
Moderate wildfire severity favors seed removal by granivores in a Mexican pine forest
iForest - Biogeosciences and Forestry, Volume 18, Issue 3, Pages 121-127 (2025)
doi: https://doi.org/10.3832/ifor4741-018
Published: May 24, 2025 - Copyright © 2025 SISEF
Research Articles
Abstract
Wildfires are critical to the ecological dynamics of communities in temperate forests. However, their frequency and severity have increased significantly in recent decades due to human activities, negatively affecting flora, fauna, and ecosystem processes. Among the primary interactions for the natural regeneration of forests is seed removal, which promotes secondary seed dispersal and predation. This study evaluated artificial seed removal under different wildfire severities in a pine forest on the eastern edge of the Trans-Mexican Volcanic Belt. An experiment of differential exclusion of vertebrates and invertebrates was carried out, considering two treatments: totally exposed seeds and seeds with exclusion of vertebrates in the unburnt forest, moderate and high severity fire. Our results showed a higher seed removal by vertebrates (birds and rodents) in the wildfire moderate severity treatment and in the time elapsed since wildfire occurrence; however, excluding vertebrates decreased the seed removal rate after the fire. Therefore, moderate-severity sites are an important factor in the communities of granivores that influence seed removal patterns in the temperate forest of Veracruz, Mexico. The findings underscore the importance of considering fire severity gradients in forest management strategies, particularly in fire-adapted ecosystems. As climate change alters fire regimes worldwide, understanding how different fire severities affect ecological interactions such as granivory is essential for promoting forest resilience and biodiversity.
Keywords
Artificial Seeds, Exclusion Experiments, Granivores, Natural Regeneration, Pine Forests, Predation
Introduction
Seed removal is a crucial process for recruiting new individuals in plant communities ([27]). It involves the relocation of seeds from one site to another by animal dispersers and accumulators. This process integrates both seed predation and secondary dispersal ([32]), contributing to the colonization of new sites, facilitating ecological succession ([19]), and enhancing genetic flow ([38]). However, seeds are susceptible to biotic and abiotic factors, such as predation and fire ([41]). Notably, fire influences recruitment rates and the establishment of various species, thereby shaping the composition of the plant communities ([34]).
Fires rapidly eliminate plant cover, including propagules, and negatively affect the abundance of seed-dispersal fauna ([6]). Additionally, fires cause habitat fragmentation, which may increase local extinction rates of both plants and animals ([2]). Although fires often have detrimental effects on ecosystems, their impact varies depending on fire severity ([26]). High-severity fires cause drastic changes in large mammal abundance and plant cover, create clearings and edge areas, and reduce the availability of shelter and forage for animals ([10]). Conversely, moderate-severity fires have a lesser impact on soil nutrients (N, P, K, and Mg), temperature, pH, and moisture, facilitating the reestablishing of herbaceous cover ([11]). The abundance of fauna, short and medium-term post-fire changes in resource availability, and vegetation structural characteristics are all influenced by fire severity and the time elapsed since the fire occurrence ([22]).
The primary seed removers in temperate forests are birds, rodents, and invertebrates ([32]). In the temperate zones of central Veracruz, small rodents such as Peromyscus spp. and Reithrodontomys spp., and birds such as Junco phaeonotus (Wagler, 1831), Tardas migratorius, Carpodacus mexicanus (P. L. Statius Müller, 1776), play a significant role in seed removal, accounting for up to 90% of the seeds that fall to the ground ([14]). In Mexico, wildfires are the third leading cause of biodiversity loss at the national level ([7]). Given the global increase in fire frequency and intensity and the ecological importance of this temperate forest ecosystem, it is imperative to evaluate wildfire effects not only on species diversity but also on ecosystem processes like seed removal. This process integrates seed predation and secondary seed dispersal, and is vital for seedling establishment and recruitment, which are critical phases in the natural post-fire regeneration of temperate forests ([21]).
Understanding seed removal patterns after fire is crucial both ecologically and for forest management, given the pivotal role seeds play in forest regeneration and ecosystem resilience. Fires, especially under changing climate conditions, can disrupt natural seed dispersal and predation dynamics, which are fundamental for recovering plant populations and maintaining biodiversity ([17]). In this context, this study aimed to evaluate seed removal under different wildfire severities in a Mexican temperate forest. It is hypothesized that: (i) seed removal will be higher with moderate fire severity compared to both unburnt forest and high-severity fire areas, due to vegetation recovery that supports the presence of granivores ([5]); (ii) vertebrates (birds and rodents) will exhibit higher seed removal rates because vegetation recovery provides protection against predators and allows granivores to expand their spatial range in search of food ([12]); (iii) the time elapsed since fire occurrence will affect seed removal by granivores ([22]).
Materials and methods
Site description
The study was conducted in the reserve San Juan del Monte reserve, a protected area dedicated to recreational and environmental education (Fig. 1a). The reserve covers 609.62 hectares on the northern slope of the Cofre de Perote volcano in Las Vigas de Ramírez, Veracruz, Mexico (19° 37′ 53.4″ N, 97° 07′ 00.1″ W; altitude 2700 m a.s.l. -[39]).
Fig. 1 - (A) Reserve for recreation and environmental education in San Juan del Monte, Veracruz, Mexico. (B) Spatial distribution of fire severity in the San Juan de Monte Reserve, Veracruz, Mexico. Surveyed sites: (a) unburnt forest (green square) without any damage from the wildfire; (b) moderate severity (light blue circle) the fire was limited to the lower layer (grasses, herbs and small shrubs), with trees showing green-yellow crown fractions; and (c) high severity (blue diamond) the tree, shrub and herb layers were burned with a layer of white ash.
