Use of fallen dead trees by Japanese squirrels within cedar plantations in northeastern Japan
iForest - Biogeosciences and Forestry, Volume 16, Issue 5, Pages 262-267 (2023)
doi: https://doi.org/10.3832/ifor4338-016
Published: Oct 19, 2023 - Copyright © 2023 SISEF
Research Articles
Abstract
Deadwood in forests plays a critical role in maintaining the ecological functions. Small mammals use deadwood, and thus deadwood can mitigate the negative impacts of plantation on small mammals. This study focused on fallen dead trees in planted forests, and aimed to verify whether fallen dead trees affect behavioral patterns of Japanese squirrels. To clarify the use of fallen dead trees by Japanese squirrels in a Japanese cedar plantation, we observed squirrel behavior by camera trap surveys at 61 survey sites. Our findings showed that fallen dead trees play a crucial role in the behavior of Japanese squirrels, serving as landmarks for movement, vigilance, resting, and hoarding sites. These functions are critical for the survival of Japanese squirrels, suggesting that fallen dead trees in planted forests have positive impacts on their microenvironment use. The increase of deadwood due to disturbances such as heavy rainfall and snowfall resulting from climate change may provide benefits to arboreal small mammals in poorly managed planted forests. Additionally, leaving some of the deadwood generated during the harvesting process in properly managed forests can improve the quality of habitat for arboreal small mammals.
Keywords
Arboreal Small Mammal, Behavioral Ecology, Coarse Woody Debris, Forestry, Deadwood, Sciurus lis
Introduction
Deadwood within forests plays a critical role in maintaining the ecological functions of forests, including nutrient cycling, energy flow, and wildlife habitat ([13], [14], [17], [7]). This accumulation can be caused by both natural and human disturbances ([29]). Natural causes include aging, imbalanced tree growth, animal predation, disease, wind, fire, lightning, environmental stress, chemical pollution, and the effects of climate change ([31]). Meanwhile, human activities such as forestry operations and mismanagement of planted forests can also contribute to the generation of deadwood through logging and the abandonment of forest residues and unused thinning material ([41]). Inadequate management practices may also weaken the wind resistance of individual trees following intensive thinning or selective cutting ([32]), leading to the growth of unhealthy trees that are more vulnerable to natural disturbances (such as wind and snow damage) and resulting in increased deadwood ([27], [18]). In Japan, recent climatic changes, including an increase in days with daily precipitation exceeding 100 mm and heavy rainfall greater than 200 mm ([20]) and projections of increased heavy snowfall risk ([26]), emphasize the importance of proper management of planted forests to reduce the impact of climate change on deadwood occurrence.
Previous studies have demonstrated the correlation between the presence of fallen dead trees and the selection of habitats and population densities by small mammals ([6], [11]). Small mammals use logs and fallen dead trees as routes for movement and escape from predators, as well as for foraging and nesting sites ([6], [8]). Thus, deadwood can mitigate the negative impacts of deforestation on small mammals. The presence of above-ground deadwood is also beneficial for arboreal small mammals. Logs within forests serve as efficient movement routes and significant food hoarding sites, even in snowy environments ([6], [3], [35], [24]). Above-ground deadwood also offers arboreal small mammals’ complex microhabitats, diverse food resources, and concealment from predators ([30], [1], [3], [24]). Disturbances resulting from above-ground deadwood have a significant impact on arboreal small mammal habitat use, and they are more likely to occur in planted forests compared to natural forests ([32]). Therefore, it is crucial to assess the effects of above-ground deadwood on arboreal small mammals in planted forests, despite the limited research currently available.
In this study, we focused on the relationship between Japanese squirrels, a small arboreal mammal endemic to Japan, and above-ground deadwood. Japanese squirrels are primarily arboreal but are known to often use the ground ([39], [37]). In the cool temperate zone, Japanese squirrels occupy parts of cedar plantations as their habitat ([15], [16]). This study focused on fallen dead trees, a common form of deadwood in planted forests, and aimed to verify whether fallen dead trees affect behavioral patterns of Japanese squirrels. To do this, we employed camera trap surveys to observe the differences in the types of behaviors exhibited by Japanese squirrels on fallen dead trees versus on the ground. If any differences were identified, we analyzed the relationship between the behavior types and the fallen dead tree size and environmental factors.
