
Assessing the environmental factors affecting the natural regeneration of Cedrus libani A. Rich. in Jawbat Burghal, Syria
iForest - Biogeosciences and Forestry, Volume 19, Issue 3, Pages 237-243 (2026)
doi: https://doi.org/10.3832/ifor4587-018
Published: Jun 14, 2026 - Copyright © 2026 SISEF
Research Articles
Abstract
Remote Sensing (RS) and Geographic Information Systems (GIS) technologies are valuable tools for managing forest ecosystems today. This study employed a Binary Logistic Regression Model to analyze the environmental factors influencing the natural regeneration of Cedrus libani A. Rich. within its native habitat in the Jawbat Burghal forest of Syria. The effects of various environmental factors (spatial and forest structure, topography, and spectral indicators) were assessed both individually and collectively to understand their impact on the probability of C. libani seedling presence in Jawbat Burghal. The model’s accuracy significantly improved when considering the combined effects of these factors, reaching 94.3%, exceeding the accuracy of the other models evaluated (none of which surpassed 85%). Higher stone density, increased broadleaved tree cover, and favorable summer soil moisture were found to promote the natural regeneration of C. libani in Jawbat Burghal.
Keywords
Natural Regeneration, Environmental Factor Modeling, Remote Sensing, Geographic Information Systems (GIS), Forest Structure, Cedrus libani
Introduction
Native populations of Lebanon cedar (Cedrus libani A. Rich.) are confined to the Eastern Mediterranean region, where they once formed extensive and magnificent forests in Syria, Lebanon, and Turkey, and were an important source of wood for successive human civilizations ([31]). Today, only small and fragmented forest populations remain as a result of more than 5.000 years of intensive human exploitation, mainly through logging, burning, and goat grazing ([7]). These populations have been genetically isolated since the last Quaternary glacial cycle; they have been threatened with extinction since the 19th century ([10]) and have been classified in national records as endangered, threatened, and at risk of extinction ([26]). In Syria, due to increasing human pressure, C. libani populations are now confined to a highly restricted area in the northern part of the Syrian coastal mountain range ([27]).
Currently, Syrian C. libani forests are degraded and fragmented, found only in the northwestern regions of Slenfeh and Jawbat Burghal ([25]). These remaining forests suffer from a severe lack of natural regeneration, threatening their long-term sustainability ([35], [18]).
Regeneration is a multiphase process involving several sequential life history stages (i.e., seeds, seedlings, saplings) connected by transitional processes (i.e., pre-dispersed seed, dispersed seed, germination, emergence, survival) which are affected by various environmental factors (e.g., litter, drought, diseases, insects, etc.) ([15]). Many environmental factors govern the success of the regeneration process, especially within microsites, defined by environmental characteristics at the spatial scale of individual seeds or seedlings ([45]). Seedling establishment is the critical stage in a plant’s life history, during which the survival and growth of the tree species are more sensitive to microenvironmental site conditions ([17]).
Satellite imagery at various spatial and temporal scales plays a vital role in studying dynamic phenomena such as natural regeneration following disturbances (climate change, fires, land-use change, volcanoes, etc.) over specific periods using vegetation biomass or remote sensing indices ([8]). Remote sensing technology has been used to monitor natural regeneration patterns in forests after fires, particularly shrub cover regeneration in the Mediterranean region ([19]).
The study aims to: (i) assess the current state of C. libani natural regeneration in its native habitat in Syria (Jawbat Burghal); (ii) identify the potential causes of regeneration weakness by developing a mathematical model to evaluate the effects of spatial and forest structure, topographic variables, and spectral indicators; (iii) predict the likelihood of C. libani occurrence in Jawbat Burghal based on environmental factors influencing its natural regeneration.
Material and methods
Study area
This study was conducted in a specific region within the natural distribution range of Cedrus libani, located in the Jawbat Burghal area of Syria’s Lattakia Governorate. The site, officially designated as the “Cedar and Fir Protected Area” since 1996, lies between 36° 10′ and 36° 17′ E and 35° 29′ and 35° 41′ N, on the eastern slopes of the northern Syrian coastal mountain range, encompassing the Slenfeh and Jawbat Burghal regions.
In Syria, C. libani naturally grows in soils developed over hard Jurassic limestone formations, composed primarily of limestone and dolomite, both calcareous rocks ([32]). These cedar forests are confined to the cold perhumid bioclimatic zone on the eastern slopes of the coastal mountain range, at elevations ranging from 900 to 1.562 m a.s.l. A detailed study by Ibrahem et al. ([22]) accurately mapped the distribution of C. libani in Syria, identifying 35.29 ha in Slenfeh and 219.43 ha in Jawbat Burghal, totaling 254.72 ha of C. libani stands across both these regions.
Image acquisition and processing
Image data used in this study were obtained from the United States Geological Survey (USGS) Earth Explorer Landsat archive (⇒ http://earthexplorer.usgs.gov/). A Landsat 8 OLI_TIRS Level 1 image, systematically and terrain-corrected (L1T/G), was acquired on August 24, 2014, with a spatial resolution of 30 m. The study site falls within Path 174, Row 35 of the World Geographic System (WGS). The image was geometrically rectified to the Universal Transverse Mercator (UTM) coordinate system, Zone 37 N, using the WGS 1984 Datum. Top-of-atmosphere (TOA) correction was applied to convert raw image data into TOA reflectance values ([44]).
A Digital Elevation Model (DEM) with a 30 m resolution was used to derive topographic variables for the study area. All satellite images were resampled to a 5 m resolution using bilinear interpolation to match the spatial resolution of the RapidEye 532 high-resolution image, acquired on June 24, 2011. Supervised classification of the RapidEye image was performed to estimate the cover of Cedrus libani, broadleaved trees, and open areas (vegetation cover < 30%) within the study site.
Field survey
In summer 2014, field surveys were conducted to evaluate natural regeneration using a random sampling method. A total of 198 sample plots were distributed along the eastern slopes of the Syrian coastal mountain range in the Jawbat Burghal region. Each plot measured 5 × 5 m and was used to record data on trees, shrubs, saplings, and seedlings.
