Assessing Pinus pinea L. resilience to three consecutive droughts in central-western Italian Peninsula
iForest - Biogeosciences and Forestry, Volume 13, Issue 3, Pages 246-250 (2020)
doi: https://doi.org/10.3832/ifor3320-013
Published: Jun 19, 2020 - Copyright © 2020 SISEF
Short Communications
Abstract
Climate projections for the Mediterranean area forecast drier and hotter conditions and increasing trend in extreme climatic events such as drought. Scientific evidences reported that extreme dry spells affected the stem growth of different Mediterranean low-elevational pine forests inducing a decrease in tree resilience, defined as the capacity to resist to environmental stress and to recover pre-disturbance functioning. Despite its ecological and economic importance, thus far no study examined Pinus pinea L. (stone pine) resilience to drought events. This research reconstructed stone pine resilience by considering resistance, recovery, and the proportion of trees showing high values of both indexes of several planted stands to three consecutive spring-summer droughts occurred during the second half of the 20th century. Local climatic conditions during dry spells modulated the species resistance and recovery. In this sense, wetter conditions promoted recovery, whereas warmer spring-summer affected stone pine resistance. Moreover, spring rather than summer droughts influenced stone pine resistance and recovery, confirming the species sensitivity to climatic conditions at the beginning of the growing season. Results indicated that while recovery did not significantly changed, the species resistance diminished along the analyzed period. Furthermore, more than 60% of the examined trees were not able to reach pre-drought growth, suggesting a moderate resilience of P. pinea to adverse climatic conditions. The results contribute to improve our understanding of stone pine growth dynamics in the climate-change context of increasing aridity actually occurring in the Mediterranean area, providing useful information for the sustainable management of these natural resources.
Keywords
Introduction
Extreme climatic events such as drought represent disturbance factors for forest ecosystems, that may induce growth decline, dieback, and mortality from the tree to the stand level ([5], [1]). Forests are considered able to tolerate gradually shifting climatic conditions, whereas weather extremes may cause loss of resilience, intended as a reduction in the capacity to recover pre-disturbance functioning ([5], [16]). Expected global changes in average climatic conditions will likely modify both frequency and intensity of climatic extremes events, highlighting the need of a deeper understanding of forest resilience to drought ([15]).
During recent years, tree-ring methods successfully contributed to reconstruct forest resilience to extreme drought in different biomes ([16], [12]). For example, Juniperus spp. distributed in the Tibetan Plateau showed an overall increase in resilience to drought in the second half of the 20th century ([11]). Temperate forests in Central Europe exhibited age/site-dependent resilience to dry spells during recent consecutive climatic extremes ([31]). In moist tropical forests, resilience to drought would depend on species-specific characteristics ([25]). Regarding Mediterranean woodlands, several works analyzed resilience to dry spells of low-elevational pine forests dominated respectively by P. halepensis and P. pinaster ([19], [26]). On the other hand, thus far no study addressed this particular topic for another important Mediterranean pine such as P. pinea (stone pine), possibly because of the species drought-tolerant characteristic and its inferred high resilience to dry spells ([21]). Nevertheless, it is well known that dry climatic conditions negatively impact stone pine growth ([17]). Furthermore, drought can trigger changes in the species physiology interacting with pollution and surfactants, affecting tree health status and survival ([27], [6], [10], [4]). Stone pine plantations play important environmental and economic roles, such as dune stabilization and nut production ([18]). These forests have been long considered as ecological desert, being often undervalued by the scientific community ([3]). Nevertheless, recent studies demonstrated their utility in maintaining local biodiversity, thus calling for conservation plans of this natural resource ([3]). A deeper understanding of the stone pine response to extreme drought would provide useful information for sustainable management of these woodlands, a mandatory task in the climatic-change scenario of increasing aridity currently developing in the Mediterranean area ([15]).
For the abovementioned reasons, this research analyzed several stone pine stands located in the coast of Central Italy aiming to: (i) reconstruct resilience to drought in term of the species resistance and recovery; and (ii) examine if consecutive dry spells cumulatively affected the species resilience.