Material and methods
Study area
This study was conducted in Tsuruoka City, northeastern Japan (38° 37′ N, 139° 50′ E - Fig. 1), located in the cool temperate zone with an annual precipitation of 2191.4 mm and an average annual temperature of 12.9 °C ([19]). The mountainous areas of the study area experience heavy snowfall with snow depths exceeding 3 m. Japanese squirrels are widely distributed in the forested sites of the study area and are well-suited for field surveys as they often use conifer plantations ([15], [16]). The study area was divided in three areas: Kaminagawa (elevation: 195-410 m a.s.l.), Kushibiki (elevation: 110-260 m), and Kinbosan (elevation: 80-170 m - Fig. 1). Typical forest types in the study area are Fagus crenata forests, Quercus crispula forests, and Japanese cedar (Cryptomeria japonica) plantations. As the focus of this study was deadwood in plantations, field surveys were conducted within cedar plantations in all three areas.
Fig. 1 - Location of the study area. Circles represent the survey sites.
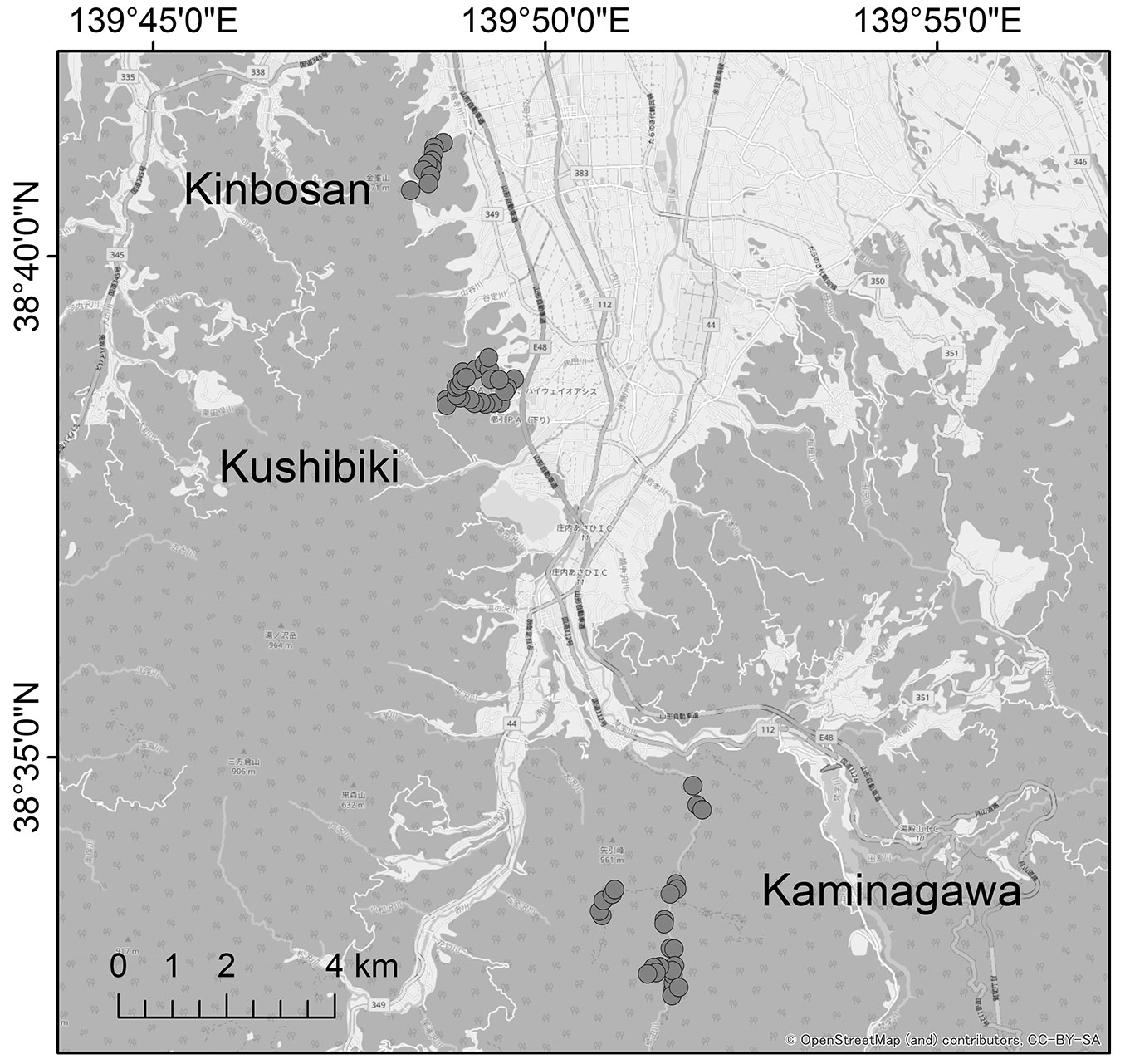
At each of the study areas, we located pre-existing fallen dead trees and established survey sites. The conditions of the fallen dead trees were defined based on previous studies ([29], [14]) as having a minimum length of 1 m and a diameter of at least 10 cm at its thickest point. The distance between the survey sites was set to be at least 100 m, taking into account the daily range of activity of Japanese squirrels ([39]). A total of 61 survey sites were established, with 26 in Kaminagawa, 25 in Kushibiki, and 10 in Kinbosan.
