What if Eurasian jay Garrulus glandarius would larder acorns instead of scatter them?
iForest - Biogeosciences and Forestry, Volume 11, Issue 5, Pages 685-689 (2018)
doi: https://doi.org/10.3832/ifor2793-011
Published: Oct 23, 2018 - Copyright © 2018 SISEF
Research Articles
Abstract
Jays usually store acorns at separate sites, but in some cases a part of jay’s caches consists of more than one acorn. Storing acorns separately (scatter hoarding) or in clusters (larder hoarding) seems to be an important factor for further survival of seeds and seedlings. Scatter hoarding is favorable for jays, but what would happen if jays prepared caches consisting of more than one acorn? We put a following question: what is the importance and impact of acorn concentration per cache for the seedlings’ mortality and their growth characteristics? In 2013 an experimental plot was established. 1400 acorns of Quercus robur L. were sewed in 600 holes in three combinations called growing mode - singly, in twos and in fours. The mortality of seedlings depended on growing mode (Z = 6.24, P < 0.001) and year (Z = -2.42, P = 0.016). In the third year of the experiment the mortality of seedlings growing from acorns sewed in fours reached almost 90%, while in the case of seedlings growing separately the mortality was stable, reaching no more than 23%. Both growing mode (F = 26.49, P < 0.001) and year (F = 52.59, P < 0.001) had a significant impact on seedling growth increment. Seedlings growing in fours had a significantly higher growth increment than these growing separately and in twos. We concluded that seedlings growing separately had a higher survival rate, but lower growth increment than those coming from acorns sewed in clumps.
Keywords
Eurasian Jay, Acorns, Scatter Hoarding, Larder Hoarding, Seedlings Survival, Seed Dispersal
Introduction
Food hoarding animals store food to meet energetic and nutritional demands during food scarcity ([26], [33]). The most spectacular relationship is known between genus Quercus spp. and birds from the family Corvidae ([33], [24]). Among European birds, jay Garrulus glandarius L. is considered to be a typical acorn eater and disperser. Because acorns allow jays to survive winter food scarcity and jays enhance the reproductive success of oaks, by some authors their relation is recognized as a symbiosis ([3]). Eurasian jays may carry acorns and other nuts 4 or 5 km to storage sites ([33]), although it seems that this distance is often much shorter ([5], [9]). Moreover, in autumn jays gather great quantities of oak acorns ranging from 2200 to 5700 ([5], [15]). Also, North American jays collect up to 3000-5000 acorns in a single year ([14]). Consequently, we may observe an effective renewing and chorological expansion of several Quercus species as a result of a close, mutualistic relationship between jays and oaks all over jays’ European range ([8], [27], [28], [25], [6], [17]).
Two main strategies of food hoarding in birds may be recognized: larder hoarding and scatter hoarding ([3], [33], [14]). Larder hoarding is the caching of all food at one site. Scatter hoarding describes the phenomenon of burying food items widely spaced. A mixed strategy with few seeds stored together in many stores scattered in space is also recognized ([33]). Jays usually store each food item (i.e., acorn) at a separate site (scatter hoarding) instead of clumped (larder hoarding), just below the ground surface in individual, widely-spaced cache sites to reduce pilferage and increase the probability of cache recovery ([3], [2]). In many systems this behaviour also contributes significantly to dispersal, germination and establishment of seedlings, when scatter hoarders fail to recover their caches ([33]). Frequently, cached seeds are not recovered and ultimately germinate.
The seed and seedling stages are the most vulnerable for most plant species and processes operating during these stages are thereby especially important for plant regeneration ([10]). The way of seed storage (separately or in clusters) seems to be an important factor for seed and seedling survival, because scattering seems to be clearly more beneficial to the trees ([11]). In the case of jays, pilferage has probably led to storing seeds separately ([2], [19]), but some authors reported that a part of jay’s caches may consist of more than one seed/acorn ([26]). Scatter hoarding strategy is favorable for jays, but what would happen if jays prepared stores of acorns consisting of more than one seed? Is mixed strategy also favorable for acorns and thus for seedling and sapling survival? Answering such questions in jay-oak interactions may have relevant implications for understanding oak renewing processes. In most papers on the effect of seed removal, seed predation and further seedling survival observations were conducted only for a short period of time with no data about the fate of seedlings. There is a lack of data concerning the fate of seedlings several years after germination. This paper reports the results of a field experiment led in three consecutive years after germination simulating the mixed strategy of acorns hoarded in caches consisting of 1, 2 and 4 acorns.