The geographic coordinates of the sample plots were recorded using a Global Positioning System (GPS) and converted into a GIS point data layer. Within each plot, the presence or absence of Cedrus libani seedlings (height < 50 cm) was recorded, as well as their density. Additionally, the number of Cedrus saplings (height ≥ 50 cm and ≤ 130 cm) and seedlings of other forestry species were registered.
For C. libani seedlings, additional measurements were taken, including height, estimated age, vitality, and microsite conditions (exposed, semi-exposed, shaded, or under the parent tree’s crown).
Data preparation for the models
The impact of environmental variables on C. libani natural regeneration was recorded across 198 plots in Jawbat Burghal, which were divided into 3 groups (see below).
Spatial and forest structure variables
Type of predominant forest cover (type): type1, C. libani; type2, conifers (except cedar); type3, Quercus spp.; type4, broadleaved (except oaks); type5, mixed forest stands; type6, open area. The tree coverage (cover, in %). Type of predominant shrub cover (shrub_type): shrub_type0, no shrubs; shrub_type1, C. libani shrubs; shrub_type2, conifers shrubs (except cedar); shrub_type3, Quercus spp.; shrub_type4, broadleaved shrubs (except oaks); shrub_type5, mixed shrub species. The shrub coverage (shrub, in %). The litter thickness (litter_thick). The litter type (litter_type): litter_type1, conifers; litter_type2, broadleaved; litter_type3, mixed conifer-broadleaved litter. The stone size (stone_size): stone_size1, small; stone_size2, medium; stone_size3, large; stone_size4, varied size. The stone coverage (stone, in %) and the fruiting intensity (fruit), according to the number of cones on the tree: (0) cones absent; (1) fewer than 20 cones; (2) number of cones between 20 and 50; (3) number of C. libani cones > 50 cones. The distance from mature cedar trees (distance_cedar), the distance from broadleaved trees (distance _broad), and the distance from forest roads (distance_ roads) were calculated in ArcGIS® v. 9.3 using Euclidean distances in Spatial Analyst Tools.
Topographic variables
The DEM was used to derive the following topographic variables layers in ArcGIS 9.3 using Spatial Analyst Tools. The elevation was classified manually into 10 classes in meters: (570-900), (900-950), (950-1000), (1000-1050), (1050-1100), (1100-1150), (1150-1200), (1200-1250), (1250-1300) and (1300-1350). The slope was divided into 5 classes (in degrees): slope1 (0-10), slope2 (10-20), slope3 (20-30), slope4 (30-40), and slope5 (40-50). The aspect was divided into 9 classes (in degrees): north (0-22.5), northeast (22.5-67.5), east (67.5-112.5), southeast (112.5-157.5), south (157.5-202.5), southwest (202.5-247.5), west (247.5-292.5), northwest (292.5-337.5), and north (337.5-360). The curvature of the earth-shaped layer was divided into 3 classes: 0, flat ground; 1, concave shape; 2, convex shapes. The hillshade layer was divided into two layers: 0 - shade, which is values less than 127° (shade), and 1 - light, which is values greater than 127° (light). The solar radiation layer was divided into 9 classes by using the natural break method (Jenks). The eastness, which is the east-west trend (sinaspect), and cell values range from -1 to +1 ([5]). The northness, which is the north-south trend (cosaspect), and cell values range from -1 to + 1 ([5]).
Remote sensing indicators
Satellite data from Landsat 8 were used to derive the following remote sensing indicators. The Normalized difference vegetation index (NDVI) was computed by the following equation ([43] - eqn. 1):
The Normalized Difference Water Index (NDWI), calculated as follows ([12] - eqn. 2):
The Normalized Difference Moisture Index (NDMI), which was computed as ([46] - eqn. 3):
The Modified Soil Adjusted Vegetation Index (MSAVI), which is defined by the following equation ([38] - eqn. 4):
The Disturbance Index (DI) was calculated as ([3] - eqn. 5):
while the simple ratio (SR) was obtained as follows ([28] - eqn. 6):
and the structure index (SI) using the equation ([11] - eqn. 7):
where NIR is the near infrared spectral wavelength, R is the red spectral wavelength, G is the green spectral wavelength, and MIR is mid-infrared spectral wavelength. The soil moisture index (SMI) was computed using the following equation ([29] - eqn. 8):
where LSTmax = a1 × (NDVI) + b1, LSTmin = a2 × (NDVI) + b2; LSTmax is the maximum earth surface temperatures corresponding to the value of the NDVI considered, LSTmin is the minimum earth surface temperatures corresponding to the value of the NDVI considered, LST is the surface temperature at the target pixel corresponding to the value of the NDVI considered; a1, a2 are the slope coefficients in the linear regression model representing the relationship between surface temperature and NDVI, and b1, b2 are constants in the linear regression model.
All final GIS database layers were in raster format. After preparing these layers, the sample plot locations were overlaid on the maps to extract the corresponding predictor variables for further analysis (see Supplementary material).
Data analysis
In order to model the effects of environmental variables on the natural regeneration of Cedrus libani in the Jawbat Burghal region, all examined spatial, forest structure, topographic, and spectral variables were treated as continuous, except for the following, which were considered categorical: dominant forest type, shrub cover, litter type, stone size, slope, aspect, hillshade, and curvature. Binary Logistic Regression (BLR) analysis was performed in SPSS® v. 20 (IBM Corp., Armonk, NY, USA) using a backward-elimination algorithm based on the maximum-likelihood ratio test. Raster-based remote sensing data were converted into ASCII format and subsequently imported into SPSS for statistical modeling of the presence or absence of C. libani seedlings.
The logistic regression model is expressed in the equation ([30] - eqn. 9):
where P is the estimated probability of occurrence of the dependent variable; its value ranges between 0 and 1, and the cutoff value is 0.5. Z is the linear relationship between the predictor variables that take values from -∞ to +∞; its values are given as follows (eqn. 10):
where b0 is the constant logistic regression model, b1-bk are slope coefficients in the logistic regression model, and X1-Xk are independent variables.