Measurement of fallen dead trees and environmental factors
During the period of August to October 2022, we conducted measurements of fallen dead trees and environmental factors at each of the 61 survey sites. Based on previous studies on the American red squirrel (Tamiasciurus hudsonicus), we selected the following measurements ([1]): fallen dead tree size, ground slope angle, mean diameter at breast height (DBH) of the five nearest cedars from the fallen dead tree, number of Japanese walnut trees (Juglans ailanthifolia), degree of understory visibility, canopy openness, and distance from the fallen dead tree to the nearest tree.
The size of the fallen dead tree (m3) was calculated as its volume assuming a cylindrical shape based on its length and diameter measurements. The diameter was measured at the thickest point ([2]). The ground slope angle (°) was measured using a Vertex™ IV (Haglöf, Sweden). The average DBH (cm) of five cedar trees, excluding standing dead trees and seedlings, was calculated as an indicator of forest age. The number of trees in a 20 m radius around the fallen dead tree was counted, based on previous studies examining hoarding and feeding sites of Japanese squirrels ([34], [4]). To assess the degree of understory visibility, we marked five points at 20 cm intervals from the ground up to 1 m and observed them from a distance of 10 m, repeating this process three times at 1 m intervals. The percentage of the number of visible points (%) was used as an index of understory visibility. The degree of canopy openness was assessed by taking hemispheric photographs at a height of 1.2 m above the ground, using a digital camera (Coolpix™ P5100, Nikon, Japan) with a fisheye lens (FC-E8, Nikon), on cloudy days during the survey period. The degree of canopy openness (%) was calculated using CanopOn2 ([33]) analysis of these photographs. The distance (in cm) from the fallen dead tree to the nearest standing tree with a height greater than 2.5 m was also measured ([1]).
Camera trap survey
To examine the use of the fallen dead trees and ground by Japanese squirrels, we performed camera traps equipped with infrared sensor cameras (BTC-6HD-MXP, Browning, UK) from August to October 2022. The camera traps were installed at each survey site for 7-14 days. The main camera trap was placed 4 m away from the fallen dead tree, at a height of approximately 70 cm from the ground, to capture the squirrels’ behavior on both the fallen dead tree and the ground. The camera was fixed in a horizontal direction 90° from the fallen dead tree, and the angle of view was adjusted to allow for both the fallen dead tree and ground to be visible. The camera trap recorded 20-second videos, with a 1-second interval between each shot. Additionally, sub-camera traps (Torophy Cam HD 20MP No Glow, Bushnell, KS, USA; Core 24MP No Glow 119938C, Bushnell; HykeCam SP2, Hyke, Japan) were installed at a 45° horizontal angle from the fallen dead tree, capturing 15-second videos. The settings of the sub-camera traps, except for the camera view direction and video length, were identical to those of the main camera traps.
In this study, we treated consecutive behaviors of Japanese squirrels captured within a 10-minute interval as being performed by the same individual, as described in previous studies ([36], [40]). For each event, we recorded the date and time of the shooting, using the data from the first video file, and the squirrels’ behavior on the fallen dead tree and the ground. Behaviors were classified as locomotion, vigilance, comfort behavior, and foraging, including food-transporting, food-searching, hoarding, and walnut manipulating, based on previous studies ([9], [22], [12], [23] - see Tab. 1). For each event, we recorded the presence or absence of each behavior type ([40]). Videos with multiple individuals were excluded because the presence of other squirrels could affect their behavior. Data from the main camera traps was used whenever possible; however, in cases where data from the main camera could not be obtained due to technical issues, data from the sub-camera traps was used.
Tab. 1 - Behavior types and definitions observed by camera trap survey. Behavioral types were defined with reference to previous studies ([9], [22], [12], [23]).
| Behavior type | Subtype | Definition |
|---|---|---|
| Locomotion | - | Behavior of moving on fallen dead trees and the ground |
| Vigilance | - | Behavior of pausing and sniffing |
| Comfort | - | Behavior of grooming, cheek rubbing, and resting |
| Foraging | Food-transporting | Behavior of transporting a walnut |
| Food-searching | Behavior of looking and digging at the ground without holding a walnut | |
| Hoarding | Behavior of looking, digging at the ground with holding a walnut, and burying walnut in the ground | |
| Walnut manipulating | Behavior of removing the rind and soil from the walnut, and putting the walnut back in the mouth. |
Statistical analysis
To investigate the differences in the behavior of Japanese squirrels on fallen dead trees and on the ground, we first conducted a chi-square test using the number of observations of each behavioral type. If significant differences were detected, we further conducted residual tests to determine the specific behaviors that differed between the fallen dead trees and the ground. The number of observations for a behavioral type was defined as the number of events in which that behavior was recorded ([40]).
To explain the effect of fallen dead tree size and environmental factors on each behavioral type, we constructed a generalized linear mixed model (GLMM). The response variable followed a binomial distribution and was represented as the presence (1) or absence (0) of each behavioral type at each camera trap event. Seven explanatory variables were used in this analysis: fallen dead tree size, ground slope angle, mean DBH, number of Japanese walnut trees, understory visibility, canopy openness, and distance to the nearest tree. We also included the shooting month as an additional explanatory variable to account for seasonal effects, and included survey sites as random effects. Due to the potential issue of multicollinearity, we calculated the correlation coefficients between the variables and found a high negative correlation (r = -0.71) between canopy openness and understory visibility. As a result, we only used the understory visibility in the analysis. No other high correlations (|r| < 0.7) were found between the explanatory variables, thus we assumed that multicollinearity did not arise ([5]). The relative importance of variables was evaluated by the absolute value of their standardized regression coefficients.
Data analysis was performed using R ver. 3.6.1 ([28]) and the package “glmmML”.
Results
A total of 578 camera days were carried out during the camera trap survey. Japanese squirrels were recorded at 34 out of 61 survey sites, yielding a total of 154 events. The behaviors of Japanese squirrels were observed 163 times on fallen dead trees and 229 times on the ground (Tab. 2). Locomotion, vigilance, comfort, hoarding, and walnut manipulating behaviors were observed on fallen dead trees. On the ground, locomotion, vigilance, comfort, food-transporting, food-searching, hoarding, and walnut manipulating behaviors were observed. The observed comfort behaviors included grooming on the ground (1 event), and cheek rubbing with a walnut in the mouth (11 events) and sprawled lying posture (1 event) on fallen dead trees. Cheek rubbing with a walnut in the mouth was only recorded at 2 sites and was repeated at the same location. Food-searching behavior was only observed on the ground, and no food-searching behavior related to fallen dead trees was observed.
Tab. 2 - Number of observed events in each behavior type and the results of a chi-square test (p-value in total) and residual tests (p-value in each behavior type).
| Behavior type | Subtype | No. of events | p-value | |
|---|---|---|---|---|
| Fallen dead tree |
Ground | |||
| Locomotion | - | 59 | 43 | <0.01 |
| Vigilance | - | 57 | 69 | 0.31 |
| Comfort | - | 13 | 1 | <0.01 |
| Foraging | Food-transporting | 27 | 42 | 0.65 |
| Food-searching | 0 | 28 | <0.01 | |
| Hoarding | 0 | 42 | <0.01 | |
| Walnut manipulating | 7 | 4 | 0.13 | |
| Total | - | 163 | 229 | <0.01 |
A chi-square test indicated that the frequency of behaviors was significantly different between fallen dead trees and the ground (χ2 = 79.149, df = 6, p < 0.01 - Tab. 2). Residual tests revealed significant differences among behavior types in locomotion and food-searching, hoarding, and comfort behaviors (p < 0.01 - Tab. 2).
