
Species interactions in pure and mixed-species stands of silver fir and European beech in Mediterranean mountains
iForest - Biogeosciences and Forestry, Volume 14, Issue 1, Pages 1-11 (2021)
doi: https://doi.org/10.3832/ifor3476-013
Published: Jan 02, 2021 - Copyright © 2021 SISEF
Research Articles
Abstract
Interactions between tree species determine the dynamics of forest communities. Spatial and temporal changes in resource availability, variation in species composition and spatial distribution of trees may alter competitive interactions between species and, therefore, affect tree growth and forest productivity. In this study, we analyzed the intra and inter-specific interactions between European beech (Fagus sylvatica L.) and silver fir (Abies alba Mill.) in southern Italy (Molise and Calabria regions), and how these interactions affect basal area increments in mixed-species and pure stands. Results showed that intra-specific interactions have a negative effect on the basal area increment, both in pure and mixed-species stands of Molise and Calabria. Basal area increment was higher influenced by intra-specific interactions in pure stands than in mixed-species stands. Silver fir in Molise showed higher basal area increment in mixed-species stand, probably in relation with stand structure and space occupation that resulted in less competition between individual trees. European beech showed high values of intra-specific interactions in pure stands, likely related to the low self-tolerance of this species and to the spatial arrangement of trees, due to canopy closure. The absence of inter-specific interactions in mixed-species stands could be explained by the sub-dominant position of European beech, which may have limited the benefit derived from niche separation and complementarity for silver fir.
Keywords
Abies alba Mill., Fagus sylvatica L., Intra-specific Interactions, Inter-specific Interactions, Stand Productivity, Tree Growth
Introduction
Interactions between trees in forest stands are often dynamics due to spatial and temporal changes in resource availability ([64], [12], [20], [21]). Competitive interactions may also increase due to changes in species composition and to different spatial patterning of trees (clustered or uniform - [37]), strongly influencing tree growth. Therefore, interactions between tree species are considered the main mechanisms responsible for the effects of niche complementarity and the most important drivers for the dynamics of forest communities.
Several studies have shown that higher tree species richness may lead to greater forest stand productivity ([67], [35]). Mixed-species stands showed higher degree of resistance and resilience to biotic and abiotic disturbances ([32], [60]), and wider range of ecosystem services in comparison to corresponding monocultures ([26]). Nevertheless, further experimental insights on the ecosystem functions of mixed-species stands and the ecological mechanisms that cause their over-yielding is required for implementing resource-efficient management strategies ([21]), particularly, in drought-prone environments ([29], [15]). Indeed, under limited resource availability, as the number of individuals in a tree population tends to fill the available growing space and the density continues to increase, competition for water (besides nutrients and light) increases as well ([63]).
Interactions between individual trees in a forest stand can be studied and identified through the use of competition indices ([16], [6], [7]). In mixed-species populations, in order to better understand the interactions that occur between tree species, a distinction between intra- and inter-specific competition indices is recommended ([18]). Several studies have found that in presence of a reduction of competition or of a facilitating effect ([18]) inter-specific interactions can advantage tree growth in mixed-species stands ([3]). Although in mixed-species stands the interactions are often less severe than in the corresponding monocultures ([13], [45]), the positive effects of neighborhood complementarity on tree growth may depend on several factors, such as stand density ([14]), site quality ([49]) and changes in growth conditions ([18]). It must be pointed out, however, that distinguishing between competition reduction and facilitation effect is almost impossible and, therefore, these two processes are often defined as complementarity ([23], [1]).
Competitive interactions between tree species, but also complementarity effects, may occur above- and/or below-ground. In forests with tree species that differ intrinsically in crown architecture and leaf phenology (e.g., deciduous vs. evergreen species, conifers vs. broadleaved trees), species-specific occupation of the growing space and partition of the intercepted radiation may play a major role in the use of complementary resources ([22], [53]). Moreover, in mixed-species stands, with shade-tolerant species and a heterogeneous spatial structure, the use of limiting resources (e.g., light) can be more efficient, since the trees in the dominated plane may take advantage of irradiance transmitted through the crown of the dominant trees, so as to survive and grow ([52], [38]). Further, also co-occurrence of species that differ in their root architecture may improve the uptake of nutrients and water ([11]). Below-ground competition does not only consider the interactions between dominant or/and sub-dominant trees with the subject tree, but also those trees whose roots occupy the root distribution area of the subject tree ([10]). For example, silver fir (Abies alba Mill.) develops a deeper root system than European beech (Fagus sylvatica L.) and adopts a strategy that allows the absorption of water during the growing season, namely in periods of high drought stress ([65]). Species-specific root uptake, but also capacity stomatal adjustment, may affect the hydraulic redistribution of soil water, particularly in drought-prone Mediterranean environments. Though unclear distinction between silver fir and European beech in the sensitivity of stomatal conductance to water potential has been reported ([33]), silver fir promptly closes the stomata to maintain leaf water potential during drought events (i.e., isohydric behavior - [39]). European beech, instead, continues to transpire even when soil water content diminishes (i.e., anisohydric behavior - [39]).
European beech has been often studied mixed with the comparatively drought-intolerant Norway spruce (Picea abies Karst. - [48]) or drought-tolerant Scots pine (Pinus sylvestris L. - [15]). However, mixed-species forests that include silver fir and European beech have recently earned attention and praise among forest researchers ([18], [10], [54]). Renewed interest for silver fir can be related to recent findings of enhanced productivity of silver fir in central Europe ([9]) and to the relatively high drought tolerance of this species, particularly in southern Italy and mountain environments ([2]). Silver fir has a geographical distribution similar to that of European beech, though restricted to mountain ranges, and grow well in the southernmost locations in the mountain ranges of Italy ([41]). Toward its southern distribution limit, silver fir can be competitive with European beech (moderately drought tolerant) and, in central Europe, a good replacement for the more drought-sensitive Norway spruce at higher elevations ([59]).
This study aimed to analyze intra- and inter-specific interactions between European beech and silver fir, and the influence these interactions may have on the growth of trees in mixed-species stands and corresponding pure stands. In order to better understand the interactions between these two major forest species, we used competition indices at the individual tree level. Individual competition indices reflect the local density of the competitors, which interact with the reference tree ([66]), giving the possibility to quantify the influence of neighboring individuals on the growth of the reference tree. We hypothesized that: (i) intra-specific competition is higher in pure stands than in mixed-species stands; and (ii) tree species mixing and stand structural complexity mitigate the negative effect of inter-specific interaction.
Materials and methods
Study sites
The study sites included two triplets located in southern Italy (Molise and Calabria regions). In particular, the following forest sites were sampled: (i) “Abeti Soprani” (Molise) and (ii) Sila National Park (“Sila Piccola”, Calabria - Fig. 1).
Fig. 1 - Location of the study sites in the southern Italy. Red triangle indicates silver fir in the pure stands (S); green circle indicates European beech in the pure stands (B); yellow square indicates the mixed-species stands (M).