According to model presented by Janzen ([12]), seed and seedling concentration has usually a negative impact on further survival and establishment of new plants. Many reports confirmed that the seedlings’ density may influence the mortality rate ([1], [13], [18], [22]) and affect biometric characteristics of the seedlings ([4]). Thus, we put a following question: what is the impact of acorn concentration per cache on the fate of seedlings (seedling mortality) and their growth characteristics? However, we suspected that growing in high densities, as seedlings usually do in early stages of their establishment, does not mean the same as growing in scattered clumps, which may be profitable for plants ([4]). Taking into consideration potential advantages of growing in clumps (i.e., grafting, mutual shading), we tested the following hypotheses: (1) the mortality rate will depend on seed concentration per cache and it will decline as acorn concentration per cache increases; (2) the height increment of seedlings will increase as acorn/seedling concentration per cache gets higher; and (3) the increment of survived seedlings will increase after the death of other seedlings from the same cache - we call it the “release effect”.
Methods
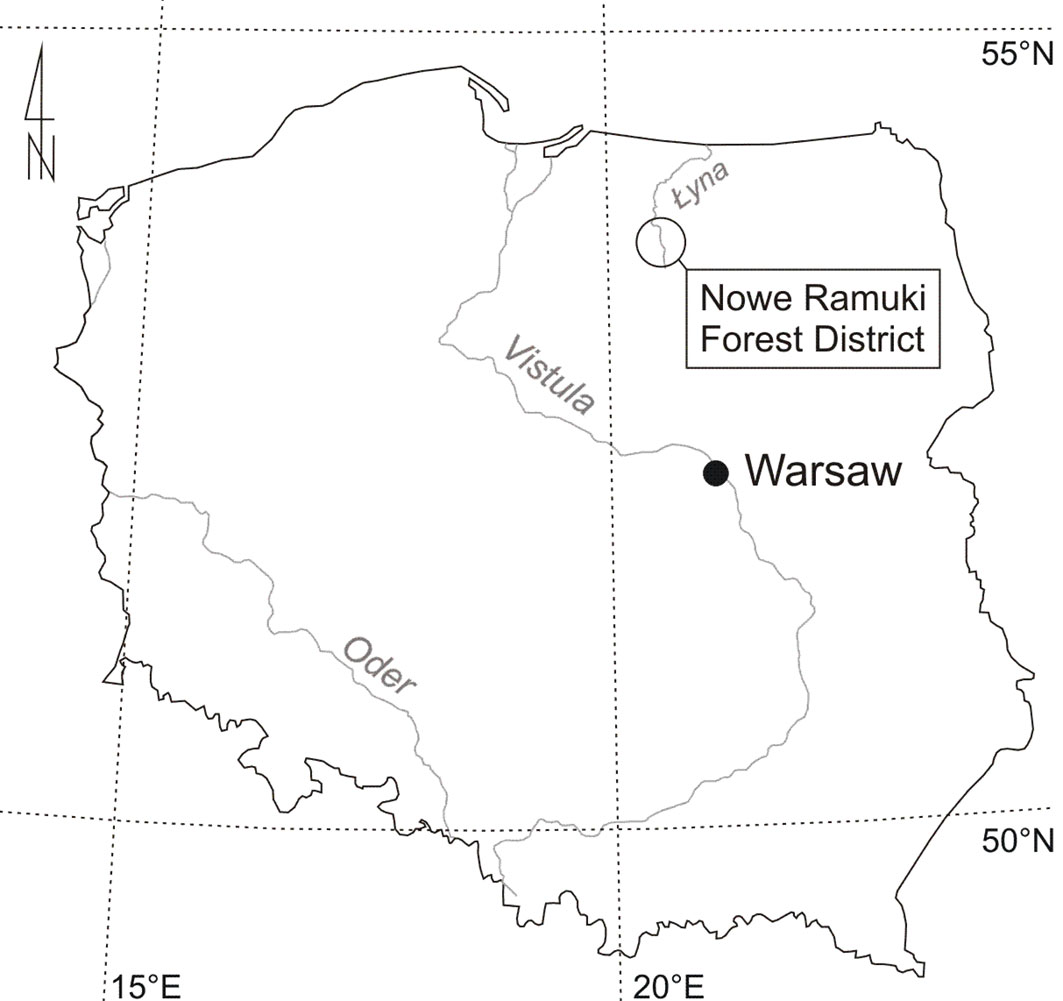
The study was carried out in the Olsztyn Lakeland (Nowe Ramuki Forest District - Fig. 1), among lowland forest complex in north-eastern Poland. It is situated in the Lyna River Valley (289 km long), 200 km north of Warsaw (53.54° to 53.69° N, 20.42° to 20.57° E; altitude up to 200 m a.s.l.) connecting two lakes: Lanskie (10.7 km2) and Pluszne (9.0 km2). In the north, soils are clay-dominated of moraine origin with developed agriculture. In the south, sands and gravel are dominant ([16]) and the landscape is mostly covered by Pinus sylvestris L. and Picea abies Karst. forests with Quercus robur L. as an admixture species. The succession of oaks by jays under the canopy of the Scots pine is evident and widespread in this region. The oak renewal is viable and numerous, mostly present as separately occurring seedlings.
