
Chitosan oligosaccharide addition affects current-year shoot of post-transplant Buddhist pine (Podocarpus macrophyllus) seedlings under contrasting photoperiods
iForest - Biogeosciences and Forestry, Volume 10, Issue 4, Pages 715-721 (2017)
doi: https://doi.org/10.3832/ifor2302-010
Published: Jul 27, 2017 - Copyright © 2017 SISEF
Research Articles
Abstract
Chitosan oligosaccharides (COS) have been used as modifiers to promote growth and mineral nutrient utilization in crop plants, but its over-year effect on current-year shoot (CYS) of juvenile trees is still unclear. In this study, Buddhist pine (Podocarpus macrophyllus) seedlings were cultured under natural and extended photoperiods with or without COS addition for one year. In the following spring, parameters of leaf length, biomass accumulation, and N content in CYS were found to be increased by COS addition under the extended photoperiod. P concentration of COS-treated seedlings was lower under longer photoperiod, but both N and P concentrations were negatively correlated with leaf length and biomass accumulation, suggesting the utilization of N and P for growth demand of CYS. The sole addition of COS mainly resulted in whole-plant P accumulation. However, when combined with the extended photoperiod, COS addition showed over-year effect on biomass accumulation and N content in CYS of transplanted Buddhist pine seedlings. Further studies are needed to confirm these results on other tree species.
Keywords
Photoperiodism, Urban Afforestation, Yew Plum Pine, Marine Oligosaccharide, Fine Root
Introduction
During the past decade, there has been a worldwide increasing interest to explore new alternative enhancers of plant growth with low environmental impacts and lower adverse side effects on human health compared to current synthetic agrochemicals. In this context, chitosan is attracting more and more attention as a biodegradable natural polysaccharide derived from chitin through deacetylation. Chitin is an abundant and water insoluble biopolymer found mainly in the hard outer skeleton of marine animals ([16]). Chitosan is comprised of 2-acetamido-2-deoxy-β-D-glucose (N-acetyl-D-glucosamine) and 2-amino-2-deoxy-β-D-glucan (D-glucosamine) attached via β-(1, 4) linkages. Chitosans include a wide group of molecules with a specific reactive amine group able to bind negatively charged molecules, e.g., proteins ([6]). Chitosan oligosaccharides (collectively oligochitosan or COS) are the enzymatic-hydrolyzed products of chitosan and can be used as plant growth regulators through seed coating (or seed dressing) and foliar spray ([6], [30], [16] and references therein). Most of the current knowledge on COS as plant modifiers was established according to studies on agricultural crops, while on woody species COS were mainly tested for biotic resistance ([15], [18]). So far, few studies have focused on the effect of COS addition on tree seedling growth. González et al. ([11]) reported a promoting effect of seaweed-derived oligo-carrageenans on growth, cell division, basal metabolism, and carbon (C) and nitrogen (N) assimilations in 3-year-old Eucalyptus globulus trees. However, different sources of marine oligosaccharides may differently affect plant growth. As COS can modulate mineral uptake in crops ([6]), a favorable effect on growth and nutrition of tree species is expected.
Due to the low growing rate, culturing highly-valued tree seedlings in the nursery usually requires a long time before transplant to meet the quality standard. Extending the photoperiod by supplementing artificial illumination at night is an effective approach to cope with this issue ([10], [29]) and promotes a faster growth of slow-growing tree seedlings ([2], [24]). Recent studies mainly focused on the effect of extended photoperiod treatments in the nursery on growth and inherent nutrient status of slow-growing ornamental tree species ([31], [32], [33]), though their over-year post-transplant performance has rarely been investigated.
Current-year shoot (CYS), including current-year buds, leaves, and twigs, is the most vulnerable tree organ ([12], [3]). It is also the main target (sink) of nutrient retranslocation from storing tissues in the spring ([20], [19]). Nitrogen (N) and phosphorus (P) are two important nutrients whose previous-year uptake and overwinter storage may affect CYS growth in the next spring ([13], [9], [21], [22]). Several studies have demonstrated the positive effect of previous nursery fertilization (e.g., exponential and late-season fertilization) on CYS growth in transplanted trees, which enhanced mineral storage and promoted nutrient retranslocation ([22], [31], [23]). Recently, Wei et al. ([31]) and Zhu et al. ([32], [33]) found that an extended photoperiod can promote growth and biomass accumulation in slow-growth tree seedlings, but resulted in the dilution of inherent N and P storage. According to Villar-Salvador et al. ([28]), an extended photoperiod may also impact CYS growth of seedlings, due to the reduction of non-structural carbohydrates storage as a result of the prolonged photosynthesis. The above studies suggested the risk of unsatisfying CYS growth in transplanted seedlings reared under a longer photoperiod. To our knowledge, none of these hypotheses have been tested so far.
Podocarpus is a large plant genus comprising 97 species of evergreens and shrubs mostly found in the Southern Hemisphere. Buddhist pine (Podocarpus macrophyllus) is widely cultivated as a garden or landscape tree in sub-tropical areas of the northern hemisphere ([31]). In addition to the aesthetical character, Buddhist pine is also a valuable species for its pharmacological uses, though its cultivation is hampered by the low growing rate of seedlings. Recently, transplanted stocks of Buddhist pine seedlings cultured under extended photoperiod have been established as pre-trials in the Zijinshan Park (34° 45′ N, 113° 40′ E), Zhengzhou City and in the Xiyuan Park (34° 39′ N, 112° 24′ E), Luoyang City, Henan Province (China), with unsatisfying results. Therefore, a better understanding of the effect of photoperiod length on seedling quality is needed to enhance CYS growth in transplanted stocks.