Materials and methods
Study area and dendrochronological sampling
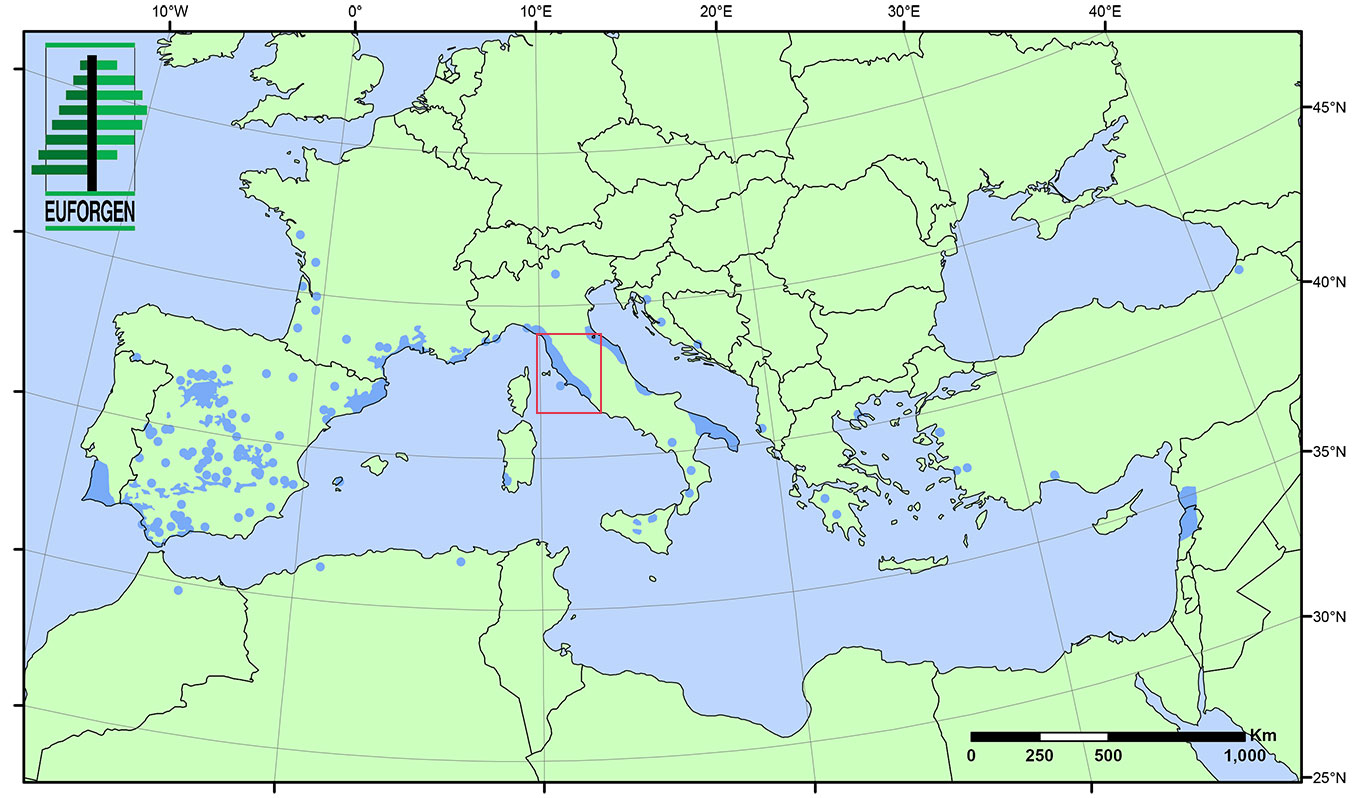
Eight stone pine stands located along the north to mid-Tyrrhenian coasts of the Italian peninsula were analyzed (Fig. 1, Tab. 1). Woodlands are pure plantations located on sandy soils and growing under typical Mediterranean climate, with summer drought and precipitation concentrated in winter months ([24], [23]).
Fig. 1 - Distribution map of Italian stone pine (Pinus pinea - [10]). The red rectangle delimits the analyzed area.