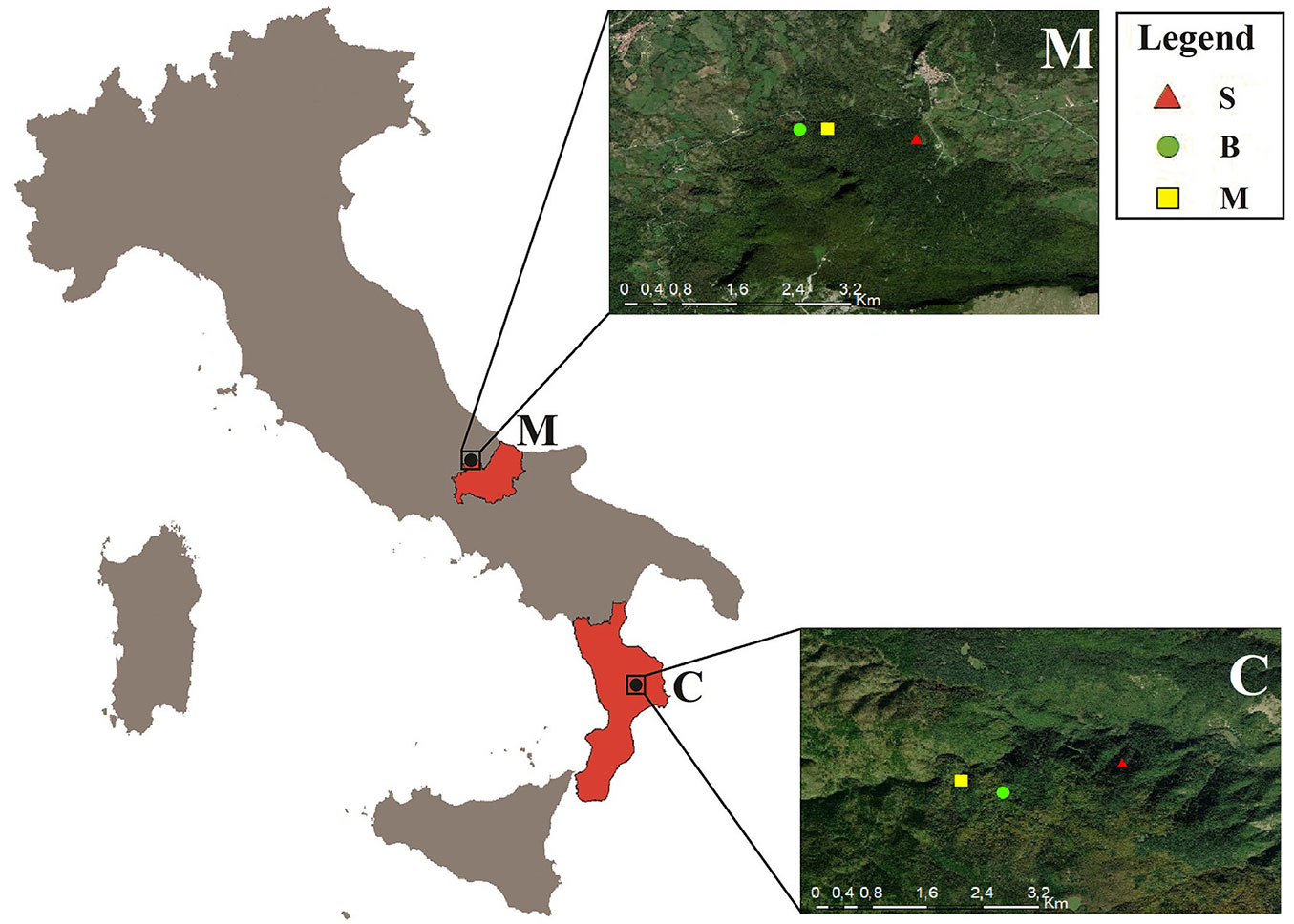
Each triplet is composed of three rectangular stands, including two pure stands of silver fir and European beech, and one mixed-species stand of these species. Stand size varied between 0.12 and 0.28 ha. Stands were selected according to the minimum number of individuals (≥ 50 trees of silver fir and/or European beech per stand) and on the elevation range (<250 m between the triplets). Furthermore, mixed and pure stands were identified on the basis of the basal area occupied by the tree species investigated inside the stand, according to the classification of Mina et al. ([43]). Stands were considered pure when the basal area of the studied tree species was at least 85%. Mixed-species stands, on the other hand, were classified as a mixture of the two species when, both species represented 85% or more of the total basal area and each of the two species contributed at least 20% of the basal area (Tab. 1). The horizontal structure was different in the two regions. Stands in Molise showed a random spatial arrangement of the trees, while in Calabria the structure was mainly clustered (Fig. 2).
Tab. 1 - Location of the study sites and structural parameters in the pure and mixed-species stands of European beech and silver fir in Molise and Calabria. (BA): stand basal area; (BAs): basal area percentage of the main species; (dg): quadratic mean diameter; (hg): quadratic mean height.
| Stand Characteristics |
Molise | Calabria | ||||||
|---|---|---|---|---|---|---|---|---|
| beechpure | firpure | beechmixed | firmixed | beechpure | firpure | beechmixed | firmixed | |
| Latitude (N) | 41°52′ 13.65″ | 41°52′ 11.02″ | 41°52′ 14.35″ | 39°09′ 13.49″ | 39°09′ 19.31″ | 39°09′ 07.96″ | ||
| Longitude (E) | 14°16′ 37.85″ | 14°17′ 26.03″ | 14°16′ 51.26″ | 16°39′ 53.05″ | 16°41′ 2.30″ | 16°40′ 12.33″ | ||
| Aspect (°) | 354 | 317 | 327 | - | 343 | 319 | 307 | - |
| Altitude (m a.s.l.) | 1182 | 1230 | 1182 | - | 1289 | 1220 | 1332 | - |
| Slope (°) | 21 | 20 | 27 | - | 24 | 37 | 28 | - |
| BA (m2 ha-1) | 33.6 | 54.4 | 45.2 | - | 59.7 | 61.5 | 74.6 | - |
| BAs (%) | 92 | 92 | 20 | 78 | 86 | 85 | 33 | 61 |
| No. trees ha-1 | 989 | 486 | 134 | 202 | 641 | 750 | 293 | 293 |
| dg (cm) | 21 | 37 | 28 | 58 | 35 | 32 | 35 | 47 |
| hg (m) | 15 | 21 | 19 | 27 | 22 | 21 | 22 | 27 |
| Soil depth (cm) | 20 | 50 | 40 | - | 50 | 70 | 90 | - |
Fig. 2 - Spatial arrangement of trees in pure and mixed-species stands in Molise (A: silver fir; B: European beech; C: mixed-species) and in Calabria (D: silver fir; E: European beech; F: mixed-species). Green indicate silver fir and violet European beech. A different colour is assigned to each species (minor species included).