The aim of this study was to compare growth and utilization of N and P in CYS of transplanted Buddhist pine seedlings cultured under contrasting photoperiods. COS was employed to test its potential interaction with photoperiod on influencing CYS growth. Our starting hypothesis was that growth and nutrient utilization in CYS were only promoted in seedlings treated with COS and reared under a longer photoperiod.
Materials and methods
Seedling materials
Buddhist pine seeds were collected from a seed orchard at Hangzhou, Zhejiang, China (30° 10′ N, 120° 20′ E) by the East China Horticulture Garden (Shuyang, Jiangsu, China). On early April 2014, seeds were transported to the Laboratory of Urban Forests and Wetlands, Northeast Institute of Geography and Agricultural Ecology (Changchun, China), Chinese Academy of Sciences, and sterilized in potassium permanganate (0.5 %, w/w) for 5 min. Seeds were then soaked for 12 h, half of them in distilled water and the other half were coated by soaking with a COS solution (Qishanbao®, GlycoBio Co., Ltd., Dalian, Liaoning, China) at a concentration of 2 mg kg-1 (w/w). Seeds were sowed in trays filled with sterilized sands for germination. On mid June 2014, germinated seeds were transferred to plug trays filled with mixed peat (Zhuangmiao® Peat Tech. Ltd., Changchun, Jilin, China) and perlite (3:1, v/v). Each trail included 32 (4 × 8) plugs of 7 × 13 cm (diameter × height). Seedlings were cultured with a sub-irrigation system implemented for tree seedling culture ([5]). Tanked water was maintained at the level of 4 cm in height by manual irrigation every day. The total number of seedlings cultured in this experiment was 1024.
Photoperiod and marine oligosaccharide addition treatments
Photoperiod and COS treatments were applied to seedlings since one week after germinated seeds were planted. Plug trays were placed in two separated rooms (16 trays with 512 seedlings per room). In the first room, photoperiod was set at 18h per day (from 6:00 a.m. to 24:00 p.m.) using 200-W plant grow lamps (Oudi® Illumination Ltd., Huzhou, Zhejiang, China) which provided a photosynthetic photon flux density (PPFD) of 400 µmol m-2 s-1 at the apical tip of seedling shoots. In the other room, a “natural” photoperiod of 11.5 h per day (from 6:00 a.m. to 17:30 p.m.) with the same PPFD was provided to seedlings using the same lamps described above. A PPFD of 400 µmol m-2 s-1 was chosen because it was close to the light saturation point (LSP) of Podocarpus spp. ([8], [25]). The natural photoperiod conditions were set based on data collected during pre-investigations carried out in the open air at Luoyang City (China), with available PPFD over light compensation point (LCP) of Buddhist pine (~20 µmol m-2 s-1).
All seedlings were fed with fertilizers at the beginning of the treatments, according to the typical exponential fertilization model employed for Buddhist pine seedlings by Wei et al. ([31] - eqn. 1):
where r is the relative addition rate required to increase NS to a final N content (NT + NS) over the different fertilizer applications (t=16). NS and NT are the initial N content of seedlings and the desired amount to be added, which were set to 1.39 mg N seedling-1 and 80 mg N seedling-1, respectively. The quantity of fertilizer for each application (Nt) was computed as follows ([31] - eqn. 2):
where Nt-1 is the cumulative amount of N added up to the current application including all previous applications. The supplied fertilizer was Peters® Professional (N-P2O5-K2O, 20-20-20, microelements added - Everris, CA Geldermalsen, Netherlands). At each fertilizer application, seedlings from COS-coated seeds were treated with exogenous COS through foliar spray, while the other seedlings were sprayed with the same volume of distilled water. All the described treatments were replicated in eight blocks. Plug trays in each room were shuffled every week to eliminate the possible edge effect. Average air temperature and relative humidity were kept at 24.9 °C and 57%, respectively, throughout the whole experiment.
In late October 2014, after the last application of fertilizers, the photoperiod was set at 11.5h (natural) for all seedlings in all rooms (including those previously subjected to extended photoperiod), the temperature was set at 10/16 °C (night/day) and relative humidity at 36-43%. These conditions are those commonly adopted in the Henan Province, in order to induce the formation of apical buds in Buddhist pine seedlings.
Transplant experiment
In late January 2015, seedlings were transplanted into plastic pots (top diameter × bottom diameter × height, 28 × 20 × 17 cm) filled with soil collected from 60-year-old northeast black pine (Pinus tabuliformis var. mukdensis) plantations in the northern part of Nanhu Park, Changchun, Jilin, China (43° 51′ N, 125° 17′ E). The soil was collected at the top 10 cm depth of the forest floor and determined for its properties, showing a texture of loamy with sand content at ~60%, bulk density of 1.42 g cm-3, pH of 6.22, hydrolyzable N of 127.94 mg kg-1, available P of 18.40 mg kg-1, and organic matter of 3.5%.
Four seedlings were transplanted in each pot, and a total of 128 plastic pots were used. Transplanted seedlings were watered according to real-time precipitations with temperatures controlled to mimic those in Zhengzhou, Henan, Central China (source data: Wunderground® - ⇒ http://www.wunderground.com).