Fig. 1 - Location of the study area in north-eastern Poland - Nowe Ramuki Forest District.
In October 2013 an experimental plot was established. The entire plot was protected from rodents with a steel net with a mesh ca. 0.4 cm attached to the ground. On the plot 600 holes were prepared (15 holes in 40 rows) and 1400 acorns of Quercus robur were sewed in them. Each hole had its own coordinates (hole number and row number) which enabled to precisely establish the fate of seedlings in each hole. We sewed acorns in three types of concentration called growing mode (one, two and four acorns per cache). This means that in 200 holes 200 acorns were sewed (single acorn per cache), in the next 200 holes 400 acorns (two acorns per cache) and 800 acorns in the following 200 holes (four acorns per cache).
In October 2014, 2015 and 2016 seedlings were counted to get information about the survival rate after each year of their growth in three different dispersal modes. After third year of the experiment (October 2016) the total height and height increment from each year were estimated with an accuracy of 1 mm in each seedling. To check if the “release effect” (see hypothesis #3) exists, we compared the growth increments of seedlings growing in twos with seedling growing singly after the death of other seedlings from the same cache (pairs) and with seedlings growing singly from the beginning of the experiment.
We used a generalized linear model with binomial error and pairwise comparison to verify the effects of growing mode, year and interaction between growing mode and year on seedlings mortality. We used two-way ANOVA with Tukey’s post-hoc test to verify the effect of growing mode, year and interaction between them on growth increment of seedlings. The data was transformed with logarithmic functions to obtain a normal or at least symmetric distribution. All analyses were carried out in R ver. 3.3.2 ([30]). The pairwise comparison was performed using the “lsmeans” package ([20]) and the data visualisation was performed by the “ggplot2” package ([35]).
Results
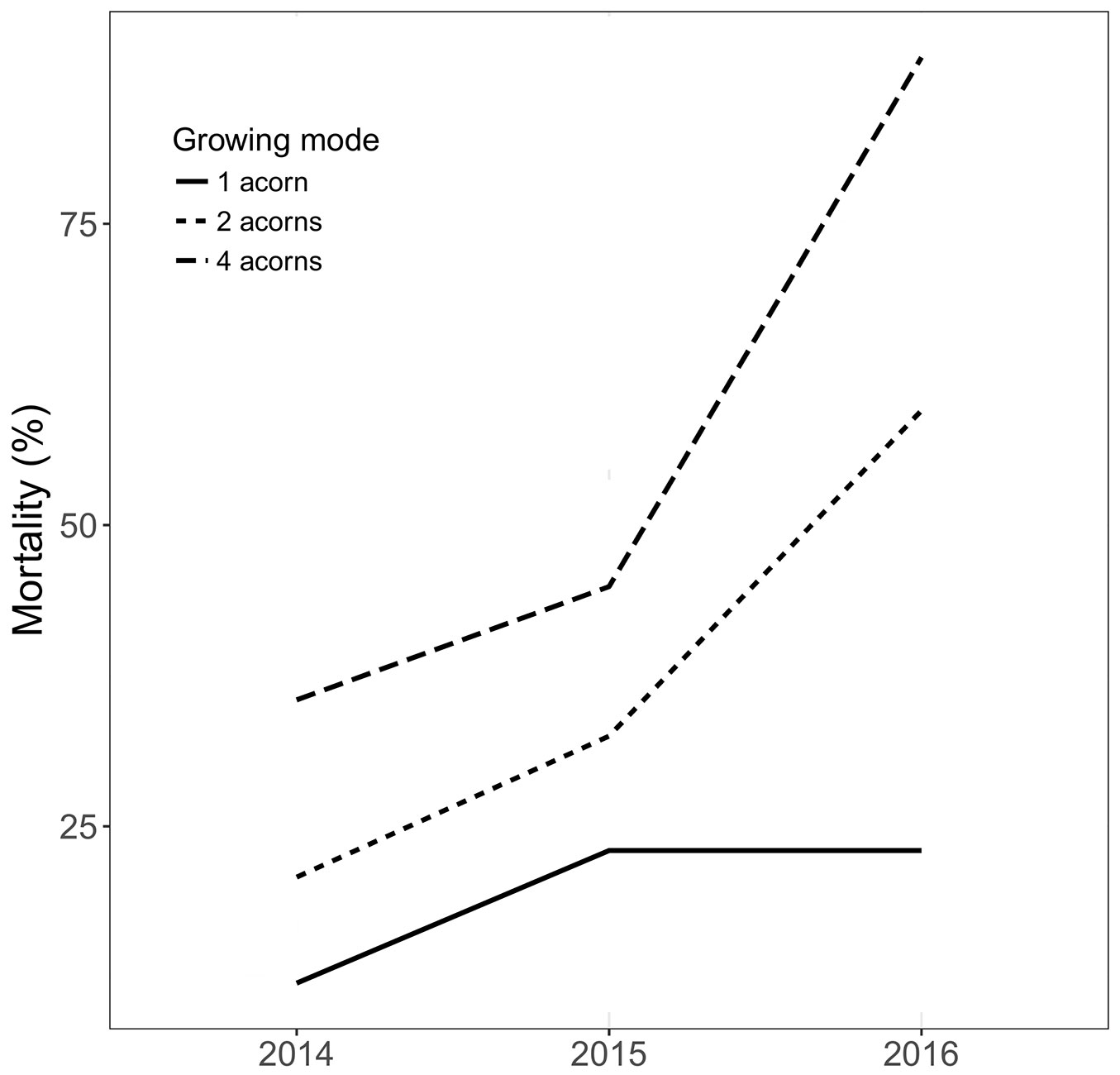
The mortality of seedlings depended significantly on the growing mode (Z = 6.24, P < 0.001) and year (Z = -2.42, P = 0.016). During all three years the greatest cumulative mortality was detected in the case of seedlings growing from acorns sewed in fours per cache (Fig. 2). The number of died down seedlings gradually increased after the first and second years (2014 and 2015), but the greatest mortality was observed after the third year (2016), when the mortality of seedlings (especially from acorns sewed in twos and in fours) suddenly increased. In the third year of the experiment, the mortality in the case of seedlings growing from acorns sewed in fours reached almost 90%, while the mortality of seedlings growing separately was stable, reaching no more than 23%.
Fig. 2 - Mortality of seedlings of Quercus robur depending on dispersal mode (1 acorn, 2 acorns, 4 acorns per cache) during three years of the experiment.