Tab. 1 - Geographical location, number of analyzed trees and time span of the tree-ring chronologies.
| Site | Lat (°N) | Long (°E) | Trees (n) | Time span |
|---|---|---|---|---|
| San Rossore | 43.72 | 10.31 | 10 | 1890-2003 |
| Cecina | 43.31 | 10.52 | 8 | 1923-2003 |
| Marina di Bibbona | 43.30 | 10.48 | 22 | 1926-1996 |
| Punta Tesorino | 43.30 | 10.48 | 23 | 1929-1996 |
| Duna Feniglia | 42.44 | 11.22 | 12 | 1925-2003 |
| Castelporziano | 41.74 | 12.40 | 20 | 1897-2003 |
| Lago di Fogliano | 41.31 | 13.03 | 6 | 1878-2004 |
| Parco del Circeo | 41.31 | 13.03 | 15 | 1945-2004 |
Six of the 8 examined plantations were sampled during 2003-2004 and earlier versions of these tree-ring chronologies have been presented in Piraino & Roig-Juñent ([23]). Ring increment data of the remaining two stands were downloaded from the ITRDB webpage (The International Tree-Ring Data Bank - ⇒ http://www.ncdc.noaa.gov/paleo/treering.html). The latter consisted of individual ring widths series from Marina di Bibbona (code ITAL030) and Punta Tesorino (ITAL032) sites, both shown in Raddi et al. ([24]). Regarding these data, we considered only tree-ring series belonging to forest stands far from the coastline, thus minimizing the possible influence of factors other than drought (e.g., shoreline erosion) on the species resilience. Details about sampling, core preparation and ring width measurements can be found in Raddi et al. ([24]) and Piraino & Roig-Juñent ([23]).
Tree growth response to drought
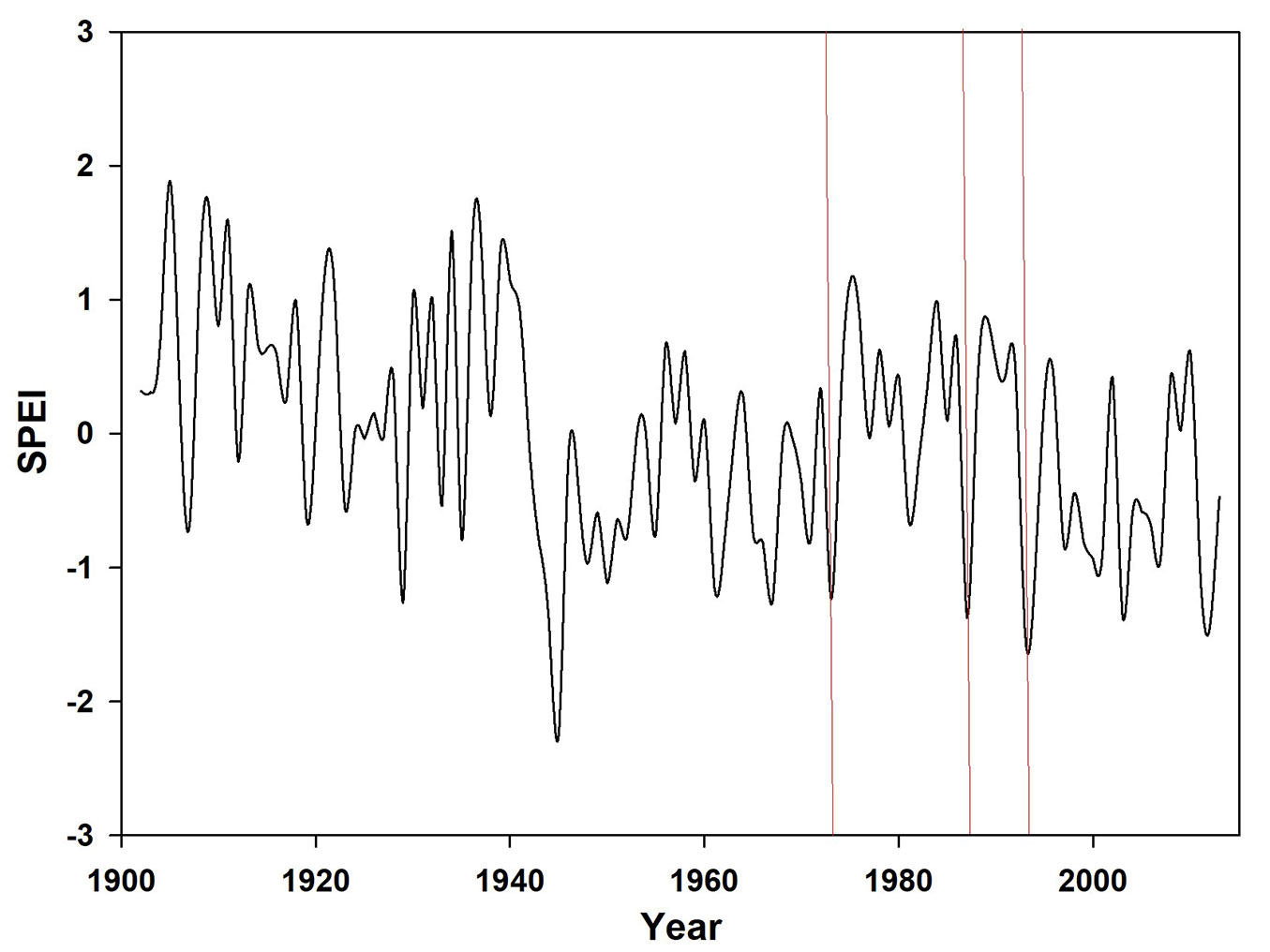
Drought was defined through the SPEI index (Standardized Precipitation Evapotranspiration Index - [29]). Based on previous researches ([22]), a regional time series of August SPEI value with a timescale of 6-months was built for the 1902-2013 period (Fig. 2). Therefore, the March-August period was considered, allowing the analysis of drought effect upon tree growth during the main drought period (summer) and the main growth season (spring). SPEI data were downloaded from the EC&D web page (⇒ http://climexp.knmi.nl/). Drought events were established as those years when SPEI index fell in the lowest 10% percentile. As Raddi et al. ([24]) ring width series ended in 1996, the three last consecutive droughts considered in this study were 1973, 1987 and 1993 (Fig. 2).
Fig. 2 - March-August SPEI of the 1902-2013 period for the examined area. Red lines represent the dry spells considered in this study.