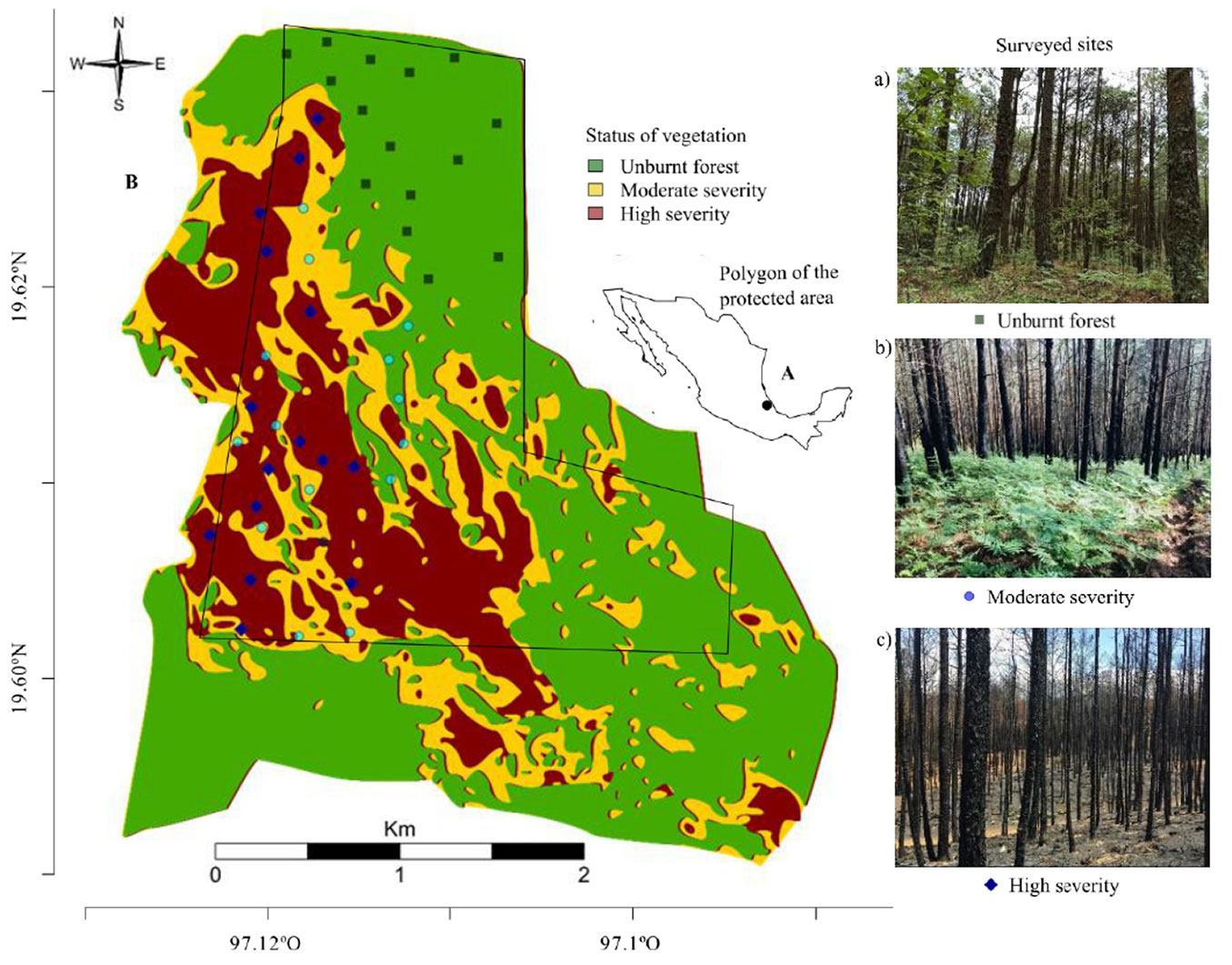
The area features heterogeneous topography, with steep hills in the southern region and plains in the north. Two forest types dominate the reserve: pine forests, primarily composed of Pinus teocote Schiede ex Schltdl, and mixed forests with Pinus patula Schl. et Cham, Pinus pseudostrobus Lindl, Quercus crassifolia Humb Bonpl, and Alnus jorullensis Kunth. In the shrub layer, dominant species include Baccharis conferta Kunth, Prunus serotina (Ehrh.), and Buddleia spp. L. The herbaceous stratum is primarily composed of Alchemilla pectinata Kunht, Archibaccharis androgyna (Brandegee) S.F. Blake, and Pteridium aquilinum (L.) Kunht. The reserve is also home to diverse wildlife, including 37 mammal species, 38 bird species, 17 reptile species, and 6 amphibian species ([16]).
Classifying wildfire severity
On March 11th, 2019, a wildfire broke out in the San Juan del Monte reserve and burned for five days, affecting approximately 350 hectares, more than 50% of the protected area (609.62 ha - Fig. 1b).
Satellite imagery and aerial photography were utilized to classify fire severity. Sentinel 2B satellite images from the European Space Agency (ESA - ⇒ https://scihub.copernicus.eu/dhus/#/home) and aerial photographs, taken four days after the fire (March 20, 2019) using a Mavic-Pro unmanned aerial vehicle (UAV) at an altitude of 50 meters, were analyzed. The UAV images, with a 3 cm/pixel resolution, were captured from five different locations.
Control points representing unburned forest, low, moderate, and high fire severity areas were extracted from the aerial images. Using RGB bands (band 2-4 from the satellite images), a spectral signature was created in ArcMap® v. 10.5 (ESRI Inc., 2016). A supervised maximum likelihood classification of the protected area was then performed ([9]). Each control point covered an area of 2500 m2 (5×5 pixels on the satellite image), with 68 control points: 15 for high severity, 15 for moderate severity, 22 for low severity, and 16 for unburned forest areas.
Classification accuracy was verified using n=30 independent points for each severity level and by constructing a confusion matrix. The overall classification error was low (< 8%), except for the low severity class, which had an error rate of 46% due to misclassification with unburned forest areas ([31]). The unburned forest also had a higher confusion rate (21%) with the low-severity class. Field visits were conducted to corroborate the classification results and account for the heterogeneous severity of wildfires.