A multicollinearity test was conducted to ensure the independence of predictor variables and eliminate redundancy prior to model implementation. Variables with a Variance Inflation Factor (VIF) > 10 were excluded from the model to enhance the reliability of parameter estimates and overall model performance
Data evaluation
The significance of the logistic regression model was assessed using the Omnibus χ2 test at a 5% significance level. Model performance was evaluated through several key indicators: Nagelkerke R2, which reflects the proportion of variance in the dependent variable explained by the independent variables; the Hosmer-Lemeshow test, where a p-value greater than 0.05 indicates good model calibration and fit; overall classification accuracy, determined based on a specified cut-off value (e.g., 0.05); and the ROC (Receiver Operating Characteristic) curve along with AUC (Area Under the Curve) values, which range from 0.5 to 1.0.
The relationship between AUC values and model prediction accuracy can be interpreted as follows: values between 0.9 and 1.0 indicate excellent performance; between 0.8 and 0.9, very good; between 0.7 and 0.8, good; between 0.6 and 0.7, moderate; and between 0.5 and 0.6, poor or weak performance ([47]).
Results and discussion
Current status of natural regeneration of C. libani
The mean density of Cedrus libani seedlings in the Jawbat Burghal forest was 11.75 individuals per 25 m2, equivalent to approximately 4.700 individuals per hectare, with a presence probability of 71% (Fig. 1). Seedling age ranged from 1 to 5 years, with an average of 3 years, while height varied between 10 and 45 cm, averaging 25 cm. Saplings were aged between 15 and 30 years, with heights ranging from 50 to 250 cm.
Fig. 1 - Sample plots of natural regeneration of C. libani in Jawbat Burghal.
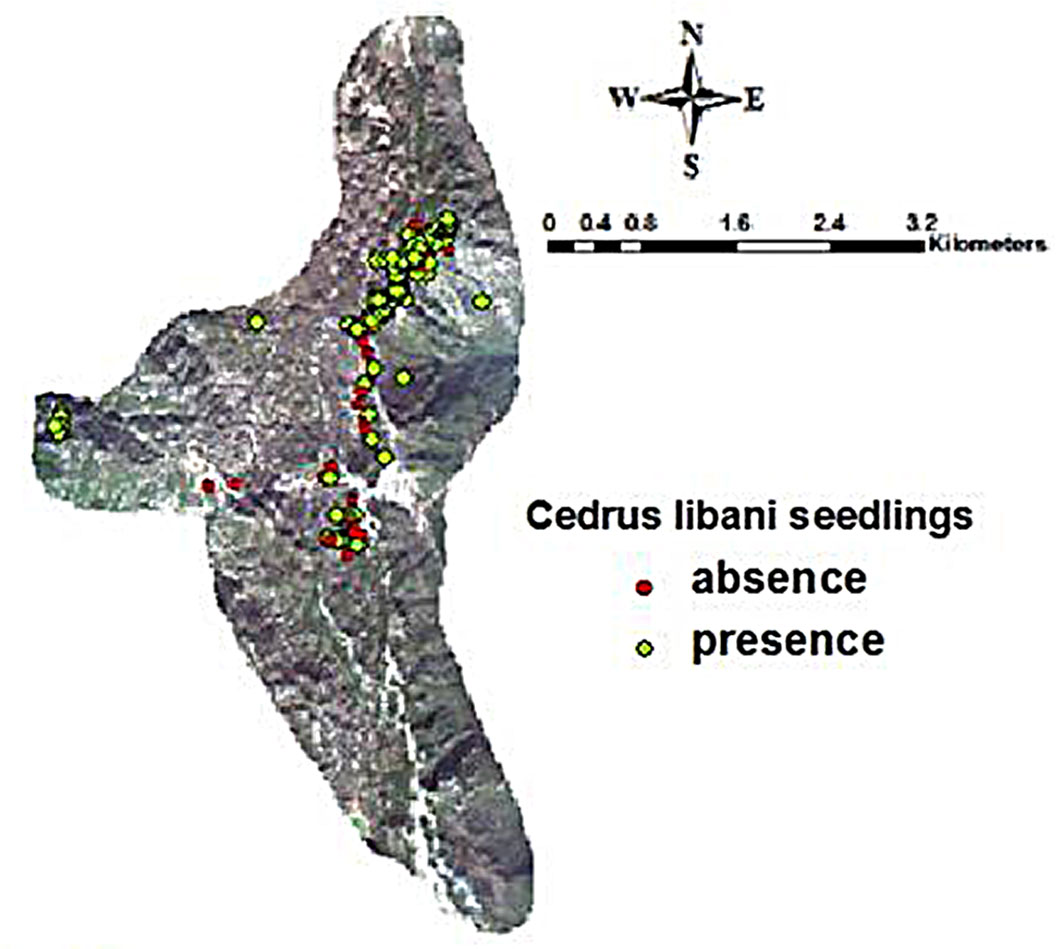
A high proportion (98%) of C. libani seedlings were healthy. Regarding microsite distribution, 71% of seedlings were found in open areas, 16% in semi-exposed areas, and 13% under cedar crowns. This pattern reflects the species’ light-demanding nature, although it can tolerate partial shade during early growth stages and occasionally at tree ages between 30 and 70 years ([7]).
Among other conifer seedlings, Juniperus oxycedrus L. and Juniperus drupacea Labill. showed the highest densities after cedar seedlings (61.5%), followed by Quercus spp. (38.5%), with an overall presence probability of 12.2% in the Jawbat Burghal forest.
The natural regeneration of C. libani in Jawbat Burghal is considered fair, with an average seedling density of 4.700 individuals ha-1. However, the average sapling density (111 ind. ha-1) remains lower than the adult tree density (330 ind. ha-1 - [40]), indicating potential limitations in seedling establishment and progression ([41], [42]). Such constraints are common in Mediterranean forests, where young woody species often experience slow growth due to climatic stress, resulting in longer transition periods from seedling to sapling than in tropical or temperate forests ([9]).