This research considered that tree resilience is expressed by its ability to resist to and to recover from extreme climatic events ([11]). Therefore, two different indexes were calculated: resistance (Rt) and recovery (Rc - [16]). In this sense, according to Lloret et al. ([16]), resistance is “considered as reversal of the reduction in ecological performance during disturbance”, and recovery corresponds to “the ability to recover relative to the damage experienced during disturbance”. The indexes were computed as follows (eqn. 1, eqn. 2):
where Rt is the resistance index, Rc is the recovery index, BAID, BAIpreD and BAIpostD are, respectively basal area index (BAI) during the drought year, mean BAI for the 3 years before and mean BAI for the 3 years following drought. BAI series were calculated from tree-ring widths through the AGE program (DPL, Dendrochronological Programme Library - [14]). Additionally to Rt and Rc, the proportion of trees showing high resistance and recovery was calculated ([11]). Trees were considered resistant and able to recovery after drought when Rt and Rc values were higher than 1 (Rt > 1 and Rc > 1).
Statistical analyses
Drought effect upon stone pine growth was assessed at both the individual and stand level. At the tree scale, changes through time of Rt and Rc were examined by the means of ANOVA with post-hoc Tukey test ([32]). Since data were not normally distributed, indexes were transformed by the Box-Cox method ([30]). At the site level, abiotic and tree biometric influence upon Rt, Rc, Rt > 1 and Rc > 1 was analyzed through correlation functions. Climatic variables were represented by total precipitation and mean temperature of spring-summer (March-August), spring (March-May) and summer (June-August) periods. These periods were selected in order to analyze the possible interaction among spring and summer climatic conditions upon the species resistance and recovery. Tree biometric parameters were growth (mean BAI) during drought year, and age. Regarding the latter, as precise age estimation of each analyzed tree was not possible, mean age was considered as the mean site chronology length when drought episodes occurred. Local precipitation and temperature data were obtained from the “Annali Idrologici” of Regione Toscana and Regione Lazio (available respectively at ⇒ http://www.sir.toscana.it/annali-idrologici and ⇒ http://www.idrografico.regione.lazio.it/std_page.aspx-Page=annali_idrologici.htm). Statistical analyses were run through the INFOSTAT software with its interface to R ([9]).