Seedling sampling and measurements
Bud burst in treated seedlings began at DOY 50 (days of year, since Jan 1st). At DOY 105 (mid-April) current-year leaves had fully expanded in all seedlings, which were then all sampled the same day. Although some leaves turned to be dark green, dark yellow, or even brown, no apparent mortality was found. One of the four seedlings was randomly sampled in each plastic pot and four seedling samples were bulked as a single group in each combined treatment (photoperiod × COS) unit.
Seedlings were carefully hand-lifted from potted soils to maintain the whole root system intact. Excavated seedlings were carefully washed to clean soil residuals using tap water, then rinsed with distilled water and dried using tissue paper. Seedling height, root-collar diameter (RCD), and number of new roots (at least 2mm in length, white and juvenile tips) were recorded. Thereafter, seedlings were divided into different organs: current-year shoots (CYS), past-year shoots (past-year leaves, twigs, and the stem), and roots. Top leaves of seedlings were greater than those at the stem base; therefore, leaf length was measured on three randomly-chosen leaves from the middle part of stem in both current-year and past-year shoots, and then averaged. The biomass of separated seedling organs was measured after oven-drying at 70 oC for 72 h. Dried samples were ground to powder and determined for total N and P concentrations in the Laboratory of Forest Silviculture, Beihua University, Jilin, Jilin, China. A sample of 0.2 g dried powder was digested in 5 ml of H2O2-H2SO4. The digestion solution was diluted to 50 ml. Seedling total N and P concentrations were determined using the Kjeldahl method and ICP-OES (Vista-MPX, Varian®, USA), respectively.
Statistical analysis
Data were analyzed as a split-plot design with eight replicates to determine the effect of photoperiod (extended vs. natural), with or without COS addition, on morphology, biomass accumulation, and N and P contents in CYS and other organs. Photoperiod was taken as the main factor, while COS treatment was nested within the main factor. All the studied variables met the assumptions of homoscedasticity and normality, thus no data transformation was applied. Analysis of variation (ANOVA) was employed to analyze the effects using the “Proc GLM” program in SAS® 9.2 (SAS Institute, Cary, NC, USA). All interactions were considered for all parameters in the analysis (Tab. 1). Differences were considered statistically significant when p<0.05. Relationships between nutrient concentrations and biomass or leaf length were analyzed by both Pearson’s and Spearman’s correlation analysis using the “Proc CORR”. Where significance was detected, data were fitted to a suitable curve using SigmaPlot® v. 12.0 (Systat Software, Inc., San José, CA, USA), and the relative coefficient of determination (R2) calculated.
Tab. 1 - F values from ANOVA testing the effect of COS addition (O), photoperiod (Ph), and their interaction (O × Ph) on seedling parameters of growth, biomass accumulation, and N and P concentrations and contents in post-transplant Buddhist pine (Podocarpus macrophyllus) seedlings. (RCD): root collar diameter; (NNR): number of new roots; (LCYL): length of current-year leaves; (LPYL): length of past-year leaves; (*): P<0.05; (**): P<0.01; (***): P<0.001.
| Seedling parameters | Source of variation | Height | RCD† | NNR | LCYL | LPYL |
|---|---|---|---|---|---|---|
| Growth | O | 1.52 | 0.92 | 2.72 | 15.98*** | 0.15 |