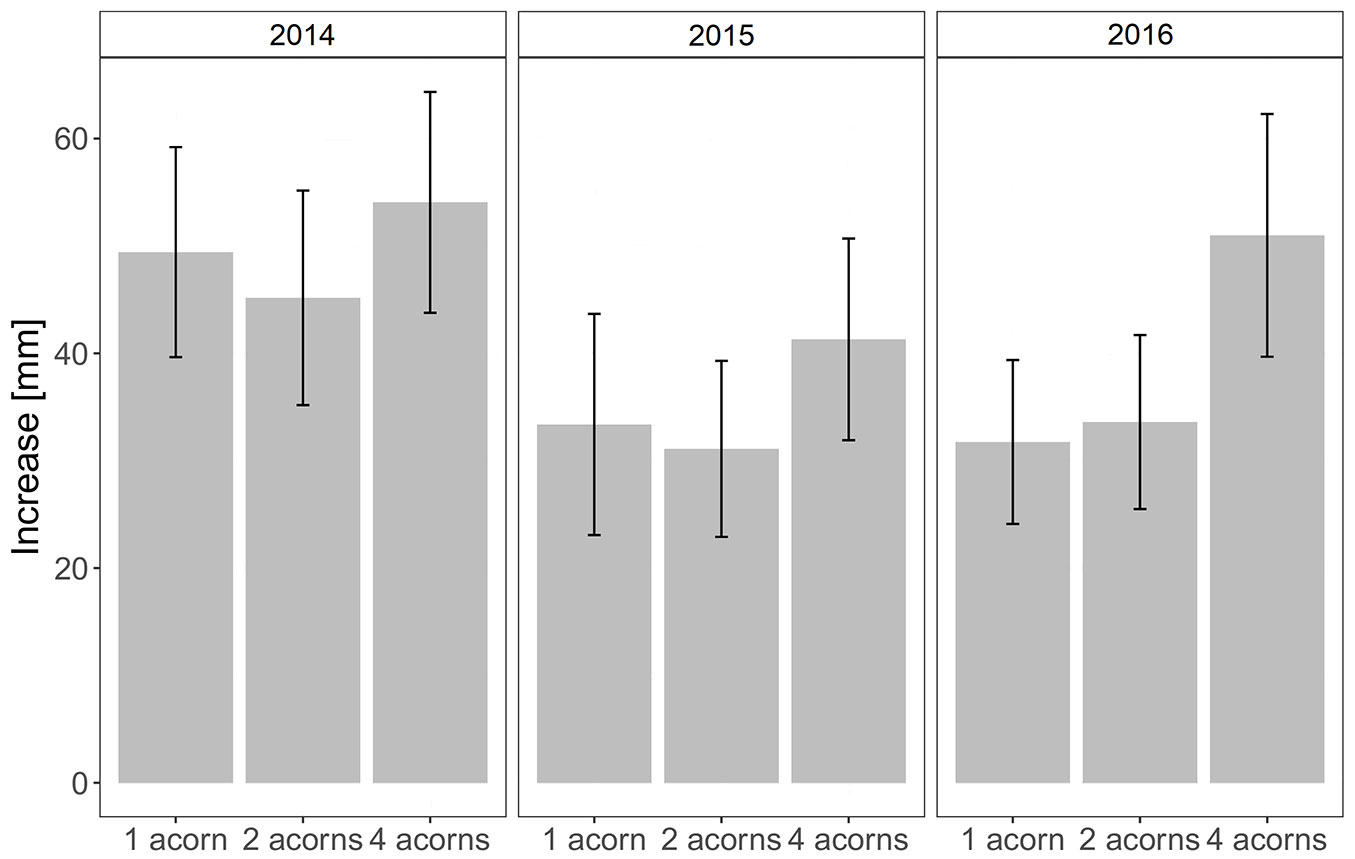
Both growing mode and year had a significant impact on seedling growth increment (Tab. 1). In comparison to seedlings growing separately and in twos, significantly higher increments were recorded in the case of seedlings coming from the acorns that were sewed in fours (Fig. 3). No difference in increment were found between seedlings growing separately and in twos. Seedlings grew best in the first year after germination. Growth increments did not differ between the next two years (Fig. 3).
Tab. 1 - Results of ANOVA on the differences in height increment of seedlings depending on growing mode (1, 2 and 4 acorns per cache) and year (2014, 2015 and 2016).
| Effect | Height increment | ||
|---|---|---|---|
| df | F | P | |
| Growing mode | 2 | 26.49 | < 0.001 |
| Year | 2 | 52.59 | < 0.001 |
| Interaction | 4 | 3.15 | 0.014 |
Fig. 3 - Height increments of seedlings of Quercus robur depending on dispersal mode (1 acorn, 2 acorns, 4 acorns per cache) and year (2014, 2015, 2016).
There were no significant differences in height increments between seedlings growing separately, in twos and growing separately after dying down of one of the pair (F = 1.11, P = 0.333). However, mean height of seedlings growing separately and those which stayed single after dying down of one of the pair were higher (114.5 vs. 113.6 mm, respectively) than mean height of seedlings still growing together (107.9 mm).