Results
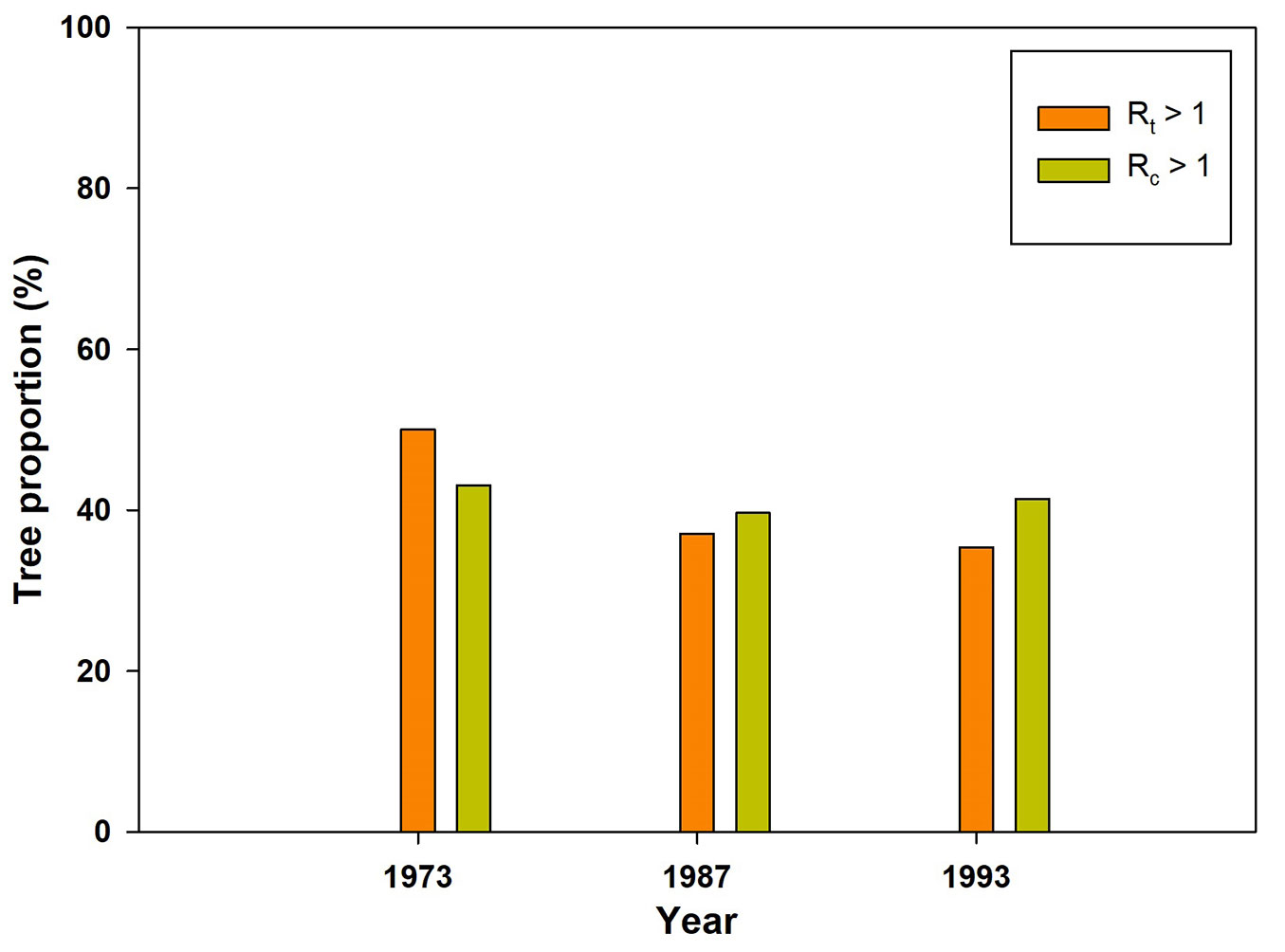
A total of 116 stone pine trees distributed along 8 sites were analyzed in this research. ANOVA showed that only Rt significantly changes along the three consecutive drought events, being values statistically lower in 1987 than in 1973 (F = 5.61, p = 0.0040 - Fig. 3). Results emerged considering proportion of trees showed a decreasing trend for the Rt > 1 index (Fig. 4). Indeed, while Rc > 1 values are constant, being respectively 43% 40% and 41% in the 1973, 1987, and 1993 dry spells, Rt > 1 showed a tendency towards lower values along the three drought events, corresponding to 50% (1973), 37% (1987) and 35% (1993) of the analyzed trees (Fig. 4).