| Ph | 9.24** | 11.40** | 8.29** | 44.73*** | 39.00*** | |
| O×Ph | 1.18 | 4.92* | 0.03 | 32.36*** | 3.68 | |
| - | - | New shoot | Old shoot | Shoot | Root | |
| N concentration | O | 0.66 | 1.13 | 25.20*** | 8.22** | |
| Ph | 0.74 | 1.93 | 25.42*** | 6.88* | ||
| O×Ph | 3.05 | 0.87 | 5.79* | 0.08 | ||
| P concentration | O | 0.01 | 4.54* | 10.52** | 0.23 | |
| Ph | 11.41** | 0.81 | 10.68** | 6.84* | ||
| O×Ph | 4.39* | 2.26 | 4.73* | 0.47 | ||
| Biomass accumulation | O | 7.51*‡ | 1.79 | 5.96* | 5.08* | |
| Ph | 24.33*** | 31.28*** | 60.06*** | 36.72*** | ||
| O×Ph | 12.57** | 4.92* | 8.78** | 5.05* | ||
| N content | O | 19.44*** | 51.98*** | 74.09*** | 22.24*** | |
| Ph | 2.54 | 5.16* | 7.86** | 8.06** | ||
| O×Ph | 4.70* | 1.93 | 5.06* | 5.19* | ||
| P content | O | 9.72** | 57.09*** | 36.74**** | 22.79*** | |
| Ph | 1.54 | 1.82 | 0.14 | 7.00* | ||
| O×Ph | 0.2 | 0.06 | 0.06 | 1.11 | ||
Results
Seedling morphology and biomass accumulation
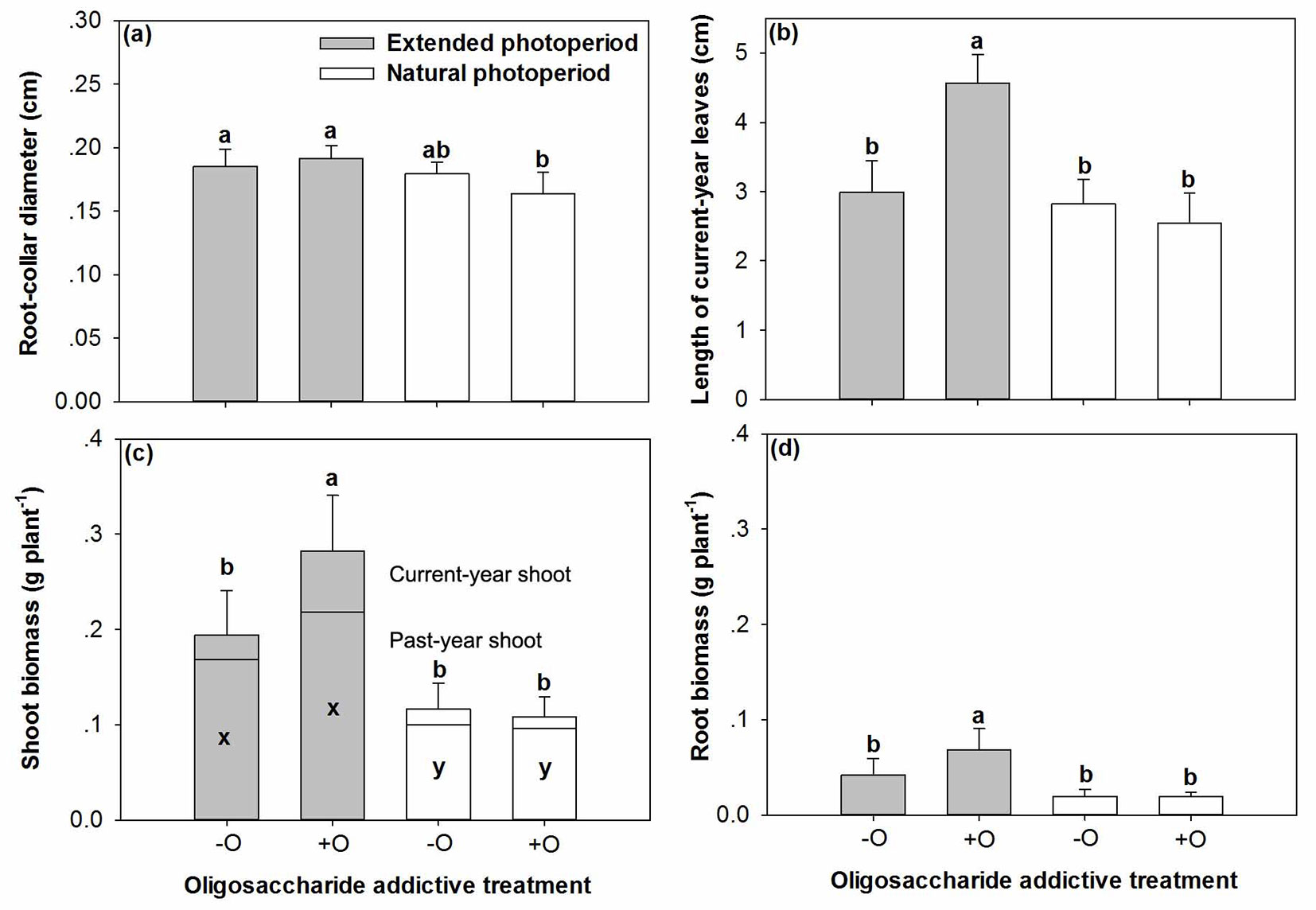
A significant interactive effect of before-transplant photoperiod and COS addition was detected on RCD (P=0.0349) and leaf length of CYS (P<0.0001). Compared to seedlings grown under the extended photoperiod, RCD declined by about 13% in those receiving COS under the natural photoperiod (Fig. 1a). Mean leaf length in CYS was greatest in seedlings treated with COS under the extended photoperiod, exceeding by 50-80% that observed in other treatments (Fig. 1b). Photoperiod had a significant effect on seedling height (P=0.0051), number of new roots (P=0.0076), and length of past-year leaves (P<0.0001), which were greater in the extended photoperiod by 17%, 35%, and 40% than those in the natural one, respectively (Tab. 2).
Fig. 1 - (a) Root-collar diameter (RCD), (b) length of current-year leaves, and biomass accumulation in shoots (c) and roots (d) of post-transplanted Buddhist pine (Podocarpus macrophyllus) seedlings in response to before-transplant photoperiod treatment (extended vs. natural) with (+O) or without (-O) COS addition. Different letters indicate significant differences (p<0.05, n=8). In (c), a and b for current-year shoot, while x and y for past-year shoot.
Tab. 2 - Average seedling height, number of new roots, length of past-year needles, N and P concentrations in root, past-year shoot N content, and root P content (± standard error) in post-transplant Buddhist pine (Podocarpus macrophyllus) seedlings cultured under extended and natural photoperiods (n=16). Different letters in the same row indicate significant differences (p<0.05).
| Parameters | Photoperiod | |
|---|---|---|
| Extended | Natural | |
| Seedling height (cm) | 7.08 ± 1.02 a | 6.05 ± 0.62 b |
| Number of new roots | 21.31 ± 4.87 a | 15.75 ± 4.76 b |
| Length of past-year leaves (cm) | 3.93 ± 0.54 a | 2.81 ± 0.34 b |
| N concentration in root (mg g-1) | 21.02 ± 3.38 a | 16.88 ± 5.00 b |
| P concentration in root (mg g-1) | 0.83 ± 0.28 b | 2.06 ± 1.58 a |
| N content in past-year shoot (mg) | 3.94 ± 1.55 a | 3.21 ± 1.03 b |
| P content in root (µg) | 33.66 ± 16.98 b | 56.94 ± 36.54 a |
Biomass was greatest in seedlings with COS addition under the extended photoperiod for both CYS (P=0.0014) and root (P=0.0327 - Fig. 1c, Fig. 1d). Contrastingly, mean past-year shoot biomass was not significantly different between treatments, regardless photoperiod length and COS addition. In general, seedlings subjected to the longer photoperiod treatment had greater biomass in past-year shoots.