Field measurements of severity indicators
Following Keeley ([20]), key biophysical indicators for fire severity were recorded at each sampling site. Unburned forest sites exhibited no fire damage, while moderate-severity sites showed trees with small portions of their canopies containing green-yellow needles and burnt stalks, but the soil’s organic layer remained mostly intact. High-severity sites, in contrast, were characterized by the complete destruction of trees, shrubs, and herbaceous layers, with deposits of white ash and charred organic matter.
Experimental design and seed removal treatments
In June of 2019, 45 plots were selected, including 15 unburnt forest sites, 15 with moderate fire severity, and 15 in areas of high severity. Each plot covered an area of 400 m2 and was separated by 230 meters to prevent overlap. All sites were within the elevation range of 2300-2500 meters.
To evaluate seed removal across wildfire severity levels, an exclusion treatment was implemented to restrict access to granivorous vertebrates while allowing the entry of invertebrates (e.g., ants). Control areas (without exclusion) were also established, permitting free access to all organisms ([8]). The exclusion treatment involved placing mesh cubes of 10 × 15 × 10 cm as confinement areas, with 0.5 cm mesh openings. These cubes were fixed to the ground using wooden stakes 30 cm long, buried approximately 10-15 cm deep. In contrast, the control treatment allowed unrestricted access to seeds by any organism in the forest.
Both treatments (exclusion and control) were placed in pairs, spaced three meters apart. In each treatment, 20 artificial seeds were placed on the ground within each of the 45 experimental plots (20 seeds × 45 plots, totaling 1800 seeds). Seeds were exposed for 72 hours after which the number of removed seeds was recorded, and the remaining seeds were retrieved to prevent further degradation. A viability and durability test was conducted under conditions similar to the study to address concerns about seed degradation due to prolonged environmental exposure. This test evaluated potential degradation indicators such as mold growth, mass loss, or decomposition. During the 72-hour exposure period, periodic checks revealed no significant signs of decomposition, physical damage, or structural loss in most cases. This procedure was repeated every three months from June 2019 to December 2020, aligning with periods of pine seed production (November-January) and of herbaceous/shrub seed availability (July-September in the rainy season, and January-April in the dry season - Tab. 1). A total of 11.160 artificial seeds were used during the study period.
Tab. 1 - Mean ± standard error of temperature and precipitation in the study area. Data correspond to 2019 and 2020 (21 months total). Temperature and precipitation data were obtained from the Las Vigas de Ramírez 30211 meteorological station (latitude 19° 37′ 53.4″ N, longitude 97° 07′ 00.1″ W) through the National Water Commission (CONAGUA - ⇒ http://www.conagua.gob.mx/).
| Seasonality | Months | Temperature (°C) | Precipitation (mm) | ||
|---|---|---|---|---|---|
| 2019 | 2020 | 2019 | 2020 | ||
| Dry season | October - February | 8.2 ± 3.2 | 7.1 ± 0.32 | 45.0 ± 32.5 | 62.6 ± 21.6 |
| Wet season | March - May | 27.0 ± 17.1 | 22.7 ± 15.6 | 55.0 ± 45.3 | 38.6 ± 8.4 |
| Water surplus | June - September | 15.0 ± 9.1 | 12.6 ± 0.29 | 250.0 ± 90.2 | 174.1 ± 63.8 |
Artificial seeds
The artificial seeds were prepared using 400 g of commercial wheat flour mixed with 200 ml of water ([1]). The mixture was combined in a 1L glass beaker (PIREX, USA) until a homogeneous consistency was achieved, kneaded into a firm dough, and rolled into a thin layer to dry for 30 minutes. Using a mold, artificial seeds were shaped to mimic natural seeds, with an average width of 3.74 ± 0.9 mm and an average height of 3.92 ± 0.04 mm ([23]).
Artificial seeds were chosen as a controlled experimental tool to study removal patterns without the influence of biological variability, such as germination, weight, size or color ([28]). Using artificial seeds offered several advantages: (i) they provide a consistent reference for testing ecological hypotheses in diverse environments, (ii) their uniformity minimized variability unrelated to seed removal interactions, and (iii) unintentional plant establishment leading to disruptions to the local ecosystem can be avoided ([24]). Although artificial seeds lack biological complexity, this trade-off was deemed acceptable for the scope of this study ([30]). Tab. 2highlights the benefits of using artificial seeds to control experimental variables and ensure reproducibility.
Tab. 2 - Specific advantages and disadvantages of artificial seeds that allow isolating key factors in the study and minimizing sources of unwanted variability.
| Advantages | Disadvantages |
|---|---|
| Artificial seeds can be designed with consistent size, shape, weight, and texture, allowing to systematically study the effects of specific variables (e.g., dispersal patterns) without the natural variability inherent in real seeds. | Artificial seeds do not replicate the biochemical properties of real seeds, such as the production of chemical signals (e.g., secondary metabolites) or interactions with soil microorganisms. |
| By eliminating biological factors such as genetic variability or differences in moisture content, artificial seeds ensure that experimental results are comparable across replicates and independent trials. | Results obtained using artificial seeds may not fully represent ecological interactions involving real seeds, especially when predator or disperser behavior is influenced by chemical or biological cues. |
| Real seeds are subject to germination, decay, or other biological processes that could interfere with the research objectives, especially in long-term studies. Artificial seeds remain stable, enabling prolonged experimentation. | Depending on the design, creating artificial seeds with specific properties can involve additional costs in terms of materials or fabrication processes. |
Data analysis
To evaluate the effects of time since the fire, treatments (control/exclusion), fire severity, and their interactions - time since the fire × fire severity, time since the fire × treatment, and fire severity × treatment - a mixed-effects binomial model was used. The response variable was the proportion of seeds removed, while the explanatory factors included time since the fire, treatment, and fire severity, as well as their pairwise interactions. Given that the study spanned 21 months and utilized 45 experimental plots, the identity of each plot was included as a random factor to account for temporal variations. Additionally, another mixed-effects binomial model was applied to assess the effect of the year (2019 vs. 2020) and fire severity on seed removal, using the same response variable and random factor.