Fig. 3 - Box plots for resistance (Rt) and recovery (Rc) index between the different considered drought years. Each box shows the values within one interquartile distance (ID 25 % above and below the median). The median is shown as a black bar. Whiskers represent values of 1.5 times the IDs and are shown as black lines. Circles represent outliers. Statistical differences are reported in the main text.
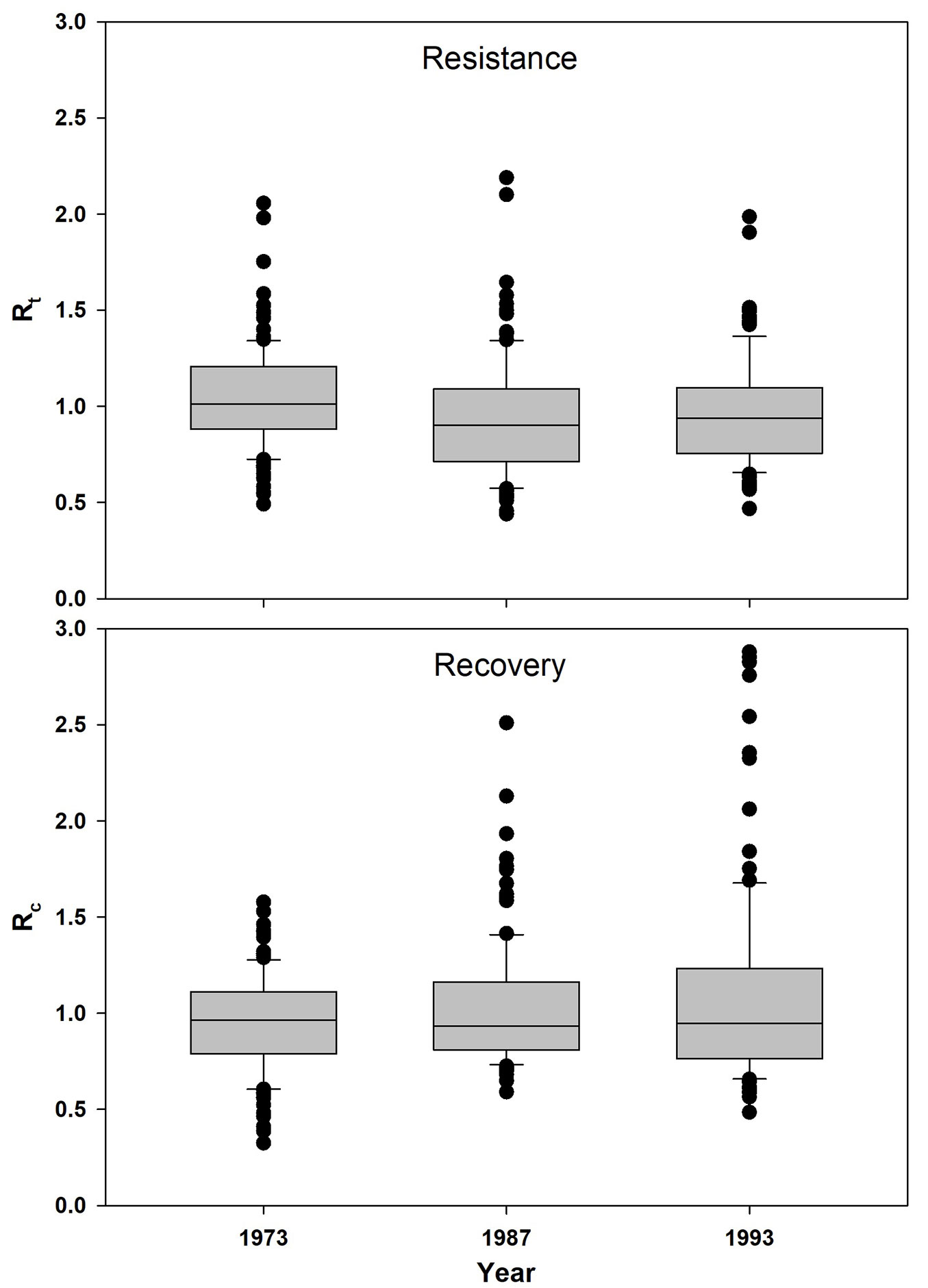
Fig. 4 - Bar plots for Rt > 1, and Rc > 1 between the different considered drought years.