N and P concentrations
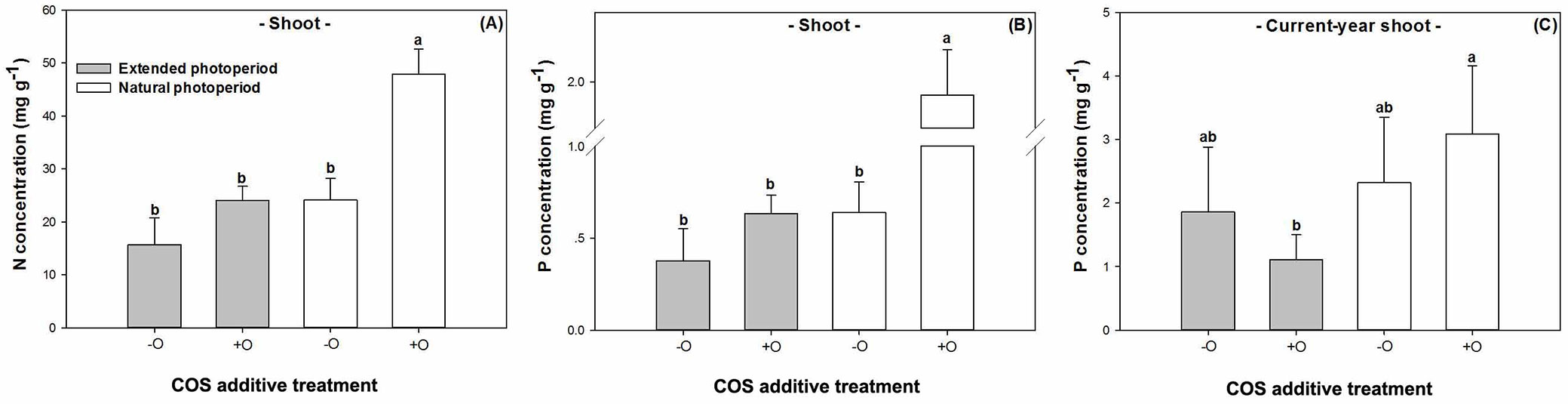
Treatments of photoperiod and COS addition had a significantly interactive effect on N concentration in shoot (P=0.0230) and P concentration in both shoot (P=0.0382) and CYS (P=0.0453 - Tab. 1). Both N and P concentrations were highest in seedlings receiving COS addition under the natural photoperiod (Fig. 2A, Fig. 2B), and P concentration in CYS for this treatment was only higher than that in seedlings receiving COS addition under the extended photoperiod (Fig. 2C). Moreover, photoperiod had a significant effect on both N and P concentrations in roots, which had higher N by 25% (P=0.0139) but lower P by 60% (P=0.0142) in the longer photoperiod (Tab. 2). In contrast, the addition of COS decreased N concentration in root by 27% (P=0.0078), but this treatment increased P concentration in past-year shoot by 25% (P=0.0421).
Fig. 2 - (A) Nitrogen (N) and (B) phosphorus (P) concentrations in shoots and P concentration in current-year shoots (C) of post-transplanted Buddhist pine (Podocarpus macrophyllus) seedlings in response to before-transplant photoperiod treatment (extended vs. natural) with (+O) or without (-O) COS addition. Different letters indicate significant differences (p<0.05, n=8).
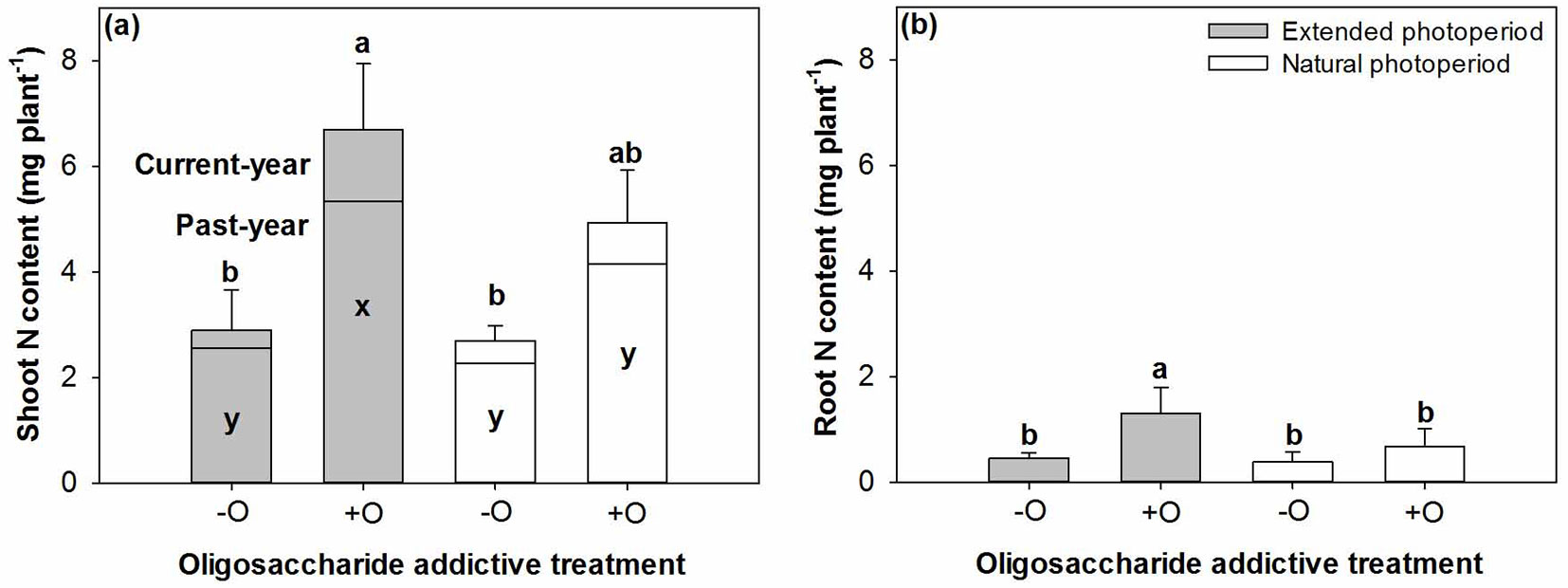
N and P contents
The responses of N content in CYS and root to combined treatments were similar to those observed for biomass (P=0.0387 and P=0.0305, respectively - Fig. 3), but N content in CYS of seedlings receiving COS was not different between the two photoperiods (Fig. 3a). Instead, photoperiod had a significant effect on N content in past-year shoot (P=0.0301) and P content in root (P=0.0132), which were higher by 23% but lower by 41% in seedlings cultured under extended photoperiod, respectively, compared with those reared under normal photoperiod (Tab. 2). The addition of COS increased not only N content in past-year shoots (P<0.0001), but also P content in all seedling organs (Tab. 3).