Logistic regression model analysis for the presence of C. libani seedlings
Spatial and forest structure model
The binary logistic regression model assessing spatial and forest structure variables was statistically significant (χ2 = 0.0001, P < 0.05), explaining 59% of the total variation in Cedrus libani seedling presence (Nagelkerke R2). The model effectively predicted the actual probability of seedling occurrence in the Jawbat Burghal forest, as confirmed by the Hosmer-Lemeshow goodness-of-fit test (χ2 = 0.965, P > 0.05). An overall classification accuracy of 82.8% indicates strong model performance (Tab. 1).
Tab. 1 - Classification accuracy of spatial and forest structure model.
| Observed | Predicted | Perc. Correct | |
|---|---|---|---|
| Abs | Pres | ||
| Absence | 37 | 10 | 78.7 |
| Presence | 17 | 93 | 84.5 |
| Overall Perc. | - | - | 82.8 |
Although tree and shrub cover did not significantly affect seedling presence, two variables emerged as significant predictors: stone coverage and distance from broadleaved trees. Their regression coefficients (B = 0.078 and 1.419, respectively - Tab. S1 in Supplementary material) suggest that higher stone coverage and greater distance from broadleaved trees positively influence the likelihood of C. libani seedling establishment. This finding aligns with Boydak ([6]), who reported that C. libani seedlings thrive on bare karstic terrain with 70%-75% stone cover, which reduces evaporation and enhances seedling survival. Additionally, distance from broadleaved trees may reduce competition for water and nutrients while providing partial shade during hot, dry periods. Nahal ([34]) observed that C. libani seedlings often establish under oak trees, benefiting from improved soil conditions and water uptake. However, broadleaved species can also act as strong competitors, especially in open areas, due to their rapid reproduction and shoot development ([4]).
The spatial relationships between plant species (competition, facilitation) can change over time; this temporal dynamics alters the relative sizes of individuals and their impacts on growth conditions and local climate fluctuations, thus the balance between competition and facilitation depends on the stages of plant life ([1]). The final logistic regression model is expressed as (eqn. 11):
where stone and distance_broad are the stone coverage (%) and the distance from broadleaved trees, respectively.
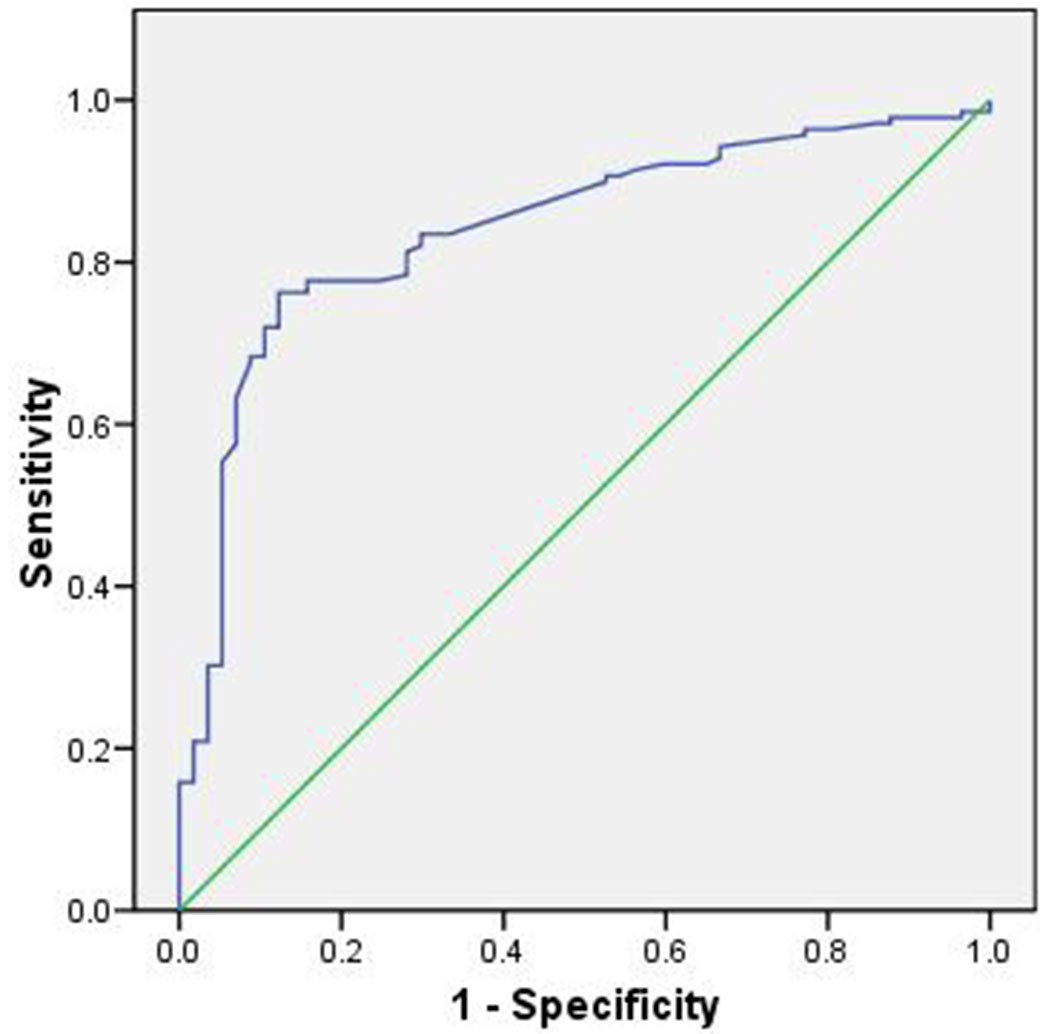
Model performance, evaluated using the ROC curve, was rated as very good, with an AUC value of 0.84, indicating high predictive accuracy (Fig. 2).
Fig. 2 - Area under the ROC curve for the spatial and forest structure model.