Correlation functions performed at the site level indicated that the species resilience to drought is climatically-driven. In this sense, Rc, and Rc > 1 are significantly and positively related to the precipitation amount of spring-summer and spring periods on one hand and of spring-summer months on the other (Tab. 2). Concerning Rt and Rt > 1, these indexes were significantly and negatively correlated to spring-summer and spring temperatures respectively. No significant correlation emerged for summer period (Tab. 2). Finally, mean age and mean BAI did not represent influencing factors upon any of the examined indexes.
Tab. 2 - Results of the Pearson’s correlation function computed between resistance (Rt) and recovery (Rc) indexes and abiotic and tree biometric variables. (T): temperature; (P): precipitation; (BAI): basal area index; ss, sp and su (subscript) refers to spring-summer, spring and summer period, respectively; (*): p<0.05.
| - | Pss | Tss | Psp | Psu | Tsp | Tsu | BAI | Age |
|---|---|---|---|---|---|---|---|---|
| Rt | 0.23 | -0.42 * | -0.05 | 0.37 | -0.40 | -0.40 | - 0.30 | - 0.24 |
| Rc | 0.44 * | 0.09 | 0.46 * | 0.14 | 0.18 | 0.01 | 0.05 | - 0.00 |
| Rt > 1 | 0.11 | -0.40 | -0.14 | 0.35 | -0.40 * | -0.37 | - 0.35 | - 0.18 |
| Rc > 1 | 0.50 * | 0.06 | 0.40 | 0.36 | 0.15 | -0.01 | 0.12 | 0.11 |
Discussion
This work assessed for the first time P. pinea resilience to drought sensu Lloret et al. ([16]). Despite 21th century extreme dry spells were not considered in this study, analyzing the possible cumulative effect of consecutive dry spells upon stone pine resistance and recovery provided novel information useful to understand the species stem growth dynamics in the current climate unstable scenario ([15]).
Stone pine resilience to drought were clearly determined by climatic conditions. The significant relations among resistance and recovery indexes on one side and temperature and precipitation on the other suggested that local cooler and wetter conditions promoted resilience. On the other hand, extreme climatic events during spring, rather than summer, affected the species resistance and recovery, in agreement with reports showing that stone pine secundary growth is mainly related to environmental conditions at the beginning of the growing season ([8]).
P. pinea is an isohidric species able to tolerate drought through different physiological mechanisms, such as root mortality, stomata control and biomass allocation ([21]). Nevertheless, marked drought conditions affect the species growth diminishing needle length, reducing sap flow, and causing significantly decrease in stem increment ([28], [17]). The positive rainfall effect upon the species recovery could be linked to increasing soil moisture and consequently higher carbohydrates production ([20]). Similarly, the negative relation between temperatures and resistance can reflect the detrimental effect of increasing evapotranspiration and soil water evaporation upon the species growth ([7]). These physiological interpretations are sound with studies showing that under severe environmental constraints, production of wood in P. pinea may be partially or even totally absent ([20]). Finally, it cannot be discarded that other variables not considered in this study, such as stand density, could modulate the species resilience to drought as in other Mediterranean pines species ([26]). Unfortunately, no data were available to retrospectively examine the influence of stand characteristics upon stone pine resilience.