The analyses were conducted in R ([29]) using the “glmer” function from the “lme4” package ([4]). Pairwise comparisons between treatments were performed using contrast methods, with standard errors estimated from the model’s linear predictors. These contrasts were implemented using the “glht” function of the “multcomp” package ([18]).
Results
From the total of 11.160 seeds used in this study, 55% were removed. Of these, 53% were removed in the control treatment, with 40% of these removals occurring in areas of moderate severity. The results showed significant effects of time since the fire, treatment type (control/exclusion), fire severity, and their interactions: time since fire × fire severity, time since fire × treatment, and fire severity × treatment, all effects were statistically significant (p < 0.05 - Tab. 3).
Tab. 3 - Summary of the effects of time since fire, treatment (exclusion/control), fire severity (unburnt forest, moderate and high severity), and their two-way interactions on artificial seed removal. (df): degrees of freedom.
| Effects | χ2 | df | p-value |
|---|---|---|---|
| Time since fire (TSF) | 42.2 | 6 | < 0.001 |
| Exclusion | 101.8 | 1 | < 0.001 |
| Fire severity | 9.7 | 2 | < 0.001 |
| TSF: Exclusion | 140.5 | 6 | < 0.001 |
| TSF: Severity | 44.7 | 12 | < 0.001 |
| Exclusion: Severity | 53.3 | 2 | < 0.001 |
Time and severity of the fire
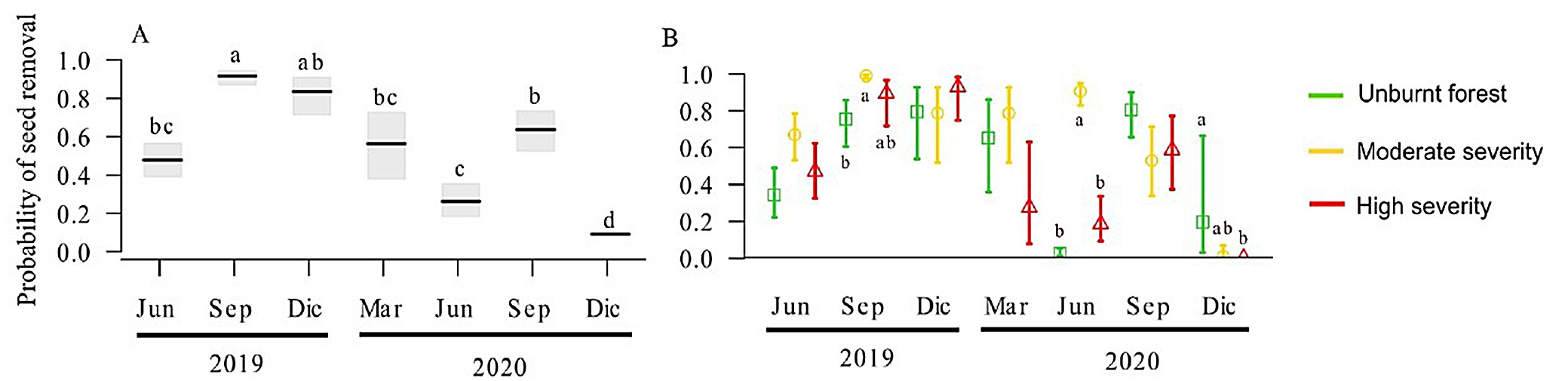
Seed removal rates varied significantly over time. The highest rates of seed removal occurred in September and December 2019 and September 2020, while the lowest rate was observed in December 2020 (P < 0.05 - Fig. 2a). Moderate fire severity consistently showed the highest seed removal rates, particularly in September 2019 and June 2020 (X2 =115, df =8, P < 0.001 - Fig. 2b). By December 2020, seed removal rates in unburned forest were significantly higher compared to those in high-severity areas (Fig. 2b).
Fig. 2 - Post-fire seed removal in March 2019 in the San Juan del Monte reserve, Las Vigas, Veracruz, Mexico. (A): Seed removal between 3 and 21 months after fire. (B): Seed removal between 3 and 21 months after fire in unburnt forest (green), moderate (yellow), and high severity (red). Different letters indicate significant differences (p < 0.05).