Topographic model
The binary logistic regression model for topographic variables was statistically significant (χ2 = 0.0001, P < 0.05), explaining 29% of the total variation in the presence of Cedrus libani seedlings (Nagelkerke R2). The model effectively predicted the probability of seedling occurrence in the Jawbat Burghal forest, as confirmed by the Hosmer-Lemeshow goodness-of-fit test (χ2 = 0.928, P > 0.05). An overall classification accuracy of 76.1% indicates that the model performs well at predicting the presence of C. libani seedlings (Tab. 2).
Tab. 2 - Classification accuracy of the topographic variables model.
| Observed | Predicted | Perc. Correct | |
|---|---|---|---|
| Abs | Pres | ||
| Absence | 15 | 42 | 26.3 |
| Presence | 5 | 135 | 96.4 |
| Overall Perc. | - | - | 76.1 |
Among the topographic variables, aspect (particularly north-facing slopes), the second slope class (10-20°), and concave landforms were statistically significant contributors to model accuracy. In general, aspect had a negative influence on seedling presence, except for southwest-facing slopes, which showed a positive effect with a regression coefficient B of 18.721 (Tab. S2 in Supplemetary material). This finding aligns with Nahal ([34]), who reported that C. libani prefers moist, wind-protected environments. Boydak ([6]) also observed higher seedling mortality on sunny exposures than on shaded ones, suggesting that seed dispersal on sunny slopes must be 1.5 to 2 times greater than on shaded slopes under similar ecological conditions.
All slope classes positively influenced seedling presence, with the second slope class (10-20°) being particularly significant. The land shape had no significant effect generally on the probability of the presence of the C. libani seedlings; the convex land shape had an insignificant positive effect (B = 0.420) while the concave land shape had a significant negative impact (B = -3.038 - Tab. S2), likely due to increased erosion risk compared to convex terrain.
C. libani is considered a fairly light-demanding species, capable of growing under partial shade during its early stages and sometimes even during the tree stage between 30 and 70 years ([23]). It can tolerate canopy cover up to 70 years of age, maintaining normal height growth, and adapts well to both vertical and lateral shading ([2]). The resulting model is as follows (eqn. 12):
where north, slope2, and curvature1 are aspect (0-22.5°), slope (10-20°), and concave shape, respectively.
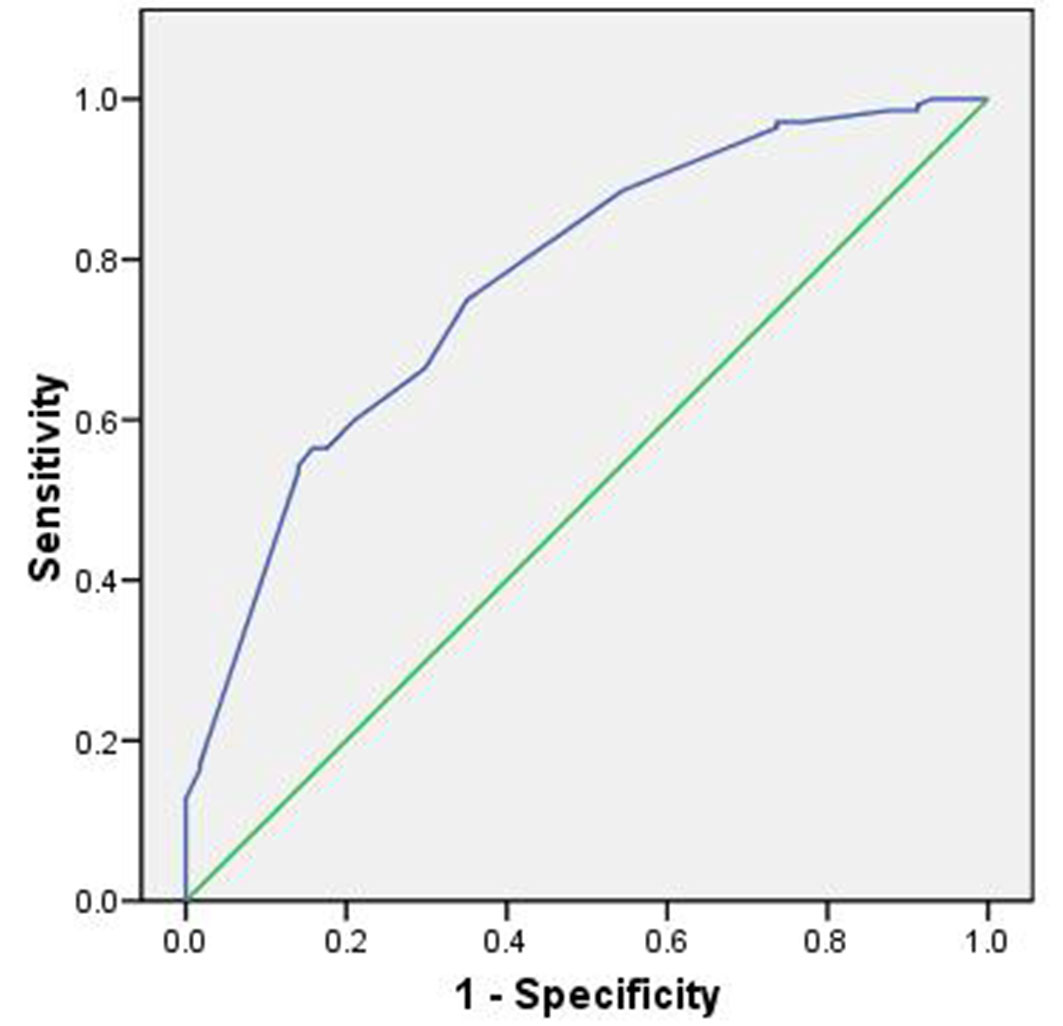
The model performance, evaluated using the ROC curve, was 0.77, indicating good predictive capability (Fig. 3).
Fig. 3 - Area under the ROC curve for the topographic model.