Apparently, no cumulative stress emerged for the species recovery, while resistance slightly decreased along the considered dry spells as showed by Rt > 1 values. In this sense, Rt > 1 in 2003 drought calculated from data belonging to Piraino & Roig-Juñent ([23]) represented 32% of examined individuals (n = 71 - data not shown), thus confirming the tendency towards lower values for this index. On the other hand, it is worth noting that approximately 60% of the analyzed stone pines were not able to reach pre-disturbance growth following drought episodes, which suggests a moderate ability of P. pinea to recover from particularly adverse climatic conditions. As stated in Fang & Zhang ([11]), using proportion of trees can provide additional information which can be hidden by analysis performed considering only mean values of the resilience indexes, minimizing the effect that few outperforming trees can have upon a regional response to drought. Finally, these results encourage future researches focusing on possible higher performance of some of the genotypes examined in this study when facing extreme drought events, an information that would surely contribute to improve the conservation of this natural resource.
Concluding, the results emerged in this study suggested that P. pinea resilience will likely be threatened by more arid conditions expected for the Mediterreanean area, with a consequent decline in the species resistance rather than recovery. This novel information should be considered in management plans, particularly in the scenario of increasing frequency of extreme drought events anticipated for the Mediterranean basin ([13], [2], [15]).
Acknowledgements
Dr. Gianluca Piovesan and Dr. Alfredo Di Filippo (Dendrology Lab, University of Tuscia, Viterbo, Italy) are both greatly acknowledge for supervising the development of earlier versions of P. pinea tree-ring chronologies. Special thanks to Sabrina Raddi and Paolo Cherubini for sharing data through the ITRDB webpage. Thanks are due also to the Authorities of the National Parks that allowed sampling. Two anonymous reviewers are greatly acknowledged for their revisions that improved the manuscript.
References
CrossRef | Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Cátedra de Dasonomía, Facultad de Ciencias Agrarias, Universidad Nacional de Cuyo, Almirante Brown 500, Chacras de Coría - Luján de Cuyo, CPA M5528AHB Mendoza (Argentina)
Corresponding author
Paper Info
Citation
Piraino S (2020). Assessing Pinus pinea L. resilience to three consecutive droughts in central-western Italian Peninsula. iForest 13: 246-250. - doi: 10.3832/ifor3320-013
Academic Editor
Tamir Klein
Paper history
Received: Dec 09, 2019
Accepted: Apr 15, 2020
First online: Jun 19, 2020
Publication Date: Jun 30, 2020
Publication Time: 2.17 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2020
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 41068
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 34262
Abstract Page Views: 3331
PDF Downloads: 2656
Citation/Reference Downloads: 9
XML Downloads: 810
Web Metrics
Days since publication: 2208
Overall contacts: 41068
Avg. contacts per week: 130.20
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2020): 15
Average cites per year: 2.50
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Links between phenology and ecophysiology in a European beech forest
vol. 8, pp. 438-447 (online: 15 December 2014)
Research Articles
Is Pinus pinea growth affected by climate change in western Anatolia?
vol. 18, pp. 93-101 (online: 28 April 2025)
Review Papers
Impacts of climate change on the establishment, distribution, growth and mortality of Swiss stone pine (Pinus cembra L.)
vol. 3, pp. 82-85 (online: 15 July 2010)
Research Articles
Response of juvenile progeny of seven forest tree species and their populations to simulated climate change-related stressors, heat, elevated humidity and drought
vol. 11, pp. 374-388 (online: 15 May 2018)
Research Articles
Predicting the effect of climate change on tree species abundance and distribution at a regional scale
vol. 1, pp. 132-139 (online: 27 August 2008)
Research Articles
Impact of climate change on radial growth of Siberian spruce and Scots pine in North-western Russia
vol. 1, pp. 13-21 (online: 28 February 2008)
Review Papers
Climate change impacts on spatial distribution, tree-ring growth, and water use of stone pine (Pinus pinea L.) forests in the Mediterranean region and silvicultural practices to limit those impacts
vol. 14, pp. 104-112 (online: 01 March 2021)
Research Articles
Impact of climate change on tree-ring growth of Scots pine, common beech and pedunculate oak in northeastern Germany
vol. 9, pp. 1-11 (online: 13 October 2015)
Research Articles
Hearing nature’s heartbeat: towards large-scale real-time forest monitoring network in Italy
vol. 18, pp. 202-211 (online: 09 August 2025)
Research Articles
Potential impacts of regional climate change on site productivity of Larix olgensis plantations in northeast China
vol. 8, pp. 642-651 (online: 02 March 2015)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords