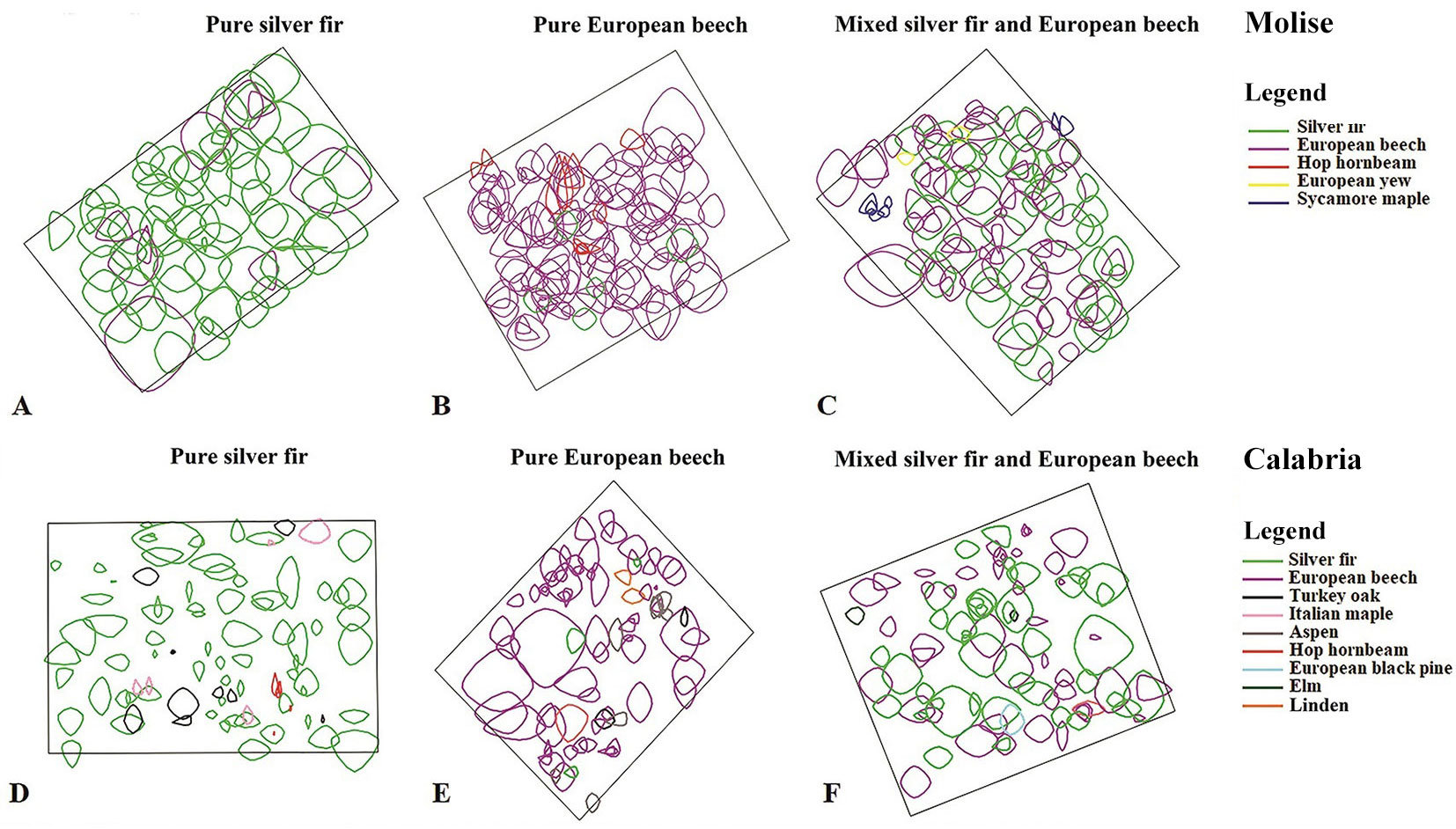
Stands have similar exposure (North, North-West) and slope, between 20° and 28°, except for pure stand of silver fir in Calabria (37°), but differ for soil type and depth. In Molise, the main soil type is Rendzina, whereas in Calabria is Distrudept. Soils in pure stands of European beech are relatively shallow and rocky in comparison with those of the mixed-species stands and the pure stands of silver fir (Tab. 1). All triplets have not been thinned since 1979-1980.
Field measurements
At the beginning of summer 2017, the structural attributes and dendrometric parameters were determined in each stand, for all living trees with a diameter ≥ 7 cm, through the Field-Map® technology and software (Institute of Forest Ecosystem Research Ltd. Jílové u Prahy, Czech Republic - ⇒ http://www.fieldmap.cz/). For each tree, the stem diameter at 1.30 m, the tree height, the crown radius in the four cardinal directions, the tree species, and the spatial position were recorded. Cores were sampled only from dominant and sub-dominant trees of European beech and silver fir.
Two cores per tree were collected at breast height (1.3 m) perpendicularly to the slope, in order to avoid compression wood (for silver fir) and tension wood (for European beech). Ring-width series were measured with a LINTAB measuring table with an accuracy of 0.01 mm, equipped with a Leica MS5® stereoscope (Leica Microsystems, Wetzlar, Germany). The analysis was performed with the TSAP software package ([57]) and subsequently were cross-dated using COFECHA ([31], [28]) to assess the measurement and dating accuracy. We successfully cross-dated 526 cores in total.
The raw chronologies were converted into tree basal area increment (BAI), according to the following standard equation (eqn. 1):
where r is the radius of the tree and t is the year of tree-ring formation.
Individual-based competition indices
To analyze the competitive dynamics within the stands, we used distance-dependent competition indices. Generally, although not for stands with stationary structures ([70]), these indices are considered useful for explaining competitive interactions between individual trees ([62]). Distance-dependent indices, in addition to the individual tree sizes, may also provide spatial information on the competitive status of an individual tree ([68]).
To calculate the distance-dependent indices, several methods can be applied ([30], [17], [66]). Three distance-dependent competition indices (CI) based on stem diameter at breast height (DBH) were used in this study: Hegyi ([30] - eqn. 2), Martin & Ek ([40] - eqn. 3) and Rouvinen & Kuuluvainen ([58] - eqn. 4).
The competition indices were calculated using the following formulas:
In the above equations, DBHi is the diameter at breast height of the subject tree i, DBHj the diameter at breast height of the competitor tree j, distij the distance between the subject tree i and the competitor tree j, and n is the number of competitors in the neighborhood zone.
In order to calculate the competition indices only European beech and silver fir were considered, being the overwhelming species in the sampled stands. The competition indices were calculated at the beginning of the growing period (2011), which allowed us to build the mixed model (see below). In order to obtain the DBHs for 2011, the basal area increments of all European beech and silver fir trees in mixed-species and pure stands were reconstructed. In detail, basal area increments in the last 5 years (2011-2016) and the thickness of the bark (in some cores also the beginning of growth in 2017) of all cored trees were considered. The 5-year basal area increment (BAI5years) and bark thickness were adapted to DBH measured in 2017 of cored trees, using linear models for each site and each species. This allowed to estimate the 5-year basal area increment (BAI5years) and the bark thickness of non-cored trees, and values were sequentially subtracted from the DBH measured in 2017 to reconstruct DBH in 2011 of non-cored European beech and silver fir trees.
The selection of competitors was determined by the fixed radius method. The competition indices (CI) values were obtained using neighborhood radii ranging 1-10 m (in 1 m increments) from the subject tree. Linear regression models were used to identify the optimal search radius and the corresponding CI was used in the subsequent analyses. Linear regression models were tested and evaluated on the basis of the constancy of variances (p > 0.05), adjusted r2, and significance of the correlation (p < 0.1). The BAI5years-CI relationship for silver fir trees was calculated only for trees with DBH ≥ 20 cm, given that the cored trees had a DBH ≥ 20 cm. European beech trees, however, had both a DBH < 20 cm and DBH ≥ 20 cm. Therefore, to assess whether the optimal search radius in European beech varied between the considered DBH values, the BAI5years-CI relationship was calculated both for all cored trees (20 cm > DBH ≥ 20 cm) and for cored trees with a DBH ≥ 20 cm only.
For each tree, the CI was divided into inter-specific and intra-specific competition CI, when trees of different species from the focal tree species (CIinter) and trees of the focal species (CIintra) were considered, respectively.
Modelling approach
To investigate the effects of competitive interactions between mixed-species and pure stands, a mixed-effects model was applied. Specifically, the basal area growth of the individual trees (cm2 year-1) was chosen as the dependent variable, being less influenced by the age of the tree than the ring-width time series ([8]). A natural logarithmic transformation of the original dependent variable was performed, usually being one of the best variables reflecting the non-linear tree growth curve ([71]).
In the model, we considered the hypothesis that the basal area increment of the subject tree is a function of the tree size (SIZE), of the competition (COMP), of site characteristics (SITE), and of species in admixture (MIXTURE - eqn. 5 - eqn. 9:
where the dependent variable is the 5-year basal area increment (BAI5years, cm2). The four independent variables were defined as:
where the variable DBH (cm) is that measured at the beginning of the growing period (2011), and Age is the tree age in the year 2011;
where the competition variables are the intra-specific (CIintra) and inter-specific (CIinter) competition indices calculated at the beginning of the growing period (2011) of European beech and silver fir trees in mixed-species and pure stands; mix-pure is the dummy variable, with mix-pure = 0 for pure species stands and mix-pure = 1 for mixed-species stands;
where the site variables are topographic features of the stand, including elevation (ALT), and slope and depth of the soil;
where BAbeech and BAfir are the basal area (measured at the beginning of the growing period - 2011) of European beech and silver fir, respectively.