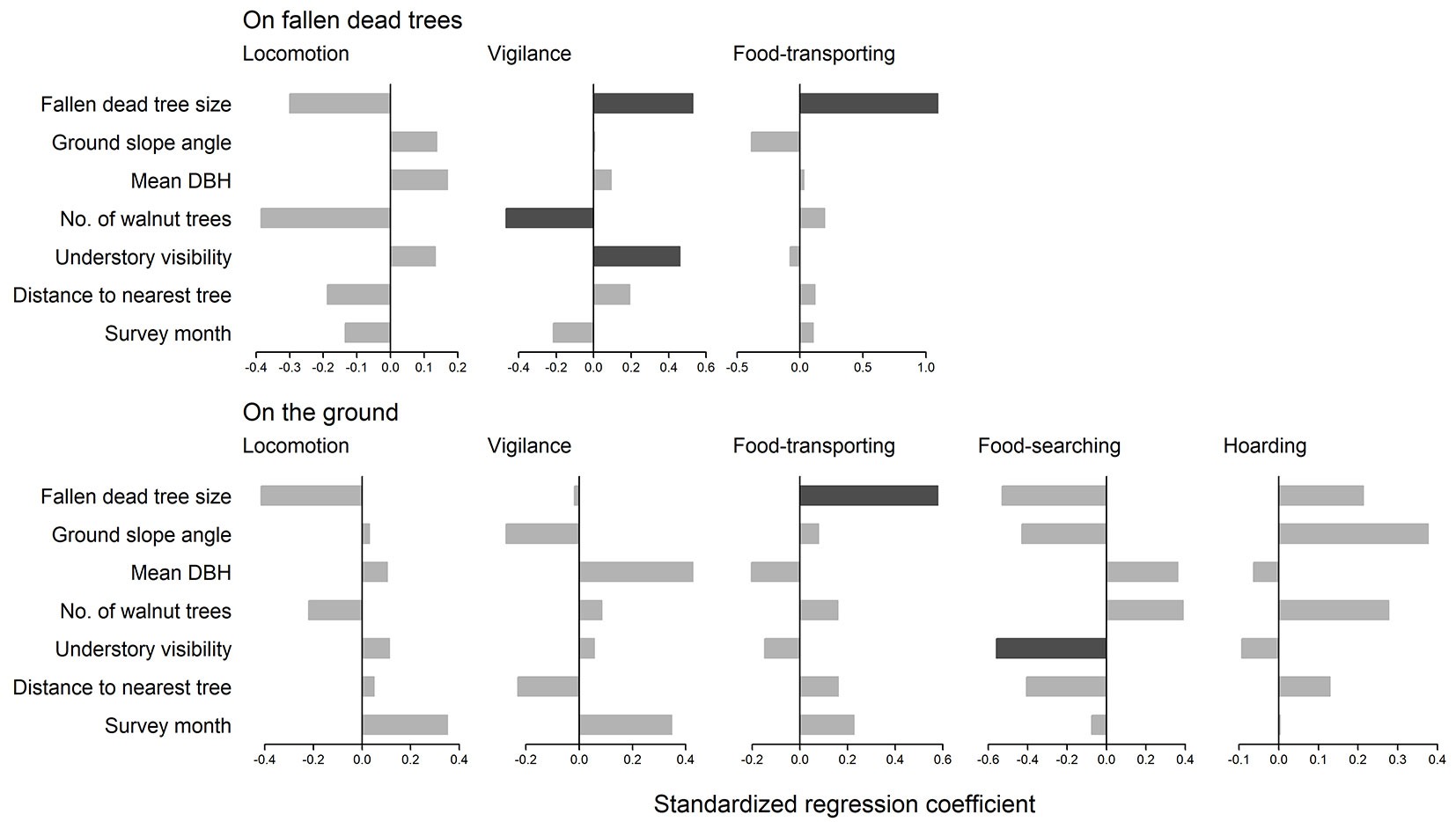
Locomotion and vigilance behaviors were observed at more than 5 sites on both fallen dead trees and the ground, while food-transporting, food-searching, and hoarding behaviors were only observed at more than 5 sites on the ground. The GLMM results showed no significant differences in locomotion behavior for either fallen dead trees or ground with regards to either variable (p ≥ 0.05 - Fig. 2). No significant differences were detected in vigilance behavior on the ground for any of the variables (p ≥ 0.05 - Tab. 2). However, significant differences were detected in vigilance behavior on fallen dead trees for three variables (p < 0.05 - Fig. 2), indicating that vigilance behavior was more frequent on larger fallen dead trees with better understory visibility and fewer nearby walnut trees. The GLMM results for vigilance behavior indicated that fallen dead tree size was the most important variable, followed by the walnut trees and understory visibility (Fig. 2). For hoarding behavior, significant differences were detected for both fallen dead trees and ground (p < 0.05 - Fig. 2), indicating that this behavior was more prevalent in areas with larger fallen dead trees. The GLMM results for food-searching behavior on the ground indicated that it was more frequent in areas with poor understory visibility (p < 0.05 - Fig. 2). No significant differences were detected in hoarding behavior on the ground for any of the variables (p ≥ 0.05 - Fig. 2).