Remote sensing indicators
The binary logistic regression model based on remote sensing indicators was statistically significant (χ2 = 0.0001, P < 0.05), explaining 23% of the total variation in the presence of Cedrus libani seedlings (Nagelkerke R2). The model effectively predicted seedling occurrence in the Jawbat Burghal forest, as confirmed by the Hosmer-Lemeshow goodness-of-fit test (χ2 = 0.80, P > 0.05). An overall classification accuracy of 68.9% suggests that the model performs well in predicting C. libani seedling presence (Tab. 3).
Tab. 3 - Classification accuracy of spectral variables model.
| Observed | Predicted | Perc. Correct | |
|---|---|---|---|
| Abs | Pres | ||
| Absence | 23 | 33 | 41.1 |
| Presence | 23 | 101 | 81.5 |
| Overall Perc. | - | - | 68.9 |
Several spectral indicators were statistically significant, including NDWI, NDMI, MSAVI, and SMI. While NDWI, NDMI, and MSAVI exhibited negative effects on seedling presence, SMI demonstrated a positive influence. The corresponding regression coefficients B were -1.552, -1.225, -1.423, 1.318, respectively (Tab. S3 in Supplementary material). Higher soil moisture levels, indicated by elevated SMI values, were positively associated with C. libani seedling presence. This may be attributed to the presence of Jawbats, which are low-lying areas that retain water, creating favorable soil conditions, and contributing to increased precipitation rates in the region ([35]). Conversely, lower values of NDWI, NDMI, and MSAVI, indicating reduced moisture and overall vegetation health, were associated with higher probabilities of seedling presence. This pattern may reflect the vulnerability of forest cover to summer water stress in Jawbat Burghal ([16]) and suggests that C. libani seedlings can thrive in areas with reduced vegetation competition and potentially greater light availability ([35]).
Additionally, the frequent fog and cloud cover in the region contribute to elevated relative humidity, thereby enhancing growing conditions for Cedrus libani even during hot, dry periods ([35]). Peyre ([36]) observed a strong correlation between high August temperatures and fog formation, which is favorable for Cedrus species. Notably, C. libani is found on the western slopes of the Lebanon mountain range overlooking the Mediterranean Sea but is absent from the eastern slopes. This absence is likely due to the absence of favorable environmental conditions, such as fog and cloud accumulation, which increase relative humidity and mitigate summer water loss, thereby supporting the survival of C. libani. The final logistic regression model is expressed as (eqn. 13):
where NDWI, NDMI, MSAVI, and SMI are the normalized difference water index, normalized difference moisture index, modified soil adjusted vegetation index, and soil moisture index, respectively.
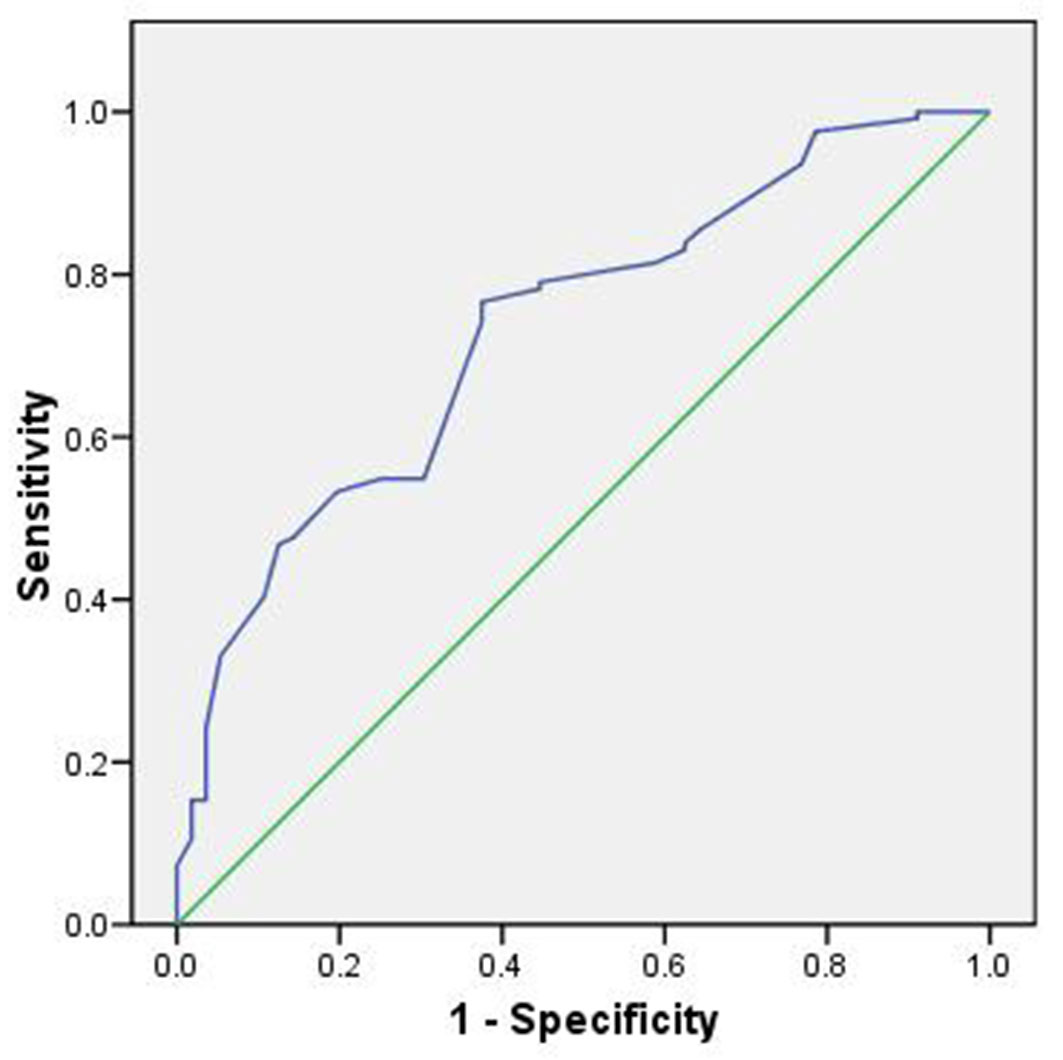
The model’s performance, evaluated using the ROC curve, demonstrated good predictive capability (AUC = 0.73 - Fig. 4).