Fig. 3 - Nitrogen (N) content in shoot (a) and root (b) of post-transplanted Buddhist pine (Podocarpus macrophyllus) seedlings in response to before-transplant photoperiod treatment (extended vs. natural) with (+O) or without (-O) COS addition. Different letters indicate significant differences (p<0.05, n=8). In (a), a and b for current-year shoot, while x and y for past-year shoot.
Tab. 3 - Average N and P concentrations and contents in different organs (± standard error) in post-transplant Buddhist pine (Podocarpus macrophyllus) seedlings cultured with or without COS addition (n=16). Different letters in the same row indicate significant differences (p<0.05).
| Parameters | COS addition | |
|---|---|---|
| With COS | Without COS | |
| N concentration in roots (mg g-1) | 16.69 ± 2.92 b | 21.21 ± 5.16 a |
| N content in past-year shoots (mg) | 4.74 ± 1.07 a | 2.41 ± 0.57 b |
| P concentration in past-year shoots (mg g-1) | 0.56 ± 0.12 a | 0.45 ± 0.15 b |
| P content in current-year shoots (µg) | 74.93 ± 49.85 a | 28.40 ± 13.87 b |
| P content in past-year shoots (µg) | 106.69 ± 25.57 a | 41.83 ± 15.36 b |
| P content in shoots (µg) | 181.62 ± 58.60 a | 70.22 ± 20.39 b |
| P content in roots (µg) | 66.30 ± 31.44 a | 24.30 ± 11.90 b |
Correlation analysis
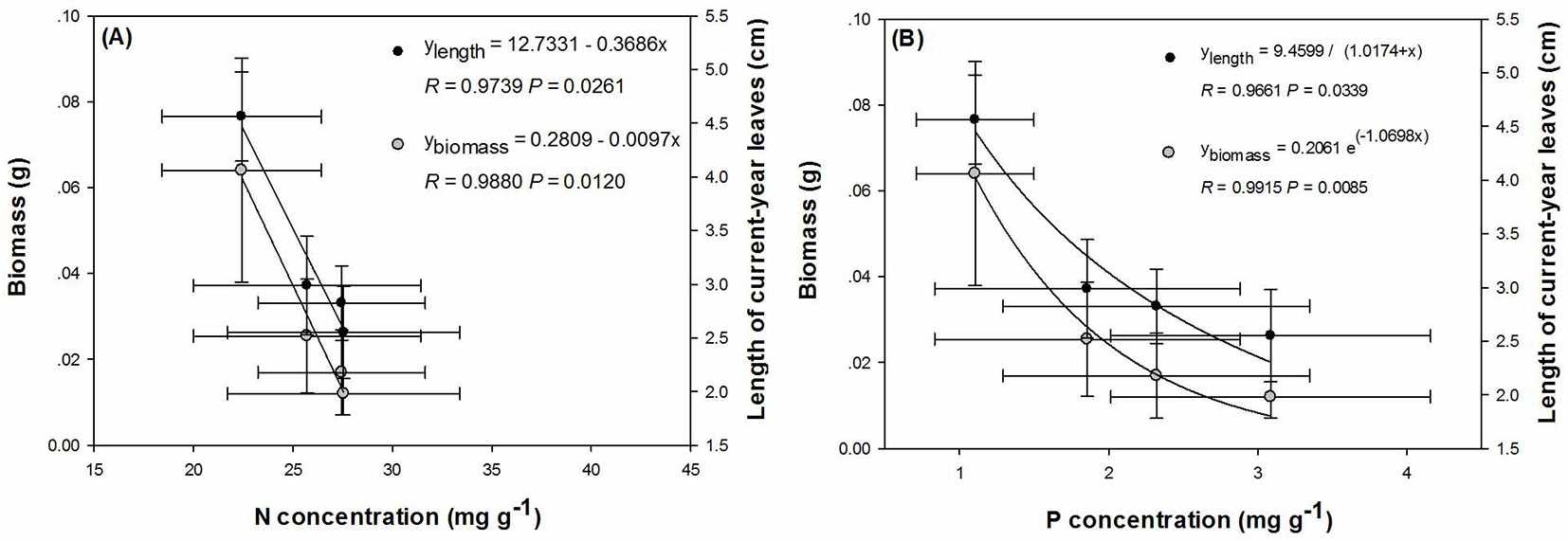
Both Pearson’s and Spearman’s correlation analyses were conducted between N and P concentrations and biomass and growth parameters. In CYS, N concentration had a negative relationship with both biomass (R=-0.9865, P=0.0135) and leaf length (R=-0.9739, P=0.0261) according to the Pearson’s correlation, while P concentration had a negative relationship with both parameters according to the Spearman’s correlation (P<0.0001 in both cases).