Fig. 2 - Standardized regression coefficients of GLMMs for each behavior type on fallen trees and the ground (n = 154). Dark gray indicates significant differences (p < 0.05), and light gray indicates no significant differences (p ≥ 0.05).
Discussion
Behavioral differences between fallen dead trees and the ground
More locomotion and comfort behaviors were observed on fallen dead trees than on the ground, indicating that Japanese squirrels exhibit different usage patterns for fallen dead trees and the ground (Tab. 2). Fallen dead trees were used as movement routes by Japanese squirrels, as previously reported in other studies ([6], [30], [1], [3]). Japanese squirrels can move quickly and efficiently over fallen dead trees, and the dry litter can help to conceal them from predators through the reduction of auditory cues ([10], [1]). The observation of comfort behaviors such as resting and rubbing only on fallen dead trees suggests that they serve as an intermediary between tree tops and the ground, offering a small resting site during locomotion. Cheek rubbing by squirrels has been linked to grooming activities during foraging ([12]), and it is possible that the animals may have been removing smudges that adhered to them while collecting walnuts. Additionally, repeated cheek rubbing at limited sites could indicate a marking behavior to help the animals familiarize themselves with their environment and orient their activities.
In contrast, food-searching and hoarding behaviors were more commonly observed on the ground, likely due to the fact that Japanese squirrels frequently use it as a hoarding site ([21], [25], [34], [38]). While previous studies have indicated that squirrels consume fungi and insects from decaying logs and fallen dead trees as a food source ([30], [3]), no foraging behavior related to fallen dead trees was observed in this study. This could be a result of the survey being conducted during the walnut fruiting period, when other food resources were scarce.
Relationship between behavioral type and fallen dead trees and environmental factors
The differences in behavior between fallen dead trees and the ground showed that fallen dead trees affect behavioral patterns of Japanese squirrels (Tab. 2). The GLMM results also indicated that fallen dead tree size and surrounding environmental factors influence squirrel behavior (Fig. 2). Fallen dead tree size was found to have the greatest impact on vigilance behavior on fallen dead trees (Fig. 2). Larger fallen dead trees offer a higher vantage point, allowing the squirrels to observe their surroundings more effectively and move more efficiently for longer periods of time. Vigilance behavior was also more common in areas with high understory visibility (Fig. 2), as these environments offer easy predator detection, particularly when large fallen dead trees are present and the understory vegetation is sparse. Japanese squirrels also tended to be more vigilant in areas with fewer nearby walnut trees (Fig. 2). This may be because squirrels have more pauses in unfamiliar locations, which decrease with repeated experience ([23]). Repeated visits to walnut trees for hoarding behavior may reduce the number of pauses near these trees.
Foraging behavior was also affected by fallen dead tree size, and walnut transporting behavior increased near large fallen dead trees (Fig. 2). Japanese squirrels hoard walnuts in a dispersed manner on the ground ([34]), and previous studies have shown that piles of logs in forests can serve as conspicuous hoarding sites for non-hibernating Hokkaido squirrels (Sciurus vulgaris orientis - [24]). Large fallen dead trees may therefore act as visible food reservoirs for Japanese squirrels. Food-searching behavior, which is a type of foraging behavior on the ground, was more prevalent in areas with poor understory visibility (Fig. 2). This behavior leaves the squirrels vulnerable to predators as they dig in the soil with their eyes focused on the ground. Japanese squirrels engage in this risky behavior in areas with dense understory vegetation to avoid predators, as they may easily hide and detect approaching predators by the sound of rustling grass.