Fig. 4 - Area under the ROC curve for the spectral model.
All combined environmental variables
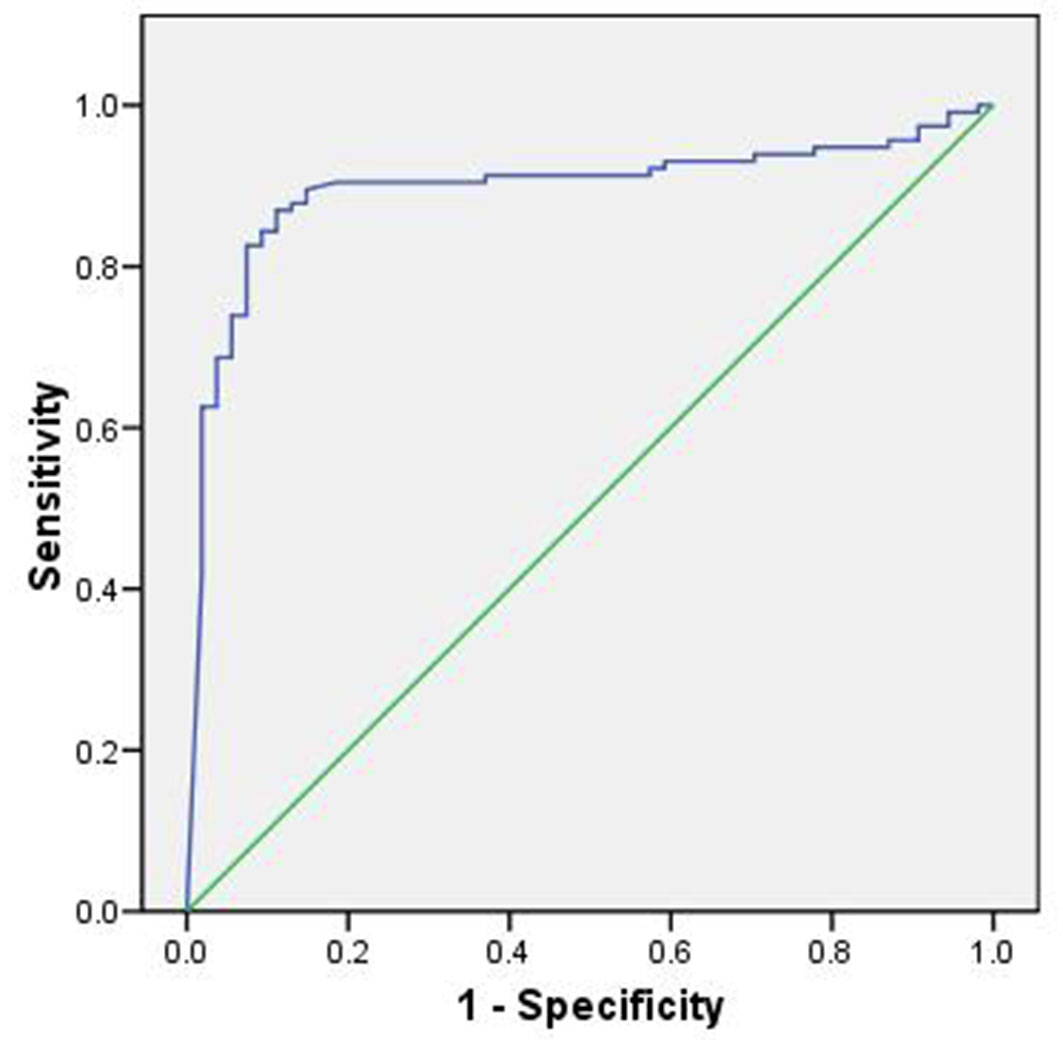
The combined binary logistic regression model, incorporating all environmental variables, was highly significant (χ2 = 0.0004, P < 0.05) and explained 87% of the total variation in the presence of Cedrus libani seedlings (Nagelkerke R²). The model demonstrated excellent predictive ability, as confirmed by the Hosmer-Lemeshow test (χ2 = 0.979, P > 0.05), and achieved an overall classification accuracy of 94.3%, indicating that the model is only 5.7% away from perfect prediction (Tab. 4). This underscores its effectiveness in estimating the actual probability of C. libani seedling presence in the Jawbat Burghal forest.
Tab. 4 - Classification accuracy of the environmental variables model. The cut value is .500
| Observed | Predicted | Perc. Correct | |
|---|---|---|---|
| Abs | Pres | ||
| Absence | 42 | 4 | 91.3 |
| Presence | 4 | 91 | 95.8 |
| Overall Perc. | - | - | 94.3 |
Several environmental variables had a statistically significant positive effect on seedling presence, including: the third slope class (20-30°), distance from broadleaved trees, soil moisture index (SMI), shrub coverage percentage, coniferous litter, broadleaved litter, and stone coverage. Their respective regression coefficients B were 40.204, 7.094, 21.684, 1.352, 10.788, 40.646, and 0.415 (Tab. S4 in Supplementary material). Conversely, the following variables had significant negative effects: elevation, eastness layer, NDMI index, C. libani tree cover, coniferous shrub cover (except cedar), broadleaved shrub cover (except oaks), and litter thickness. Their regression coefficients B were -10.252, -12.293, -13.814, -36.676, -28.400, -67.792, -9.652, respectively (Tab. S4 in Supplementary material).