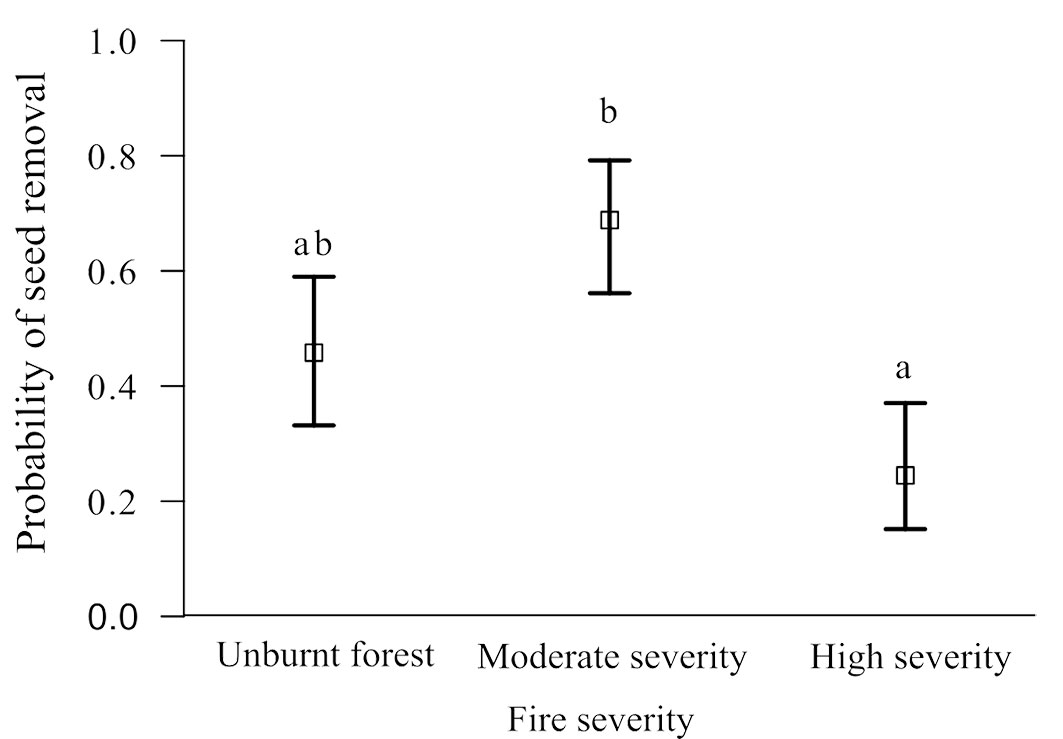
In the overall analysis, areas with moderate fire severity exhibited significantly higher seed removal rates compared to the areas of high severity. However, seed removal rates in unburned forest areas were intermediate and did not significantly differ from those in moderate and high-severity areas (P > 0.05 - Fig. 3).
Fig. 3 - Post-fire seed removal rate in forest with different fire severity. Different letters indicate significant differences (p < 0.05).
Treatments and severity of the fire
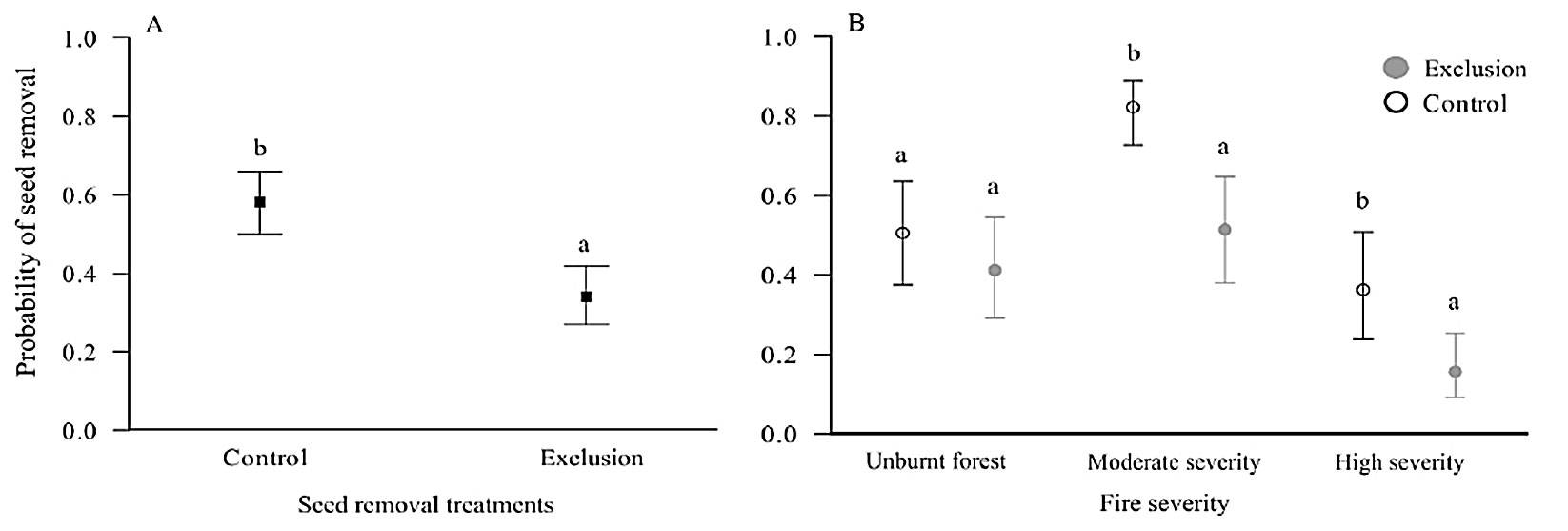
The exclusion treatment reduced the seed removal rate by 46% compared to the control (χ2 = 99, df = 1, p < 0.001 - Fig. 4a). This effect was observed in areas of moderate and high severity, but not in the unburnt forest (χ2 = 43, df = 2, p < 0.001 - Fig. 4b).
Fig. 4 - (A) Vertebrate exclusion experimental treatment. (B) Experimental vertebrate exclusion treatment, control (open circle) and exclusion (solid circle), in unburnt forest, moderate and high severity. Different letters indicate significant differences (p < 0.05).