Regression analysis revealed that the relationships between N concentration vs. biomass and leaf length in CYS were best described by linear models (Fig. 4A), while the prediction of P concentration in seedling CYS using either parameters as independent variables were found to follow an exponential-decay model (Fig. 4B).
Fig. 4 - Regression analysis of biomass or foliar length in CYS using (A) nitrogen (N) and (B) phosphorus (P) concentrations as predictors in post-transplanted Buddhist pine (Podocarpus macrophyllus) seedlings.
Discussion
Previous studies revealed that photoperiod does not affect root-collar diameter (RCD) in Buddhist pine seedlings ([31], [32], [33]). Accordingly, our results showed an increase in RCD under the extended photoperiod only with COS addition, which had no effect on seedling height. These findings contradict to those in González et al. ([11]) and Dzung et al. ([7]), wherein COS increased the growth of aerial tree parts. However, our results concur with those by Kollárová et al. ([17]) on mung bean (Vigna radiate [L.] Wilczek) seedlings.
In our study, over 85% of seedling height was due to the first year stem growth, and mean height increased in seedlings subjected to extended photoperiod. An enhanced height growth induced by a longer photoperiod was also reported in Buddhist pine and Japanese maple (Acer palmatum) by other studies ([31], [33]). Moreover, we observed a greater number of new roots in seedlings cultured under the extended photoperiod. This was in contrast to Zhu et al. ([33]), where seedlings were checked during the first year of culture and new roots competed with shoots for source utilization. Nonetheless, we observed new root production also in transplanted seedlings, where the effect of a longer photoperiod results in a larger storage of resources which are readily available for growth in the next spring. According to Villar-Salvador et al. ([28]), conifers strongly rely on current photosynthesis and partly on N supply for early new root growth. This suggested that photoperiod might affect new root egression through the effect on current photosynthesis, though deeper studies are needed to elucidate the specific mechanism.
Leaf area affects plant productivity by increasing sunlight interception ([4]). Leaf area of a coniferous tree mostly depends on needle elongation, and leaves usually grew rapidly in length at the beginning of the growing season ([26]). In our study, COS addition increased leaf length under extended photoperiod, and these results are consistent with those on biomass accumulation in all organs (Fig. 1C). This suggests that COS may have enhanced photosynthesis rate by increasing current leaf length in seedlings grown under extended photoperiod, thus promoting their photosynthetic production. Watt & McGregor ([29]) found that second-year needle length was increased by first-year prolonged photoperiod treatment in four northern conifers seedlings. This disagreed with our results on leaf length in CYS, unless COS was added, suggesting that COS addition affects the growth of current tissues in seedlings subjected to extended photoperiod. Hansen & Beck ([14]) reported that oligosaccharides decreased drastically in Scots pine (Pinus sylvestris L.) trees in spring, when new tissues were intensively growing. This evidence supports the possible function of exogenous COS as a modifier of needle growth.
The promotion of current biomass accumulation in Buddhist pine seedlings subjected to extended photoperiod has been reported ([31], [33]). Such effect persists over the second-year regardless COS addition; indeed, first-year stem biomass accounted for about 60% of second-year biomass in Buddhist pine seedlings, and first-year stem was the only organ with greater biomass accumulation under the longer photoperiod without COS addition in the second year. However, when COS was added biomass for all organs was increased in the extended photoperiod treatment. According to previous analysis, the increase of biomass in CYS mainly resulted from the elongation of current leaves. Contrastingly, no effect of COS addition was found on the number of new roots in Buddhist pine seedlings.
Both N and P concentrations in shoots treated with COS declined in seedlings subjected to extended photoperiod, revealing the symptom of nutrient dilution. Generally, nutrient dilution tends to happen in fall, when seedling growth continued during hardening without sufficient nutrient supply, which could lead to the degradation of seedling quality and lowered post-transplant performances ([1]). Therefore, nutrient dilution in shoots may be considered as a serious drawback caused by the exposure of seedlings to the extended photoperiod. However, shoot N and P concentrations in our study seemed to be acceptable for transplanted seedling growth, as nutrients were utilized for biomass accumulation and leaf elongation in CYS (Fig. 4). It is worthy to notice that only with COS addition nutrient concentration was diluted in seedling shoots. These results, along with those about growth and biomass accumulation in CYS, suggest that first-year COS addition promoted the utilization of N and P in shoots for CYS growth. On the other hand, the longer photoperiod induced higher N concentration in annual organs, but less P retained in roots (Tab. 2). This finding is the opposite of that reported by Wei et al. ([31]), in one-year-old Buddhist pine seedlings, where N concentration was lower and P concentration was higher in annual organs under the longer photoperiod. In the first year a large amount of N was allocated to shoots and roots for utilization, while most P was retained in annual organs. Nevertheless, the N demand by CYS was not so compelling under the extended photoperiod in the second year as a large amount of N was retained in the roots.
In seedling treated with COS, we found both lower N concentration in roots and greater N content in first-year shoots, which could be caused by two separated processes. The decline of N concentration in roots was driven by the N utilization for CYS growth in response to COS addition, because of both CYS growth and increase of shoot N concentration in COS-treated seedlings. On the other hand, the greater N content observed in the old stem of seedlings likely resulted from biomass accumulation therein, as nutrient content is the result of nutrient concentration by biomass. In contrast, the higher P content in old stems after COS addition was mainly attributed to the promotion of P concentration, but it did not lead to biomass accumulation. Further, with the addition of COS P uptake and accumulation was found to be enhanced in all seedling organs. COS addition had also been found to promote P uptake in Maize (Zea mays - [27]) and coffee (Coffea canephora var. Robusta - [7]).
Conclusions
Current understanding of the effect of COS on plants was mainly based on studies on herbaceous plants, and few information exists on woody species. In the present study, a two-year experiment was conducted on Buddhist pine seedlings treated with COS addition under contrasting photoperiods. Under the extended photoperiod COS addition increased leaf length in CYS as well as biomass accumulation and N content at the whole-plant scale. In contrast, addition of COS decreased RCD under the natural photoperiod treatment, but it had no effect on seedling height. Both N and P concentrations in seedling shoots were increased by COS addition under the extended photoperiod, and they were also negatively correlated with leaf length and biomass accumulation in CYS.
In conclusion, the effect of extended photoperiod on growth and nutrient utilization would diminish in CYS of transplanted Buddhist pine seedlings, unless COS was added during culture. Both seed coating and leaf spraying with COS were found to result in similar effects on plant growth.
This study is the first trial to determine the over-year effect of COS on tree seedlings under contrasting photoperiods. Additional studies on different tree species are needed to confirm our results and avoid possible species-specific limits of this study.
Acknowledgments
This study was supported by the National Natural Science Foundation of China (31600496) and The National Key Research and Development Program of China (2016YFC0500307).
References
Authors’ Info
Authors’ Affiliation
Northeast Institute of Geography and Agricultural Ecology, Chinese Academy of Sciences, Changchun 130102 (China)
Corresponding author
Paper Info
Citation
Wang Z, Zhao Y, Wei H (2017). Chitosan oligosaccharide addition affects current-year shoot of post-transplant Buddhist pine (Podocarpus macrophyllus) seedlings under contrasting photoperiods. iForest 10: 715-721. - doi: 10.3832/ifor2302-010
Academic Editor
Claudia Cocozza
Paper history
Received: Dec 10, 2016
Accepted: May 29, 2017
First online: Jul 27, 2017
Publication Date: Aug 31, 2017
Publication Time: 1.97 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2017
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 49841
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 41509
Abstract Page Views: 3350
PDF Downloads: 3858
Citation/Reference Downloads: 14
XML Downloads: 1110
Web Metrics
Days since publication: 3247
Overall contacts: 49841
Avg. contacts per week: 107.45
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2017): 26
Average cites per year: 2.89
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Conservation of Betula oycoviensis, an endangered rare taxon, using vegetative propagation methods
vol. 13, pp. 107-113 (online: 23 March 2020)
Technical Reports
De novo adventitious root formations in mini-cuttings of Azadirachta indica in response to different rooting media and auxin treatments
vol. 8, pp. 558-564 (online: 09 December 2014)
Research Articles
Forest litter as the mulch improving growth and ectomycorrhizal diversity of bare-root Scots pine (Pinus sylvestris) seedlings
vol. 8, pp. 394-400 (online: 20 August 2014)
Research Articles
Use of alternative containers for promoting deep rooting of native forest species used for dryland restoration: the case of Acacia caven
vol. 10, pp. 776-782 (online: 02 September 2017)
Research Articles
Fine root morphological traits and production in coniferous- and deciduous-tree forests with drained and naturally wet nutrient-rich organic soils in hemiboreal Latvia
vol. 16, pp. 165-173 (online: 08 June 2023)
Research Articles
Modeling the fine root biomass dispersion using a special influence function
vol. 1, pp. 141-144 (online: 25 November 2008)
Research Articles
Fine root production and distribution in the tropical rainforests of south-western Cameroon: effects of soil type and selective logging
vol. 3, pp. 130-136 (online: 27 September 2010)
Research Articles
Fertilisation of Quercus seedlings inoculated with Tuber melanosporum: effects on growth and mycorrhization of two host species and two inoculation methods
vol. 10, pp. 267-272 (online: 13 December 2016)
Research Articles
Forecasting the field performance of Austrian pine seedlings using morphological attributes
vol. 10, pp. 99-107 (online: 13 October 2016)
Technical Reports
Effects of different mechanical treatments on Quercus variabilis, Q. wutaishanica and Q. robur acorn germination
vol. 8, pp. 728-734 (online: 05 May 2015)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