Conclusions
In this study, we investigated the use of fallen dead trees by Japanese squirrels in cedar plantations. Our findings showed that fallen dead trees play a crucial role in the behavior of Japanese squirrels, serving as landmarks for locomotion, vigilance, resting, and hoarding sites. These functions are critical for the survival of Japanese squirrels, suggesting that fallen dead trees in planted forests have positive impacts on their use of microenvironments. However, it is advisable to conduct further research to determine the behavior of Japanese squirrels in other seasons and other types of deadwood to provide a comprehensive understanding of deadwood function.
The results of this study support previous studies that found fallen dead trees positively affect small mammals in both well-managed and poorly managed planted forests ([6], [30], [1], [3], [8], [35], [11], [24]). The increase of deadwood due to disturbances such as heavy rainfall and snowfall resulting from climate change may provide benefits to arboreal small mammals in poorly managed planted forests. Additionally, leaving some of the deadwood generated during the harvesting process in properly managed forests can improve the quality of habitat for arboreal small mammals. Future research can assess the value of planted forests for conservation management by examining the impact of deadwood on Japanese squirrel populations and other arboreal small mammals.
Acknowledgements
This study was conducted with the permission of the owner of the forests in the study area. We thank the staff of the Experimental Forest of Yamagata University, Japan and the associates of Yamagata University for supporting our field survey.
Author Contributions
SH and MUS designed the study, SH conducted data collection and analyses, and SH and MUS wrote the manuscript.
References
Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Graduate School of Agricultural Sciences, Yamagata University, Tsuruoka 9978555 (Japan)
Faculty of Agriculture, Yamagata University, Tsuruoka 9978555 (Japan)
Corresponding author
Paper Info
Citation
Honda S, Saito MU (2023). Use of fallen dead trees by Japanese squirrels within cedar plantations in northeastern Japan. iForest 16: 262-267. - doi: 10.3832/ifor4338-016
Academic Editor
Mirko Di Febbraro
Paper history
Received: Mar 01, 2023
Accepted: Jul 23, 2023
First online: Oct 19, 2023
Publication Date: Oct 31, 2023
Publication Time: 2.93 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2023
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 19045
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 14459
Abstract Page Views: 2601
PDF Downloads: 1582
Citation/Reference Downloads: 2
XML Downloads: 401
Web Metrics
Days since publication: 981
Overall contacts: 19045
Avg. contacts per week: 135.90
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
(No citations were found up to date. Please come back later)
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Impact of the four-eyed fir bark beetle (Polygraphus proximus Blandf.) on coarse woody debris stock and carbon budget of fir forests in Central Siberia
vol. 19, pp. 209-218 (online: 08 June 2026)
Research Articles
Long-term effects of single-tree selection cutting management on coarse woody debris in natural mixed beech stands in the Caspian forest (Iran)
vol. 10, pp. 652-658 (online: 20 June 2017)
Research Articles
Carbon stock in Kolli forests, Eastern Ghats (India) with emphasis on aboveground biomass, litter, woody debris and soils
vol. 4, pp. 61-65 (online: 05 April 2011)
Research Articles
Decomposition of Norway spruce and European larch coarse woody debris (CWD) in relation to different elevation and exposure in an Alpine setting
vol. 9, pp. 154-164 (online: 28 August 2015)
Research Articles
Amount and distribution of coarse woody debris in pine ecosystems of north-western Spain, Russia and the United States
vol. 7, pp. 53-60 (online: 28 October 2013)
Review Papers
Linking deadwood traits with saproxylic invertebrates and fungi in European forests - a review
vol. 11, pp. 423-436 (online: 18 June 2018)
Research Articles
Effects of silvicultural thinning on stand structure and coarse woody debris in the deciduous Arasbaran forest, Iran
vol. 16, pp. 377-384 (online: 19 December 2023)
Short Communications
Gliding patterns of Siberian flying squirrels in relation to forest structure
vol. 12, pp. 114-117 (online: 11 February 2019)
Research Articles
Effect of topography on tree species composition and volume of coarse woody debris in an Oriental beech (Fagus orientalis Lipsky) old growth forests, northern Iran
vol. 9, pp. 658-665 (online: 17 March 2016)
Research Articles
Rewilding beech-dominated temperate forest ecosystems: effects on carbon stocks and biodiversity indicators
vol. 18, pp. 1-9 (online: 02 February 2025)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