Discussion
Oaks benefit from food hoarding animals that bury their acorns in suitable sites where they may establish a new plant ([33]). The survival rate of seeds stored in jay caches is higher, because of reduced predation from granivory species and pest pressure which prevents seedlings from drying ([10], [29], [14]). Jays deposit acorns in shallow subsurface caches in soil or between the soil and the litter layer and cache depth is constrained by the length of birds’ bill ([33]). In particular, seed burial by animals strongly affects seedling establishment in many plant species. Seedlings that emerge from acorns buried in the soil or deposited close to bare soil (under litter layer) have a better start and a higher chance to survive than acorns that had fallen straight from the tree and were not buried ([31]). We tried to answer the question what was the effect of scatter or larder hoarding on seedling survival and seedling growth.
The most obvious benefit of scatter hoarding over larder hoarding is not necessarily solely related to robbery by food competitors. Seedling mortality in caches can be attributed to a variety of factors ([11], [33]). Seedlings’ death can be caused by storage at sites inappropriate for establishment; seedlings are damaged by insects, pathogens, seed predators, seedling herbivory, animals digging up the germinants and by intraspecific competition for water and other resources ([11], [33]). Moreover, seedlings’ density affect seedling survival and recruitment ([1], [13], [18], [22]). The neighborhood of conspecifics may increase the competition for resources (nutrient and water). In this case, especially in periods of resource demands, the mortality may be higher. In research carried out by McBride et al. ([23]) the mortality during drought was much higher in 10 acorn clumps (55.2%) than in the case of single acorns (23.9%). Our findings also confirm that saplings may have higher mortality, when growing in groups than separately (90% vs. 23%). We also found that the mortality increased as acorn concentration per cache increased, thus hypothesis #1 was not confirmed (Fig. 2).
We conclude that the seed/seedling concentration per cache increases the mortality rate in comparison to plants growing separately. Bossema ([3]) in his research found that planting in clumps or separately did not influence the viability of acorns. Therefore the chance of acorns rotting was not influenced by the presence of other acorns. The results cited above show that in some cases growing in clumps did not affect the survival rate of seedlings. Saplings’ roots may contact (by grafting) and improve the nutrient intake, when growing in groups. Bullock ([4]) suggested that seedlings in aggregations may profit from i.e. mutual shading or grafting. Grafting was commonly observed among individuals/ seedlings growing in clumps ([4]). It may be that in some cases seedlings and future saplings growing separately (no grafting observed) may have higher mortality than these growing in groups ([4]). On the other hand, we have to remember that the death of the epicotyl does not mean the death of a whole seedling. In many cases in our experiment, seedlings revealed an apparent death by epicotyl caesura. Such effect may reappear in the following years, preventing the young plant’s efficient growth; nonetheless, the roots are still alive and may produce shoots again.
Many studies revealed that the mortality rate among seeds and seedlings is usually density dependent ([7], [11], [33]). Some authors reported that the consequences of seed aggregation in caches may result in higher seedling survival and thus by distinct multi-trunked growth forms of adult trees ([4], [21]). During seedling establishment, the number of stems per clump has been found to be negatively related to clump age for some species ([32], [34]) and in consecutive years the clumps consisting of more than 1-2 stems became rare ([33]). The intraspecific competition for water between seedlings from a cache seems to be more distinctive in species that occupy arid sites. Vander Wall ([33]) reported that mature piñon pines are rarely found in multiple-stemmed clumps like whitebark pine and limber pine, which occupy relatively more mesic sites. Multi-stemmed clumps in such conditions are usually reduced to a single seedling within a few growing years. As a consequence of high mortality in seedlings that are normally clump-dispersed, usually no more than one adult plant from a single clump survives ([11], [33]).
We found it interesting that the biggest height increment of seedlings was observed in the case of acorns sewed in fours (i.e., hypothesis #2 was confirmed). So, the effect of seedling clumps on quantity (growth increment) and quality (mortality) characteristics seems to be reciprocal. Bullock ([4]) also reported that seedlings growing in clumps had a higher biomass than those growing separately. We did not find support for the third hypothesis, where we stated that the increment of survived seedling increases after the death of other seedlings from the same cache. Seedlings from numerous caches had better growth increments, so there are no grounds for the “release effect” to occur.