The models (silver fir mixed-pure and European beech mixed-pure) were developed with competition variables conveying intra- and inter-specific species interactions. The models for silver fir in mixed-species and pure stands were built with 55 trees, while the models for European beech in mixed-species and pure stands were built with 50 trees. To test whether there was variability in the basal area increment between stands, we fitted a linear mixed model, including the stand (intercept) as a random effect.
Prior to fit the models through the “lmer” function of the R package “lme4” ([5]), the incidence of multicollinearity between the independent variables was evaluated, with the “VIF” function of the R package “car” ([24]). The threshold used to assess multicollinearity was VIF = 2.5 ([19]). After sub-setting the independent variables according to multicollinearity, models were fitted through the likelihood criterion (ML - [46]) and compared using the Akaike’s information criterion (AIC - see Appendix 1 in Supplementary material). We discarded redundant explanatory variables through the “chisq.test” function of the standard “statistics” package in R. The best models were fitted using the limited maximum likelihood method (REML), with a significance threshold of p < 0.05. Normality and homogeneity of the residuals of the final models were visually inspected by plotting the residuals against fitted values ([72] - see Fig. S1 in Supplementary material). Conditional R2 was obtained through the “r.squared GLMM” function of the “MuMIn” package ([4]).
Statistical analyses
The differences between mixed-species and pure stands, in terms of competition and basal area increment, were assessed through the analysis of variance (ANOVA). Analyses were performed for each stand to identify significant correlations between the competition indices calculated at the beginning of the growing period (inter- and intra-specific) and basal area increment. For multiple comparisons of subgroups of variables, Tukey’s post-hoc test was performed. All statistical analyses were performed using the R software package (⇒ https://www.r-project.org/).
Results
Stand structure
Stands showed differences both in tree age (Tab. 2) and stem diameter distribution (Fig. 3). Age differences were observed between Molise and Calabria, as generally younger trees were found in Calabria (Tab. 2).
Tab. 2 - Tree age (range), average diameter at breast height (DBH) of sampled trees, 5-year basal area increment (BAI5years) with the standard deviation (SD), and number of trees and cores used in the study.
| Site | Stand | Tree age (yrs) |
DBH ± SD (cm) |
BAI5years ± SD (cm2 yr-1) |
Cores/trees |
|---|---|---|---|---|---|
| Molise | beechpure | 50-140 | 22 ± 6 | 6.71 ± 5.32 | 110/60 |
| firpure | 57-171 | 42 ± 8 | 13.58 ± 18.55 | 86/44 | |
| beechmixed | 50-119 | 27 ± 11 | 17.26 ± 17.32 | 46/31 | |
| firmixed | 76-161 | 59 ± 14 | 31.24 ± 18.26 | 38/19 | |
| Calabria | beechpure | 50-109 | 41 ± 15 | 13.21 ± 13.35 | 78/46 |
| firpure | 39-82 | 37 ± 12 | 15.57 ± 14.52 | 82/43 | |
| beechmixed | 53-128 | 40 ± 14 | 14.53 ± 14.45 | 38/25 | |
| firmixed | 51-108 | 52 ± 16 | 27.53 ± 17.12 | 46/28 |
Fig. 3 - DBH distribution classes of all species in pure and mixed-species stands in Molise (A: silver fir; B: European beech; C: mixed-species) and in Calabria (D: silver fir; E: European beech; F: mixed-species). The size of DBH classes was 10 cm both in pure and mixed-species stands. Tree species are referred to by colours in the legend.
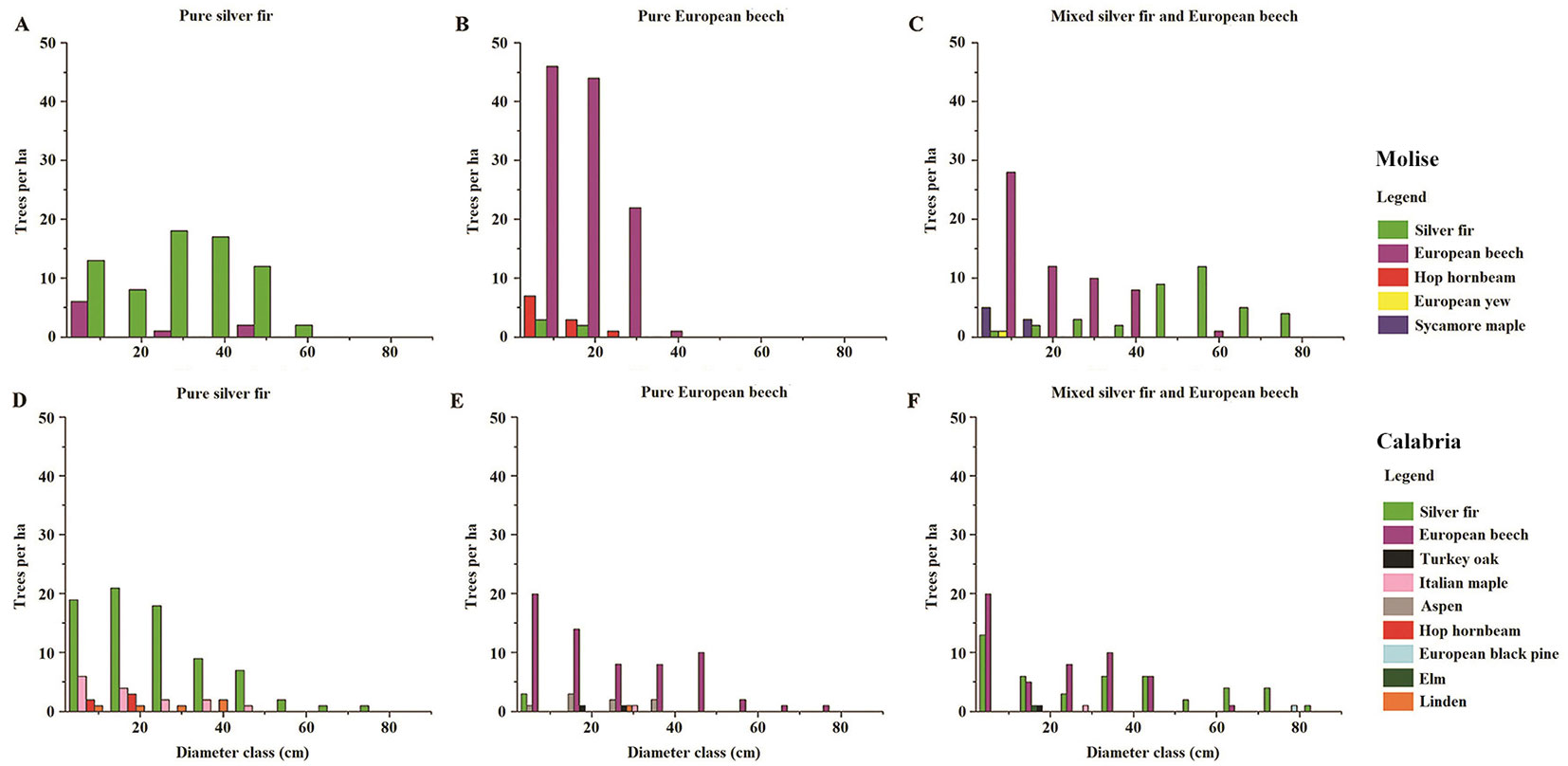
Considering all the species in each stand, diameter distribution showed a negative exponential pattern in almost all sites, except for pure silver fir in Molise (Fig. 3). DBH had a continuous distribution, with a maximum value of 80 cm, except for silver fir (maximum DBH of 60 cm - Fig. 3A) and European beech (40 cm - Fig. 3B) in pure stands, in Molise. Variation in tree height also showed differences between the stands in Molise and those in Calabria (Fig. S2 in Supplementary material). In Molise, trees of silver fir were taller than those of European beech. In Calabria, instead, only in mixed-species stand, trees of silver fir grew taller than those of European beech, the first species occupying the dominant layer while the second the sub-dominated layer.
Selection of competition index and identification of optimal search radius
Among the competition indices that were tested, the Hegyi index (CIHeg), based on the relationship between the stem radial diameters and the distances between the subject tree and the competitors, explained the highest proportion of variability in basal area increment (Tab. 3). The Martin and Ek index (CIME) was the second best, while the Rouvinen and Kuuluvainen (CIRK) was the least appropriate for these stands. CIHeg explained a high proportion of variability in basal area increment for European beech (up to 72% in mixed-species and pure stands) than for silver fir, except for European beech in mixed-species stand in Calabria (from 26% to 39%).
Tab. 3 - Range of R2 calculated for relationships between the 5-years basal area increment (BAI5years) of species and individual competition indices. (CIHeg): Hegyi index; (CIME): Martin and Ek index; (CIRK): Rouvinen and Kuuluvainen index.
| Site | Competition Index |
firpure | beechpure | firmixed | beechmixed |
|---|---|---|---|---|---|
| Molise | CI Heg | 0.15-0.40 | 0.08-0.50 | 0.16-0.47 | 0.44-0.72 |
| CI ME | 0.05-0.39 | 0.00-0.41 | 0.14-0.40 | 0.28-0.63 | |
| CI RK | 0.03-0.21 | 0.00-0.20 | 0.01-0.19 | 0.02-0.14 | |
| Calabria | CI Heg | 0.12-0.45 | 0.29-0.58 | 0.00-0.53 | 0.26-0.39 |
| CI ME | 0.03-0.44 | 0.13-0.21 | 0.00-0.39 | 0.08-0-33 | |
| CI RK | 0.00-0.43 | 0.04-0.08 | 0.01-0.20 | 0.00-0.03 |
The optimal search radii were selected according to the highest prediction accuracy of the linear regression models (i.e., constancy of variances, adjusted r2, and significance of the correlation - Tab. 4) obtained from the selected competition index (CIHeg). The optimal search radii varied according to the species and stands examined (Tab. 4). In Molise, the highest accuracies of the linear regression model for silver fir were reached for optimal search radii of 10 m. In Calabria, silver fir in pure and mixed-species stands, however, reached the highest prediction accuracy of the linear regression model for an optimal search radius of 9 m and 8 m, respectively.
Tab. 4 - Optimal search radii selected on the basis of the best values of linear regression models, considering DBH ≥ 20 cm. (CV): Constancy of variances (p > 0.05); (r2): adjusted r2; (p): significance of the correlation (p < 0.1); (n): number of tree samples; R (m): optimal radius.
| Sites | Stands | CV | r 2 | p | n | R (m) |
|---|---|---|---|---|---|---|
| Molise | beechpure | 0.80 | 0.47 | < 0.01 | 22 | 7 |
| firpure | 1.76 | 0.36 | <0.01 | 18 | 10 | |
| beechmixed | 0.13 | 0.69 | <0.01 | 13 | 9 | |
| firmixed | 0.45 | 0.43 | <0.01 | 17 | 10 | |
| Calabria | beechpure | 0.78 | 0.38 | 0.03 | 10 | 7 |
| firpure | 3.14 | 0.39 | <0.01 | 14 | 9 | |
| beechmixed | 3.38 | 0.30 | 0.07 | 9 | 5 | |
| firmixed | 1.12 | 0.46 | 0.02 | 9 | 8 |
Considering only trees with DBH ≥ 20 cm, in Molise the optimal search radius for European beech in pure and mixed-species stands was 7 m and 9 m, respectively (Tab. 4). In Calabria, the optimal search radii for European beech in pure and mixed-species stands was 7 m and 5 m, respectively (Tab. 4). The same optimal search radii were identified also for all sampled trees inside the stand (Tab. S1).
Individual-level competition and BAI of mixed-species vs. pure stands
Tab. 5shows the competition indices (CIHeg) and the 5-years basal area increment calculated for each stand. Both intra-specific (CI2011_intra) and inter-specific (CI2011_inter) indices showed higher levels of competition in pure stands of Molise than in pure stands of Calabria. European beech in mixed-species stands had higher values of intra- and inter-specific competition in Molise (0.44 ± 0.25 and 0.99 ± 0.32, respectively) than in Calabria (0.30 ± 0.33 and 0.48 ± 0.35, respectively). In contrast, silver fir in mixed-species stands showed higher values of intra- and inter-specific competition in Calabria (0.83 ± 0.43 and 0.49 ± 0.26, respectively) than in Molise (0.49 ± 0.39 and 0.34 ± 0.11, respectively).
Tab. 5 - Intra- and inter-specific CIHeg in the mixed-species stands for European beech and silver fir at the beginning of the growing period (CI2011_intra and CI2011_inter) and basal area increment of the last 5 years (BAI5years) for the analyzed species.
| Site | Stand | CI2011_intra | CI2011_inter | BAI5years (cm2 yr-1) |
|---|---|---|---|---|
| Molise | beechpure | 1.83 ± 0.57 | 0.09 ± 0.14 | 8.57 ± 4.59 |
| firpure | 1.89 ± 0.57 | 0.10 ± 0.12 | 10.12 ± 8.13 | |
| beechmixed | 0.44 ± 0.25 | 0.99 ± 0.32 | 19.52 ± 10.93 | |
| firmixed | 0.49 ± 0.39 | 0.34 ± 0.11 | 33.10 ± 19.55 | |
| Calabria | beechpure | 0.79 ± 0.54 | 0.02 ± 0.02 | 22.98 ± 13.01 |
| firpure | 1.94 ± 0.47 | 0 | 14.23 ± 9.42 | |
| beechmixed | 0.30 ± 0.33 | 0.48 ± 0.35 | 17.13 ± 17.36 | |
| firmixed | 0.83 ± 0.43 | 0.49 ± 0.26 | 28.13 ± 17.17 |
The 5-years basal area increment (BAI5years) in pure stands was higher in Calabria than in Molise. Silver fir (33.10 ± 19.55) and European beech (19.52 ± 10.93) in mixed-species stands of Molise showed higher BAI5years values than in pure stands. In Calabria, instead, only silver fir in mixed-species stand (28.13 ± 17.17) had higher BAI5years values than in pure stand (14.23 ± 9.42).
Growth models
The selection of the best intra-specific and inter-specific interaction models was based on the lowest values of AIC (Tab. S2 and Tab. S3 in Supplementary material).
Intra-specific interactions models
The best intra-specific interaction models for silver fir mixed-pure, four independent variables were used: CI2011_intra, slope, DBH2 and mix-pure (Tab. S4). In the model for European beech mixed-pure, six independent variables were used: CI2011_intra, BAbeech, age, soil depth, DBH2 and mix-pure (Tab. S4). In the final model for silver fir mixed-pure, only CI2011_intra affected the basal area increment significantly (Tab. 6); the basal area increment decreased with increasing CI2011_intra. Furthermore, the model showed differences between stands, with R2 of 0.51, when all effects were included, and R2 of 0.50, when only fixed effects were considered.
Tab. 6 - Parameter estimates and fitting statistics for the best intra-specific interaction models through the limited maximum likelihood method (REML). (CI2011_intra): intra-specific interactions; (DBH2): diameter at breast height squared (cm2); (Soil_depth): depth of the soil; (VarPlot): Plot (intercept) variance; (VarRes): Residual variance; (SD): standard deviation; (AIC); Akaike’s information criterion (mixed effect).
| Stand Type |
Fixed effects | Random effects | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Factor | Est. | SE | Prob. | VarPlot (SD) |
VarRes (SD) |
R2m | R2c | AIC | |
| Silver fir mixed-pure |
CI2011_intra | -0.693 | 0.102 | <0.001 | 0.011 (0.106) |
0.465 (0.682) |
0.500 | 0.510 | 126.537 |
| European beech mixed-pure |
CI2011_intra | -0.667 | 0.122 | <0.001 | 0.000 (0.000) |
0.513 (0716) |
0.600 | 0.600 | 147.014 |
| DBH 2 | 0.595 | 0.12 | <0.001 | ||||||
| Soil_depth | -0.45 | 0.126 | <0.001 | ||||||
In the final model for European beech mixed-pure, CI2011_intra, DBH2 and Soil_ depth affected the basal area increment significantly (Tab. 6); the basal area increment decreased with increasing CI2011_intra and Soil_depth. DBH2 showed direct responses to basal area increment. Furthermore, the model did not show differences between stands, with an R2 of 0.60, both when all effects were included and if only fixed effects were considered.