Time with treatments
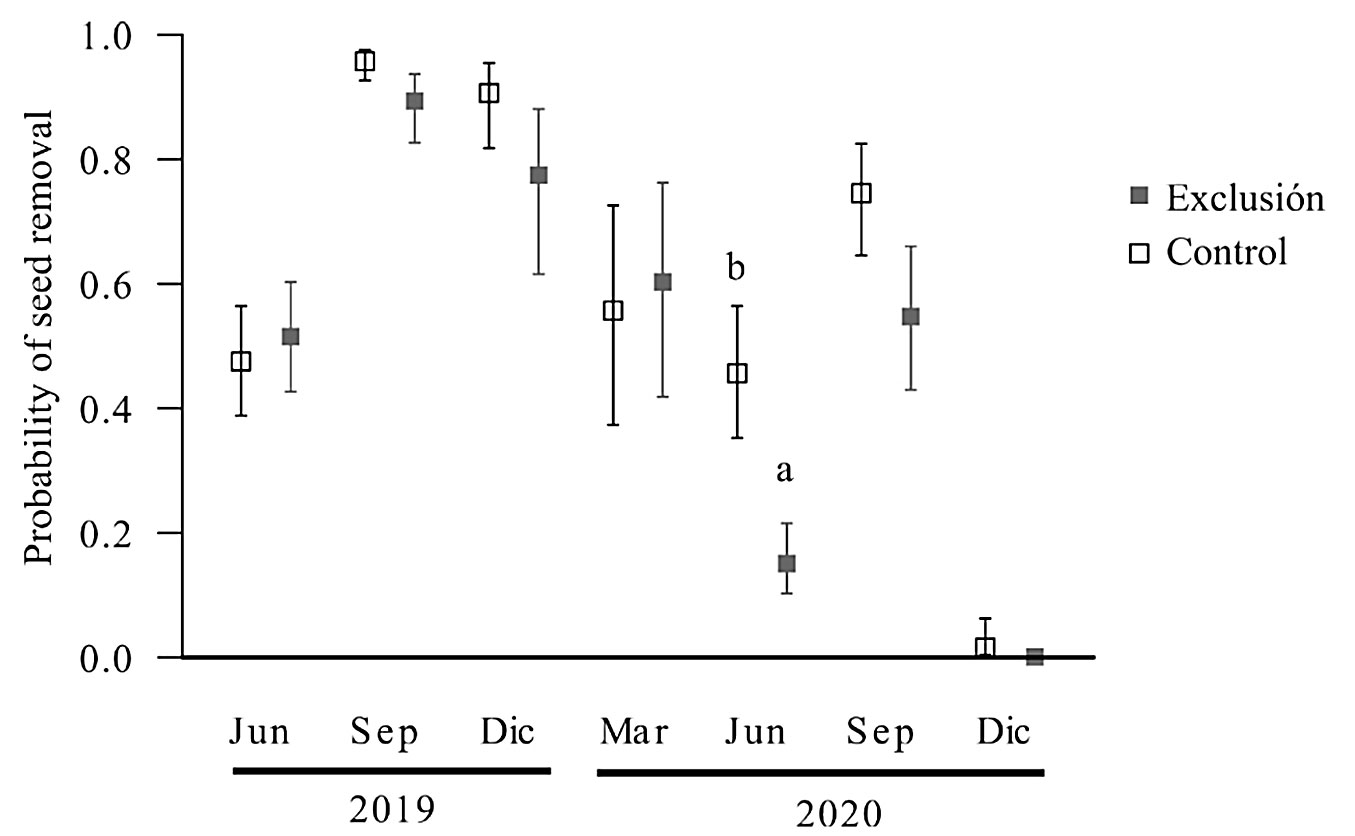
The interaction between the control/exclusion treatment and time since the fire showed significant differences only in June 2020, when the control treatment exhibited a higher seed removal rate than the exclusion treatment (χ2 = 122, df = 6, p < 0.001 - Fig. 5).
Fig. 5 - Seed removal after the March 2019 fire in the San Juan del Monte reserve, Las Vigas, Veracruz, Mexico. Experimental vertebrate exclusion treatment between 3 and 21 months post-fire. Different letters indicate significant differences (p < 0.05).
Years and severity of the fire
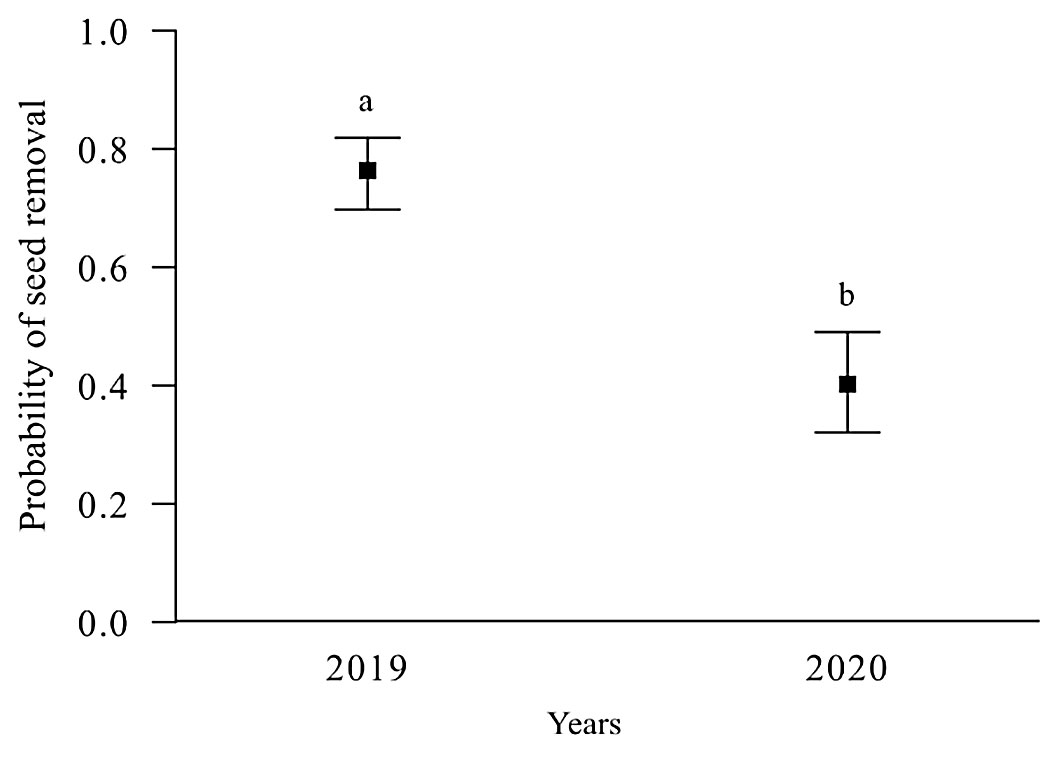
The mixed-effects model revealed significant differences between years (χ2 = 12, df = 1, p < 0.001 - Fig. 6). Seed removal rates were higher in June and December 2019 compared to March-December 2020. However, no significant differences were found between unburned forest, moderate severity and high severity areas (p > 0.05).
Fig. 6 - Seed removal following the March 2019 fire in the San Juan del Monte reserve, Las Vigas, Veracruz, Mexico in 2019 and 2020. Different letters indicate significant differences (p < 0.05).
Discussion
This study evaluated the effects of fire severity on seed removal activity and the potential removal of natural seeds through a vertebrate and invertebrate exclusion experiment. Results reveal that moderate fire severity significantly influenced seed removal patterns 21 months after the San Juan de Monte reserve wildfires. Differences in vegetation structure, fire severity, and the time since the fire all affected seed removal rates and the organisms involved. Vertebrates were particularly critical in seed removal in the moderate-severity areas during the first year after the wildfire.
Wildfires modify vegetation structure, which in turn impacts granivorous-mediated seed removal ([36]). Less affected areas may act as refugees for regeneration, influenced by variations in temperature, humidity, and disperser activity. This aligns with prediction (i) of this study, as rapid vegetation recovery in moderate-severity areas, dominated by P. aquilinum (fern) and Muhlenbergia macroura L. (grass), Arbutus xalapensis (Kunth), and Baccharis conferta (Kunth) Hitchc. (shrub)- created micro-habitats with specific conditions (temperature, light, and moisture) favorable to granivores like Reithrodontomys sp., Dendroica occidentalis (J. K. Townsend, 1837), Wilsonia pusilla (A. Wilson, 1811), and Turdus grayi (Bonaparte, 1838), among others ([13], [31]).
Greater seed removal in areas affected by moderate fire severity can significantly influence the initial regeneration and successional trajectories of the forest. This process favors less-consumed species, enhances biodiversity, and accelerates the return to a mature forest state ([10]). While low-severity fires generally do not cause substantial changes to flora and fauna, enabling complete seed removal. Moderate fire severity negatively impacts large mammals, soil temperature, and pH levels. However, it also increases the availability of key nutrients (e.g., phosphorus, potassium, and magnesium) while preserving seed trees and supporting small mammals. These conditions allow for seed removal, seedling recruitment, and the continuation of forest regeneration and successional processes. On the other hand, high-severity fires severely disrupt seed trees, seed banks, organic matter, soil moisture, depth, and nutrient availability, thereby impairing seed availability, seed removal, and seedling establishment ([11]).
These findings are consistent with other studies conducted in temperate forests, which indicate that vertebrates, particularly birds and rodents, are the primary agents of seed removal ([5], [12]). Granivorous rodents display foraging patterns closely linked to vegetation compositions ([5], [12]). For example, Peromyscus melanotis and Mus musculus Linnaeus exhibit foraging behaviors influenced by vegetation composition, moving between fragments based on disturbance levels and resource availability ([13], [15]). Dense vegetation, such as that found in moderate-severity areas, provides cover that reduces predation risk, while open areas (e.g., high-severity sites) expose granivores to predators like eagles, owls, and snakes ([3])
Granivorous birds also contribute significantly to seed removal, with their mobility and use of vegetation as perch sites enhancing their foraging efficiency ([40]). For example, the ability to fly enables birds to move rapidly between different forage zones and cover greater distances by utilizing foliage profiles that serve as perch sites ([15]). In this study, moderate-severity sites were characterized by trees with green-yellow canopies and herbaceous vegetation. In contrast, high-severity sites were destroyed and characterized by completely charred trees, shrubs, and herbaceous vegetation. Additionally, Flores-Peredo ([13]) reported in the same study area that granivorous birds, such as Junco phaeonotus (Wagler, 1831), Tardas migratorius, Carpodacus mexicanus (P. L. Statius Müller, 1776), Spizella passerine (Bechstein, 1798), and Poocetes gramineus, play a fundamental role in seed removal and dispersal, primarily during the day. This observation could explain the higher seed removal values found in moderate-severity areas.
The time elapsed since the fire began influenced seed removal, supporting prediction (iii). Overall, the seed removal rate was higher in 2019 (September). Post-fire fire changes in plant coverage lead to decreased food resources and altered activity patterns as birds must forage at a greater distance from their nests ([33]). Conversely, when serotine cones open and release seeds due to the fire effect, the presence and abundance of granivorous birds are favored. In contrast, the nectarivorous, insectivorous, and frugivorous birds decrease ([25]). It was further observed that the recovery and restructuring of the plant community mediates the recovery of granivorous communities and their levels of seed removal. Factors such as understory plant density, herbaceous coverage, and number of mammals synergistically influenced resource availability within the ecosystem; these dynamics contribute to the lower seed removal observed in 2020 ([37]).
Seed removal is a critical ecological process influencing plant regeneration and ecosystem resilience. However, this process is increasingly vulnerable to the combined pressures of climate change and shifting fire regimes ([35]). These global phenomena alter the interactions between plants and animals, mediating seed dispersal and predation, with profound implications for forest dynamics, biodiversity, and ecosystem recovery. The interplay between climate change, fire regimes, and seed removal is a complex and pressing issue. These forces disrupt the delicate balance of ecosystems, modifying habitats and altering the behavior, distribution, and abundance of species involved in seed removal. Such disruptions hinder forest regeneration and threaten biodiversity ([17]). Addressing these challenges requires innovative research and conservation strategies to prevent forest resilience and the vital ecological processes that forests support.
