in a pine forest in Southern Italy
vol. 15, pp. 451-457 (online: 03 November 2022)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