The positive relationship between shrub coverage and seedling emergence aligns with findings from Mediterranean forest studies. For example, Gómez-Aparicio et al. ([14]) demonstrated a strong correlation between shrub presence and the emergence and survival of Acer opalus ssp. granatense seedlings. Similarly, Keyes & Maguire ([24]) reported that shrub cover enhances the survival of Pinus ponderosa seedlings during the critical early weeks of establishment, particularly in summer when mortality rates are highest. Forest litter may inhibit initial seedling emergence but contributes positively to survival and long-term establishment ([21]). The final regression model is expressed as (eqn. 14):
where slope3 = slope (20°-30°), distance_ broad = distance from broadleaved trees, SMI = soil moisture index, shrub = shrub coverage (%), litter_type1 = coniferous litter, litter_type2 = broadleaved litter, stone = stone coverage (%), elevation = elevation (m a.s.l.), eastness = eastness layer, NDMI = normalized difference moisture index, type1 = C. libani tree cover, type_ shrub2 = coniferous cover (except cedar shrub), type_ shrub4 = broadleaved shrub cover (except oaks), litter_thick = litter thickness.
The model’s performance, as assessed by the ROC curve, yielded an AUC of 0.90, indicating that the environmental model was highly effective (Fig. 5).
Fig. 5 - Area under the ROC curve for the environmental model.
While shrub cover may not directly enhance seedling emergence rates, it significantly improves overall regeneration by reducing mortality during the first summer ([24]). This pattern is consistent with many woody species in Mediterranean environments, where seedlings often survive and grow under shrub canopies, away from mature trees ([13]).
The quality and quantity of forest litter vary with forest structure and composition, as well as with gradients in light, water, and temperature. These factors interact to influence litter decomposition rates and nutrient accumulation in the soil, which in turn affect species regeneration, seedling survival and growth, and the spatial distribution of forest species ([37]). Miina & Saksa ([33]) suggest that litter thickness should be reduced to less than 3 cm to improve seedling establishment.
The spatial distribution of C. libani seedlings is shaped by a complex interplay of environmental factors, including light availability, litter characteristics, soil moisture, and canopy structure. Canopy cover influences microclimatic conditions and resource availability, thereby affecting seedling distribution through facilitation and competition ([20]).
Optimal forest cover density is considered a key factor in promoting seedling emergence and survival in Mediterranean forests. By modifying abiotic conditions, such as reducing evaporation and enabling deeper root access to soil moisture, appropriate canopy density enhances seedling resistance to summer drought ([39]).
Conclusion
This study demonstrated that several environmental factors positively influence the presence of Cedrus libani seedlings in the Jawbat Burghal forest. These include low elevations, steep slopes (20-30°), greater distance from broadleaved trees, proximity to forest roads, open areas, increased shrub coverage, the presence of coniferous and broadleaved litter, and higher stone coverage. In contrast, factors such as dense cedar tree cover, coniferous shrubs (excluding cedar), broadleaved shrubs (excluding oaks), and excessive litter thickness negatively affect seedling emergence.
Given the harsh environmental conditions in Jawbat Burghal, particularly water stress and soil erosion, effective canopy management is essential. Regulating understory shrub density and reducing litter thickness can enhance seedling survival and establishment.
The environmental models developed in this study offer valuable insights into the ecological drivers of C. libani regeneration. These models provide a scientific basis for prioritizing forest management interventions to promote natural regeneration. Despite the limited distribution of C. libani forests in Syria, their ecological and floristic significance highlights the urgency of conservation efforts. Sustaining population dynamics and regeneration should be a central focus for conservation managers, and the predictive models presented here can serve as practical tools to guide strategic decision-making.
References
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Higher Institute for Environmental Research, Latakia (Tishreen) University (Syria)
Department of Ecology and Forest, Latakia (Tishreen) University (Syria)
Corresponding author
Paper Info
Citation
Ibrahem A, Koubaily E, Thabeet A (2026). Assessing the environmental factors affecting the natural regeneration of Cedrus libani A. Rich. in Jawbat Burghal, Syria. iForest 19: 237-243. - doi: 10.3832/ifor4587-018
Academic Editor
Maurizio Marchi
Paper history
Received: Feb 14, 2024
Accepted: Dec 06, 2025
First online: Jun 14, 2026
Publication Date: Jun 30, 2026
Publication Time: 6.33 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2026
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 1041
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 189
Abstract Page Views: 562
PDF Downloads: 244
Citation/Reference Downloads: 0
XML Downloads: 46
Web Metrics
Days since publication: 47
Overall contacts: 1041
Avg. contacts per week: 155.04
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Jul 2026)
(No citations were found up to date. Please come back later)
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Stand structure and regeneration of Cedrus libani (A. Rich) in Tannourine Cedar Forest Reserve (Lebanon) affected by cedar web-spinning sawfly (Cephalcia tannourinensis, Hymenoptera: Pamphiliidae).
vol. 11, pp. 300-307 (online: 13 April 2018)
Review Papers
Remote sensing-supported vegetation parameters for regional climate models: a brief review
vol. 3, pp. 98-101 (online: 15 July 2010)
Review Papers
Accuracy of determining specific parameters of the urban forest using remote sensing
vol. 12, pp. 498-510 (online: 02 December 2019)
Review Papers
Remote sensing of selective logging in tropical forests: current state and future directions
vol. 13, pp. 286-300 (online: 10 July 2020)
Research Articles
Estimation of above-ground biomass and sequestered carbon of Taurus Cedar (Cedrus libani L.) in Antalya, Turkey
vol. 6, pp. 278-284 (online: 01 July 2013)
Technical Reports
Detecting tree water deficit by very low altitude remote sensing
vol. 10, pp. 215-219 (online: 11 February 2017)
Technical Reports
Remote sensing of american maple in alluvial forests: a case study in an island complex of the Loire valley (France)
vol. 13, pp. 409-416 (online: 16 September 2020)
Research Articles
Geostatistical techniques for estimating aboveground biomass in eastern Amazonia
vol. 19, pp. 85-93 (online: 13 March 2026)
Review Papers
Remote sensing support for post fire forest management
vol. 1, pp. 6-12 (online: 28 February 2008)
Research Articles
Afforestation monitoring through automatic analysis of 36-years Landsat Best Available Composites
vol. 15, pp. 220-228 (online: 12 July 2022)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