Conclusions
Our results demonstrated higher seed removal by vertebrates (birds and rodents) in areas affected by moderate fire severity. These findings highlight the significant role of moderate fire severity in shaping granivorous communities and influencing seed removal patterns in the temperate forest of Veracruz, Mexico. Artificial seeds proved a valuable methodological approach, particularly in scenarios where fruiting seasons are disrupted due to the severity and intensity of fires, or where there is limited knowledge of local fruiting plant species. While the absence of natural seeds somewhat constrains this method, it offers a broader understanding of ecosystem functioning patterns by minimizing seed preference and size biases. This approach provides critical insights into the ecological processes that drive forest recovery and resilience in the face of environmental disturbances.
Author Contributions
DACZ: Conceptualization, Data curation, Formal analysis, Writing - original draft. JCI, JFAM, JDCZ: Supervision, Writing - review and editing. JLAV, LABG, SAS, JFDN: Supervision, Writing - review and editing. DACZ: Funding acquisition, Project administration. DACZ: Writing - review and editing.
Funding
This work was supported by the Veracruz Council for Scientific Research and Technological Development (COVEICYDET-8.8 2SO 2020) and a CONAHCYT grant awarded to DACZ (609138).
Data Availability Statement
The data presented in this study are available upon request. Due to privacy concerns, they are not publicly available.
Acknowledgements
We thank Ismael Guzman for their invaluable assistance in field campaigns and sorting samples. Diana Ramírez provided us with remote sensing maps of the affected area. A special thanks to SEDEMA-Xalapa for the logistic support and lodging facilities in the SJM Protected Area.
Declaration of Interest
The authors declare that they have no known competing financial interests or personal relationships that could influence the work reported in this paper.
References
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
CrossRef | Gscholar
Authors’ Info
Authors’ Affiliation
Department of Forest Ecology, Interdisciplinary Research Group at Sechium edule in México (GISeM), Agustín Melgar 10, Texcoco, 56160, México (Mexico)
Department of Botany, Colegio de Postgraduados, Iturbide 73, 78622, San Luis Potosí (Mexico)
Department of Plant Phisiology, Universidad Autónoma de Chiapas, Entronque Carretera Costera y Pueblo de Huehuetán, 30660, Chiapas (Mexico)
Department of Forestry Genetic Resources, Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias, Boulevard de La Biodiversidad 400, 47600, Jalisco (Mexico)
Department of Forestry Management and Agroecology, Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias, Carretera Iguala-Tuxpan, 40000, Guerrero (Mexico)
Department of Sustainable Ecosystems, Universidad Autónoma Chapingo, Carretera Federal Huatusco-Xalapa Km. 6.5, 94100, Veracruz (Mexico)
José F Díaz-Nájera 0000-0001-7181-9425
Department of Forestry Management, Colegio Superior Agropecuario del Estado de Guerrero, 40000, Guerrero (Mexico)
Corresponding author
Paper Info
Citation
Cadena-Zamudio DA, Cadena-Iñiguez J, Aguirre-Medina JF, Cadena-Zamudio JD, Arispe-Vázquez JL, Barrera-Guzmán LA, Ayvar-Serna S, Díaz-Nájera JF (2025). Moderate wildfire severity favors seed removal by granivores in a Mexican pine forest. iForest 18: 121-127. - doi: 10.3832/ifor4741-018
Academic Editor
Marco Borghetti
Paper history
Received: Oct 06, 2024
Accepted: Feb 06, 2025
First online: May 24, 2025
Publication Date: Jun 30, 2025
Publication Time: 3.57 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2025
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 6859
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 2447
Abstract Page Views: 2067
PDF Downloads: 2102
Citation/Reference Downloads: 0
XML Downloads: 243
Web Metrics
Days since publication: 414
Overall contacts: 6859
Avg. contacts per week: 115.97
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
(No citations were found up to date. Please come back later)
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Review Papers
Problems and solutions to cork oak (Quercus suber L.) regeneration: a review
vol. 16, pp. 10-22 (online: 09 January 2023)
Research Articles
Short- and long-term natural regeneration after windthrow disturbances in Norway spruce forests in Bulgaria
vol. 11, pp. 675-684 (online: 23 October 2018)
Research Articles
The impact of seed predation and browsing on natural sessile oak regeneration under different light conditions in an over-aged coppice stand
vol. 9, pp. 569-576 (online: 04 April 2016)
Research Articles
Seed trait and rodent species determine seed dispersal and predation: evidences from semi-natural enclosures
vol. 8, pp. 207-213 (online: 28 August 2014)
Research Articles
Post-fire effects and short-term regeneration dynamics following high-severity crown fires in a Mediterranean forest
vol. 5, pp. 93-100 (online: 30 May 2012)
Research Articles
Post-fire recovery of Abies cephalonica forest communities: the case of Mt Parnitha National Park, Attica, Greece
vol. 11, pp. 757-764 (online: 15 November 2018)
Research Articles
Post-fire recovery of the plant community in Pinus brutia forests: active vs. indirect restoration techniques after salvage logging
vol. 11, pp. 635-642 (online: 04 October 2018)
Research Articles
Modelling natural regeneration of Oak in Saxony, Germany: identifying factors influencing the occurrence and density of regeneration
vol. 16, pp. 47-52 (online: 16 February 2023)
Research Articles
Methods for predicting Sitka spruce natural regeneration presence and density in the UK
vol. 12, pp. 279-288 (online: 23 May 2019)
Research Articles
Stand dynamics and natural regeneration in silver fir (Abies alba Mill.) plantations after traditional rotation age
vol. 7, pp. 313-323 (online: 08 April 2014)
iForest Database Search
Google Scholar Search
Citing Articles
Search By Author
- DA Cadena-Zamudio
- J Cadena-Iñiguez
- JF Aguirre-Medina
- JD Cadena-Zamudio
- JL Arispe-Vázquez
- LA Barrera-Guzmán
- S Ayvar-Serna
- JF Díaz-Nájera
Search By Keywords