In contrast to clump-dispersal (larder hoarding), the distinctive feature of scatter-dispersal for plant demography is that seeds normally germinate from several centimeters to many meters away from other seeds ([11]). Not all studies reported that growing separately is better for sapling survival, but in some cases scatter-hoarding strategy of dispersal seems to be more effective and convenient from the ecological and parental tree point of view ([12]). Scattering of single acorns into separate caches seems to be the most effective anti-pathogen and anti-granivory strategy we can imagine. Scattering of hoarded food is also important for jays, since patchy distribution of caches containing single acorns are less susceptible to granivory. This evidence suggests that both participants (plant and bird) benefit from this strategy which is a kind of trade-off between costs and benefits ([33]). Plants gain a higher seed and seedling survival and a more efficient dispersal effect (acorns cached separately), while the growth rate of seedlings is lower than in clumps. Scatter hoarding birds benefit lower hazard of cache loss from granivory, but must invest more energy in scattering the caches and retrieving them. Scattering seems to be clearly more beneficial for seedlings due to decreased mortality. Answering the question from the title, we can confirm that in the case of Quercus robur it is much more favourable for seedlings’ survival that jays scatter their acorns rather than clump them.
Acknowledgements
We are grateful to the Forest Managers Janusz Jeznach and Grzegorz Wanat and to the staff of Nowe Ramuki Forest District, for great help, supporting and enabling us to conduct the research. We are also grateful to Fiona Milne-Rostkowska for the final language correction. The study was supported by grant number 240110 financed by the Polish Ministry of Science and Higher Education.
References
Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Department of Plant Ecology and Environmental Protection, Adam Mickiewicz University, Umultowska 89, 61-614 Poznan (Poland)
Department of Forest Ecology, Forest Research Institute, Braci Lesnej 3, 05-090 Sekocin Stary (Poland)
Department of Forest Botany, Poznan University of Life Sciences, Wojska Polskiego 71D, PoznaÅÂ 60-625 (Poland)
Institute of Zoology, Poznan University of Life Sciences, Wojska Polskiego 71C, Poznan 60-625 (Poland)
Corresponding author
Paper Info
Citation
Kurek P, Dobrowolska D, Wiatrowska B, Dylewski L (2018). What if Eurasian jay Garrulus glandarius would larder acorns instead of scatter them?. iForest 11: 685-689. - doi: 10.3832/ifor2793-011
Academic Editor
Massimo Faccoli
Paper history
Received: Mar 20, 2018
Accepted: Aug 02, 2018
First online: Oct 23, 2018
Publication Date: Oct 31, 2018
Publication Time: 2.73 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2018
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 50700
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 43015
Abstract Page Views: 3450
PDF Downloads: 3261
Citation/Reference Downloads: 6
XML Downloads: 968
Web Metrics
Days since publication: 2807
Overall contacts: 50700
Avg. contacts per week: 126.43
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2018): 4
Average cites per year: 0.50
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
The effect of seed size on seed fate in a subtropical forest, southwest of China
vol. 9, pp. 652-657 (online: 04 April 2016)
Research Articles
Seed trait and rodent species determine seed dispersal and predation: evidences from semi-natural enclosures
vol. 8, pp. 207-213 (online: 28 August 2014)
Research Articles
Dispersal and hoarding of sympatric forest seeds by rodents in a temperate forest from northern China
vol. 7, pp. 70-74 (online: 18 November 2013)
Short Communications
Evidence of Alectoris chukar (Aves, Galliformes) as seed dispersal and germinating agent for Pistacia khinjuk in Balochistan, Pakistan
vol. 14, pp. 378-382 (online: 22 August 2021)
Research Articles
The impact of seed predation and browsing on natural sessile oak regeneration under different light conditions in an over-aged coppice stand
vol. 9, pp. 569-576 (online: 04 April 2016)
Research Articles
Use of fallen dead trees by Japanese squirrels within cedar plantations in northeastern Japan
vol. 16, pp. 262-267 (online: 19 October 2023)
Research Articles
Inter- and intra-annual patterns of seed rain in the black spruce stands of Quebec, Canada
vol. 10, pp. 189-195 (online: 13 December 2016)
Short Communications
Differential adaptations in nursery seedlings from diverse Chilean provenances of Peumus boldus Mol.
vol. 9, pp. 409-413 (online: 29 January 2016)
Short Communications
Towards an optimal sampling effort for paternity analysis in forest trees: what do the raw numbers tell us?
vol. 5, pp. 18-25 (online: 27 February 2012)
Research Articles
Seed germination traits of Pinus heldreichii in two Greek populations and implications for conservation
vol. 15, pp. 331-338 (online: 24 August 2022)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