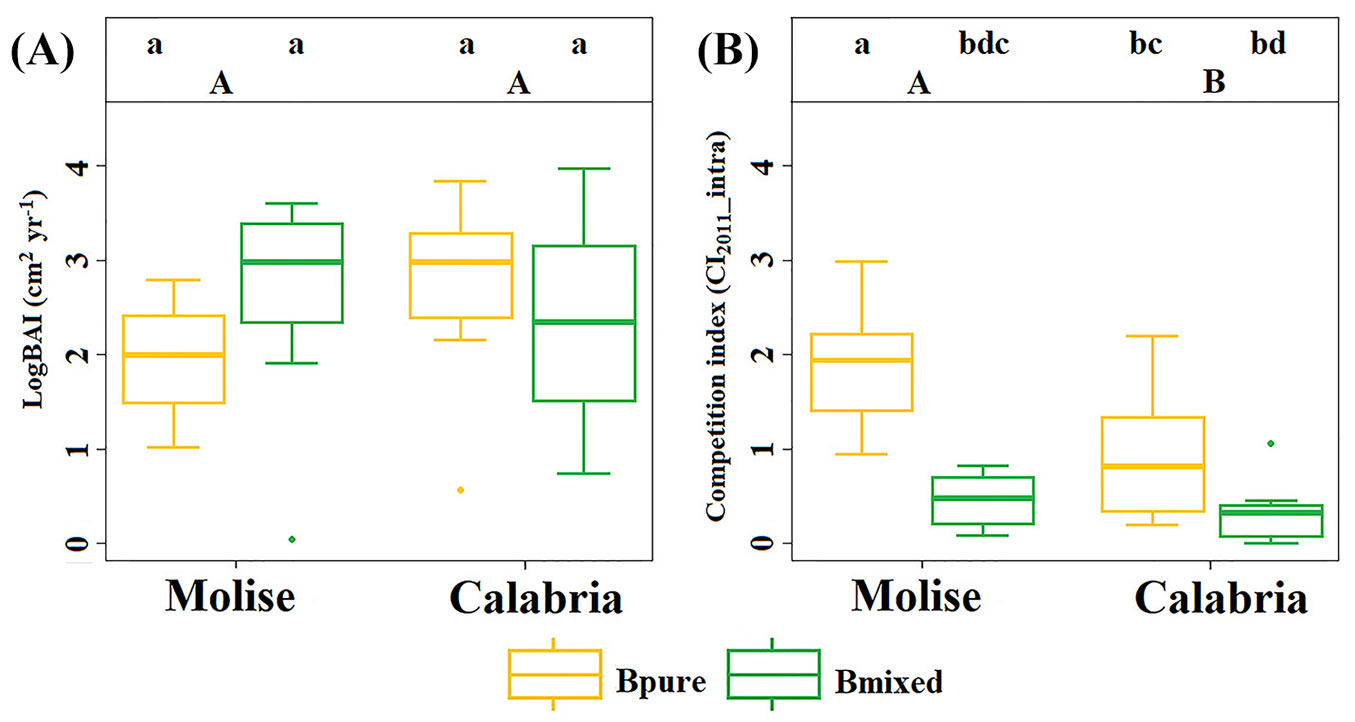
Basal area increment (BAI5years) revealed that only silver fir in mixed-species stand of Molise differed significantly from the corresponding pure stand (p < 0.05 - Fig. 4A). In both Molise and Calabria, the basal area increments of European beech did not show significant differences between mixed-species and pure stands (Fig. 5A). The competition indices (CI2011_intra), on the other hand, differed significantly between pure and mixed-species stands of Molise and Calabria (p < 0.05 - Fig. 4B, Fig. 5B). In Molise and Calabria, silver fir in mixed-species stands differed significantly from silver fir in pure stands (p < 0.05 - Fig. 4B). Values of CI2011_intra were higher in mixed-species and pure stands of European beech in Molise than in Calabria (Fig. 5B).
Fig. 4 - Basal area increment (logBAI) and competition indices (CI2011_intra) of silver fir in mixed-species and pure stands, considering the last 5 years and the beginning of the growing period. Smixed refers to mixed-species stands (Calabria and Molise) and Spure to pure stands (Calabria and Molise). Capital letters indicate differences between the sites for basal area increment and competition indices, and small letters indicate differences between species (ANOVA and post-hoc tests, p < 0.05). The vertical lines represent the “whiskers” for the 5th and 95th percentiles of data distribution.
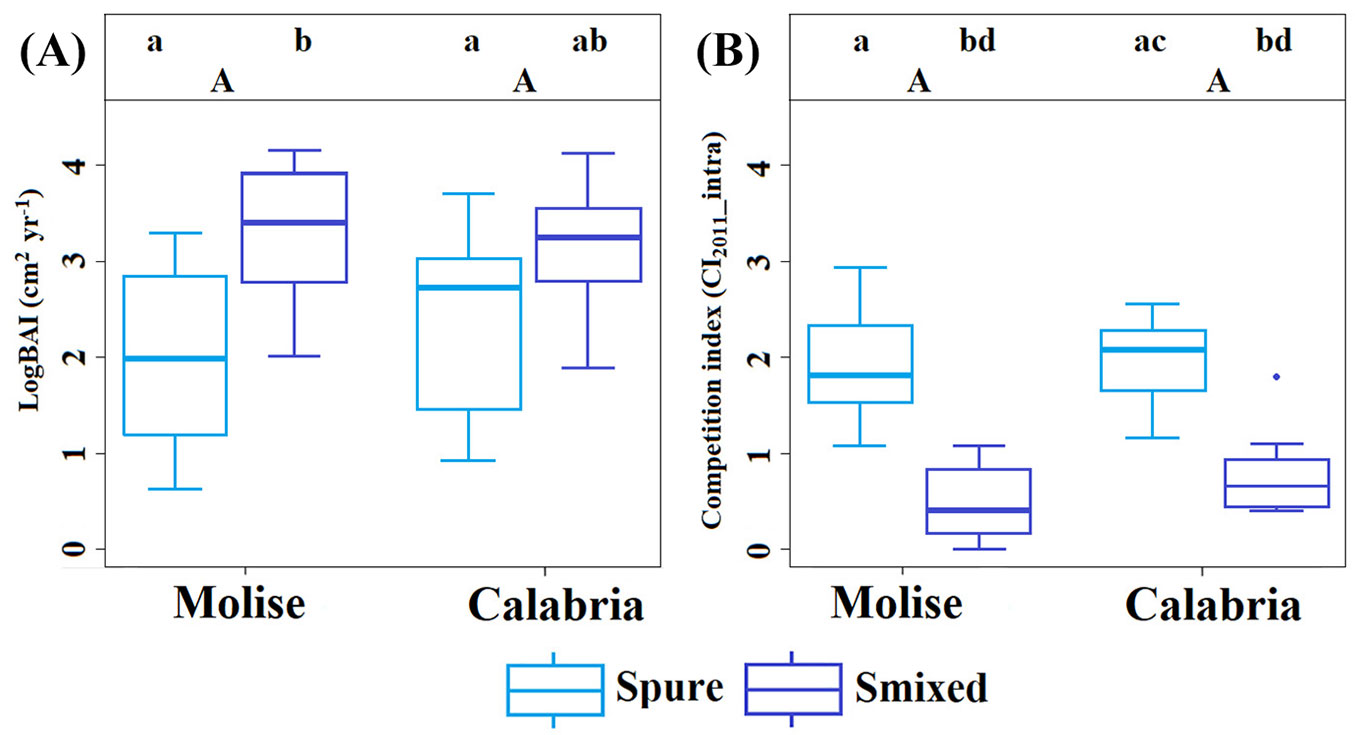
Fig. 5 - Basal area increment (logBAI) and competition indices (CI2011_intra) of European beech in mixed-species and pure stands, considering the last 5 years and the beginning of the growing period. Bmixed refers to mixed-species stands (Calabria and Molise) and Bpure to pure stands (Calabria and Molise). Capital letters indicate differences between sites for each competition index, intra- and inter-specific (ANOVA and post-hoc tests, p < 0.05). The vertical lines represent the “whiskers” for the 5th and 95th percentiles of data distribution.
Inter-specific interactions models
Even for inter-specific interactions, the best models for silver fir mixed-pure, four independent variables were used: CI2011_inter, slope, DBH2 and mix-pure (Tab. S5). In the model for European beech mixed-pure, six independent variables were used: CI2011_inter, BAbeech, age, Soil_depth, DBH2 and mix-pure (Tab. S5). In the final model for silver fir mixed-pure, only DBH2 significantly affected the basal area increment (Tab. 7). DBH2 showed direct responses to tree basal area increment. The model showed also differences between stands, with an R2 of 0.46, when all effects were included, and R2 of 0.36, when only fixed effects were considered.
Tab. 7 - Parameter estimates and fitting statistics for the best inter-specific interaction models through the limited maximum likelihood method (REML). (DBH2): diameter at breast height squared (cm2); (Age): tree age (years); (Soil_depth): depth of the soil; (BAbeech): basal area occupied by competitors of European beech; (VarPlot): Plot (intercept) variance; (VarRes): Residual variance; (SD): standard deviation; (AIC); Akaike’s information criterion (mixed effect).
| Stand Type |
Fixed effects | Random effects | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Factor | Est. | SE | Prob. | VarPlot (SD) |
VarRes (SD) |
R2m | R2c | AIC | |
| Silver fir mixed-pure |
DBH 2 | 0.562 | 0.114 | <0.001 | 0.093 (0.305) |
0.479 (0.692) |
0.360 | 0.460 | 130.837 |
| European beech mixed-pure |
DBH 2 | 0.678 | 0.125 | <0.001 | 0.000 (0.000) |
0.564 (0.751) |
0.570 | 0.570 | 141.972 |
| Age | 0.192 | 0.113 | 0.097 | ||||||
| Soil_depth | -0.456 | 0.135 | 0.001 | ||||||
| BA beech | -0.541 | 0.122 | <0.001 | ||||||
In the final model for European beech mixed-species and pure stands, BAbeech, DBH2 and Soil_depth affected the basal area increment significantly (Tab. 7); the basal area increment decreased with increasing BAbeech and Soil_depth. DBH2 showed direct responses to basal area increment. The model did not show differences between stands, with an R2 of 0.57, both when all effects were included and if only fixed effects were considered.
Discussion
Influence of intra-specific interactions on trees basal area increment in mixed-species and pure stands
We analyzed the effect of interactions between tree species on the basal area increment of uneven-aged mixed-species and pure stands of European beech and silver fir, implementing growth models and competition indices at the individual tree level. The final models showed strong influence of intra-specific interactions (CI2011_intra) on the basal area increment (BAI5years) of both species (Tab. 6). The dummy variable mix-pure (Tab. S4 in Supplementary material) indicates the influence of variables related to the pure and mixed-species stands on the basal area increment (BAI5years). The absence of significance of this variable, suggests that intra-specific interactions (CI2011_intra) had high influence on the basal area increment (BAI5years) of both species in mixed-species and pure stands. Therefore, European beech and silver fir in mixed-species and pure stands were more affected by competition with conspecific than allospecific neighbors. Several studies indicated that European beech is a strong self-competitor ([49], [51], [10]), with high intra-specific competition for above-ground ([55]) and below-ground ([56], [18], [44]) resources. In literature studies on silver fir focused mainly on drought response ([34]), while are rather scarce in terms of competitive interactions between mixed-species and pure stands.
Analysis of variance (ANOVA) showed that the influence of intra-specific interactions (CI2011_intra) was higher in pure stands of silver fir and European beech than in mixed-species stands (Fig. 4B, Fig. 5B), supporting our first hypothesis. In Molise, ANOVA highlighted significantly higher increase in the basal area increment (BAI5years) of silver fir in mixture than in monoculture (Fig. 4A). In accordance with our results, Toïgo et al. ([65]) observed an increase in the basal area increment of silver fir more marked in mixed-species stands than in pure stands. Crown architecture, vertical positioning within the canopy and root morphology may lead to differences in resources accessibility between pure and mixed-species stands, and, therefore, a more efficient use of water and light ([36], [23]). However, in our case, higher basal area increments of silver fir trees in the mixed-species stand in Molise, compared to the corresponding pure stand (Fig. 4A) might depend on differences in stand structure (Fig. 3). Silver fir trees in the mixed-species stand were bigger (DBH > 50 cm) than in the pure stand (DBH < 50 cm), which might explain the reduction in competitive interactions and, consequently the higher basal area increment of silver fir trees in the mixed-species stand.
European beech, instead, showed high tree-to-tree competition in pure stands, most probably related to the low self-tolerance of this species ([47]), which is attenuate in mixed-species stands ([50]). For example, Mölder & Leuschner ([45]) found a lower impact of intra-specific interactions on the basal area increment of European beech in mixed-species forests than in pure stands. This less negative impact of intra-specific interactions was related to the lower number of European beech trees among competitors, confirming that European beech is a strong self-competitor. High levels of intra-specific interactions may also depend on high degree of canopy closure in pure stands of European beech ([49]). In our case, significantly higher intra-specific competition levels (CI2011_intra) were observed in pure stand of European beech, in Molise, which had closed canopy (Fig. 2B), in comparison with the corresponding one in Calabria (Fig. 2E). A closed-canopy may derive from different spatial arrangement of trees, random (in Molise) vs. clustered (in Calabria). In fact, mixed-species and pure stands of European beech showed higher influence of intra-specific interactions (CI2011_intra) in Molise than in Calabria (Fig. 5B). Gap recruitment of small competing trees leading to clustered regeneration may occur more frequently in drier sites ([61]). Edaphic heterogeneity and local aridity may also cause clumped spatial distribution of trees.
Several studies found that the stand-level spatial arrangement of trees may influence the competitive intensity between species ([25]), especially in uneven-aged and uneven-structured stands ([69]), as well as in those stands with trees differing in crown morphology, crown efficiency and space occupation ([42]). This would suggest that the forest structure (e.g., spatial arrangement, crown morphology, etc.) may regulate tree-to-tree competition and tree growth in structurally diverse forests ([25]).
Intra- and inter-specific interactions in mixed-species stand
Inter-specific interaction (Tab. 7), was not the most important variable to explain the competitive interactions in mixed stands. This suggests that inter-specific interactions (CI2011_inter) in mixed-species stands did not significantly influence the basal area increment, supporting our second hypothesis. Gazol et al. ([27]) observed higher growth rates in silver fir admixed with European beech. Other studies of intra- and inter-specific interactions on fir-beech mixtures indicated either no effects of mixing on basal area increment of either species ([18], [59]) or positive mixing effects for beech, whereas positive effects for silver fir could be restricted to sites of lower productivity ([65]). A negative influence of intra-specific interactions (in comparison with inter-specific interactions) on the growth of European beech was confirmed in the present study. However, the effect of these interactions on the growth of silver fir needs to be further explored, in view of contradicting results of previous studies ([18], [10], [44]). Indeed, Mina et al. ([44]) found that the growth of silver fir was negatively affected by the presence of European beech. In contrast, Del Río et al. ([18]) revealed that silver fir was more affected by intra-specific than inter-specific competition, suggesting the presence of some degree of complementarity between European beech and silver fir. Bošela et al. ([10]) noted that the growth of silver fir was positively influenced by admixture with European beech, showing facilitating effect or a reduction in competition of European beech on silver fir. Evidence of inter-specific facilitation or reduction in tree-to-tree competition could be linked to differences in functional traits and phenological patterns of silver fir vs. European beech ([18]). For example, compartmentalization of resource utilization may allow a different strategy for root water uptake ([34]). Complementary crown architectures and contrasting physiological adaptations may facilitate canopy expansion (European beech) and improve crown light interception in spring and autumn (silver fir - [38]). This suggests that, in mixed-species stands, tree species with complementary traits may increase the exploitation of resources in comparison with those species with similar traits ([55]). According to Pretzsch & Schütze ([55]), improved resource use efficiency (e.g., light, water and nutrients) in mixed-species stands is attained when a complex vertical structure (small and large individuals of both species) occurs. In our case, the absence of inter-specific interactions (CI2011_inter) can be related to the smaller diameter in European beech than in silver fir, in the mixed-species stands of Molise and Calabria (Fig. 3). These differences between European beech and silver fir were also observed for the variation in tree height, silver fir being taller than European beech (Fig. S2 in Supplementary material). Therefore, the sub-dominant structural position of European beech within mixed-species stands may determine a reduction in competition for silver fir.
Overall, forest management that promotes multi-layered uneven-sized species mixtures might allow maintaining these Mediterranean mountain forests continuously, increasing their resilience and productivity. Although the importance of species mixture is recognized, management of mixed-species uneven-aged forests at the range edge of species distribution needs to be optimized for effective application. Indeed, the sustainability of species mixture might become an issue in drought-prone environments ([15]), in which a stable species distribution could be negatively affected by increasing disturbance frequency and intensity. For a continuous delivery of ecosystem services of mixed-species mountain forests, the selective felling might regulate the occupation of the growing space of the species and enhance the development of the stands towards a multi-layered structure, making them more stable against natural hazards and ensuring the protective functions of the forests, for soils and biodiversity, as well as the maintenance of carbon stocks.
Conclusion
The growth models showed that intra-specific interactions influenced the basal area increment more than inter-specific interactions. In particular, intra-specific interactions negatively influenced the basal area increment of European beech and silver fir, both in pure and mixed-species stands of Molise and Calabria. In accordance with our first hypothesis, higher intra-specific interactions were observed in pure stands of both species than in corresponding mixed-species stands. Silver fir showed relatively lower levels of intra-specific competition, with higher basal area increment in mixed-species stands. This higher basal area increment of individual trees in mixed-species stands can be explained by the distinct stand structure, which may have increased space occupation and improved resource utilization. For European beech, high values of intra-specific interactions could be linked to its low self-tolerance and to the spatial arrangement of trees within the stands, following canopy closure. The absence of inter-specific interactions in mixed-species stands was probably related to the sub-dominant position of European beech, which may have reduced the benefit derived from niche separation and complementarity for silver fir. These results support our second hypothesis, providing useful insights on the structural and functional dynamics that occur in uneven-aged mixed-species forests vs. pure stands of European beech and silver fir. This suggest that mixtures of European beech and silver fir, a natural species combination in mountain regions, should be promoted as an option for mountain ranges in southern Europe. However, further studies with a larger number of samples are needed to draw more definitive conclusions, so as to help evaluate forest adaptation measures under global change scenarios.
Acknowledgements
The authors are grateful to Carabinieri Forestali of Petilia Policastro (KR, Italy) for their help in selecting the study sites. This study generated from the COST (European Cooperation in Science and Technology) Action CLIMO (“Climate-Smart Forestry in Mountain Regions” - CA15226) financially supported by the EU Framework Programme for Research and Innovation HORIZON 2020.
References
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Gscholar
Online | Gscholar
CrossRef | Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Vittorio Garfì 0000-0002-6980-2920
Di Febbraro Mirko 0000-0001-8898-7046
Department of Biosciences and Territory, University of Molise, c.da Fonte Lappone, 86090 Pesche, IS (Italy)
Michele Dalponte 0000-0001-9850-8985
Lorenzo Frizzera 0000-0003-3150-8395
Damiano Gianelle 0000-0001-7697-5793
Department of Sustainable Agro-ecosystems and Bioresources, Research and Innovation Centre, Fondazione E. Mach, v. E. Mach 1, 38010 San Michele all’Adige, TN (Italy)
Roberto Tognetti 0000-0002-7771-6176
The EFI Project Centre on Mountain Forests (MOUNTFOR), v. Edmund Mach 1, 38010, San Michele all’Adige, TN (Italy)
Department of Agricultural, Environmental and Food Sciences, University of Molise, v. Francesco De Sanctis, 86100 Campobasso (Italy)
Corresponding author
Paper Info
Citation
Versace S, Garfì V, Dalponte M, Febbraro Mirko D, Frizzera L, Gianelle D, Tognetti R (2021). Species interactions in pure and mixed-species stands of silver fir and European beech in Mediterranean mountains. iForest 14: 1-11. - doi: 10.3832/ifor3476-013
Academic Editor
Giorgio Vacchiano
Paper history
Received: Apr 20, 2020
Accepted: Oct 26, 2020
First online: Jan 02, 2021
Publication Date: Feb 28, 2021
Publication Time: 2.27 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2021
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 42081
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 32672
Abstract Page Views: 4589
PDF Downloads: 4043
Citation/Reference Downloads: 6
XML Downloads: 771
Web Metrics
Days since publication: 2045
Overall contacts: 42081
Avg. contacts per week: 144.04
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Jul 2026)
Total number of cites (since 2021): 20
Average cites per year: 3.33
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Pinus halepensis Mill. in the Mediterranean region: a review of ecological significance, growth patterns, and soil interactions
vol. 18, pp. 30-37 (online: 15 February 2025)
Research Articles
Stand dynamics and natural regeneration in silver fir (Abies alba Mill.) plantations after traditional rotation age
vol. 7, pp. 313-323 (online: 08 April 2014)
Research Articles
Inter- and intra-annual patterns of seed rain in the black spruce stands of Quebec, Canada
vol. 10, pp. 189-195 (online: 13 December 2016)
Research Articles
Optimizing silviculture in mixed uneven-aged forests to increase the recruitment of browse-sensitive tree species without intervening in ungulate population
vol. 11, pp. 227-236 (online: 12 March 2018)
Research Articles
Growth dynamics and productivity of pure and mixed Castanea sativa Mill. and Pseudotsuga menziesii (Mirb.) Franco plantations in northern Portugal
vol. 7, pp. 92-102 (online: 18 December 2013)
Research Articles
Extreme climatic events, biotic interactions and species-specific responses drive tree crown defoliation and mortality in Italian forests
vol. 17, pp. 300-308 (online: 30 September 2024)
Research Articles
The response of intra-annual stem circumference increase of young European beech provenances to 2012-2014 weather variability
vol. 9, pp. 960-969 (online: 23 June 2016)
Research Articles
Patterns of genetic diversity in European beech (Fagus sylvatica L.) at the eastern margins of its distribution range
vol. 10, pp. 916-922 (online: 10 December 2017)
Research Articles
Influence of inorganic salts on biomass production, biochemical composition, and bioethanol production of Populus alba
vol. 13, pp. 566-574 (online: 07 December 2020)
Research Articles
Two Populus deltoides W.Bartram ex Marshall clones cope differentially with sodium salinity stress
vol. 18, pp. 259-266 (online: 10 October 2025)
iForest Database Search
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords







